

Chemical and Biological Properties of Biochanin A and Its Pharmaceutical Applications


Abstract
:1. Introduction
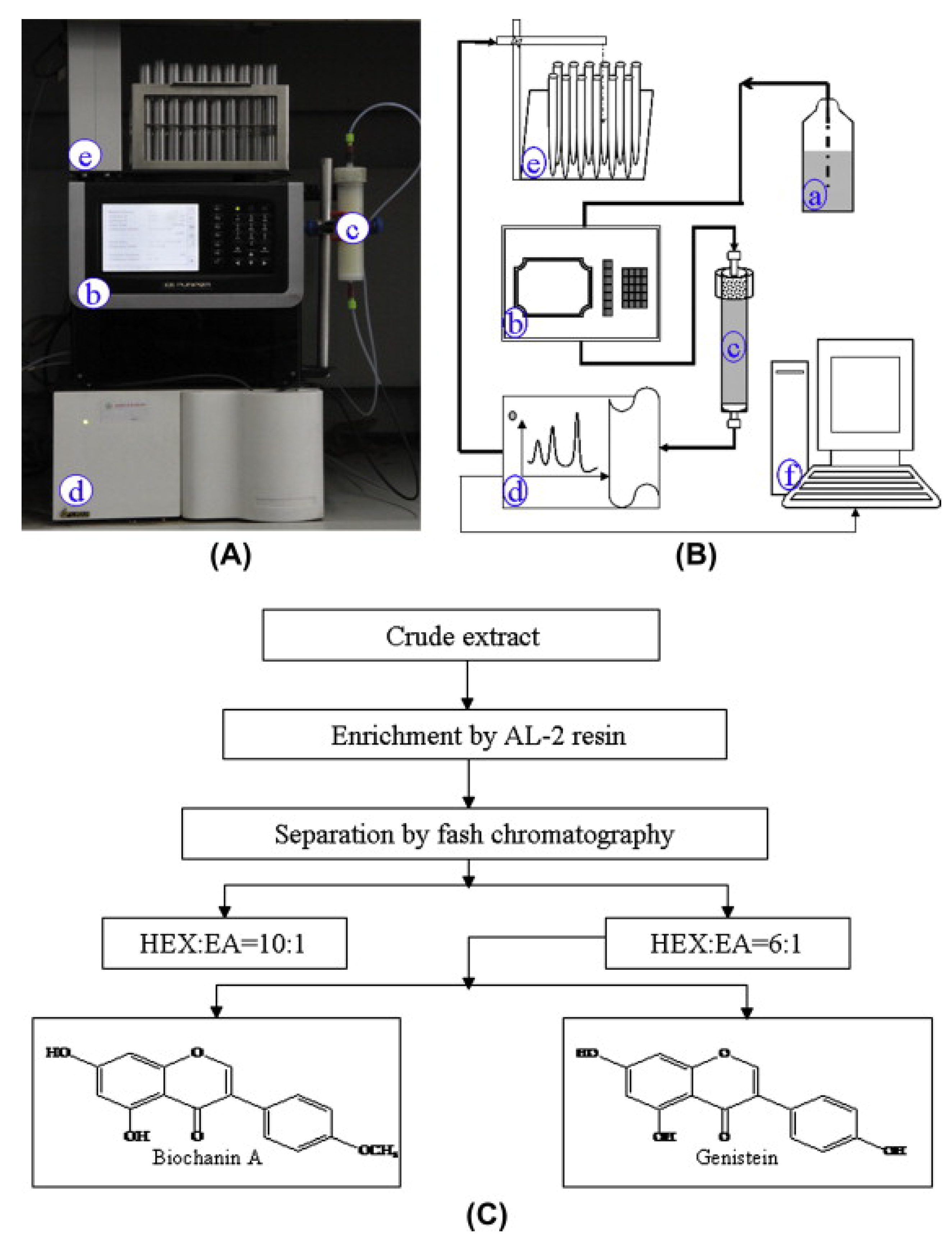
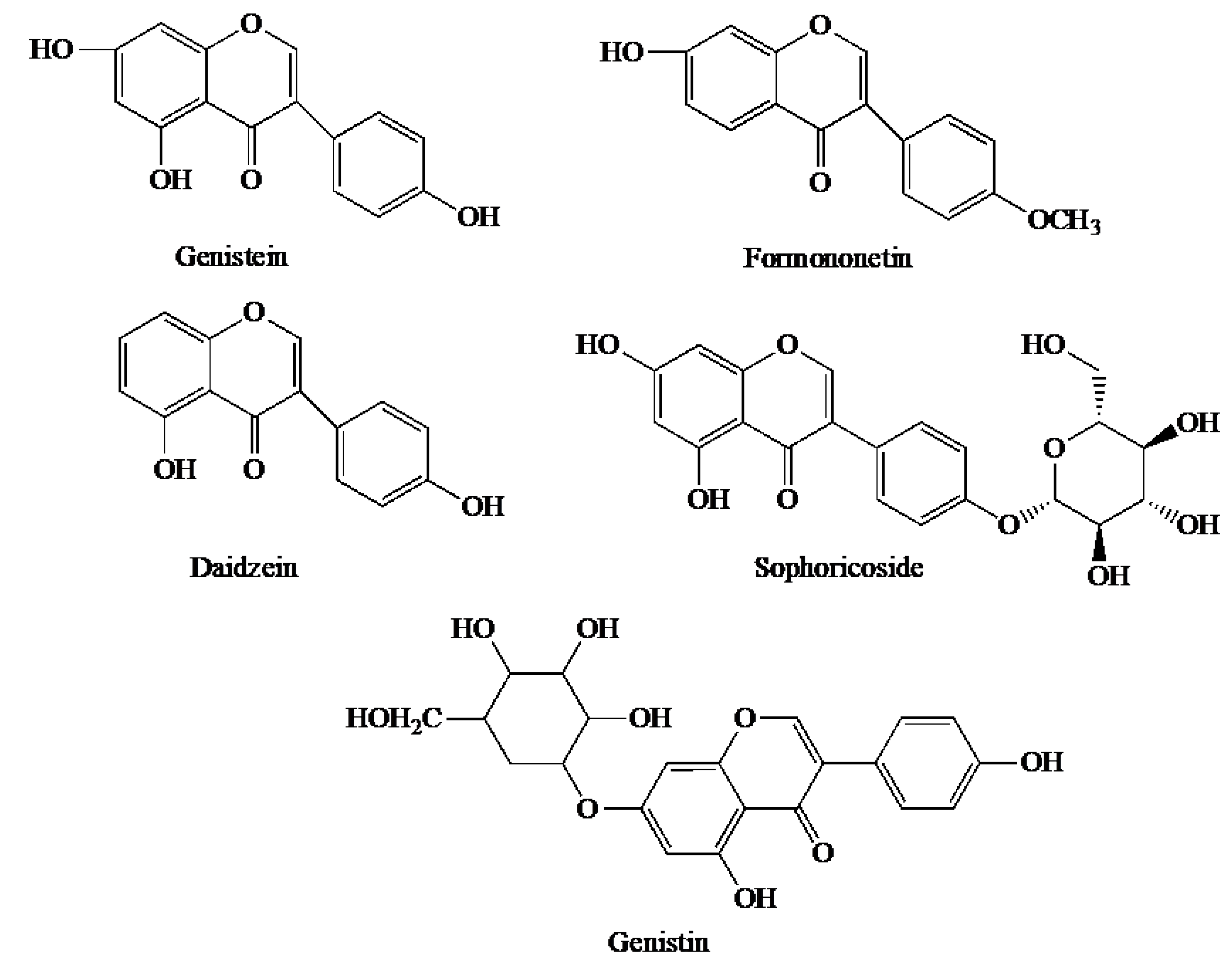
2. Chemical Properties and Extraction of BCA
3. Physiological Functions of BCA
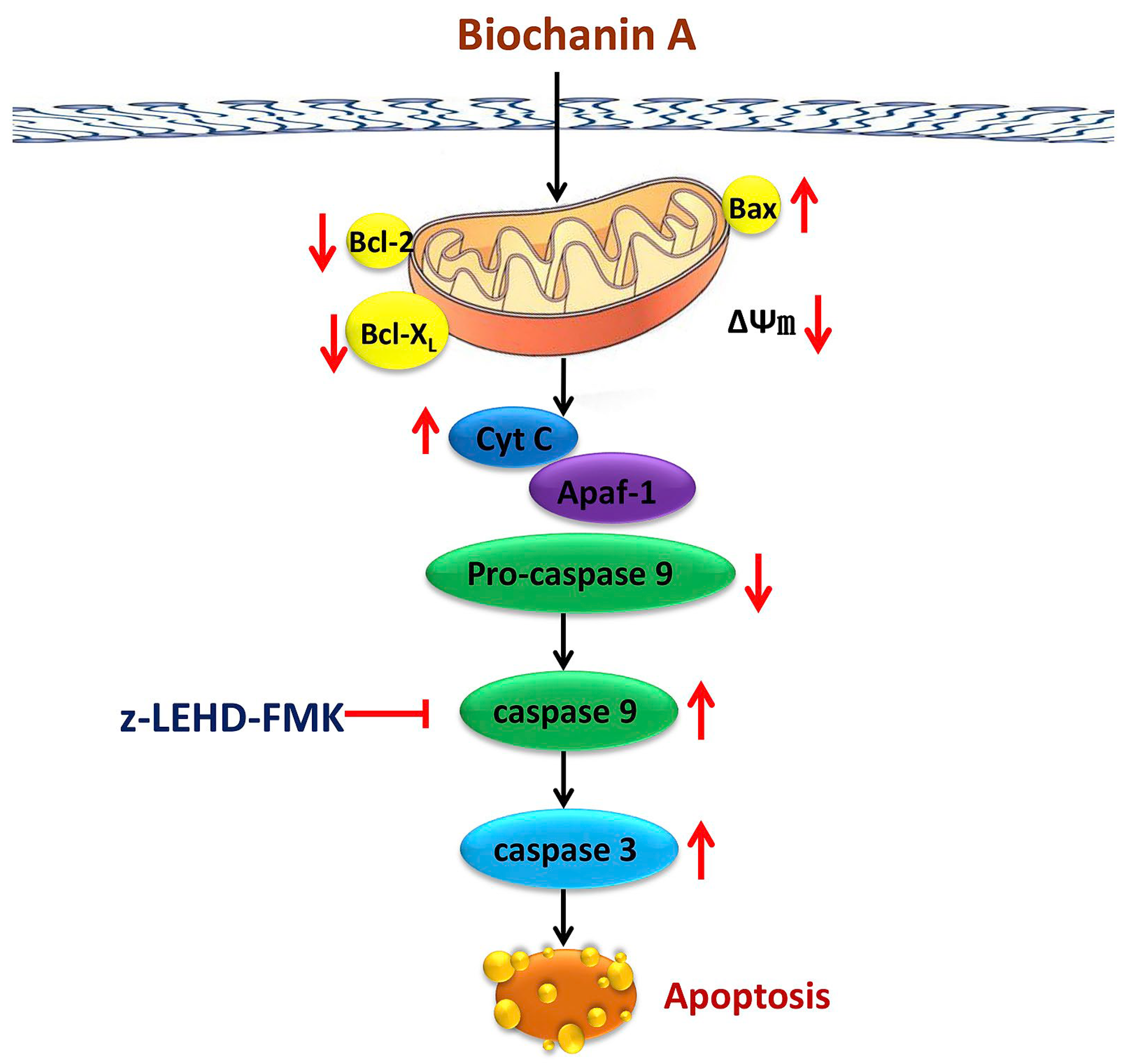
3.1. Anti-Cancer Function
3.2. Anti-Oxidant Effects
3.3. Anti-Inflammatory Function
4. Metabolism and Strategies to Enhance the Bioavailability of BCA
5. Opportunities and Challenges for Applications of BCA and Its Sources
6. Concluding Remarks and Outlooks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purushothaman, R.; Krishnamurthy, L.; Upadhyaya, H.D.; Vadez, V.; Varshney, R.K. Genotypic variation in soil water use and root distribution and their implications for drought tolerance in chickpea. Funct. Plant Biol. 2017, 44, 235–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grasso, N.; Lynch, N.L.; Arendt, E.K.; O’Mahony, J.A. Chickpea protein ingredients: A review of composition, functionality, and applications. Compr. Rev. Food Sci. Food Saf. 2022, 21, 435–452. [Google Scholar] [CrossRef]
- Ying-hua, F.; Zheng-lei, L.; Ying-jie, L. Research progress on chickpea resource and isoflavones. Storage Process 2021, 21, 130–135. [Google Scholar] [CrossRef]
- Krizova, L.; Dadakova, K.; Kasparovska, J.; Kasparovsky, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Yao, Y.; Zhu, Y.; Ren, G. Isoflavone content and composition in chickpea (Cicer arietinum L.) sprouts germinated under different conditions. J. Agric. Food Chem. 2015, 63, 2701–2707. [Google Scholar] [CrossRef]
- Chukwumah, Y.C.; Walker, L.T.; Verghese, M.; Ogutu, S. Effect of frequency and duration of ultrasonication on the extraction efficiency of selected isoflavones and trans-resveratrol from peanuts (Arachis hypogaea). Ultrason. Sonochem. 2009, 16, 293–299. [Google Scholar] [CrossRef]
- Hsu, Y.N.; Shyu, H.W.; Hu, T.W.; Yeh, J.P.; Lin, Y.W.; Lee, L.Y.; Yeh, Y.T.; Dai, H.Y.; Perng, D.S.; Su, S.H.; et al. Anti-proliferative activity of biochanin A in human osteosarcoma cells via mitochondrial-involved apoptosis. Food Chem. Toxicol. 2018, 112, 194–204. [Google Scholar] [CrossRef]
- Kagan, I.A.; Goff, B.M.; Flythea, M.D. Soluble phenolic compounds in different cultivars of red clover and alfalfa, and their implication for protection against proteolysis and ammonia production in ruminants. Nat. Prod. Commun. 2015, 10, 1263–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zgorka, G. Pressurized liquid extraction versus other extraction techniques in micropreparative isolation of pharmacologically active isoflavones from Trifolium L. species. Talanta 2009, 79, 46–53. [Google Scholar] [CrossRef]
- Butkute, B.; Dagilyte, A.; Benetis, R.; Padarauskas, A.; Ceseviciene, J.; Olsauskaite, V.; Lemeziene, N. Mineral and phytochemical profiles and antioxidant activity of herbal material from two temperate astragalus species. Biomed. Res. Int. 2018, 2018, 6318630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.R.; Kim, H.J.; Yu, S.H.; Lee, B.S.; Jeon, S.Y.; Lee, J.J.; Lee, Y.C. Combination of red clover and hops extract improved menopause symptoms in an ovariectomized rat model. Evid.-Based Complement. Alternat. Med. 2020, 2020, 7941391. [Google Scholar] [CrossRef]
- Castellano, G.; Torrens, F. Quantitative structure-antioxidant activity models of isoflavonoids: A theoretical study. Int. J. Mol. Sci. 2015, 16, 12891–12906. [Google Scholar] [CrossRef] [Green Version]
- Beszterda, M.; Kasperkowiak, M.; Franski, R. Comment on the published data concerning the identification of biochanin A and prunetin by LC/ESI-MS. Talanta 2020, 211, 120733. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-H.; Mei, Y.-H.; Wang, F.; Liu, X.-S.; Chen, Y. A novel and efficient method combining SFE and liquid–liquid extraction for separation of coumarins from Angelica dahurica. Sep. Purif. Technol. 2011, 77, 397–401. [Google Scholar] [CrossRef]
- Flores-Espinosa, R.M.; Ortíz-Oliveros, H.B.; Olguín, M.T.; Perusquia-Cueto, M.R.; Gallardo-San-Vicente, R. Separation and treatment of ion-exchange resins used in cleaning systems of a research nuclear reactor. Chem. Eng. J. 2012, 188, 71–76. [Google Scholar] [CrossRef]
- Cagliero, C.; Galli, S.; Galli, M.; Elmi, I.; Belluce, M.; Zampolli, S.; Sgorbini, B.; Rubiolo, P.; Bicchi, C. Conventional and enantioselective gas chromatography with microfabricated planar columns for analysis of real-world samples of plant volatile fraction. J. Chromatogr. A 2016, 1429, 329–339. [Google Scholar] [CrossRef]
- Ma, F.-Y.; Luo, M.; Zhao, C.-J.; Li, C.-Y.; Wang, W.; Gu, C.-B.; Wei, Z.-F.; Zu, Y.-G.; Fu, Y.-J. Simple and efficient preparation of biochanin A and genistein from Dalbergia odorifera T. Chen leaves using macroporous resin followed by flash chromatography. Sep. Purif. Technol. 2013, 120, 310–318. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Sun, X.; Liu, Y.-G.; Han, R.-M.; Lan, L.; Chen, H.-B.; Sun, X.-R.; Li, Q.; Tang, Y.-L. Interaction of isoflavones with different structures and transferrin. Spectrosc. Lett. 2016, 49, 596–601. [Google Scholar] [CrossRef]
- Fokialakis, N.; Alexi, X.; Aligiannis, N.; Siriani, D.; Meligova, A.K.; Pratsinis, H.; Mitakou, S.; Alexis, M.N. Ester and carbamate ester derivatives of Biochanin A: Synthesis and in vitro evaluation of estrogenic and antiproliferative activities. Bioorg. Med. Chem. 2012, 20, 2962–2970. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Martín, L.; Bustamante-Rangel, M.; Delgado-Zamarreño, M.M. Classification of lentils, chickpeas and beans based on their isoflavone content. Food Anal. Methods 2016, 10, 1191–1201. [Google Scholar] [CrossRef]
- Oza, M.J.; Kulkarni, Y.A. Biochanin A improves insulin sensitivity and controls hyperglycemia in type 2 diabetes. Biomed. Pharmacother. 2018, 107, 1119–1127. [Google Scholar] [CrossRef]
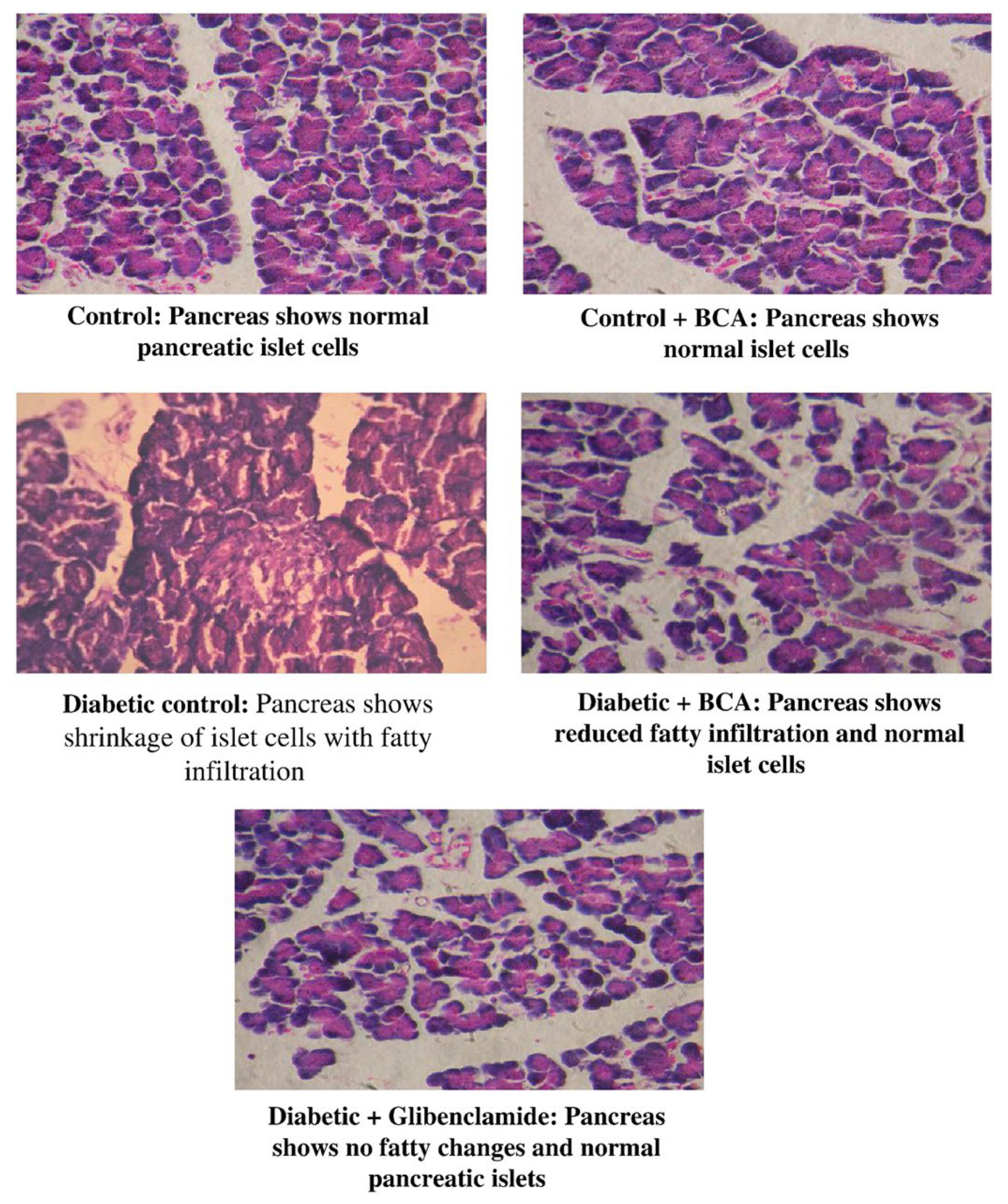
- Harini, R.; Ezhumalai, M.; Pugalendi, K.V. Antihyperglycemic effect of biochanin A, a soy isoflavone, on streptozotocin-diabetic rats. Eur. J. Pharmacol. 2012, 676, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Thors, L.; Burston, J.J.; Alter, B.J.; McKinney, M.K.; Cravatt, B.F.; Ross, R.A.; Pertwee, R.G.; Gereau, R.W.t.; Wiley, J.L.; Fowler, C.J. Biochanin A, a naturally occurring inhibitor of fatty acid amide hydrolase. Br. J. Pharmacol. 2010, 160, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Yan, L.T.; Yao, Z.; Xiong, G.Y. Biochanin A regulates cholesterol metabolism further delays the progression of nonalcoholic fatty liver disease. Diabetes Metab. Syndr. Obes. 2021, 14, 3161–3172. [Google Scholar] [CrossRef]
- Youn, K.; Park, J.H.; Lee, J.; Jeong, W.S.; Ho, C.T.; Jun, M. The identification of biochanin A as a potent and selective β-site APP-cleaving enzyme 1 (Bace1) inhibitor. Nutrients 2016, 8, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirone, M.; Zompetta, C.; Tarasi, D.; Frati, L.; Faggioni, A. Infection of human T lymphoid cells by human herpesvirus 6 is blocked by two unrelated protein tyrosine kinase inhibitors, biochanin A and herbimycin. AIDS Res. Hum. Retrovir. 1996, 12, 1629–1634. [Google Scholar] [CrossRef]
- Sklenickova, O.; Flesar, J.; Kokoska, L.; Vlkova, E.; Halamova, K.; Malik, J. Selective growth inhibitory effect of biochanin A against intestinal tract colonizing bacteria. Molecules 2010, 15, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Liang, J.C.; Wang, X.L.; Li, Z.H.; Wang, W.; Guo, N.; Wu, X.P.; Shen, F.G.; Xing, M.X.; Liu, L.H.; et al. In vitro synergy of biochanin A and ciprofloxacin against clinical isolates of Staphylococcus aureus. Molecules 2011, 16, 6656–6666. [Google Scholar] [CrossRef]
- Hanski, L.; Genina, N.; Uvell, H.; Malinovskaja, K.; Gylfe, A.; Laaksonen, T.; Kolakovic, R.; Makila, E.; Salonen, J.; Hirvonen, J.; et al. Inhibitory activity of the isoflavone biochanin A on intracellular bacteria of genus Chlamydia and initial development of a buccal formulation. PLoS ONE 2014, 9, e115115. [Google Scholar] [CrossRef]
- Xu, J.; Yang, X.; Pan, J.; Fan, H.; Mei, J.; Hua, D. Biochanin A suppresses tumor progression and PD-L1 expression via inhibiting ZEB1 expression in colorectal cancer. J. Oncol. 2022, 2022, 3224373. [Google Scholar] [CrossRef]
- Li, Y.; Yu, H.; Han, F.; Wang, M.; Luo, Y.; Guo, X. Biochanin A induces S phase arrest and apoptosis in lung cancer cells. Biomed. Res. Int. 2018, 2018, 3545376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehdev, V.; Lai, J.C.; Bhushan, A. Biochanin A modulates cell viability, invasion, and growth promoting signaling pathways in HER-2-positive breast cancer cells. J. Oncol. 2009, 2009, 121458. [Google Scholar] [CrossRef] [Green Version]
- Liang, F.; Cao, W.; Huang, Y.; Fang, Y.; Cheng, Y.; Pan, S.; Xu, X. Isoflavone biochanin A, a novel nuclear factor erythroid 2-related factor 2 (Nrf2)-antioxidant response element activator, protects against oxidative damage in HepG2 cells. Biofactors 2019, 45, 563–574. [Google Scholar] [CrossRef]
- Wu, Q.; Shang, Y.; Shen, T.; Liu, F.; Zhang, W. Biochanin A protects SH-SY5Y cells against isoflurane-induced neurotoxicity by suppressing oxidative stress and apoptosis. Neurotoxicology 2021, 86, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Q.; Jin, Z.Y.; Li, G.H. Biochanin A protects dopaminergic neurons against lipopolysaccharide-induced damage through inhibition of microglia activation and proinflammatory factors generation. Neurosci. Lett. 2007, 417, 112–117. [Google Scholar] [CrossRef]
- Zhang, S.; Niu, Y.; Yang, Z.; Zhang, Y.; Guo, Q.; Yang, Y.; Zhou, X.; Ding, Y.; Liu, C. Biochanin A alleviates gingival inflammation and alveolar bone loss in rats with experimental periodontitis. Exp. Ther. Med. 2020, 20, 251. [Google Scholar] [CrossRef]
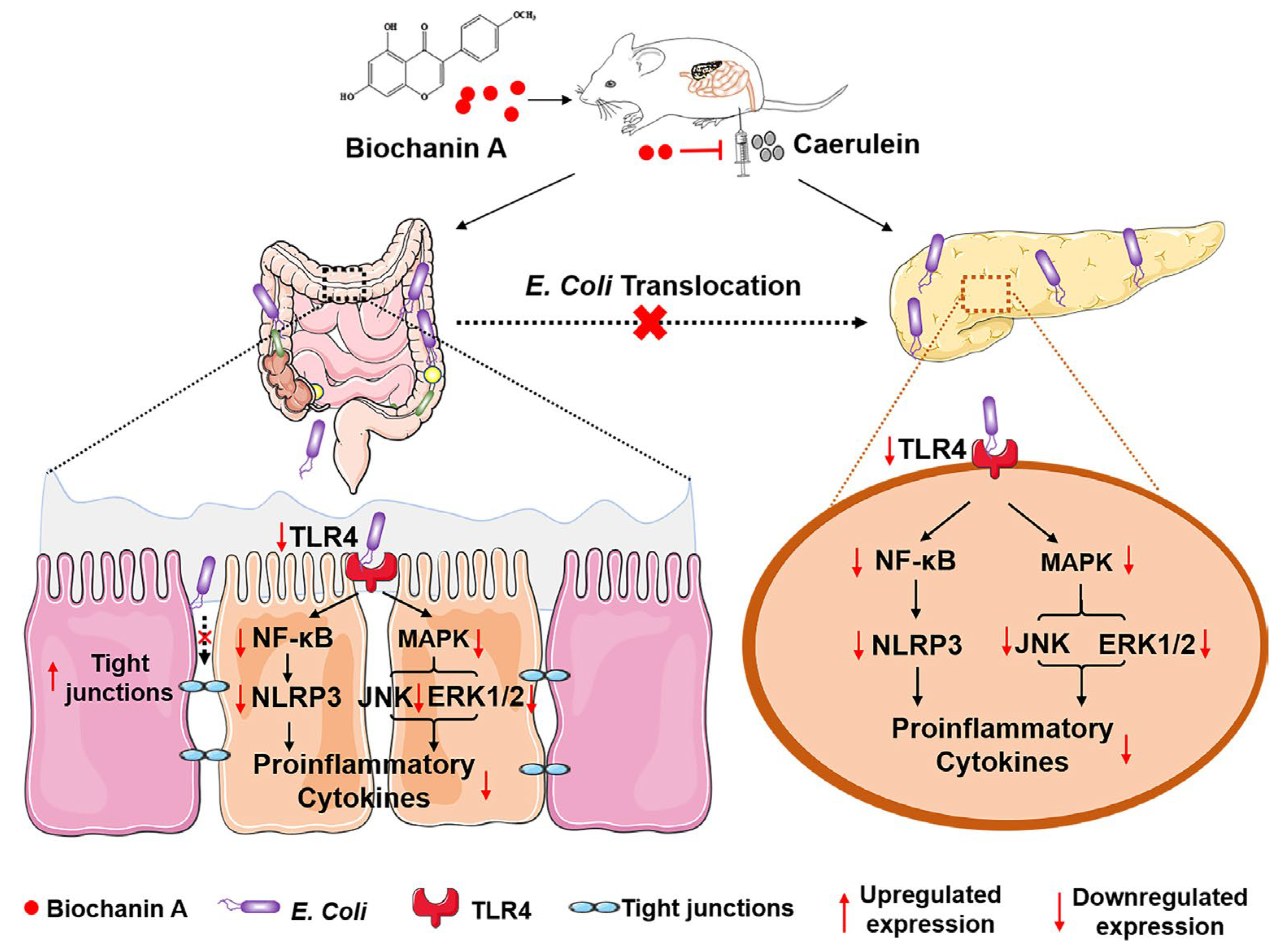
- Pan, X.; Ye, L.; Ren, Z.; Li, J.; Li, B.; Pan, L.L.; Sun, J. Biochanin A ameliorates caerulein-induced acute pancreatitis and associated intestinal injury in mice by inhibiting TLR4 signaling. J. Nutr. Biochem. 2022, 113, 109229. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Global Health Estimates 2020: Deaths by Cause, Age, Sex, by Country and by Region, 2000–2019; World Health Organization (WHO): Geneva, Switzerland, 2020. [Google Scholar]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Puthli, A.; Tiwari, R.; Mishra, K.P. Biochanin A enhances the radiotoxicity in colon tumor cells in vitro. J. Environ. Pathol. Toxicol. Oncol. 2013, 32, 189–203. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.J.; Chen, Y.M. Biochanin A extirpates the epithelial-mesenchymal transition in a human lung cancer. Exp. Ther. Med. 2018, 15, 2830–2836. [Google Scholar] [CrossRef] [Green Version]
- Cho, I.A.; You, S.J.; Kang, K.R.; Kim, S.G.; Oh, J.S.; You, J.S.; Lee, G.J.; Seo, Y.S.; Kim, D.K.; Kim, C.S.; et al. Biochanin-A induces apoptosis and suppresses migration in FaDu human pharynx squamous carcinoma cells. Oncol. Rep. 2017, 38, 2985–2992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaina, V.K.; Eedara, A.; Svs, S.P.; Jadav, S.S.; Chilaka, S.; Sistla, R.; Andugulapati, S.B. Anti-cancer activity of biochanin A against multiple myeloma by targeting the CD38 and cancer stem-like cells. Process Biochem. 2022, 123, 11–26. [Google Scholar] [CrossRef]
- Dong, Q.; Wang, D.; Li, L.; Wang, J.; Li, Q.; Duan, L.; Yin, H.; Wang, X.; Liu, Y.; Yuan, G.; et al. Biochanin A sensitizes glioblastoma to temozolomide by inhibiting autophagy. Mol. Neurobiol. 2022, 59, 1262–1272. [Google Scholar] [CrossRef]
- Jalaludeen, A.M.; Ha, W.T.; Lee, R.; Kim, J.H.; Do, J.T.; Park, C.; Heo, Y.T.; Lee, W.Y.; Song, H. Biochanin A ameliorates arsenic-induced hepato- and hematotoxicity in rats. Molecules 2016, 21, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haridevamuthu, B.; Guru, A.; Murugan, R.; Sudhakaran, G.; Pachaiappan, R.; Almutairi, M.H.; Almutairi, B.O.; Juliet, A.; Arockiaraj, J. Neuroprotective effect of biochanin A against bisphenol A-induced prenatal neurotoxicity in zebrafish by modulating oxidative stress and locomotory defects. Neurosci. Lett. 2022, 790, 136889. [Google Scholar] [CrossRef]
- Lancaster, L.H.; de Andrade, J.A.; Zibrak, J.D.; Padilla, M.L.; Albera, C.; Nathan, S.D.; Wijsenbeek, M.S.; Stauffer, J.L.; Kirchgaessler, K.U.; Costabel, U. Pirfenidone safety and adverse event management in idiopathic pulmonary fibrosis. Eur. Respir. Rev. 2017, 26, 170057. [Google Scholar] [CrossRef] [Green Version]
- Andugulapati, S.B.; Gourishetti, K.; Tirunavalli, S.K.; Shaikh, T.B.; Sistla, R. Biochanin-A ameliorates pulmonary fibrosis by suppressing the TGF-β mediated EMT, myofibroblasts differentiation and collagen deposition in in vitro and in vivo systems. Phytomedicine 2020, 78, 153298. [Google Scholar] [CrossRef]
- Lee, P.J.; Papachristou, G.I. New insights into acute pancreatitis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 479–496. [Google Scholar] [CrossRef]
- Heinonen, S.M.; Wahala, K.; Adlercreutz, H. Identification of urinary metabolites of the red clover isoflavones formononetin and biochanin A in human subjects. J. Agric. Food Chem. 2004, 52, 6802–6809. [Google Scholar] [CrossRef]
- Tolleson, W.H.; Doerge, D.R.; Churchwell, M.I.; Marques, M.M.; Roberts, D.W. Metabolism of biochanin A and formononetin by human liver microsomes in vitro. J. Agric. Food Chem. 2002, 50, 4783–4790. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Zhang, X.; Zhang, Y.; Ma, Y.; Li, L.; Li, D.; Zhang, L.; Zhang, Z. Comprehensive study of the in vivo and in vitro metabolism of dietary isoflavone biochanin A based on UHPLC-Q-TOF-MS/MS. J. Agric. Food Chem. 2019, 67, 12481–12495. [Google Scholar] [CrossRef] [PubMed]
- Tay, K.C.; Tan, L.T.; Chan, C.K.; Hong, S.L.; Chan, K.G.; Yap, W.H.; Pusparajah, P.; Lee, L.H.; Goh, B.H. Formononetin: A review of its anticancer potentials and mechanisms. Front. Pharmacol. 2019, 10, 820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawat, S.; Pathak, S.; Gupta, G.; Singh, S.K.; Singh, H.; Mishra, A.; Gilhotra, R. Recent updates on daidzein against oxidative stress and cancer. EXCLI J. 2019, 18, 950–954. [Google Scholar] [CrossRef]
- Choi, Y.R.; Shim, J.; Kim, M.J. Genistin: A novel potent anti-adipogenic and anti-lipogenic agent. Molecules 2020, 25, 2042. [Google Scholar] [CrossRef] [PubMed]
- Yanjun, L.; Nan, Z.; Jiaqi, W.; Shengguo, Z. Research progress on biological functions of biochanin A. Chin. J. Anim. Nutr. 2022, 34, 2177–2185. [Google Scholar] [CrossRef]
- Yu, C.; Zhang, P.; Lou, L.; Wang, Y. Perspectives regarding the role of biochanin A in humans. Front. Pharmacol. 2019, 10, 793. [Google Scholar] [CrossRef] [Green Version]
- Moon, Y.J.; Sagawa, K.; Frederick, K.; Zhang, S.; Morris, M.E. Pharmacokinetics and bioavailability of the isoflavone biochanin A in rats. AAPS J. 2006, 8, E433–E442. [Google Scholar] [CrossRef] [Green Version]
- Ling, X.B. Breviscapine Alleviates transforming growth factor-β 1-induced epithelial-mesenchymal transition of human alveolar epithelium cells A549 via inhibiting PI3K/AKT/GSK3 β pathway. J. Biol. Reg. Homeos. Agents 2022, 36, 611–623. [Google Scholar] [CrossRef]
- Liu, Q.Q.; Peng, H.Y.; Wang, Z.A. Convergence to nonlinear diffusion waves for a hyperbolic-parabolic chemotaxis system modelling vasculogenesis. J. Differ. Equ. 2022, 314, 251–286. [Google Scholar] [CrossRef]
- Lou, J.S.; Zhao, L.P.; Huang, Z.H.; Chen, X.Y.; Xu, J.T.; Tai, W.C.S.; Tsim, K.W.K.; Chen, Y.T.; Xie, T. Ginkgetin derived from Ginkgo biloba leaves enhances the therapeutic effect of cisplatin via ferroptosis-mediated disruption of the Nrf2/HO-1 axis in EGFR wild-type non-small-cell lung cancer. Phytomedicine 2021, 80, 153370. [Google Scholar] [CrossRef]
- Sui, X.B.; Zhang, R.N.; Liu, S.P.; Duan, T.; Zhai, L.J.; Zhang, M.M.; Han, X.M.; Xiang, Y.; Huang, X.X.; Lin, H.M.; et al. RSL3 drives ferroptosis through GPX4 inactivation and ROS production in colorectal cancer. Front. Pharmacol. 2018, 9, 1371. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Qu, Y.Y.; Liu, L.; Qiao, Y.N.; Geng, H.R.; Lin, Y.; Xu, W.; Cao, J.; Zhao, J.Y. Homocysteine inhibits pro-insulin receptor cleavage and causes insulin resistance via protein cysteine-homocysteinylation. Cell Rep. 2021, 37, 109821. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.J.; Deng, P.; Peng, C.E.; Ji, H.Y.; Mao, L.F.; Peng, L.Z. Extraction, structure and immunoregulatory activity of low molecular weight polysaccharide from Dendrobium officinale. Polymers 2022, 14, 2899. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.H.; Chen, X.Y.; Zhu, L.Y.; Liu, W.; Jiang, L. Programming an orthogonal self-assembling protein cascade based on reactive peptide-protein pairs for in vitro enzymatic trehalose production. J. Agr. Food Chem. 2022, 70, 4690–4700. [Google Scholar] [CrossRef]
- Jin, H.Y.; Wang, Z.A. Global stabilization of the full attraction-repulsion Keller-Segel system. Discret. Cont. Dyn.-A 2020, 40, 3509–3527. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Shi, J.P.; Wang, Z.A. Pattern formation of the attraction-repulsion Keller-Segel system. Discret. Cont. Dyn.-B 2013, 18, 2597–2625. [Google Scholar] [CrossRef]
- Zhang, C.; Li, J.X.; Xiao, M.; Wang, D.; Qu, Y.; Zou, L.; Zheng, C.A.; Zhang, J.M. Oral colon-targeted mucoadhesive micelles with enzyme-responsive controlled release of curcumin for ulcerative colitis therapy. Chin. Chem. Lett. 2022, 33, 4924–4929. [Google Scholar] [CrossRef]
- Zeng, Q.B.; Bie, B.L.; Guo, Q.N.; Yuan, Y.P.; Han, Q.; Han, X.C.; Chen, M.W.; Zhang, X.; Yang, Y.H.; Liu, M.L.; et al. Hyperpolarized Xe NMR signal advancement by metal-organic framework entrapment in solution. Proc. Natl. Acad. Sci. USA 2020, 117, 17558–17563. [Google Scholar] [CrossRef] [PubMed]
- Han, H.K.; Lee, B.J.; Lee, H.K. Enhanced dissolution and bioavailability of biochanin A via the preparation of solid dispersion: In vitro and in vivo evaluation. Int. J. Pharm. 2011, 415, 89–94. [Google Scholar] [CrossRef]
- Wu, X.; Ge, W.; Shao, T.; Wu, W.; Hou, J.; Cui, L.; Wang, J.; Zhang, Z. Enhancing the oral bioavailability of biochanin A by encapsulation in mixed micelles containing Pluronic F127 and Plasdone S630. Int. J. Nanomed. 2017, 12, 1475–1483. [Google Scholar] [CrossRef] [Green Version]
- Sachdeva, C.; Mishra, N.; Sharma, S. Development and characterization of enteric-coated microparticles of biochanin A for their beneficial pharmacological potential in estrogen deficient-hypertension. Drug Deliv. 2016, 23, 2044–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajic, I.; Stojanovic, S.; Ristic, I.; Ilic-Stojanovic, S.; Pilic, B.; Nesic, A.; Najman, S.; Dinic, A.; Stanojevic, L.; Urosevic, M.; et al. Electrospun poly(lactide) fibers as carriers for controlled release of biochanin A. Pharmaceutics 2022, 14, 528. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, I.L.; Savic, I.M.; Popsavin, M.M.; Rakic, S.J.; Mihajilov-Krstev, T.M.; Ristic, I.S.; Eric, S.P.; Savic-Gajic, I.M. Preparation, characterization and antimicrobial activity of inclusion complex of biochanin A with (2-hydroxypropyl)-beta-cyclodextrin. J. Pharm. Pharmacol. 2018, 70, 1485–1493. [Google Scholar] [CrossRef] [PubMed]
- Mallis, L.; Sarkahian, A.; Harris, H.; Zhang, M.; McConnell, O. Determination of rat oral bioavailability of soy-derived phytoestrogens using an automated on-column extraction procedure and electrospray tandem mass spectrometry. J. Chromatogr. B 2003, 796, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Yangzom, P.; Amruthanand, S.; Sharma, M.; Mahajan, S.; Lingaraju, M.C.; Parida, S.; Sahoo, M.; Kumar, D.; Singh, T.U. Subacute 28 days oral toxicity study of kaempferol and biochanin-A in the mouse model. J. Biochem. Mol. Toxicol. 2022, 36, e23090. [Google Scholar] [CrossRef]
- Li, P.; Li, M.; Lou, X.; Zhao, B.; Ma, Q.; Bian, Y.; Mi, X. Evaluation of hypoglycemic activity and sub-acute toxicity of the novel biochanin A-chromium(III) complex. Molecules 2022, 27, 5786. [Google Scholar] [CrossRef]
- Almeida, I.M.; Rodrigues, F.; Sarmento, B.; Alves, R.C.; Oliveira, M.B. Isoflavones in food supplements: Chemical profile, label accordance and permeability study in Caco-2 cells. Food Funct. 2015, 6, 938–946. [Google Scholar] [CrossRef]
- Acevedo Martinez, K.A.; Yang, M.M.; Gonzalez de Mejia, E. Technological properties of chickpea (Cicer arietinum): Production of snacks and health benefits related to type-2 diabetes. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3762–3787. [Google Scholar] [CrossRef]
- Rachwa-Rosiak, D.; Nebesny, E.; Budryn, G. Chickpeas-composition, nutritional value, health benefits, application to bread and snacks: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Alarcon-Valdez, C.; Milan-Carrillo, J.; Cardenas-Valenzuela, O.G.; Mora-Escobedo, R.; Bello-Perez, L.A.; Reyes-Moreno, C. Infant food from quality protein maize and chickpea: Optimization for preparing and nutritional properties. Int. J. Food Sci. Nutr. 2005, 56, 273–285. [Google Scholar] [CrossRef]
- Acharjee, S.; Sarmah, B.K. Biotechnologically generating ‘super chickpea’ for food and nutritional security. Plant Sci. 2013, 207, 108–116. [Google Scholar] [CrossRef]
- Herrera, A.C.; Gonzalez de Mejia, E. Feasibility of commercial breadmaking using chickpea as an ingredient: Functional properties and potential health benefits. J. Food Sci. 2021, 86, 2208–2224. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Yang, J.; Pan, Z.; Zhang, G.; Chen, B.; Li, S.; Xiao, J.; Tan, F.; Wang, Z.; Chen, P.; et al. Biochanin A protects against iron overload associated knee osteoarthritis via regulating iron levels and NRF2/System xc-/GPX4 axis. Biomed. Pharmacother. 2023, 157, 113915. [Google Scholar] [CrossRef] [PubMed]
- Hur, H.; Rafii, F. Biotransformation of the isoflavonoids biochanin A, formononetin, and glycitein by Eubacterium limosum. FEMS Microbiol. Lett. 2000, 192, 21–25. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Mechanism | Ref. |
|---|---|---|
| Anti-cancer effect |
| [30] |
| [31] | |
| [32] | |
| [7] | |
| Anti-oxidant effect |
| [33] |
| [34] | |
| [35] | |
| Anti-inflammatory effect |
| [36] |
| [37] | |
| Hypoglycemic effect |
| [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Z.-J.; Lai, W.-F. Chemical and Biological Properties of Biochanin A and Its Pharmaceutical Applications. Pharmaceutics 2023, 15, 1105. https://doi.org/10.3390/pharmaceutics15041105
Feng Z-J, Lai W-F. Chemical and Biological Properties of Biochanin A and Its Pharmaceutical Applications. Pharmaceutics. 2023; 15(4):1105. https://doi.org/10.3390/pharmaceutics15041105
Chicago/Turabian StyleFeng, Zhen-Jie, and Wing-Fu Lai. 2023. "Chemical and Biological Properties of Biochanin A and Its Pharmaceutical Applications" Pharmaceutics 15, no. 4: 1105. https://doi.org/10.3390/pharmaceutics15041105