
The Secretome of Irradiated Peripheral Mononuclear Cells Attenuates Hypertrophic Skin Scarring
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Patient Material
2.3. Animals
2.4. Full-Thickness Wound and Scarring Model in Mice
2.5. Production of Irradiated Mononuclear Cell Secretome (PBMCsec)
2.6. PBMCsec Injection into Mouse Scars
2.7. PBMCsec Topical Application on Mouse Scars
2.8. Ex Vivo Skin and Scar Stimulation
2.9. Skin and Scar PBMCsec Stimulation, Cell Isolation, and Droplet-Based scRNAseq
2.10. Cell–Gene Matrix Preparation and Downstream Analysis
2.11. Gene Ontology (GO) Calculation and Dot Plots
2.12. GSEA Matrisome Dot Plots
2.13. TGFβ Injection Fibrosis Model in Mouse Skin
2.14. Isolation of Primary Skin FBs
2.15. Western Blots
2.16. Immunofluorescence, H&E, and EvG Staining
2.17. TGFβ1-Induced Myofibroblast Differentiation
2.18. Elastase Assay
2.19. ELISA
3. Results
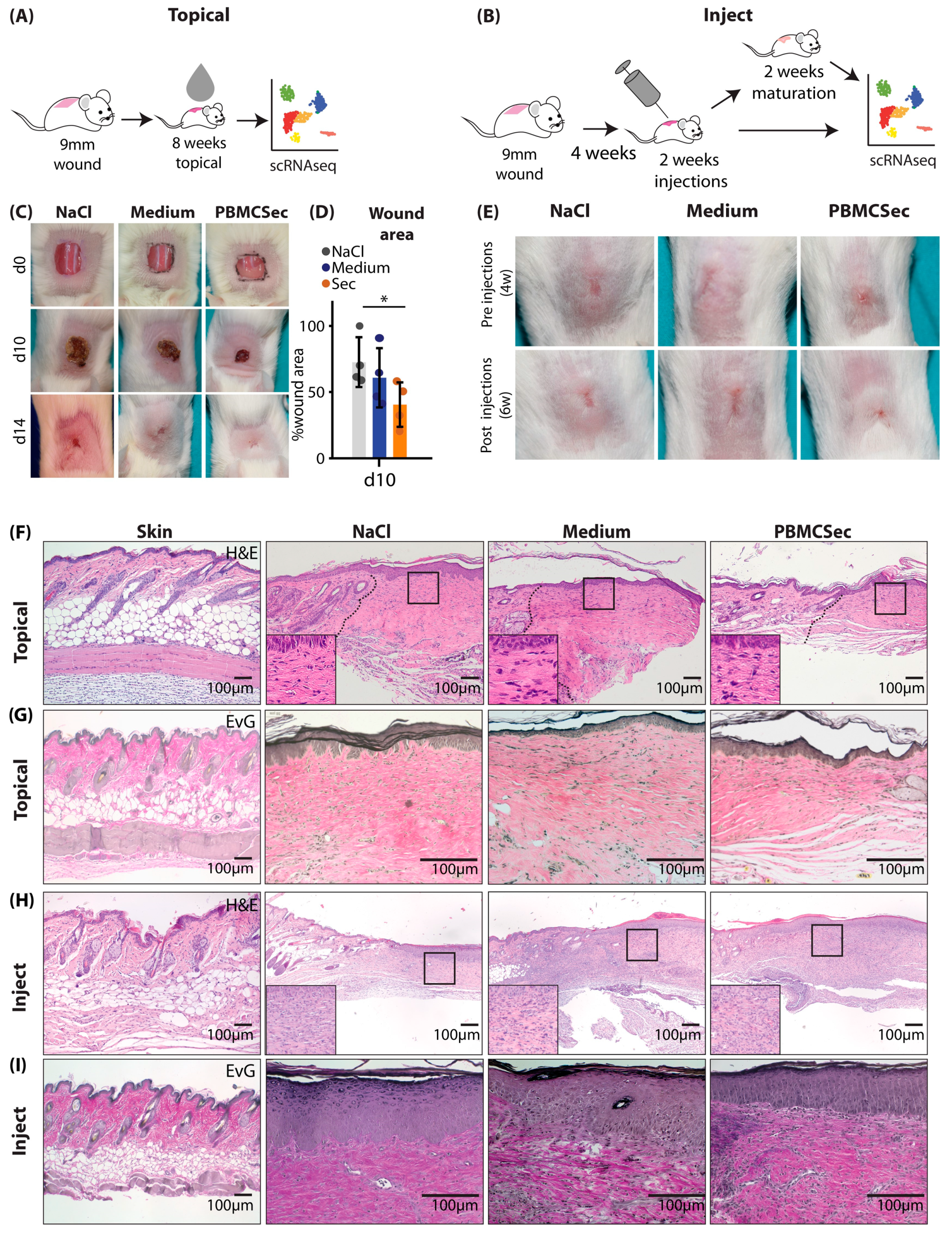
3.1. PBMCsec Improves Scar Formation in Mice after Topical Treatment during Wound Healing and Intradermal Injection of Preformed Scars
3.2. PBMCsec Induces Significant Changes in the Transcriptome after Topical and Intradermal Application
3.3. PBMCsec Significantly Alters the Matrisome
3.4. Scars Treated with PBMCsec Ex Vivo Show Strong Similarities to Mouse Models
3.5. PBMCsec Abolishes Myofibroblast Differentiation In Vitro
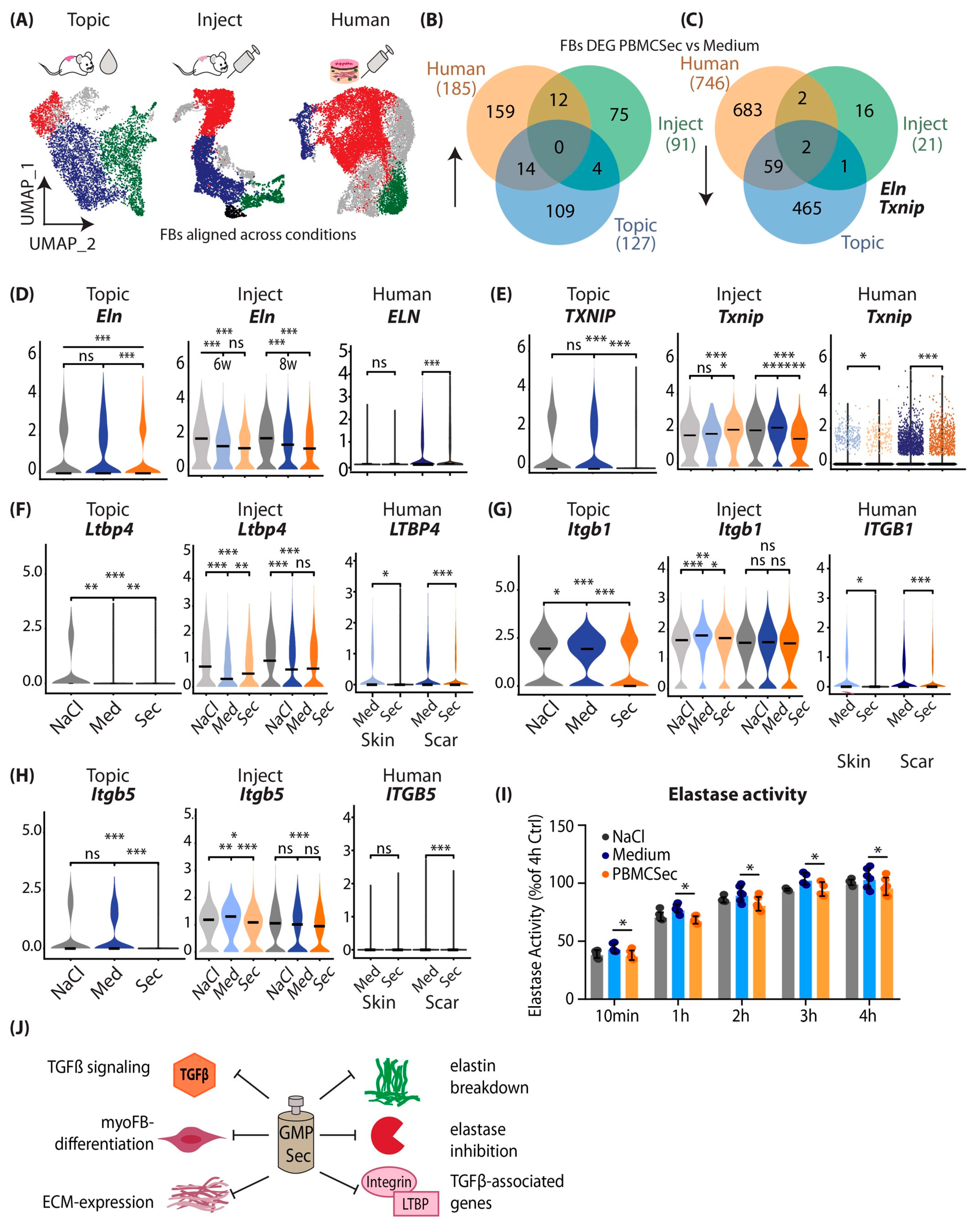
3.6. Combined Analysis of Murine and Human scRNAseq Datasets Reveals Elastin and TXNIP as Joint Key Players of Beneficial PBMCsec Effects
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bayat, A.; McGrouther, D.A.; Ferguson, M.W.J. Skin scarring. BMJ 2003, 326, 88–92. [Google Scholar] [CrossRef]
- Gauglitz, G.G.; Korting, H.C.; Pavicic, T.; Ruzicka, T.; Jeschke, M.G. Hypertrophic scarring and keloids: Pathomechanisms and current and emerging treatment strategies. Mol. Med. 2011, 17, 113–125. [Google Scholar] [CrossRef]
- Bock, O.; Schmid-Ott, G.; Malewski, P.; Mrowietz, U. Quality of life of patients with keloid and hypertrophic scarring. Arch. Dermatol. Res. 2006, 297, 433–438. [Google Scholar] [CrossRef]
- Van Loey, N.E.; Bremer, M.; Faber, A.W.; Middelkoop, E.; Nieuwenhuis, M.K. Itching following burns: Epidemiology and predictors. Br. J. Dermatol. 2008, 158, 95–100. [Google Scholar] [CrossRef]
- Jiang, D.; Rinkevich, Y. Scars or regeneration?-dermal fibroblasts as drivers of diverse skin wound responses. Int. J. Mol. Sci. 2020, 21, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karppinen, S.M.; Heljasvaara, R.; Gullberg, D.; Tasanen, K.; Pihlajaniemi, T. Toward understanding scarless skin wound healing and pathological scarring. F1000Research 2019, 8, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorstandlechner, V.; Laggner, M.; Copic, D.; Klas, K.; Direder, M.; Chen, Y.; Golabi, B.; Haslik, W.; Radtke, C.; Tschachler, E.; et al. The serine proteases dipeptidyl-peptidase 4 and urokinase are key molecules in human and mouse scar formation. Nat. Commun. 2021, 12, 6242. [Google Scholar] [CrossRef] [PubMed]
- Sidgwick, G.P.; McGeorge, D.; Bayat, A. A comprehensive evidence-based review on the role of topicals and dressings in the management of skin scarring. Arch. Dermatol. Res. 2015, 307, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Takeo, M.; Lee, W.; Ito, M. Wound healing and skin regeneration. Cold Spring Harb. Perspect. Med. 2015, 5, a023267. [Google Scholar] [CrossRef]
- Mack, M. Inflammation and fibrosis. Matrix Biol. 2018, 68–69, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef]
- Nabai, L.; Pourghadiri, A.; Ghahary, A. Hypertrophic scarring: Current knowledge of predisposing factors, cellular and molecular mechanisms. J. Burn. Care Res. 2020, 41, 48–56. [Google Scholar] [CrossRef]
- Lee, H.J.; Jang, Y.J. Recent understandings of biology, prophylaxis and treatment strategies for hypertrophic scars and keloids. Int. J. Mol. Sci. 2018, 19, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthonissen, M.; Daly, D.; Janssens, T.; Van den Kerckhove, E. The effects of conservative treatments on burn scars: A systematic review. Burns 2016, 42, 508–518. [Google Scholar] [CrossRef]
- Kafka, M.; Collins, V.; Kamolz, L.P.; Rappl, T.; Branski, L.K.; Wurzer, P. Evidence of invasive and noninvasive treatment modalities for hypertrophic scars: A systematic review. Wound Repair Regen. 2017, 25, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Xu, S.; Pan, Z.; Deng, J.; Li, X.; Pan, F. Comparative efficacy and safety of common therapies in keloids and hypertrophic scars: A systematic review and meta-analysis. Aesthetic Plast. Surg. 2019, 44, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Sato, C.; Yamamoto, Y.; Funayama, E.; Furukawa, H.; Oyama, A.; Murao, N.; Hosono, H.; Kawakubo, K.; Sakamoto, N.; Ohnishi, S. Conditioned medium obtained from amnion-derived mesenchymal stem cell culture prevents activation of keloid fibroblasts. Plast. Reconstr. Surg. 2018, 141, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Z.; Huang, Z.; Liang, L.; Chen, M. Chyle fat-derived stem cells conditioned medium inhibits hypertrophic scar fibroblast activity. Ann. Plast. Surg. 2019, 83, 271–277. [Google Scholar] [CrossRef]
- Hu, C.H.; Tseng, Y.W.; Chiou, C.Y.; Lan, K.C.; Chou, C.H.; Tai, C.S.; Huang, H.D.; Hu, C.W.; Liao, K.H.; Chuang, S.S.; et al. Bone marrow concentrate-induced mesenchymal stem cell conditioned medium facilitates wound healing and prevents hypertrophic scar formation in a rabbit ear model. Stem Cell Res. Ther. 2019, 10, 275. [Google Scholar] [CrossRef]
- Ren, Y.; Deng, C.L.; Wan, W.D.; Zheng, J.H.; Mao, G.Y.; Yang, S.L. Suppressive effects of induced pluripotent stem cell-conditioned medium on in vitro hypertrophic scarring fibroblast activation. Mol. Med. Rep. 2015, 11, 2471–2476. [Google Scholar] [CrossRef] [Green Version]
- Kanji, S.; Das, H. Advances of stem cell therapeutics in cutaneous wound healing and regeneration. Mediat. Inflamm. 2017, 2017, 5217967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bojanic, C.; To, K. Mesenchymal stem cell therapy in hypertrophic and keloid scars. Cell Tissue Res. 2021, 383, 915–930. [Google Scholar] [CrossRef] [PubMed]
- Beer, L.; Mildner, M.; Ankersmit, H.J. Cell secretome based drug substances in regenerative medicine: When regulatory affairs meet basic science. Ann. Transl. Med. 2017, 5, 170. [Google Scholar] [CrossRef] [Green Version]
- Korf-Klingebiel, M.; Kempf, T.; Sauer, T.; Brinkmann, E.; Fischer, P.; Meyer, G.P.; Ganser, A.; Drexler, H.; Wollert, K.C. Bone marrow cells are a rich source of growth factors and cytokines: Implications for cell therapy trials after myocardial infarction. Eur. Heart J. 2008, 29, 2851–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mildner, M.; Hacker, S.; Haider, T.; Gschwandtner, M.; Werba, G.; Barresi, C.; Zimmermann, M.; Golabi, B.; Tschachler, E.; Ankersmit, H.J. Secretome of peripheral blood mononuclear cells enhances wound healing. PLoS ONE 2013, 8, e60103. [Google Scholar] [CrossRef] [Green Version]
- Wagner, T.; Traxler, D.; Simader, E.; Beer, L.; Narzt, M.-S.; Gruber, F.; Madlener, S.; Laggner, M.; Erb, M.; Vorstandlechner, V.; et al. Different pro-angiogenic potential of γ-irradiated pbmc-derived secretome and its subfractions. Sci. Rep. 2018, 8, 18016. [Google Scholar] [CrossRef] [Green Version]
- Hacker, S.; Mittermayr, R.; Nickl, S.; Haider, T.; Lebherz-Eichinger, D.; Beer, L.; Mitterbauer, A.; Leiss, H.; Zimmermann, M.; Schweiger, T.; et al. Paracrine factors from irradiated peripheral blood mononuclear cells improve skin regeneration and angiogenesis in a porcine burn model. Sci. Rep. 2016, 6, 25168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simader, E.; Beer, L.; Laggner, M.; Vorstandlechner, V.; Gugerell, A.; Erb, M.; Kalinina, P.; Copic, D.; Moser, D.; Spittler, A.; et al. Tissue-regenerative potential of the secretome of gamma-irradiated peripheral blood mononuclear cells is mediated via tnfrsf1b-induced necroptosis. Cell Death Dis. 2019, 10, 729. [Google Scholar] [CrossRef] [Green Version]
- Hoetzenecker, K.; Assinger, A.; Lichtenauer, M.; Mildner, M.; Schweiger, T.; Starlinger, P.; Jakab, A.; Berényi, E.; Pavo, N.; Zimmermann, M.; et al. Secretome of apoptotic peripheral blood cells (aposec) attenuates microvascular obstruction in a porcine closed chest reperfused acute myocardial infarction model: Role of platelet aggregation and vasodilation. Basic Res. Cardiol. 2012, 107, 292. [Google Scholar] [CrossRef] [Green Version]
- Kasiri, M.M.; Beer, L.; Nemec, L.; Gruber, F.; Pietkiewicz, S.; Haider, T.; Simader, E.M.; Traxler, D.; Schweiger, T.; Janik, S.; et al. Dying blood mononuclear cell secretome exerts antimicrobial activity. Eur. J. Clin. Investig. 2016, 46, 853–863. [Google Scholar] [CrossRef]
- Altmann, P.; Mildner, M.; Haider, T.; Traxler, D.; Beer, L.; Ristl, R.; Golabi, B.; Gabriel, C.; Leutmezer, F.; Ankersmit, H.J. Secretomes of apoptotic mononuclear cells ameliorate neurological damage in rats with focal ischemia. F1000Research 2014, 3, 131. [Google Scholar] [CrossRef] [PubMed]
- Haider, T.; Hoftberger, R.; Ruger, B.; Mildner, M.; Blumer, R.; Mitterbauer, A.; Buchacher, T.; Sherif, C.; Altmann, P.; Redl, H.; et al. The secretome of apoptotic human peripheral blood mononuclear cells attenuates secondary damage following spinal cord injury in rats. Exp. Neurol. 2015, 267, 230–242. [Google Scholar] [CrossRef] [Green Version]
- Ankersmit, H.J.; Hoetzenecker, K.; Dietl, W.; Soleiman, A.; Horvat, R.; Wolfsberger, M.; Gerner, C.; Hacker, S.; Mildner, M.; Moser, B.; et al. Irradiated cultured apoptotic peripheral blood mononuclear cells regenerate infarcted myocardium. Eur. J. Clin. Investig. 2009, 39, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Laggner, M.; Acosta, G.S.; Kitzmüller, C.; Copic, D.; Gruber, F.; Altenburger, L.M.; Vorstandlechner, V.; Gugerell, A.; Direder, M.; Klas, K.; et al. The secretome of irradiated peripheral blood mononuclear cells attenuates activation of mast cells and basophils. EBioMedicine 2022, 81, 104093. [Google Scholar] [CrossRef] [PubMed]
- Laggner, M.; Copic, D.; Nemec, L.; Vorstandlechner, V.; Gugerell, A.; Gruber, F.; Peterbauer, A.; Ankersmit, H.J.; Mildner, M. Therapeutic potential of lipids obtained from γ-irradiated pbmcs in dendritic cell-mediated skin inflammation. EBioMedicine 2020, 55, 102774. [Google Scholar] [CrossRef]
- Simader, E.; Traxler, D. Safety and tolerability of topically administered autologous, apoptotic pbmc secretome (APOSEC) in dermal wounds: A randomized phase 1 trial (MARSYAS I). Sci. Rep. 2017, 7, 6216. [Google Scholar] [CrossRef] [Green Version]
- Gugerell, A.; Gouya-Lechner, G.; Hofbauer, H.; Laggner, M.; Trautinger, F.; Almer, G.; Peterbauer-Scherb, A.; Seibold, M.; Hoetzenecker, W.; Dreschl, C.; et al. Safety and clinical efficacy of the secretome of stressed peripheral blood mononuclear cells in patients with diabetic foot ulcer—Study protocol of the randomized, placebo-controlled, double-blind, multicenter, international phase ii clinical trial marsyas ii. Trials 2021, 22, 10. [Google Scholar]
- Fearmonti, R.M.; Bond, J.E.; Erdmann, D.; Levin, L.S.; Pizzo, S.V.; Levinson, H. The modified patient and observer scar assessment scale: A novel approach to defining pathologic and nonpathologic scarring. Plast. Reconstr. Surg. 2011, 127, 242–247. [Google Scholar] [CrossRef] [Green Version]
- Laggner, M.; Gugerell, A.; Bachmann, C.; Hofbauer, H.; Vorstandlechner, V.; Seibold, M.; Gouya Lechner, G.; Peterbauer, A.; Madlener, S.; Demyanets, S.; et al. Reproducibility of gmp-compliant production of therapeutic stressed peripheral blood mononuclear cell-derived secretomes, a novel class of biological medicinal products. Stem Cell Res. Ther. 2020, 11, 9. [Google Scholar] [CrossRef] [Green Version]
- Gugerell, A.; Sorgenfrey, D.; Laggner, M.; Raimann, J.; Peterbauer, A.; Bormann, D.; Suessner, S.; Gabriel, C.; Moser, B.; Ostler, T.; et al. Viral safety of aposec. Blood Transfus. 2019, 18, 30–39. [Google Scholar]
- Stuart, T.; Butler, A.; Hoffman, P.; Hafemeister, C.; Papalexi, E.; Mauck, W.M., 3rd; Hao, Y.; Stoeckius, M.; Smibert, P.; Satija, R. Comprehensive integration of single-cell data. Cell 2019, 177, 1888–1902.e1821. [Google Scholar] [CrossRef]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411. [Google Scholar] [CrossRef] [PubMed]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly parallel genome-wide expression profiling of individual cells using nanoliter droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [Green Version]
- Luecken, M.D.; Theis, F.J. Current best practices in single-cell rna-seq analysis: A tutorial. Mol. Syst. Biol. 2019, 15, e8746. [Google Scholar] [CrossRef]
- Rojahn, T.B.; Vorstandlechner, V.; Krausgruber, T.; Bauer, W.M.; Alkon, N.; Bangert, C.; Thaler, F.M.; Sadeghyar, F.; Fortelny, N.; Gernedl, V.; et al. Single-cell transcriptomics combined with interstitial fluid proteomics defines cell type-specific immune regulation in atopic dermatitis. J. Allergy Clin. Immunol. 2020, 146, 1056–1069. [Google Scholar] [CrossRef]
- Hammond, T.R.; Dufort, C.; Dissing-Olesen, L.; Giera, S.; Young, A.; Wysoker, A.; Walker, A.J.; Gergits, F.; Segel, M.; Nemesh, J.; et al. Single-cell rna sequencing of microglia throughout the mouse lifespan and in the injured brain reveals complex cell-state changes. Immunity 2019, 50, 253–271.e256. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabib, T.; Morse, C.; Wang, T.; Chen, W.; Lafyatis, R. Sfrp2/dpp4 and fmo1/lsp1 define major fibroblast populations in human skin. J. Investig. Dermatol. 2017, 138, 802–810. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, Y.; Min, P. Single-cell and bulk transcriptome data integration reveals dysfunctional cell types and aberrantly expressed genes in hypertrophic scar. Front. Genet. 2021, 12, 806740. [Google Scholar] [CrossRef]
- Ma, L.; Gan, C.; Huang, Y.; Wang, Y.; Luo, G.; Wu, J. Comparative proteomic analysis of extracellular matrix proteins secreted by hypertrophic scar with normal skin fibroblasts. Burn. Trauma 2014, 2, 76–83. [Google Scholar]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The extracellular matrix: Tools and insights for the “omics” era. Matrix Biol. 2016, 49, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Direder, M.; Weiss, T.; Copic, D.; Vorstandlechner, V.; Laggner, M.; Pfisterer, K.; Mildner, C.S.; Klas, K.; Bormann, D.; Haslik, W.; et al. Schwann cells contribute to keloid formation. Matrix Biol. 2022, 108, 55–76. [Google Scholar] [CrossRef] [PubMed]
- Thielitz, A.; Vetter, R.W.; Schultze, B.; Wrenger, S.; Simeoni, L.; Ansorge, S.; Neubert, K.; Faust, J.; Lindenlaub, P.; Gollnick, H.P.; et al. Inhibitors of dipeptidyl peptidase iv-like activity mediate antifibrotic effects in normal and keloid-derived skin fibroblasts. J. Investig. Dermatol. 2008, 128, 855–866. [Google Scholar] [CrossRef] [Green Version]
- Hinz, B. Myofibroblasts. Exp. Eye Res. 2016, 142, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Alanazi, Y.F. Elastic fibre proteins in elastogenesis and wound healing. Int. J. Mol. Sci. 2022, 23, 4087. [Google Scholar] [CrossRef]
- Khan, Z.; Marshall, J.F. The role of integrins in tgfβ activation in the tumour stroma. Cell Tissue Res. 2016, 365, 657–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.T.; Urban, Z. Ltbp4 in health and disease. Genes 2021, 12, 795. [Google Scholar] [CrossRef] [PubMed]
- Godwin, A.R.F.; Singh, M.; Lockhart-Cairns, M.P.; Alanazi, Y.F.; Cain, S.A.; Baldock, C. The role of fibrillin and microfibril binding proteins in elastin and elastic fibre assembly. Matrix Biol. 2019, 84, 17–30. [Google Scholar] [CrossRef]
- Lu, J.; Liu, Q.; Wang, L.; Tu, W.; Chu, H.; Ding, W.; Jiang, S.; Ma, Y.; Shi, X.; Pu, W.; et al. Increased expression of latent tgf-β-binding protein 4 affects the fibrotic process in scleroderma by tgf-β/smad signaling. Lab. Investig. 2017, 97, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Buczek-Thomas, J.A.; Lucey, E.C.; Stone, P.J.; Chu, C.L.; Rich, C.B.; Carreras, I.; Goldstein, R.H.; Foster, J.A.; Nugent, M.A. Elastase mediates the release of growth factors from lung in vivo. Am. J. Respir. Cell Mol. Biol. 2004, 31, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Arjunan, S.; Gan, S.U.; Choolani, M.; Raj, V.; Lim, J.; Biswas, A.; Bongso, A.; Fong, C.Y. Inhibition of growth of asian keloid cells with human umbilical cord wharton′s jelly stem cell-conditioned medium. Stem Cell Res. Ther. 2020, 11, 78. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.H.; Tseng, Y.W.; Lee, C.W.; Chiou, C.Y.; Chuang, S.S.; Yang, J.Y.; Lee, O.K. Combination of mesenchymal stem cell-conditioned medium and botulinum toxin type a for treating human hypertrophic scars. J. Plast. Reconstr. Aesthetic Surg. 2020, 73, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ren, J.; Su, L.; Cheng, S.; Zhou, J.; Ye, X.; Dong, Y.; Sun, S.; Qi, F.; Liu, Z.; et al. Human adipose tissue-derived stem cells inhibit the activity of keloid fibroblasts and fibrosis in a keloid model by paracrine signaling. Burns 2018, 44, 370–385. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, S.; Mancini, E.; Xu, L.; Moore, A.; Jahanbani, F.; Hebestreit, K.; Srinivasan, R.; Li, X.; Devarajan, K.; Prélot, L. , et al. Heterogeneity in old fibroblasts is linked to variability in reprogramming and wound healing. Nature 2019, 574, 553–558. [Google Scholar] [CrossRef]
- Solé-Boldo, L.; Raddatz, G.; Schütz, S.; Mallm, J.-P.; Rippe, K.; Lonsdorf, A.S.; Rodríguez-Paredes, M.; Lyko, F. Single-cell transcriptomes of the human skin reveal age-related loss of fibroblast priming. Commun. Biol. 2020, 3, 188. [Google Scholar] [CrossRef] [Green Version]
- Joost, S.; Annusver, K.; Jacob, T.; Sun, X.; Dalessandri, T.; Sivan, U.; Sequeira, I.; Sandberg, R.; Kasper, M. The molecular anatomy of mouse skin during hair growth and rest. Cell Stem Cell 2020, 26, 441–457.e447. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, C.Y.; Yang, S.C.; Alalaiwe, A.; Fang, J.Y. Laser ablation and topical drug delivery: A review of recent advances. Expert Opin. Drug Deliv. 2019, 16, 937–952. [Google Scholar] [CrossRef] [PubMed]
- Aich, K.; Singh, T.; Dang, S. Advances in microneedle-based transdermal delivery for drugs and peptides. Drug Deliv. Transl. Res. 2022, 12, 1556–1568. [Google Scholar] [CrossRef]
- Qu, F.; Geng, R.; Liu, Y.; Zhu, J. Advanced nanocarrier- and microneedle-based transdermal drug delivery strategies for skin diseases treatment. Theranostics 2022, 12, 3372–3406. [Google Scholar] [CrossRef]
- Naba, A.; Clauser, K.R.; Hoersch, S.; Liu, H.; Carr, S.A.; Hynes, R.O. The matrisome: In silico definition and in vivo characterization by proteomics of normal and tumor extracellular matrices. Mol. Cell. Proteom. 2012, 11, M111.014647. [Google Scholar] [CrossRef] [Green Version]
- Tellman, T.V.; Dede, M.; Aggarwal, V.A.; Salmon, D.; Naba, A.; Farach-Carson, M.C. Systematic analysis of actively transcribed core matrisome genes across tissues and cell phenotypes. Matrix Biol. 2022, 111, 95–107. [Google Scholar] [CrossRef]
- The biology of extracellular matrix proteins in hypertrophic scarring. Adv. Wound Care 2022, 11, 234–254. [CrossRef] [PubMed]
- Mithieux, S.M.; Weiss, A.S. Elastin. Adv. Protein Chem. 2005, 70, 437–461. [Google Scholar] [PubMed]
- Heinz, A. Elastases and elastokines: Elastin degradation and its significance in health and disease. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 252–273. [Google Scholar] [CrossRef]
- Copic, D.; Direder, M.; Schossleitner, K.; Laggner, M.; Klas, K.; Bormann, D.; Ankersmit, H.J.; Mildner, M. Paracrine factors of stressed peripheral blood mononuclear cells activate proangiogenic and anti-proteolytic processes in whole blood cells and protect the endothelial barrier. Pharmaceutics 2022, 14, 1600. [Google Scholar] [CrossRef] [PubMed]
- Zani, M.L.; Tanga, A.; Saidi, A.; Serrano, H.; Dallet-Choisy, S.; Baranger, K.; Moreau, T. Slpi and trappin-2 as therapeutic agents to target airway serine proteases in inflammatory lung diseases: Current and future directions. Biochem. Soc. Trans. 2011, 39, 1441–1446. [Google Scholar] [CrossRef]
- Zhou, J.; Chng, W.J. Roles of thioredoxin binding protein (TXNIP) in oxidative stress, apoptosis and cancer. Mitochondrion 2013, 13, 163–169. [Google Scholar] [CrossRef]
- Saad, I.; Fournier, C.T.; Wilson, R.L.; Lakshmanan, R.; Selvaraju, V.; Thirunavukkarasu, M.; Alexander Palesty, J.; McFadden, D.W.; Maulik, N. Thioredoxin-1 augments wound healing and promote angiogenesis in a murine ischemic full-thickness wound model. Surgery 2018, 164, 1077–1086. [Google Scholar] [CrossRef]
- Han, Y.Y.; Gu, X.; Yang, C.Y.; Ji, H.M.; Lan, Y.J.; Bi, Y.Q.; Si, R.; Qu, J.; Cheng, M.H.; Gao, J. Protective effect of dimethyl itaconate against fibroblast-myofibroblast differentiation during pulmonary fibrosis by inhibiting TXNIP. J. Cell. Physiol. 2021, 236, 7734–7744. [Google Scholar] [CrossRef]
- Hsiao, P.F.; Huang, Y.T.; Lu, P.H.; Chiu, L.Y.; Weng, T.H.; Hung, C.F.; Wu, N.L. Thioredoxin-interacting protein regulates keratinocyte differentiation: Implication of its role in psoriasis. FASEB J. 2022, 36, e22313. [Google Scholar] [CrossRef]
- Popova, A.P.; Bozyk, P.D.; Goldsmith, A.M.; Linn, M.J.; Lei, J.; Bentley, J.K.; Hershenson, M.B. Autocrine production of tgf-beta1 promotes myofibroblastic differentiation of neonatal lung mesenchymal stem cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 298, L735–L743. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhang, Q.; Gao, Y.; Tan, M.; Zheng, R.; Zhao, L.; Zhang, X. Induced pluripotent stem cell-conditioned medium suppresses pulmonary fibroblast-to-myofibroblast differentiation via the inhibition of tgf-β1/smad pathway. Int. J. Mol. Med. 2018, 41, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. Tgf-β: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Akboua, H.; Eghbalzadeh, K.; Keser, U.; Wahlers, T.; Paunel-Görgülü, A. Impaired non-canonical transforming growth factor-β signalling prevents profibrotic phenotypes in cultured peptidylarginine deiminase 4-deficient murine cardiac fibroblasts. J. Cell. Mol. Med. 2021, 25, 9674–9684. [Google Scholar] [CrossRef]
- Kim, D.-Y.; Lee, S.-H.; Fu, Y.; Jing, F.; Kim, W.-Y.; Hong, S.-B.; Song, J.-A.; Choe, H.; Ryu, H.J.; Kim, M.; et al. Del-1, an endogenous inhibitor of tgf-β activation, attenuates fibrosis. Front. Immunol. 2020, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Zomer, H.D.; Trentin, A.G. Skin wound healing in humans and mice: Challenges in translational research. J. Dermatol. Sci. 2018, 90, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound repair and regeneration: Mechanisms, signaling, and translation. Sci. Transl. Med. 2014, 6, 265sr6. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Mirza, R.; Kwon, Y.; DiPietro, L.A.; Koh, T.J. The murine excisional wound model: Contraction revisited. Wound Repair Regen. 2015, 23, 874–877. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.M.; Adams, D.H.; Greenwood, J.E.; Anderson, P.J.; Cowin, A.J. A novel murine model of hypertrophic scarring using subcutaneous infusion of bleomycin. Plast. Reconstr. Surg. 2014, 133, 69–78. [Google Scholar] [CrossRef]
- Seo, B.F.; Lee, J.Y.; Jung, S.N. Models of abnormal scarring. BioMed Res. Int. 2013, 2013, 423147. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vorstandlechner, V.; Copic, D.; Klas, K.; Direder, M.; Golabi, B.; Radtke, C.; Ankersmit, H.J.; Mildner, M. The Secretome of Irradiated Peripheral Mononuclear Cells Attenuates Hypertrophic Skin Scarring. Pharmaceutics 2023, 15, 1065. https://doi.org/10.3390/pharmaceutics15041065
Vorstandlechner V, Copic D, Klas K, Direder M, Golabi B, Radtke C, Ankersmit HJ, Mildner M. The Secretome of Irradiated Peripheral Mononuclear Cells Attenuates Hypertrophic Skin Scarring. Pharmaceutics. 2023; 15(4):1065. https://doi.org/10.3390/pharmaceutics15041065
Chicago/Turabian StyleVorstandlechner, Vera, Dragan Copic, Katharina Klas, Martin Direder, Bahar Golabi, Christine Radtke, Hendrik J. Ankersmit, and Michael Mildner. 2023. "The Secretome of Irradiated Peripheral Mononuclear Cells Attenuates Hypertrophic Skin Scarring" Pharmaceutics 15, no. 4: 1065. https://doi.org/10.3390/pharmaceutics15041065