
Prenatal Exposure to Δ9-Tetrahydrocannabinol Affects Hippocampus-Related Cognitive Functions in the Adolescent Rat Offspring: Focus on Specific Markers of Neuroplasticity
 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
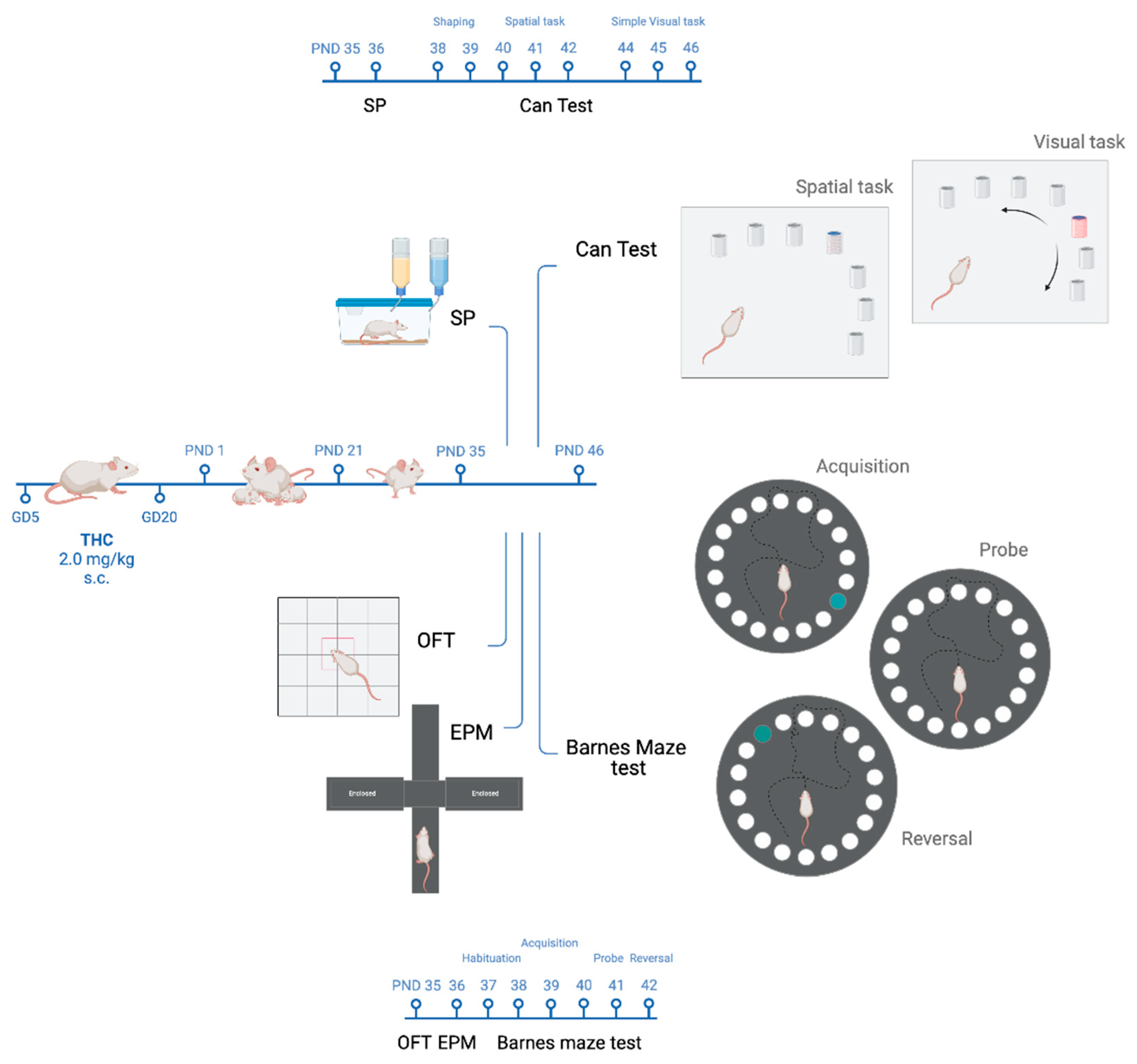
2.1. Animals and Treatment
2.2. Drugs
2.3. Behavioural Procedures
2.4. Open Field Test
2.5. Elevated Plus Maze Test
2.6. Sucrose Preference Test
2.7. Can Test
2.7.1. Experimental Design
Shaping Period
Spatial Task
Simple Visual Task
2.8. Barnes Maze Test
2.8.1. Experimental Design
Habituation
Acquisition Phase
Probe Task
Reversal Task
2.9. Tissue Collection and qRT-PCR Procedure
2.10. Statistical Analysis
3. Results
3.1. Behavioural Assessment of the Offspring
3.1.1. Prenatal THC Exposure Increases Locomotor Activity and Does Not Modify Exploration in the Adolescent Offspring
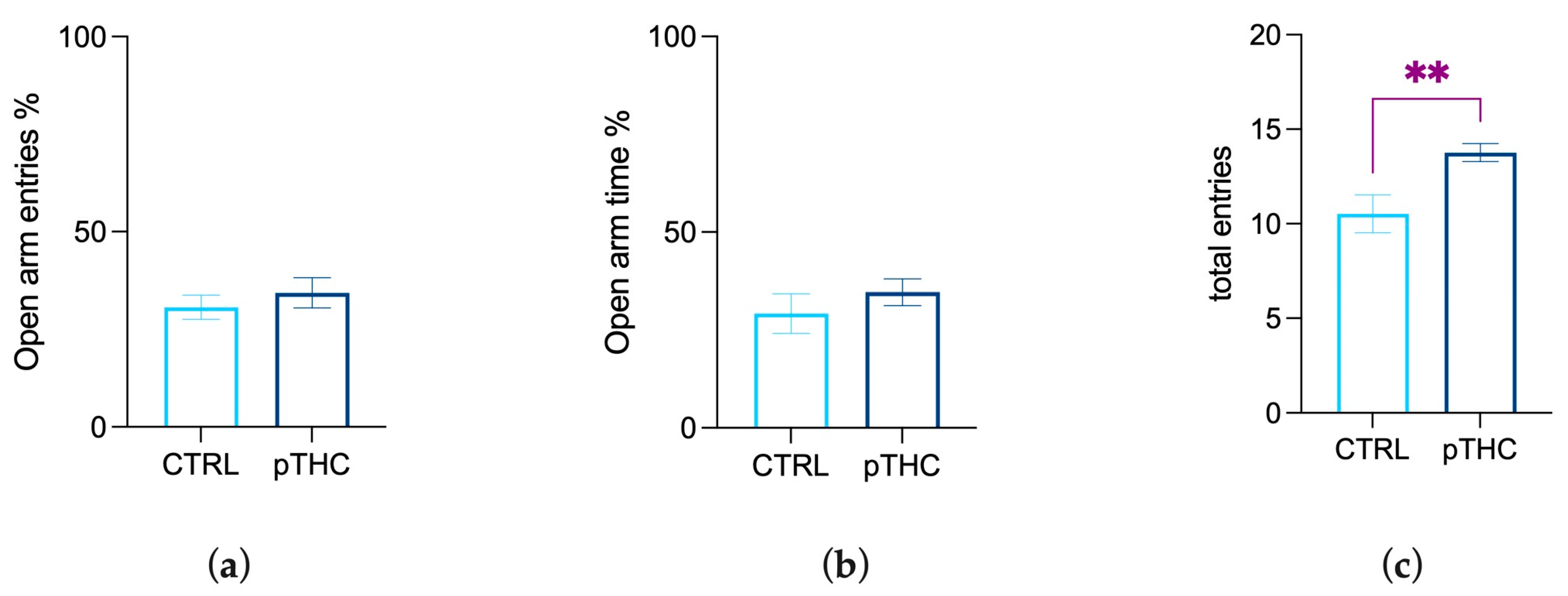
3.1.2. Prenatal THC Exposure Does Not Impair Emotional Reactivity in the Adolescent Offspring
3.1.3. Prenatal THC Exposure Does Not Impact the Responsivity to Natural Reinforcing Stimuli
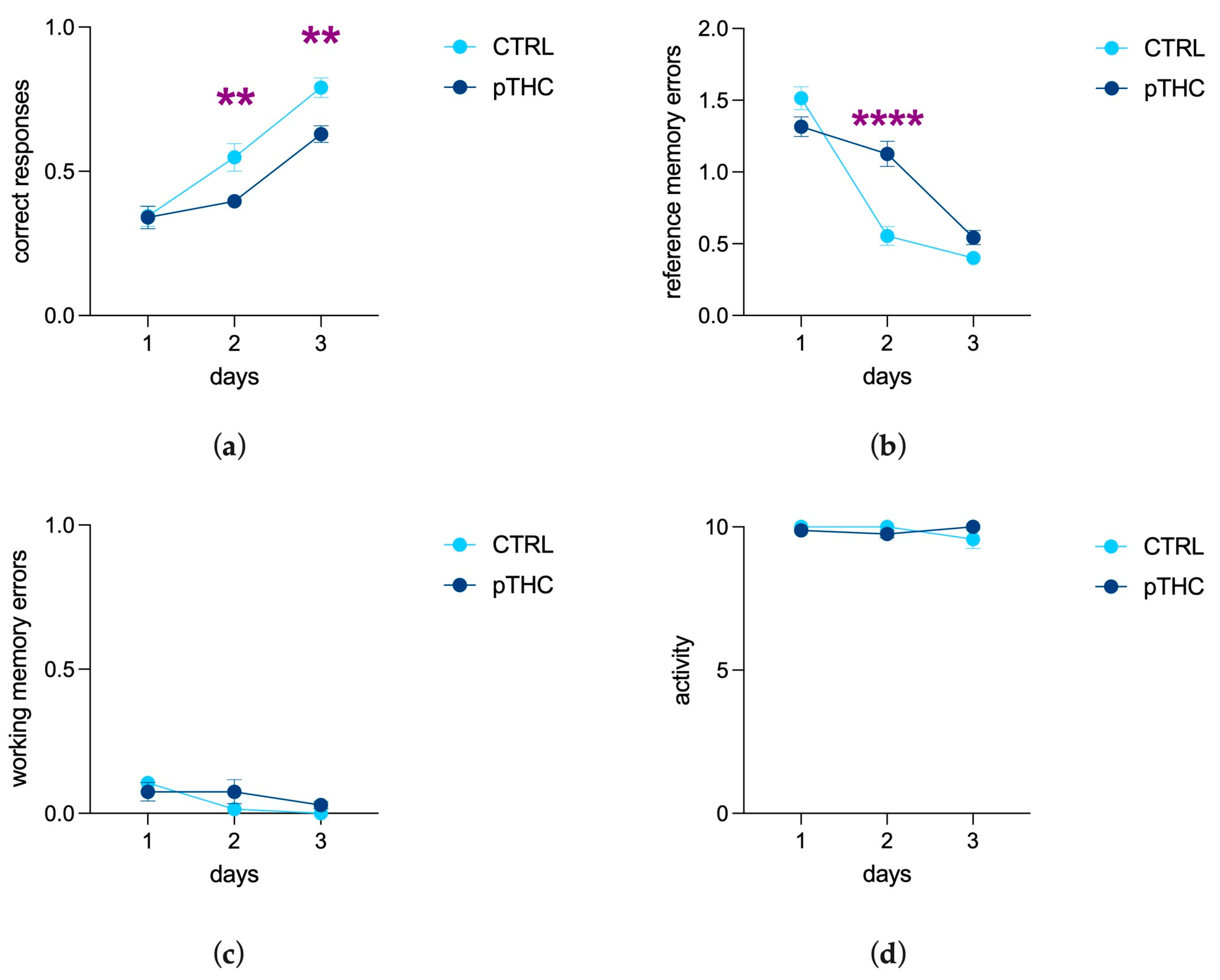
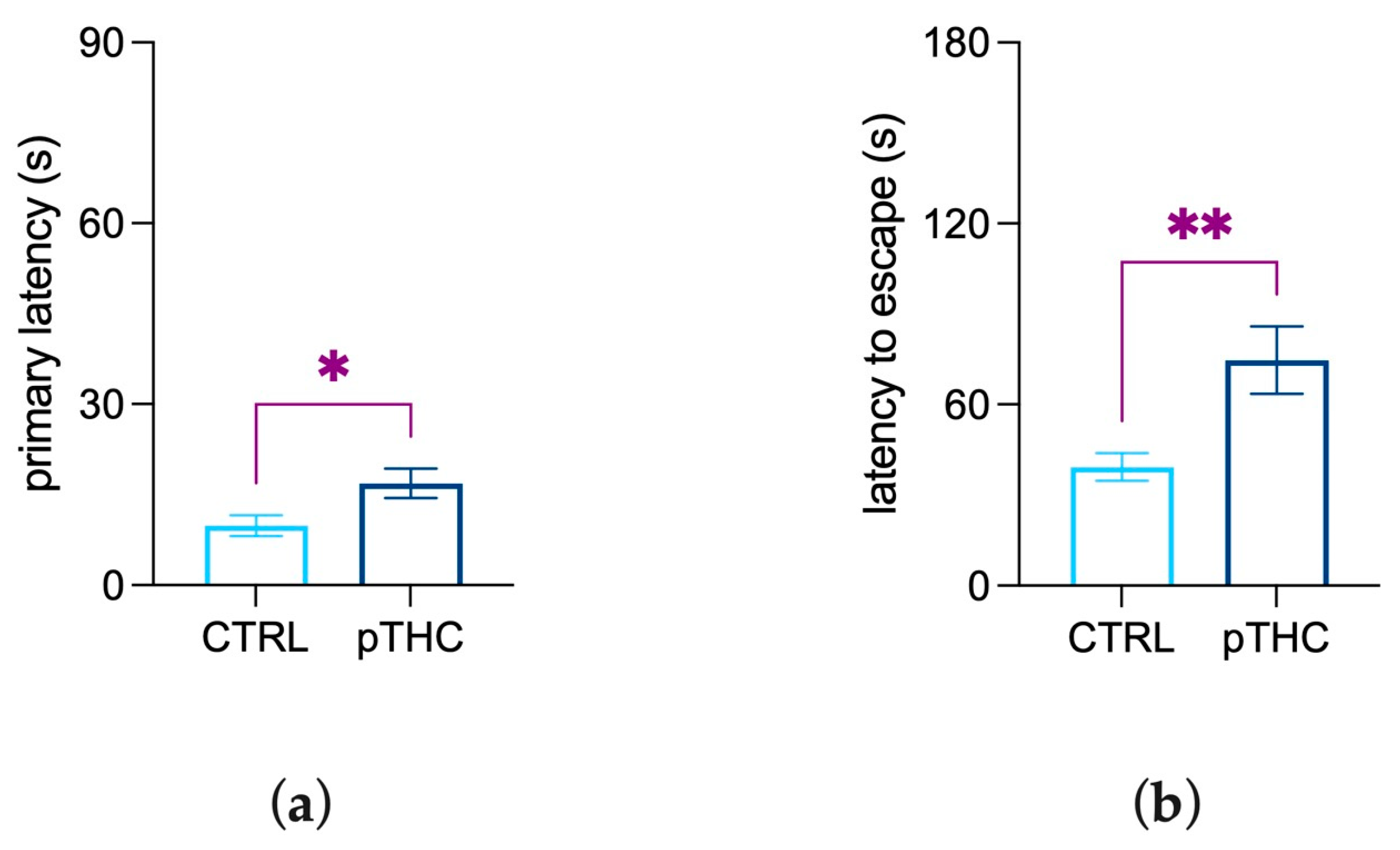
3.1.4. Prenatal THC Exposure Impairs Spatial Memory and Object Recognition Memory of the Adolescent Offspring in a Reinforce-Motivated Can Test
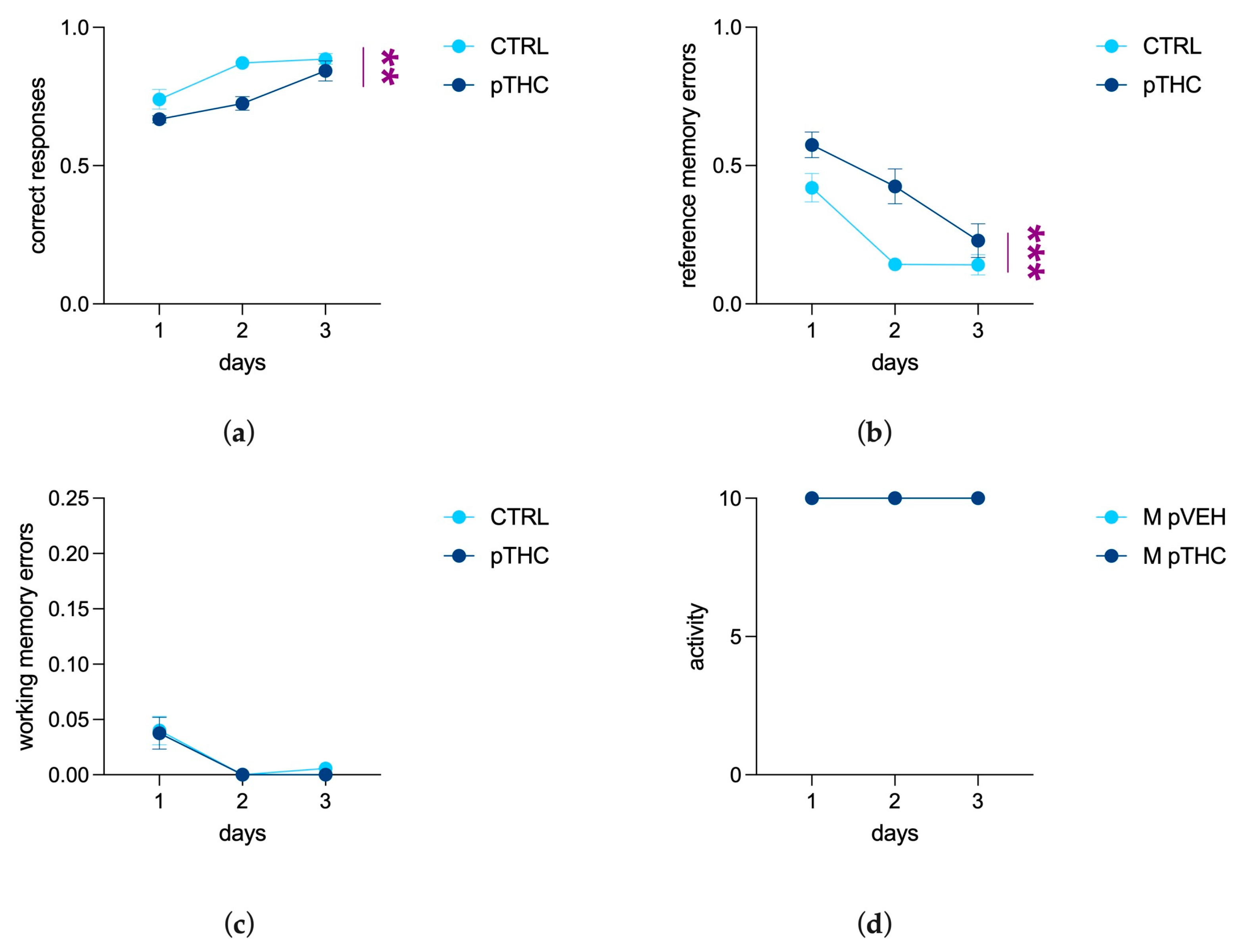
3.1.5. Prenatal THC Exposure Impairs Spatial Memory and Reversal Learning in an Aversive Context
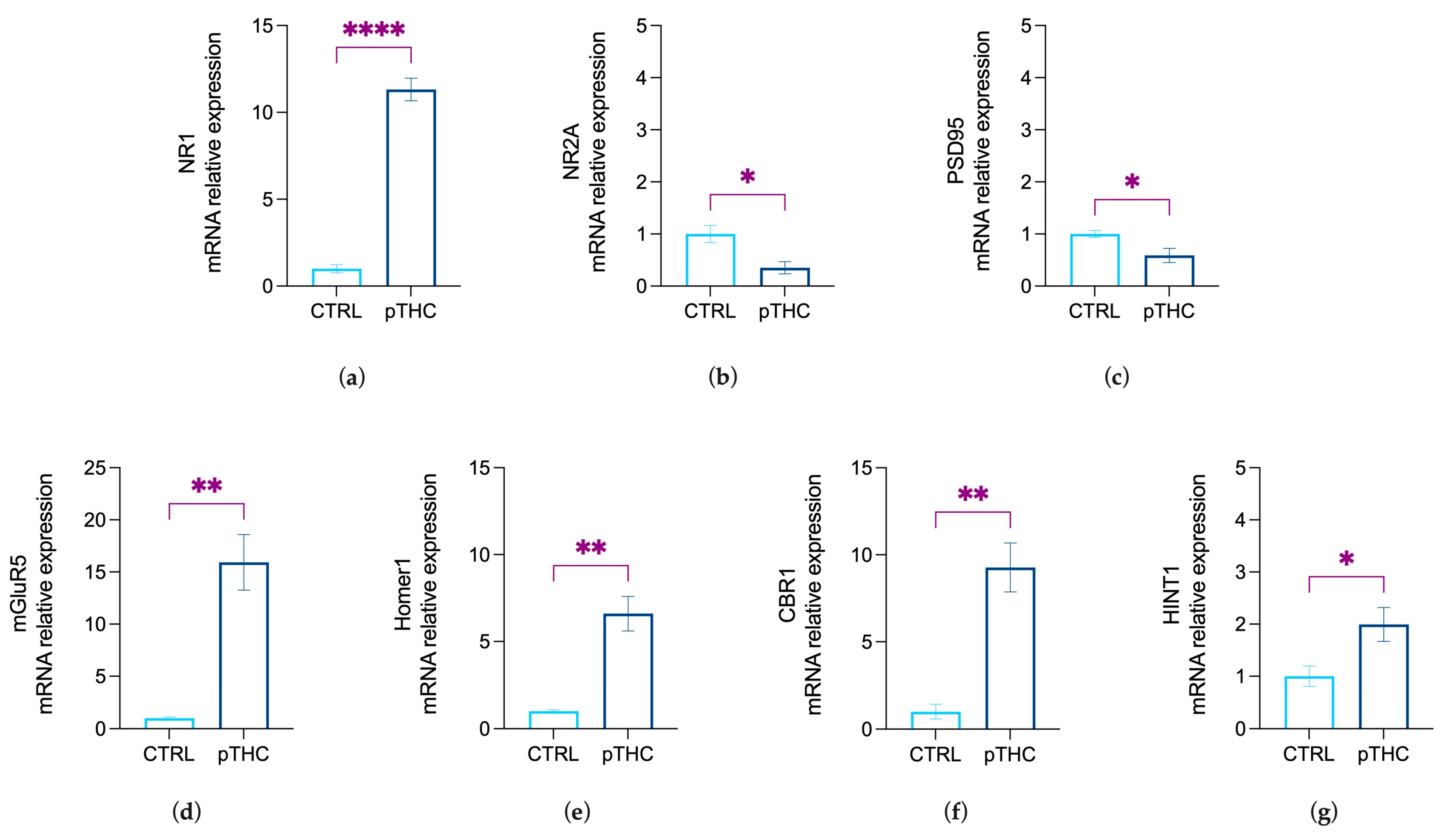
3.2. Prenatal THC Exposure Modifies Markers of Synaptic Plasticity in the Hippocampus of Adolescent Offspring
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Metz, T.D.; Borgelt, L.M. Marijuana Use in Pregnancy and While Breastfeeding. Obstet. Gynecol. 2018, 132, 1198–1210. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, K.L.; Day, N.L.; Rubio, D.; Jarlenski, M.; Arnold, R.M. Beliefs and attitudes regarding prenatal marijuana use: Perspectives of pregnant women who report use. Drug Alcohol Depend. 2019, 196, 14–20. [Google Scholar]
- Fitzcharles, M.A.; Eisenberg, E. Medical cannabis: A forward vision for the clinician. Eur. J. Pain 2018, 22, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klumpers, L.E.; Thacker, D.L. A Brief Background on Cannabis: From Plant to Medical Indications. J. AOAC Int. 2019, 102, 412–420. [Google Scholar] [CrossRef]
- Weisman, J.M.; Rodríguez, M. A systematic review of medical students’ and professionals’ attitudes and knowledge regarding medical cannabis. J. Cannabis. Res. 2021, 3, 47. [Google Scholar] [CrossRef]
- Romero, J.; Garcia-Palomero, E.; Berrendero, F.; Garcia-Gil, L.; Hernandez, M.L.; Ramos, J.A.; Fernández-Ruiz, J.J. Atypical location of cannabinoid receptors in white matter areas during rat brain development. Synapse 1997, 26, 317–323. [Google Scholar] [CrossRef]
- Berrendero, F.; García-Gil, L.; Hernández, M.L.; Romero, J.; Cebeira, M.; de Miguel, R.; Ramos, J.A.; Fernández-Ruiz, J.J. Localization of mRNA expression and activation of signal transduction mechanisms for cannabinoid receptor in rat brain during fetal development. Development 1998, 125, 3179–3188. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Berrendero, F.; Hernández, M.L.; Ramos, J.A. The endogenous cannabinoid system and brain development. Trends Neurosci. 2000, 23, 14–20. [Google Scholar] [CrossRef]
- Fride, E.; Gobshtis, N.; Dahan, H.; Weller, A.; Giuffrida, A.; Ben-Shabat, S. The endocannabinoid system during development: Emphasis on perinatal events and delayed effects. Vitam. Horm. 2009, 81, 139–158. [Google Scholar]
- Martin, M.; Ledent, C.; Parmentier, M.; Maldonado, R.; Valverde, O. Involvement of CB1 cannabinoid receptors in emotional behaviour. Psychopharmacology 2002, 159, 379–387. [Google Scholar] [CrossRef]
- Brancato, A.; Lavanco, G.; Cavallaro, A.; Plescia, F.; Cannizzaro, C. The use of the Emotional-Object Recognition as an assay to assess learning and memory associated to an aversive stimulus in rodents. J. Neurosci. Methods 2016, 274, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Lavanco, G.; Castelli, V.; Brancato, A.; Tringali, G.; Plescia, F.; Cannizzaro, C. The endocannabinoid-alcohol crosstalk: Recent advances on a bi-faceted target. Clin. Exp. Pharmacol. Physiol. 2018, 45, 889–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-mediated control of synaptic transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef] [PubMed]
- Heifets, B.D.; Castillo, P.E. Endocannabinoid signaling and long-term synaptic plasticity. Annu. Rev. Physiol. 2009, 71, 283–306. [Google Scholar] [CrossRef] [Green Version]
- Busquets Garcia, A.; Soria-Gomez, E.; Bellocchio, L.; Marsicano, G. Cannabinoid receptor type-1: Breaking the dogmas. F1000Research 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Oliveira da Cruz, J.F.; Busquets-Garcia, A.; Zhao, Z.; Varilh, M.; Lavanco, G.; Bellocchio, L.; Robin, L.; Cannich, A.; Julio-Kalajzić, F.; Lesté-Lasserre, T.; et al. Specific Hippocampal Interneurons Shape Consolidation of Recognition Memory. Cell Rep. 2020, 32, 108046. [Google Scholar] [CrossRef]
- Wilson, R.I.; Nicoll, R.A. Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature 2001, 410, 588–592. [Google Scholar] [CrossRef]
- Diana, M.A.; Marty, A. Endocannabinoid-mediated short-term synaptic plasticity: Depolarization-induced suppression of inhibition (DSI) and depolarization-induced suppression of excitation (DSE). Br. J. Pharmacol. 2004, 142, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Straiker, A.; Mackie, K. Metabotropic suppression of excitation in murine autaptic hippocampal neurons. J. Physiol. 2007, 578 Pt 3, 773–785. [Google Scholar] [CrossRef]
- Carlson, G.; Wang, Y.; Alger, B.E. Endocannabinoids facilitate the induction of LTP in the hippocampus. Nat. Neurosci. 2002, 5, 723–724. [Google Scholar] [CrossRef]
- Djurisic, M.; Brott, B.K.; Saw, N.L.; Shamloo, M.; Shatz, C.J. Activity-dependent modulation of hippocampal synaptic plasticity via PirB and endocannabinoids. Mol. Psychiatry 2019, 24, 1206–1219. [Google Scholar] [CrossRef] [PubMed]
- Castillo, P.E.; Younts, T.J.; Chávez, A.E.; Hashimotodani, Y. Endocannabinoid signaling and synaptic function. Neuron 2012, 76, 70–81. [Google Scholar] [CrossRef] [Green Version]
- Silva-Cruz, A.; Carlström, M.; Ribeiro, J.A.; Sebastião, A.M. Dual Influence of Endocannabinoids on Long-Term Potentiation of Synaptic Transmission. Front. Pharmacol. 2017, 8, 921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, G.; Walder, K.; Kloiber, S.; Amminger, P.; Berk, M.; Bortolasci, C.C.; Maes, M.; Puri, B.K.; Carvalho, A.F. The endocannabinoidome in neuropsychiatry: Opportunities and potential risks. Pharmacol. Res. 2021, 70, 105729. [Google Scholar] [CrossRef] [PubMed]
- Monyer, H.; Sprengel, R.; Schoepfer, R.; Herb, A.; Higuchi, M.; Lomeli, H.; Burnashev, N.; Sakmann, B.; Seeburg, P.H. Heteromeric NMDA receptors: Molecular and functional distinction of subtypes. Science 1992, 256, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, H.; Singh, S.K.; Mancusso, R.; Gouaux, E. Subunit arrangement and function in NMDA receptors. Nature 2005, 438, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Sengar, A.S.; Li, H.; Zhang, W.; Leung, C.; Ramani, A.K.; Saw, N.M.; Wang, Y.; Tu, Y.; Ross, P.J.; Scherer, S.W.; et al. Control of Long-Term Synaptic Potentiation and Learning by Alternative Splicing of the NMDA Receptor Subunit GluN1. Cell Rep. 2019, 29, 4285–4294.e5. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Huang, F.S.; Abbas, A.K.; Wigstrom, H. Role of NMDA receptor subtypes in different forms of NMDA-dependent synaptic plasticity. BMC Neurosci. 2007, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Baez, M.V.; Cercato, M.C.; Jerusalinsky, D.A. NMDA Receptor Subunits Change after Synaptic Plasticity Induction and Learning and Memory Acquisition. Neural Plast. 2018, 2018, 5093048. [Google Scholar] [CrossRef]
- Dong, Y.N.; Waxman, E.A.; Lynch, D.R. Interactions of postsynaptic density-95 and the NMDA receptor 2 subunit control calpain-mediated cleavage of the NMDA receptor. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 11035–11045. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Levy, J.M.; Hou, A.; Winters, C.A.; Azzam, R.; Sousa, A.A.; Leapman, R.D.; Nicoll, R.A.; Reese, T.S. PSD-95 family MAGUKs are essential for anchoring AMPA and NMDA receptor complexes at the postsynaptic density. Proc. Natl. Acad. Sci. USA 2015, 112, E6983–E6992. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Muñoz, M.; Cortés-Montero, E.; Pozo-Rodrigálvarez, A.; Sánchez-Blázquez, P.; Garzón-Niño, J. The ON:OFF switch, σ1R-HINT1 protein, controls GPCR-NMDA receptor cross-regulation: Implications in neurological disorders. Oncotarget 2015, 6, 35458–35477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Blázquez, P.; Rodríguez-Muñoz, M.; Garzón, J. The cannabinoid receptor 1 associates with NMDA receptors to produce glutamatergic hypofunction: Implications in psychosis and schizophrenia. Front. Pharmacol. 2014, 4, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Muñoz, M.; Sánchez-Blázquez, P.; Merlos, M.; Garzón-Niño, J. Endocannabinoid control of glutamate NMDA receptors: The therapeutic potential and consequences of dysfunction. Oncotarget 2016, 7, 55840–55862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevaleyre, V.; Castillo, P.E. Heterosynaptic LTD of hippocampal GABAergic synapses: A novel role of endocannabinoids in regulating excitability. Neuron 2003, 38, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi-Yamaguchi, Y.; Furuichi, T. The Homer family proteins. Genome Biol. 2007, 8, 206. [Google Scholar] [CrossRef] [Green Version]
- Batista, E.M.; Doria, J.G.; Ferreira-Vieira, T.H.; Alves-Silva, J.; Ferguson, S.S.; Moreira, F.A.; Ribeiro, F.M. Orchestrated activation of mGluR5 and CB1 promotes neuroprotection. Mol. Brain 2016, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Castelli, V.; Brancato, A.; Cavallaro, A.; Lavanco, G.; Cannizzaro, C. Homer2 and Alcohol: A Mutual Interaction. Front. Psychiatry 2017, 8, 268. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, D.M.; Mathias, C.W.; Dawes, M.A.; Furr, R.M.; Charles, N.E.; Liguori, A.; Shannon, E.E.; Acheson, A. Impulsivity, attention, memory, and decision-making among adolescent marijuana users. Psychopharmacology 2013, 226, 307–319. [Google Scholar] [CrossRef]
- Jung, K.M.; Astarita, G.; Zhu, C.; Wallace, M.; Mackie, K.; Piomelli, D. A key role for diacylglycerol lipase-alpha in metabotropic glutamate receptor-dependent endocannabinoid mobilization. Mol. Pharmacol. 2007, 72, 612–621. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.C.; Mackie, K. Review of the Endocannabinoid System. Biol. Psychiatry. Cogn. Neurosci. Neuroimaging 2021, 6, 607–615. [Google Scholar] [CrossRef]
- Scheyer, A.F.; Melis, M.; Trezza, V.; Manzoni, O.J.J. Consequences of perinatal cannabis exposure. Trends Neurosci. 2019, 42, 871–884. [Google Scholar] [CrossRef]
- DiNieri, J.A.; Hurd, Y.L. Rat models of prenatal and adolescent cannabis exposure. Methods Mol. Biol. 2012, 829, 231–242. [Google Scholar]
- Brancato, A.; Castelli, V.; Lavanco, G.; Marino, R.; Cannizzaro, C. In utero Δ9-tetrahydrocannabinol exposure confers vulnerability towards cognitive impairments and alcohol drinking in the adolescent offspring: Is there a role for neuropeptide Y? J. Psychopharmacol. 2020, 34, 663–679. [Google Scholar] [CrossRef]
- Fatemi, S.H.; Folsom, T.D. The neurodevelopmental hypothesis of schizophrenia, revisited. Schizophr. Bull. 2009, 35, 528–548. [Google Scholar] [CrossRef]
- Germann, M.; Brederoo, S.G.; Sommer, I.E.C. Abnormal synaptic pruning during adolescence underlying the development of psychotic disorders. Curr. Opin. Psychiatry 2021, 34, 222–227. [Google Scholar] [CrossRef]
- Battaglia, F.P.; Benchenane, K.; Sirota, A.; Pennartz, C.M.; Wiener, S.I. The hippocampus: Hub of brain network communication for memory. Trends Cogn. Sci. 2011, 15, 310–318. [Google Scholar] [CrossRef]
- Frau, R.; Miczán, V.; Traccis, F.; Aroni, S.; Pongor, C.I.; Saba, P.; Serra, V.; Sagheddu, C.; Fanni, S.; Congiu, M.; et al. Prenatal THC exposure produces a hyperdopaminergic phenotype rescued by pregnenolone. Nat. Neurosci. 2019, 22, 1975–1985. [Google Scholar] [CrossRef] [Green Version]
- Drazanova, E.; Ruda-Kucerova, J.; Kratka, L.; Stark, T.; Kuchar, M.; Maryska, M.; Drago, F.; Starcuk, Z., Jr.; Micale, V. Different effects of prenatal MAM vs. perinatal THC exposure on regional cerebral blood perfusion detected by Arterial Spin Labelling MRI in rats. Sci. Rep. 2019, 9, 6062. [Google Scholar] [CrossRef] [Green Version]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-Like Behavior. Methods Mol. Biol. 2019, 1916, 99–103. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Dalvi, A. Anxiety, defence and the elevated plus-maze. Neurosci. Biobehav. Rev. 1997, 21, 801–810. [Google Scholar] [CrossRef]
- Gawel, K.; Gibula, E.; Marszalek-Grabska, M.; Filarowska, J.; Kotlinska, J.H. Assessment of spatial learning and memory in the Barnes maze task in rodents-methodological consideration. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Serchov, T.; van Calker, D.; Biber, K. Sucrose Preference Test to Measure Anhedonic Behaviour in Mice. Bio-protocol 2016, 6, e1958. [Google Scholar] [CrossRef] [Green Version]
- Popoviç, M.; Biessels, G.J.; Isaacson, R.L.; Gispen, W.H. Learning and memory in streptozotocin-induced diabetic rats in a novel spatial/object discrimination task. Behav. Brain Res. 2001, 122, 201–207. [Google Scholar] [CrossRef]
- Castelli, V.; Lavanco, G.; Brancato, A.; Plescia, F. Targeting the Stress System During Gestation: Is Early Handling a Protective Strategy for the Offspring? Front. Behav. Neurosci. 2020, 14, 9. [Google Scholar] [CrossRef]
- Brancato, A.; Castelli, V.; Cavallaro, A.; Lavanco, G.; Plescia, F.; Cannizzaro, C. Pre-conceptional and Peri-Gestational Maternal Binge Alcohol Drinking Produces Inheritance of Mood Disturbances and Alcohol Vulnerability in the Adolescent Offspring. Front. Psychiatry 2018, 9, 150. [Google Scholar] [CrossRef] [Green Version]
- Brancato, A.; Castelli, V.; Lavanco, G.; Tringali, G.; Micale, V.; Kuchar, M.; D’Amico, C.; Pizzolanti, G.; Feo, S.; Cannizzaro, C. Binge-like Alcohol Exposure in Adolescence: Behavioural, Neuroendocrine and Molecular Evidence of Abnormal Neuroplasticity…and Return. Biomedicines 2021, 9, 1161. [Google Scholar] [CrossRef]
- Plescia, F.; Sardo, P.; Rizzo, V.; Cacace, S.; Marino, R.A.; Brancato, A.; Ferraro, G.; Carletti, F.; Cannizzaro, C. Pregnenolone sulphate enhances spatial orientation and object discrimination in adult male rats: Evidence from a behavioural and electrophysiological study. Behav. Brain Res. 2014, 258, 193–201. [Google Scholar] [CrossRef]
- Sweatt, J.D. Chapter 4—Rodent Behavioral Learning and Memory Models. In Mechanisms of Memory, 2nd ed.; Sweatt, J.D., Ed.; Academic Press: London, UK, 2010; pp. 76–103. [Google Scholar] [CrossRef]
- Gibula-Tarlowska, E.; Wydra, K.; Kotlinska, J.H. Deleterious Effects of Ethanol, Δ(9)-Tetrahydrocannabinol (THC), and Their Combination on the Spatial Memory and Cognitive Flexibility in Adolescent and Adult Male Rats in the Barnes Maze Task. Pharmaceutics 2020, 12, 654. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]
- Volkow, N.D.; Han, B.; Compton, W.M.; McCance-Katz, E.F. Self-reported Medical and Nonmedical Cannabis Use Among Pregnant Women in the United States. JAMA 2019, 322, 167–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Salas-Quiroga, A.; García-Rincón, D.; Gómez-Domínguez, D.; Valero, M.; Simón-Sánchez, S.; Paraíso-Luna, J.; Aguareles, J.; Pujadas, M.; Muguruza, C.; Callado, L.F.; et al. Long-term hippocampal interneuronopathy drives sex-dimorphic spatial memory impairment induced by prenatal THC exposure. Neuropsychopharmacology 2020, 45, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Bara, A.; Manduca, A.; Bernabeu, A.; Borsoi, M.; Serviado, M.; Lassalle, O.; Murphy, M.; Wager-Miller, J.; Mackie, K.; Pelissier-Alicot, A.L.; et al. Sex-dependent effects of in utero cannabinoid exposure on cortical function. eLife 2018, 7, e36234. [Google Scholar] [CrossRef]
- Mereu, G.; Fà, M.; Ferraro, L.; Cagiano, R.; Antonelli, T.; Tattoli, M.; Ghiglieri, V.; Tanganelli, S.; Gessa, G.L.; Cuomo, V. Prenatal exposure to a cannabinoid agonist produces memory deficits linked to dysfunction in hippocampal long-term potentiation and glutamate release. Proc. Natl. Acad. Sci. USA 2003, 100, 4915–4920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, L.F.; Schenk, F. Unpacking the cognitive map: The parallel map theory of hippocampal function. Psychol. Rev. 2003, 110, 285–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrager, Y.; Bayley, P.J.; Bontempi, B.; Hopkins, R.O.; Squire, L.R. Spatial memory and the human hippocampus. Proc. Natl. Acad. Sci. USA 2007, 104, 2961–2966. [Google Scholar] [CrossRef] [Green Version]
- Schiller, D.; Eichenbaum, H.; Buffalo, E.A.; Davachi, L.; Foster, D.J.; Leutgeb, S.; Ranganath, C. Memory and space: Towards an understanding of the cognitive map. J. Neurosci. 2015, 35, 13904–13911. [Google Scholar] [CrossRef] [Green Version]
- Wise, L.E.; Thorpe, A.J.; Lichtman, A.H. Hippocampal CB(1) receptors mediate the memory impairing effects of Delta(9)-tetrahydrocannabinol. Neuropsychopharmacology 2009, 34, 2072–2080. [Google Scholar] [CrossRef]
- Cannich, A.; Wotjak, C.T.; Kamprath, K.; Hermann, H.; Lutz, B.; Marsicano, G. CB1 cannabinoid receptors modulate kinase and phosphatase activity during extinction of conditioned fear in mice. Learn. Mem. 2004, 11, 625–632. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira Alvares, L.; de Oliveira, L.F.; Camboim, C.; Diehl, F.; Genro, B.P.; Lanziotti, V.B.; Quillfeldt, J.A. Amnestic effect of intrahippocampal AM251, a CB1-selective blocker, in the inhibitory avoidance, but not in the open field habituation task, in rats. Neurobiol. Learn. Mem. 2005, 83, 119–124. [Google Scholar] [CrossRef]
- Wilson, W.H.; Ellinwood, E.H.; Mathew, R.J.; Johnson, K. Effects of marijuana on performance of a computerized cognitive-neuromotor test battery. Psychiatry Res. 1994, 51, 115–125. [Google Scholar] [CrossRef]
- D’Souza, D.C.; Perry, E.; MacDougall, L.; Ammerman, Y.; Cooper, T.; Wu, Y.T.; Braley, G.; Gueorguieva, R.; Krystal, J.H. The psychotomimetic effects of intravenous delta-9-tetrahydrocannabinol in healthy individuals: Implications for psychosis. Neuropsychopharmacology 2004, 29, 1558–1572. [Google Scholar] [CrossRef]
- Goldschmidt, L.; Day, N.L.; Richardson, G.A. Effects of prenatal marijuana exposure on child behavior problems at age 10. Neurotoxicol. Teratol. 2000, 22, 325–336. [Google Scholar] [CrossRef]
- Pope, H.G.; Yurgelun-Todd, D. The residual cognitive effects of heavy marijuana use in college students. JAMA 1996, 275, 521–527. [Google Scholar] [CrossRef]
- Lallai, V.; Manca, L.; Sherafat, Y.; Fowler, C.D. Effects of Prenatal Nicotine, THC, or Co-Exposure on Cognitive Behaviors in Adolescent Male and Female Rats. Nicotine Tob. Res. 2022, 24, 1150–1160. [Google Scholar] [CrossRef]
- Meunier, M.; Bachevalier, J.; Mishkin, M.; Murray, E.A. Effects on visual recognition of combined and separate ablations of the entorhinal and perirhinal cortex inrhesus monkeys. J. Neurosci. 1993, 13, 5418–5432. [Google Scholar] [CrossRef]
- Eacott, M.J.; Machin, P.E.; Gaffan, E.A. Elemental and configural visual discrimina-tion learning following lesions to perirhinal cortex in the rat. Behav. Brain Res. 2001, 124, 55–70. [Google Scholar] [CrossRef]
- Higuchi, S.; Miyashita, Y. Formation of mnemonic neuronal responses to visualpaired associates in inferotemporal cortex is impaired by perirhinal andentorhinal lesions. Proc. Natl. Acad. Sci. USA 1996, 93, 739–743. [Google Scholar] [CrossRef] [Green Version]
- Naber, P.A.; Witter, M.P.; Lopez da Silva, F.H. Perirhinal cortex input to the hippocampus in the rat: Evidence for parallel pathways, both direct and indirect. A combined physiological and anatomical study. Eur. J. Neurosci. 1999, 11, 4119–4133. [Google Scholar] [CrossRef]
- Canning, K.J.; Leung, L.S. Lateral entorhinal, perirhinal, and amygdala-entorhinal transition projections to hippocampal CA1 and dentate gyrus in the rat: A current source density study. Hippocampus 1997, 7, 643–655. [Google Scholar] [CrossRef]
- Park, S.B.; Lim, H.Y.; Lee, E.Y.; Yoo, S.W.; Jung, H.S.; Lee, E.; Sun, W.; Lee, I. The fasciola cinereum subregion of the hippocampus is important for the acquisition of visual contextual memory. Prog. Neurobiol. 2022, 210, 102217. [Google Scholar] [CrossRef]
- Silver, H.; Feldman, P. Evidence for sustained attention and working memory in schizophrenia sharing a common mechanism. J. Neuropsychiatry Clin. Neurosci. 2005, 17, 391–398. [Google Scholar] [CrossRef]
- Genaro, G.; Schmidek, W.R. Exploratory activity of rats in three different environments. Ethology 2000, 106, 849–859. [Google Scholar] [CrossRef]
- Azrin, N.H.; Hutchinson, R.R.; Hake, D.F. Attack, avoidance, and escape reactions to aversive shock. J. Exp. Anal. Behav. 1967, 10, 131–148. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.A.; Stempel, A.V.; Vale, R.; Branco, T. Cognitive Control of Escape Behaviour. Trends Cogn. Sci. 2019, 23, 334–348. [Google Scholar] [CrossRef] [Green Version]
- Harrison, F.E.; Hosseini, A.H.; McDonald, M.P. Endogenous anxiety and stress responses in water maze and Barnes maze spatial memory tasks. Behav. Brain Res. 2009, 198, 247–251. [Google Scholar] [CrossRef] [Green Version]
- McHail, D.G.; Valibeigi, N.; Dumas, T.C. A Barnes maze for juvenile rats delineates the emergence of spatial navigation ability. Learn. Mem. 2018, 25, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Antonelli, T.; Tanganelli, S.; Tomasini, M.C.; Finetti, S.; Trabace, L.; Steardo, L.; Sabino, V.; Carratu, M.R.; Cuomo, V.; Ferraro, L. Long-term effects on cortical glutamate release induced by prenatal exposure to the cannabinoid receptor agonist (R)-(+)-[2,3-dihydro-5-methyl-3-(4-morpholinyl-methyl)pyrrolo [1,2,3-de]-1,4-benzoxazin-6-yl]-1-naphthalenylmethanone: An in vivo microdialysis study in the awake rat. Neuroscience 2004, 124, 367–375. [Google Scholar]
- Castaldo, P.; Magi, S.; Gaetani, S.; Cassano, T.; Ferraro, L.; Antonelli, T.; Amoroso, S.; Cuomo, V. Prenatal exposure to the cannabinoid receptor agonist WIN 55,212-2 increases glutamate uptake through overexpression of GLT1 and EAAC1 glutamate transporter subtypes in rat frontal cerebral cortex. Neuropharmacology 2007, 53, 369–378. [Google Scholar] [CrossRef]
- Cull-Candy, S.; Brickley, S.; Farrant, M. NMDA receptor subunits: Diversity, development and disease. Curr. Opin. Neurobiol. 2001, 11, 327–335. [Google Scholar] [CrossRef]
- Flores-Soto, M.E.; Chaparro-Huerta, V.; Escoto-Delgadillo, M.; Vazquez-Valls, E.; González-Castañeda, R.E.; Beas-Zarate, C. Estructura y función de las subunidades del receptor a glutamato tipo NMDA [Structure and function of NMDA-type glutamate receptor subunits]. Neurologia 2012, 27, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Rebola, N.; Srikumar, B.N.; Mulle, C. Activity-dependent synaptic plasticity of NMDA receptors. J. Physiol. 2010, 588 Pt 1, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Otaño, I.; Ehlers, M.D. Learning from NMDA receptor trafficking: Clues to the development and maturation of glutamatergic synapses. Neuro-Signals 2004, 13, 175–189. [Google Scholar] [CrossRef]
- Yashiro, K.; Philpot, B.D. Regulation of NMDA receptor subunit expression and its implications for LTD, LTP, and metaplasticity. Neuropharmacology 2008, 55, 1081–1094. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Clemente, A.; Nicoll, R.A.; Roche, K.W. Diversity in NMDA receptor composition: Many regulators, many consequences. Neuroscientist 2013, 19, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Duan, J. The C-terminus of NMDAR GluN1-1a Subunit Translocates to Nucleus and Regulates Synaptic Function. Front. Cell Neurosci. 2018, 12, 334. [Google Scholar] [CrossRef]
- Guilarte, T.R.; McGlothan, J.L. Hippocampal NMDA receptor mRNA undergoes subunit specific changes during developmental lead exposure. Brain Res. 1998, 790, 98–107. [Google Scholar] [CrossRef]
- Fumagalli, F.; Caffino, L.; Vogt, M.A.; Frasca, A.; Racagni, G.; Sprengel, R.; Gass, P.; Riva, M.A. AMPA GluR-A receptor subunit mediates hippocampal responsiveness in mice exposed to stress. Hippocampus 2011, 21, 1028–1035. [Google Scholar] [CrossRef]
- Laurie, D.J.; Seeburg, P.H. Ligand affinities at recombinant N-methyl-D-aspartate receptors depend on subunit composition. Eur. J. Pharmacol. 1994, 268, 335–345. [Google Scholar] [CrossRef]
- Di Bartolomeo, M.; Stark, T.; Maurel, O.M.; Iannotti, F.A.; Kuchar, M.; Ruda-Kucerova, J.; Piscitelli, F.; Laudani, S.; Pekarik, V.; Salomone, S.; et al. Crosstalk between the transcriptional regulation of dopamine D2 and cannabinoid CB1 receptors in schizophrenia: Analyses in patients and in perinatal Δ9-tetrahydrocannabinol-exposed rats. Pharmacol. Res. 2021, 164, 105357. [Google Scholar] [CrossRef]
- García-Gil, L.; Romero, J.; Ramos, J.A.; Fernández-Ruiz, J.J. Cannabinoid receptor binding and mRNA levels in several brain regions of adult male and female rats perinatally exposed to delta9-tetrahydrocannabinol. Drug Alcohol Depend. 1999, 55, 127–136. [Google Scholar] [CrossRef]
- Keimpema, E.; Mackie, K.; Harkany, T. Molecular model of cannabis sensitivity in developing neuronal circuits. Trends Pharmacol. Sci. 2011, 32, 551–561. [Google Scholar] [CrossRef] [Green Version]
- Berghuis, P.; Rajnicek, A.M.; Morozov, Y.M.; Ross, R.A.; Mulder, J.; Urbán, G.M.; Monory, K.; Marsicano, G.; Matteoli, M.; Canty, A.; et al. Hardwiring the brain: Endocannabinoids shape neuronal connectivity. Science 2007, 316, 1212–1216. [Google Scholar] [CrossRef] [Green Version]
- Vargish, G.A.; Pelkey, K.A.; Yuan, X.; Chittajallu, R.; Collins, D.; Fang, C.; McBain, C.J. Persistent inhibitory circuit defects and disrupted social behaviour following in utero exogenous cannabinoid exposure. Mol. Psychiatry 2017, 22, 56–67. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Sánchez, A.; Sánchez-Blázquez, P.; Rodríguez-Muñoz, M.; Garzón, J. HINT1 protein cooperates with cannabinoid 1 receptor to negatively regulate glutamate NMDA receptor activity. Mol. Brain Res. 2013, 6, 42. [Google Scholar] [CrossRef] [Green Version]
- Koussounadis, A.; Langdon, S.; Um, I.; Harrison, D.J.; Smith, V.A. Relationship between differentially expressed mRNA and mRNA-protein correlations in a xenograft model system. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence | Product |
|---|---|---|
| Gapdh | GTTTGTGATGGGTGTGAACC (Forward) CTTCTGAGTGGCAGTGATG (Reverse) | |
| NMDAR NR1 subunit-Grin1 | Rn_Grin1_1_SG QuantiTect Primer Assay (QT00182287) | |
| NMDAR NR2 subunit-Grin2A | Rn_Grin2a_1_SG QuantiTect Primer Assay (QT00379281) | |
| PSD-95-Dlg4 | Rn_Dlg4_1_SG QuantiTect Primer Assay (QT00183414) | |
| mGluR5-Grm5 | Rn_Grm5_1_SG QuantiTect Primer Assay (QT01081549) | |
| Homer1-HOM1 | CTTCACAGGAATCAGCAGGAG (Forward) GTCCCATTGATACTTTCTGGTG (Reverse) | |
| CB1R-Cnr1 | Rn_Cnr1_1_SG QuantiTect Primer Assay (QT00191737) | |
| Histidine triad nucleotide-binding protein 1 (HINT1) | Rn_Hint1_1_SG QuantiTect Primer Assay (QT01602713) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castelli, V.; Lavanco, G.; Feo, S.; D’Amico, C.; Micale, V.; Kuchar, M.; Plescia, F.; Brancato, A.; Cannizzaro, C. Prenatal Exposure to Δ9-Tetrahydrocannabinol Affects Hippocampus-Related Cognitive Functions in the Adolescent Rat Offspring: Focus on Specific Markers of Neuroplasticity. Pharmaceutics 2023, 15, 692. https://doi.org/10.3390/pharmaceutics15020692
Castelli V, Lavanco G, Feo S, D’Amico C, Micale V, Kuchar M, Plescia F, Brancato A, Cannizzaro C. Prenatal Exposure to Δ9-Tetrahydrocannabinol Affects Hippocampus-Related Cognitive Functions in the Adolescent Rat Offspring: Focus on Specific Markers of Neuroplasticity. Pharmaceutics. 2023; 15(2):692. https://doi.org/10.3390/pharmaceutics15020692
Chicago/Turabian StyleCastelli, Valentina, Gianluca Lavanco, Salvatore Feo, Cesare D’Amico, Vincenzo Micale, Martin Kuchar, Fulvio Plescia, Anna Brancato, and Carla Cannizzaro. 2023. "Prenatal Exposure to Δ9-Tetrahydrocannabinol Affects Hippocampus-Related Cognitive Functions in the Adolescent Rat Offspring: Focus on Specific Markers of Neuroplasticity" Pharmaceutics 15, no. 2: 692. https://doi.org/10.3390/pharmaceutics15020692