
Essential Oils from Côa Valley Lamiaceae Species: Cytotoxicity and Antiproliferative Effect on Glioblastoma Cells

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Essential Oils Isolation
2.3. Gas Chromatography-Mass Spectrometry (GC-MS)
2.4. Scanning Electron Microscopy (SEM)
2.5. Cell Culture
2.6. Cell Viability
2.7. Cell Death Assay
2.8. Cell Cycle Progression Assay
2.9. Statistical Analysis
3. Results and Discussion
3.1. Essential Oils Composition
3.2. Secretory Structures Morphology
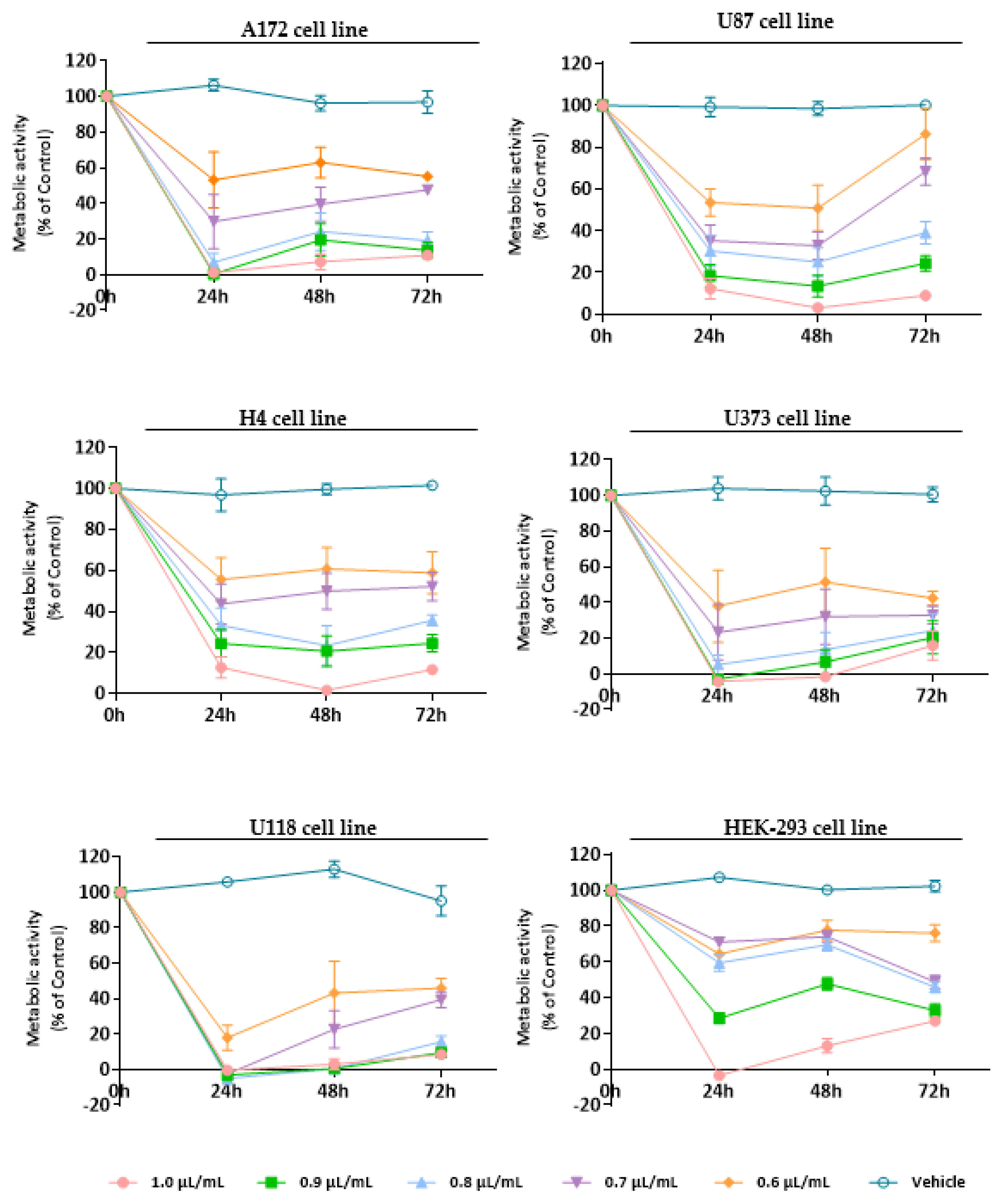
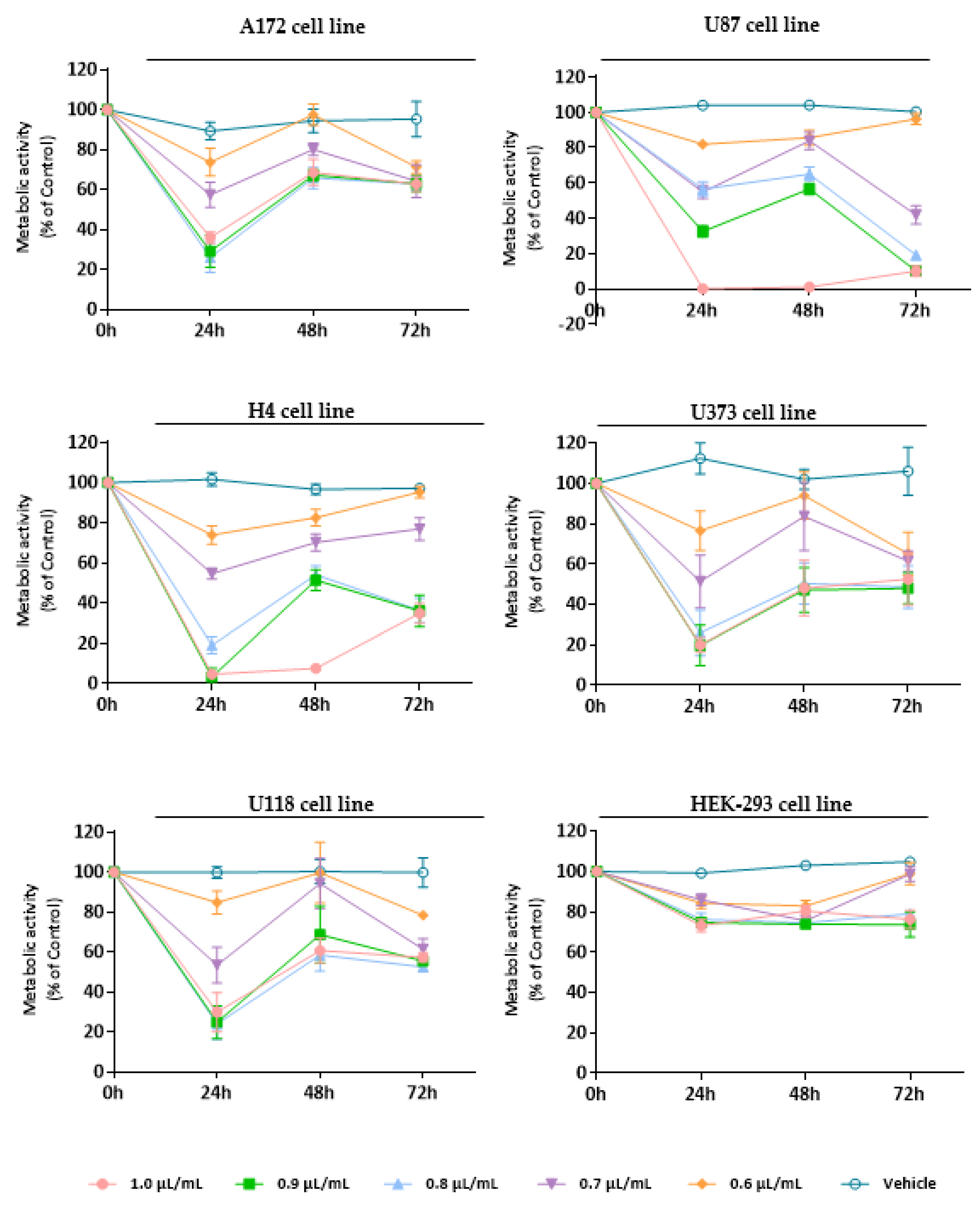
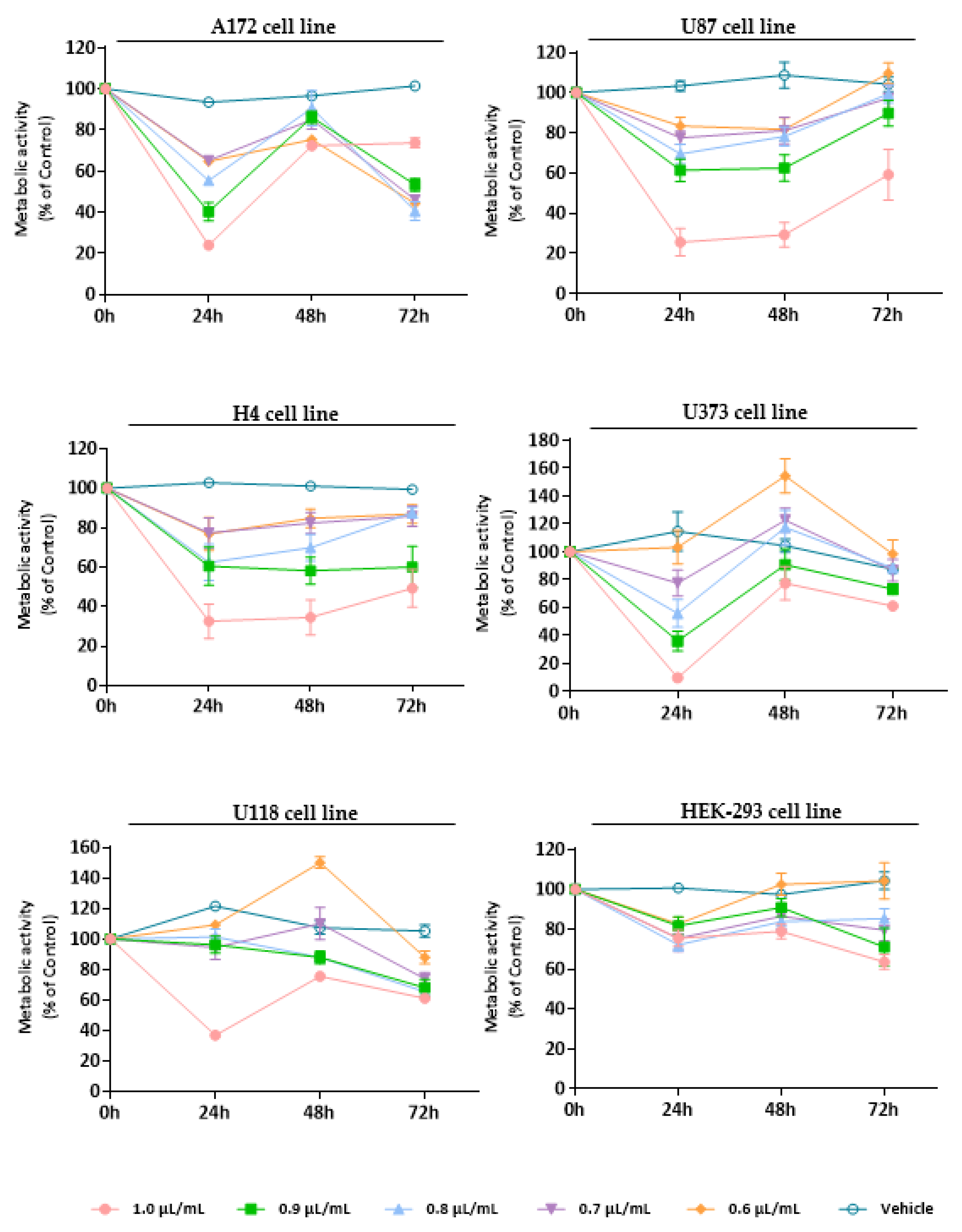
3.3. Evaluation of the Cytotoxic Effect of Essential Oils
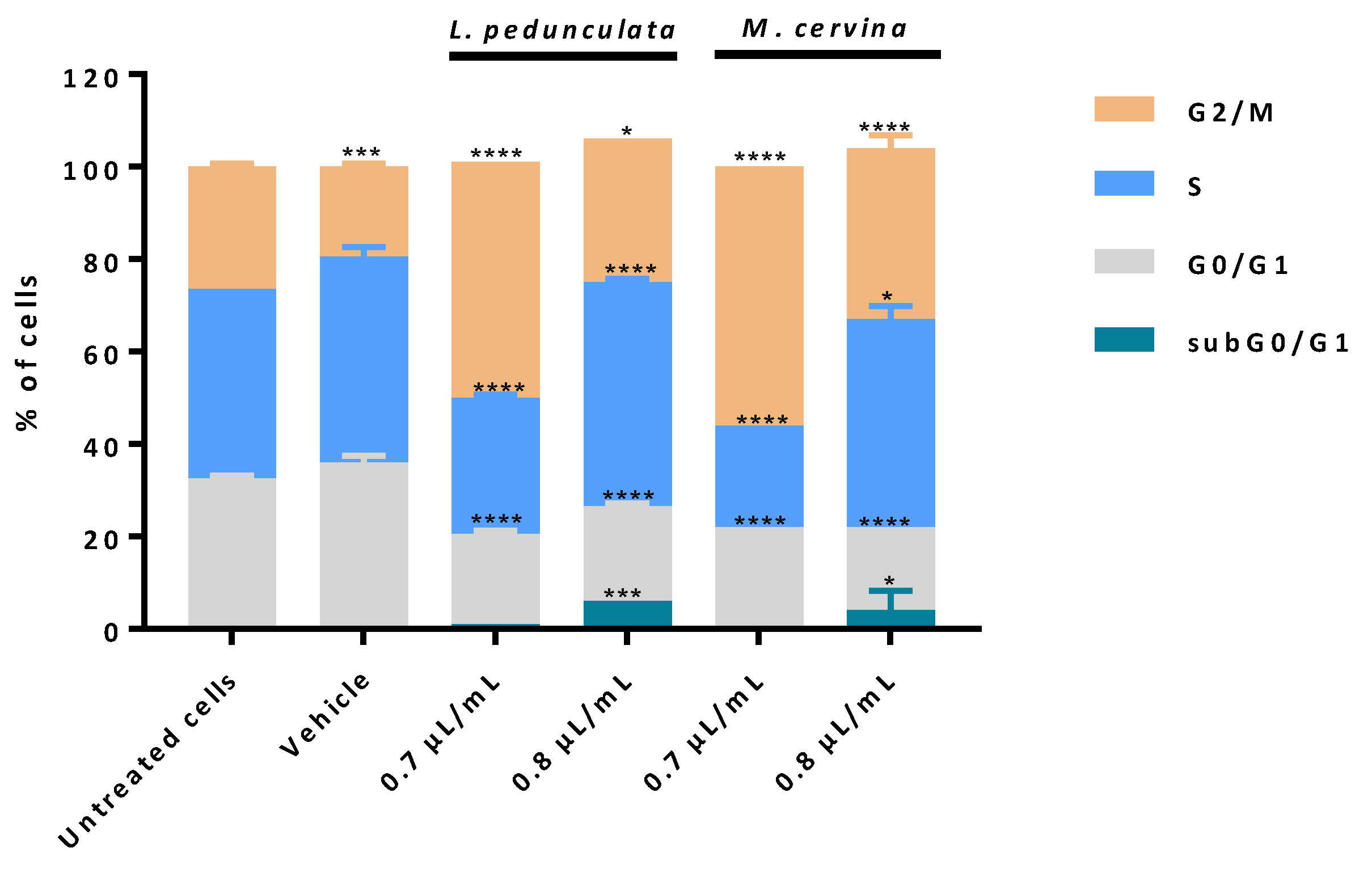
3.4. Assessment of Cell Death Profile and Cell Cycle Progression of Essential Oils
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A Summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Waqar, M.; Trifiletti, D.M.; McBain, C.; O’Connor, J.; Coope, D.J.; Akkari, L.; Quinones-Hinojosa, A.; Borst, G.R. Early Therapeutic Interventions for Newly Diagnosed Glioblastoma: Rationale and Review of the Literature. Curr. Oncol. Rep. 2022, 24, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Martín, E.M.; Magalhães, M.; Díaz-Lanza, A.M.; Marques, M.P.; Princiotto, S.; Gómez, A.M.; Efferth, T.; Cabral, C.; Rijo, P. Phytochemical Study and Antiglioblastoma Activity Assessment of Plectranthus hadiensis (Forssk.) Schweinf. Ex Sprenger Var. Hadiensis Stems. Molecules 2022, 27, 3813. [Google Scholar] [CrossRef] [PubMed]
- Rong, L.; Li, N.; Zhang, Z. Emerging Therapies for Glioblastoma: Current State and Future Directions. J. Exp. Clin. Cancer Res. 2022, 41, 142. [Google Scholar] [CrossRef]
- Fernandes, C.; Costa, A.; Osório, L.; Lago, R.C.; Linhares, P.; Carvalho, B.; Caeiro, C. Current Standards of Care in Glioblastoma Therapy. In Glioblastoma; Exon Publications: Brisbane, Australia, 2017. [Google Scholar]
- le Rhun, E.; Preusser, M.; Roth, P.; Reardon, D.A.; van den Bent, M.; Wen, P.; Reifenberger, G.; Weller, M. Molecular Targeted Therapy of Glioblastoma. Cancer Treat. Rev. 2019, 80, 101896. [Google Scholar] [CrossRef]
- Wen, P.Y.; Packer, R.J. The 2021 WHO Classification of Tumors of the Central Nervous System: Clinical Implications. Neuro Oncol. 2021, 23, 1215–1217. [Google Scholar] [CrossRef]
- Magalhães, M.; Manadas, B.; Efferth, T.; Cabral, C. Chemoprevention and Therapeutic Role of Essential Oils and Phenolic Compounds: Modeling Tumor Microenvironment in Glioblastoma. Pharmacol. Res. 2021, 169, 105638. [Google Scholar] [CrossRef]
- Blowman, K.; Magalhães, M.; Lemos, M.F.L.; Cabral, C.; Pires, I.M. Anticancer Properties of Essential Oils and Other Natural Products. Evid. Based Complement. Altern. Med. 2018, 2018, 3149362. [Google Scholar] [CrossRef]
- de Mesquita, L.S.S.; Luz, T.R.S.A.; Mesquita, J.W.C.d.; Coutinho, D.F.; Amaral, F.M.M.d.; Ribeiro, M.N.d.S.; Malik, S. Exploring the Anticancer Properties of Essential Oils from Family Lamiaceae. Food Rev. Int. 2019, 35, 105–131. [Google Scholar] [CrossRef]
- Beeby, E.; Magalhães, M.; Lemos, M.F.L.; Pires, I.M.; Cabral, C. Cytotoxic Effects of Ridolfia segetum (L.) Moris Phytoproducts in Cancer Cells. J. Ethnopharmacol. 2021, 267, 113515. [Google Scholar] [CrossRef]
- Beeby, E.; Magalhães, M.; Poças, J.; Collins, T.; Lemos, M.F.L.; Barros, L.; Ferreira, I.C.F.R.; Cabral, C.; Pires, I.M. Secondary Metabolites (Essential Oils) from Sand-Dune Plants Induce Cytotoxic Effects in Cancer Cells. J. Ethnopharmacol. 2020, 258, 112803. [Google Scholar] [CrossRef] [PubMed]
- Howes, M.-J.R. The Evolution of Anticancer Drug Discovery from Plants. Lancet Oncol. 2018, 19, 293–294. [Google Scholar] [CrossRef] [PubMed]
- Negre-Zakharov, F.; Long, M.C.; Dudareva, N. Floral Scents and Fruit Aromas Inspired by Nature. In Plant-derived Natural Products: Synthesis, Function, and Application; Springer: New York, NY, USA, 2009. [Google Scholar]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Russo, R.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Exploitation of Cytotoxicity of Some Essential Oils for Translation in Cancer Therapy. Evid. Based Complement. Altern. Med. 2015, 2015, 397821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viktorová, J.; Stupák, M.; Řehořová, K.; Dobiasová, S.; Hoang, L.; Hajšlová, J.; van Thanh, T.; van Tri, L.; van Tuan, N.; Ruml, T. Lemon Grass Essential Oil Does Not Modulate Cancer Cells Multidrug Resistance by Citral—Its Dominant and Strongly Antimicrobial Compound. Foods 2020, 9, 585. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Fan, D.; Xu, Y.; Li, X.; Yuan, J.; Yang, Q.; Zhou, X.; Lu, J.; Zhang, C.; Han, J.; et al. Curcumol Enhances the Sensitivity of Doxorubicin in Triple-Negative Breast Cancer via Regulating the MiR-181b-2-3p-ABCC3 Axis. Biochem. Pharmacol. 2020, 174, 113795. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Li, T.; Chen, L.; Peng, S.; Liao, W.; Bai, R.; Zhao, X.; Yang, H.; Wu, C.; Zeng, H.; et al. Essential Oils from Inula Japonica and Angelicae Dahuricae Enhance Sensitivity of MCF-7/ADR Breast Cancer Cells to Doxorubicin via Multiple Mechanisms. J. Ethnopharmacol. 2016, 180, 18–27. [Google Scholar] [CrossRef]
- Zhou, Y.; Shen, J.; Xia, L.; Wang, Y. Curcuma Zedoaria (Berg.) Rosc. Essential Oil and Paclitaxel Synergistically Enhance the Apoptosis of SKOV3 Cells. Mol. Med. Rep. 2015, 12, 1253–1257. [Google Scholar] [CrossRef] [Green Version]
- Ambrož, M.; Matoušková, P.; Skarka, A.; Zajdlová, M.; Žáková, K.; Skálová, L. The Effects of Selected Sesquiterpenes from Myrica Rubra Essential Oil on the Efficacy of Doxorubicin in Sensitive and Resistant Cancer Cell Lines. Molecules 2017, 22, 1021. [Google Scholar] [CrossRef] [Green Version]
- Zare, E.; Jamali, T.; Ardestani, S.K.; Kavoosi, G. Synergistic Effect of Zataria Multiflora Essential Oil on Doxorubicin-Induced Growth Inhibition of PC3 Cancer Cells and Apoptosis. Complement. Ther. Clin. Pract. 2021, 42, 101286. [Google Scholar] [CrossRef]
- Lampronti, I.; Saab, A.M.; Gambari, R. Antiproliferative Activity of Essential Oils Derived from Plants Belonging to the Magnoliophyta Division. Int. J. Oncol. 2006, 29, 989–995. [Google Scholar] [CrossRef]
- Pudziuvelyte, L.; Stankevicius, M.; Maruska, A.; Petrikaite, V.; Ragazinskiene, O.; Draksiene, G.; Bernatoniene, J. Chemical Composition and Anticancer Activity of Elsholtzia Ciliata Essential Oils and Extracts Prepared by Different Methods. Ind. Crop. Prod. 2017, 107, 90–96. [Google Scholar] [CrossRef]
- Zuzarte, M.R.; Dinis, A.M.; Cavaleiro, C.; Salgueiro, L.R.; Canhoto, J.M. Trichomes, Essential Oils and in Vitro Propagation of Lavandula Pedunculata (Lamiaceae). Ind. Crop. Prod. 2010, 32, 580–587. [Google Scholar] [CrossRef]
- Lopes, C.L.; Pereira, E.; Soković, M.; Carvalho, A.M.; Barata, A.M.; Lopes, V.; Rocha, F.; Calhelha, R.C.; Barros, L.; Ferreira, I.C.F.R. Phenolic Composition and Bioactivity of Lavandula pedunculata (Mill.) Cav. Samples from Different Geographical Origin. Molecules 2018, 23, 1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neves, J.M.; Matos, C.; Moutinho, C.; Queiroz, G.; Gomes, L.R. Ethnopharmacological Notes about Ancient Uses of Medicinal Plants in Trás-Os-Montes (Northern of Portugal). J. Ethnopharmacol. 2009, 124, 270–283. [Google Scholar] [CrossRef]
- Miguel, M.; Barros, L.; Pereira, C.; Calhelha, R.C.; Garcia, P.A.; Castro, M.Á.; Santos-Buelga, C.; Ferreira, I.C.F.R. Chemical Characterization and Bioactive Properties of Two Aromatic Plants: Calendula officinalis L. (Flowers) and Mentha cervina L. (Leaves). Food Funct. 2016, 7, 2223–2232. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, L.; Monteiro, P.; Póvoa, O.; Teixeira, G.; Moldão, M.; Figueiredo, A.C.; Monteiro, A. Morphology of Secretory Structures and Essential Oil Composition in Mentha cervina L. from Portugal. Flavour Fragr. J. 2008, 23, 340–347. [Google Scholar] [CrossRef]
- Rodrigues, L.; Duarte, A.; Figueiredo, A.C.; Brito, L.; Teixeira, G.; Moldão, M.; Monteiro, A. Chemical Composition and Antibacterial Activity of the Essential Oils from the Medicinal Plant Mentha cervina L. Grown in Portugal. Med. Chem. Res. 2011, 21, 3485–3490. [Google Scholar] [CrossRef]
- Barros, L.; Heleno, S.A.; Carvalho, A.M.; Ferreira, I.C.F.R. Lamiaceae Often Used in Portuguese Folk Medicine as a Source of Powerful Antioxidants: Vitamins and Phenolics. LWT 2010, 43, 544–550. [Google Scholar] [CrossRef]
- Méndez-Tovar, I.; Sponza, S.; Asensio-S-Manzanera, M.C.; Novak, J. Contribution of the Main Polyphenols of Thymus mastichina subsp. mastichina to Its Antioxidant Properties. Ind. Crop. Prod. 2015, 66, 291–298. [Google Scholar] [CrossRef]
- Rodrigues, M.; Lopes, A.C.; Vaz, F.; Filipe, M.; Alves, G.; Ribeiro, M.P.; Coutinho, P.; Araujo, A.R.T.S. Thymus mastichina: Composition and Biological Properties with a Focus on Antimicrobial Activity. Pharmaceuticals 2020, 13, 479. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Tovar, I.; Martín, H.; Santiago, Y.; Ibeas, A.; Herrero, B.; Asensio-S.-Manzanera, M.C. Variation in Morphological Traits among Thymus mastichina (L.) L. Populations. Genet. Resour. Crop. Evol. 2015, 62, 1257–1267. [Google Scholar] [CrossRef]
- Arantes, S.M.; Piçarra, A.; Guerreiro, M.; Salvador, C.; Candeias, F.; Caldeira, A.T.; Martins, M.R. Toxicological and Pharmacological Properties of Essential Oils of Calamintha nepeta, Origanum virens and Thymus mastichina of Alentejo (Portugal). Food Chem. Toxicol. 2019, 133, 110747. [Google Scholar] [CrossRef] [PubMed]
- Taghouti, M.; Martins-Gomes, C.; Schäfer, J.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Chemical Characterization and Bioactivity of Extracts from Thymus mastichina: A Thymus with a Distinct Salvianolic Acid Composition. Antioxidants 2020, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Albano, S.M.; Miguel, M.G. Biological Activities of Extracts of Plants Grown in Portugal. Ind. Crop. Prod. 2011, 33, 338–343. [Google Scholar] [CrossRef]
- Gordo, J.; Máximo, P.; Cabrita, E.; Lourenço, A.; Oliva, A.; Almeida, J.; Filipe, M.; Cruz, P.; Barcia, R.; Santos, M.; et al. Thymus mastichina: Chemical Constituents and Their Anti-Cancer Activity. Nat. Prod. Commun. 2012, 7, 1491–1494. [Google Scholar] [CrossRef] [Green Version]
- Leal, F.; Coelho, A.C.; Soriano, T.; Alves, C.; Matos, M. Fungicide Activity of Thymus mastichina and Mentha rotundifolia in Plants in Vitro. J. Med. Food 2013, 16, 273. [Google Scholar] [CrossRef]
- Council of Europe. European Pharmacopoeia (Ph. Eur.), 9th ed.; Council of Europe: Strasbourg, France, 2016. [Google Scholar]
- Spréa, R.M.; Fernandes, Â.; Calhelha, R.C.; Pereira, C.; Pires, T.C.S.P.; Alves, M.J.; Canan, C.; Barros, L.; Amaral, J.S.; Ferreira, I.C.F.R. Chemical and Bioactive Characterization of the Aromatic Plant: Levisticum Officinale W.D.J. Koch: A Comprehensive Study. Food Funct. 2020, 11, 1292–1303. [Google Scholar] [CrossRef]
- Marques, M.P.; Martin, D.; Bosch, M.; Martins, J.; Biswal, A.; Zuzarte, M.; de Carvalho, L.B.; Canhoto, J.; da Costa, R. Unveiling the Compositional Remodelling of Arbutus unedo L. Fruits during Ripening. Sci. Hortic. 2022, 303, 111248. [Google Scholar] [CrossRef]
- Magalhães, M.; Farinha, D.; de Lima, M.C.P.; Faneca, H. Increased Gene Delivery Efficiency and Specificity of a Lipid-Based Nanosystem Incorporating a Glycolipid. Int. J. Nanomed. 2014, 9, 4979–4989. [Google Scholar] [CrossRef]
- Magalhães, M.; Jorge, J.; Gonçalves, A.C.; Sarmento-Ribeiro, A.B.; Carvalho, R.; Figueiras, A.; Santos, A.C.; Veiga, F. MiR-29b and Retinoic Acid Co-Delivery: A Promising Tool to Induce a Synergistic Antitumoral Effect in Non-Small Cell Lung Cancer Cells. Drug Deliv. Transl. Res. 2020, 10, 1367–1380. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, M.; Almeida, M.; Tavares-da-Silva, E.; Roleira, F.M.F.; Varela, C.; Jorge, J.; Gonçalves, A.C.; Carvalho, R.A.; Veiga, F.; Santos, A.C.; et al. MiR-145-Loaded Micelleplexes as a Novel Therapeutic Strategy to Inhibit Proliferation and Migration of Osteosarcoma Cells. Eur. J. Pharm. Sci. 2018, 123, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.; Gonçalves, S.; Valentão, P.; Andrade, P.B.; Almeida, C.; Nogueira, J.M.F.; Romano, A. Metabolic Profile and Biological Activities of Lavandula pedunculata Subsp. lusitanica (Chaytor) Franco: Studies on the Essential Oil and Polar Extracts. Food Chem. 2013, 141, 2501–2506. [Google Scholar] [CrossRef] [PubMed]
- Zuzarte, M.; Gonåalves, M.J.; Cavaleiro, C.; Dinis, A.M.; Canhoto, J.M.; Salgueiro, L.R. Chemical Composition and Antifungal Activity of the Essential Oils of Lavandula pedunculata (Miller) Cav. Chem. Biodivers. 2009, 6, 1283–1292. [Google Scholar] [CrossRef]
- Politi, M.; Rodrigues, C.L.; Gião, M.S.; Pintado, M.E.; Castro, P.M.L. Antioxidant Principles and Volatile Constituents from the North-Western Iberian Mint “Erva-Peixeira”, Mentha cervina. Nat. Prod. Commun. 2008, 3, 2065–2068. [Google Scholar] [CrossRef] [Green Version]
- Helal, I.M.; El-Bessoumy, A.; Al-Bataineh, E.; Joseph, M.R.P.; Rajagopalan, P.; Chandramoorthy, H.C.; ben Hadj Ahmed, S. Antimicrobial Efficiency of Essential Oils from Traditional Medicinal Plants of Asir Region, Saudi Arabia, over Drug Resistant Isolates. Biomed. Res. Int. 2019, 2019, 8928306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, M.J.; Vicente, A.M.; Cavaleiro, C.; Salgueiro, L. Composition and Antifungal Acticvity of the Essential Oil of Mentha cervia from Portugal. Nat. Prod. Res. 2007, 21, 867–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Abu-Darwish, M.S.; Tarawneh, A.H.; Cabral, C.; Gadetskaya, A.V.; Salgueiro, L.; Hosseinabadi, T.; Rajabi, S.; Chanda, W.; Sharifi-Rad, M.; et al. Thymus spp. Plants-Food Applications and Phytopharmacy Properties. Trends Food Sci. Technol. 2019, 85, 287–306. [Google Scholar] [CrossRef]
- Salgueiro, L.R.; Vila, R.; Tomàs, X.; Cañigueral, S.; da Cunha, A.P.; Adzet, T. Composition and Variability of the Essential Oils of Thymus Species from Section Mastichina from Portugal. Syst. Ecol. 1997, 25, 659–672. [Google Scholar] [CrossRef]
- Faleiro, M.L.; Miguel, M.G.; Ladeiro, F.; Venâncio, F.; Tavares, R.; Brito, J.C.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G. Antimicrobial Activity of Essential Oils Isolated from Portuguese Endemic Species of Thymus. Lett. Appl. Microbiol. 2003, 36, 35–40. [Google Scholar] [CrossRef]
- Miguel, G.; Simões, M.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Carvalho, L. Composition and Antioxidant Activities of the Essential Oils of Thymus caespititius, Thymus camphoratus and Thymus mastichina. Food Chem. 2004, 86, 183–188. [Google Scholar] [CrossRef]
- Araujo, A.R.T.S.; Périno, S.; Fernandez, X.; Cunha, C.; Rodrigues, M.; Ribeiro, M.P.; Jordao, L.; Silva, L.A.; Rodilla, J.; Coutinho, P.; et al. Solvent-free Microwave Extraction of Thymus mastichina Essential Oil: Influence on Their Chemical Composition and on the Antioxidant and Antimicrobial Activities. Pharmaceuticals 2021, 14, 709. [Google Scholar] [CrossRef] [PubMed]
- Cutillas, A.B.; Carrasco, A.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Thymus mastichina L. Essential Oils from Murcia (Spain): Composition and Antioxidant, Antienzymatic and Antimicrobial Bioactivities. PLoS ONE 2018, 13, e0190790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy, 5th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2017. [Google Scholar]
- John Wiley & Sons NIST 05 Mass Spectral Library, 8th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006.
- Bosabalidis, A.M.; Tsekos, I. Glandular Hair Formation in Origanum Species. Ann. Bot. 1984, 53, 559–563. [Google Scholar] [CrossRef]
- Luca, T.; Napoli, E.; Privitera, G.; Musso, N.; Ruberto, G.; Castorina, S. Antiproliferative Effect and Cell Cycle Alterations Induced by Salvia Officinalis Essential Oil and Its Three Main Components in Human Colon Cancer Cell Lines. Chem. Biodivers. 2020, 17, e2000309. [Google Scholar] [CrossRef]
- de Lima, V.T.; Vieira, M.C.; Kassuya, C.A.L.; Cardoso, C.A.L.; Alves, J.M.; Foglio, M.A.; de Carvalho, J.E.; Formagio, A.S.N. Chemical Composition and Free Radical-Scavenging, Anticancer and Anti-Inflammatory Activities of the Essential Oil from Ocimum Kilimandscharicum. Phytomedicine 2014, 21, 1298–1302. [Google Scholar] [CrossRef]
- Elansary, H.O.; Abdelgaleil, S.A.M.; Mahmoud, E.A.; Yessoufou, K.; Elhindi, K.; El-Hendawy, S. Effective Antioxidant, Antimicrobial and Anticancer Activities of Essential Oils of Horticultural Aromatic Crops in Northern Egypt. BMC Complement. Altern. Med. 2018, 18, 214. [Google Scholar] [CrossRef]
- Rahimifard, N.; Hajimehdipoor, H.; Hedayati, M.H.; Bagheri, O.; Pishehvar, H.; Ajani, Y. Cytotoxic Effects of Essential Oils and Extracts of Some Mentha Species on Vero, Hela and Hep2 Cell Lines. J. Med. Plants 2010, 9, 88–92. [Google Scholar]
- Shehab, N.G.; Abu-Gharbieh, E. Constituents and Biological Activity of the Essential Oil and the Aqueous Extract of Micromeria fruticosa (L.) Druce Subsp. Serpyllifolia. Pak. J. Pharm. Sci. 2012, 25, 687–692. [Google Scholar]
- Loizzo, M.R.; Tundis, R.; Menichini, F.; Saab, A.M.; Statti, G.A.; Menichini, F. Cytotoxic Activity of Essential Oils from Labiatae and Lauraceae Families against in Vitro Human Tumor Models. Anticancer. Res. 2007, 27, 3293–3299. [Google Scholar]
- Gelles, J.D.; Edward Chipuk, J. Robust High-Throughput Kinetic Analysis of Apoptosis with Real-Time High-Content Live-Cell Imaging. Cell Death Dis. 2016, 7, e2493. [Google Scholar] [CrossRef] [PubMed]
- Rieger, A.M.; Nelson, K.L.; Konowalchuk, J.D.; Barreda, D.R. Modified Annexin V/Propidium Iodide Apoptosis Assay for Accurate Assessment of Cell Death. J. Vis. Exp. 2011, e2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taruvai Kalyana Kumar, R.; Liu, S.; Minna, J.D.; Prasad, S. Monitoring Drug Induced Apoptosis and Treatment Sensitivity in Non-Small Cell Lung Carcinoma Using Dielectrophoresis. Biochim. Biophys. Acta (BBA) Gen. Subj. 2016, 1860, 1877–1883. [Google Scholar] [CrossRef] [PubMed]
- Duymuş, H.G.; Çiftçi, G.A.; Yildirim, Ş.U.; Demirci, B.; Kirimer, N. The Cytotoxic Activity of Vitex agnus castus L. Essential Oils and Their Biochemical Mechanisms. Ind. Crop. Prod. 2014, 55, 33–42. [Google Scholar] [CrossRef]
- Azimi, M.; Mehrzad, J.; Ahmadi, A.; Ahmadi, E.; Ghorbani Ranjbary, A. Apoptosis Induced by Ziziphora Tenuior Essential Oil in Human Colorectal Cancer Cells. Biomed. Res. Int. 2021, 2021, 5522964. [Google Scholar] [CrossRef]
- Magalhães, M.; Domínguez-Martín, E.M.; Jorge, J.; Gonçalves, A.C.; Díaz-Lanza, A.M.; Manadas, B.; Efferth, T.; Rijo, P.; Cabral, C. Parvifloron D-Based Potential Therapy for Glioblastoma: Inducing Apoptosis via the Mitochondria Dependent Pathway. Front. Pharmacol. 2022, 13, 1006832. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
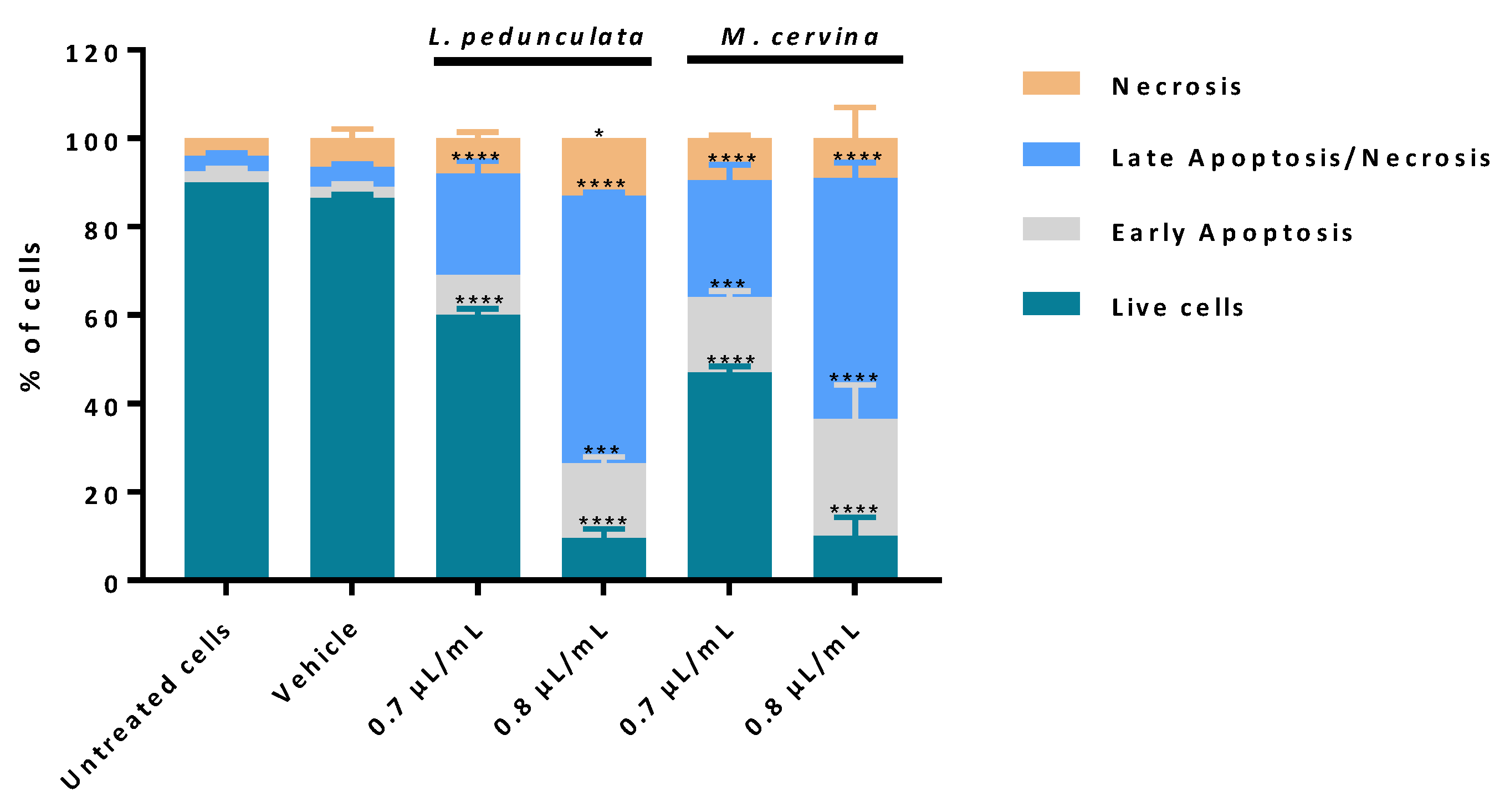{kind=link}
{kind=link}
| Compound | RT | LRI a | LRI b | LP c | MC d | TM e |
|---|---|---|---|---|---|---|
| Tricyclene | 13.29 | 914 | 921 | 0.39 | - | 0.06 |
| α-Thujene | 13.61 | 920 | 924 | - | - | 0.18 |
| α-Pinene | 13.91 | 925 | 932 | 6.9 | 0.66 | 4.1 |
| Camphene | 14.65 | 940 | 946 | 4.00 | - | 1.4 |
| 2,4(10)-Thujadiene | 14.97 | 946 | 953 | 0.357 | - | - |
| Sabinene | 15.99 | 965 | 969 | 0.022 | 0.45 | 3.2 |
| β-Pinene | 16.12 | 968 | 974 | 0.107 | 1.139 | 5.2 |
| 2,3-Dehydro-1,8-cineole | 16.88 | 983 | 988 | - | - | 0.34 |
| β-Myrcene | 16.98 | 984 | 988 | - | 0.2 | 1.1 |
| Δ3-Carene | 17.89 | 1002 | 1008 | 1.21 | - | - |
| 4-Carene | 18.24 | 1009 | 1018 * | 0.048 | 0.85 | 0.27 |
| p-Cymene | 18.52 | 1014 | 1020 | 0.28 | - | - |
| o-Cymene | 18.66 | 1017 | 1022 | 0.53 | 0.36 | 0.61 |
| Limonene | 18.8 | 1019 | 1025 | 0.53 | 1.46 | - |
| 1,8-Cineole | 19.03 | 1024 | 1026 | 10.9 | - | 57.00 |
| cis-β-Ocimene | 19.41 | 1031 | 1032 | 0.18 | 0.161 | 0.18 |
| trans-β-Ocimene | 19.93 | 1041 | 1044 | - | - | 1.7 |
| γ-Terpinene | 20.44 | 1051 | 1053 | 0.092 | 1.6 | 0.61 |
| cis-Sabinene hydrate | 20.86 | 1059 | 1065 | - | 3.4 | 0.54 |
| cis-Linalool oxide | 21.14 | 1065 | 1067 | 0.68 | - | 0.68 |
| Camphenilone | 21.65 | 1074 | 1078 | 0.45 | - | 0.45 |
| Fenchone | 21.9 | 1079 | 1083 | 5.36 | - | 5.36 |
| p-Mentha-2,4(8)-diene | 21.97 | 1081 | 1085 | - | 0.4 | 0.41 |
| trans-Sabinene hydrate | 22.45 | 1090 | 1096 * | - | 0.28 | 0.28 |
| Linalool | 22.56 | 1091 | 1095 | 1.9 | 0.18 | 1.29 |
| Fenchol | 23.25 | 1105 | 1114 | 0.072 | - | 0.072 |
| cis-p-Menth-2-en-1-ol | 23.62 | 1113 | 1120 * | - | 0.23 | 0.144 |
| α-Campholenal-(+)2-pinen-7-one | 23.85 | 1118 | 1122 + 1124 | 0.62 | - | 0.62 |
| trans-Pinocarveol | 24.50 | 1131 | 1135 | 0.19 | - | 0.17 |
| trans-p-Menth-2-en-1-ol | 24.55 | 1132 | 1139 * | - | 0.24 | - |
| Camphor | 24.73 | 1135 | 1141 | 39.00 | - | 1.2 |
| Menthone | 25.24 | 1146 | 1148 | - | 0.12 | - |
| Pinocarvone | 25.66 | 1154 | 1160 | 0.12 | - | 0.102 |
| Menthofuran | 25.74 | 1156 | 1159 | - | 0.19 | - |
| endo-Borneol | 25.88 | 1159 | 1165 | 2.4 | - | 4.8 |
| Isopulegone | 26.33 | 1168 | - | - | 2.55 | - |
| Terpinen-4-ol | 26.41 | 1169 | 1174 | 0.66 | 5.7 | 1.8 |
| m-Cymen-8-ol | 26.64 | 1174 | 1176 | 0.6 | - | - |
| p-Cymen-8-ol | 26.79 | 1177 | 1179 | 1.1 | - | - |
| α-Terpineol | 27.07 | 1182 | 1186 | 0.69 | 0.5 | 7.2 |
| Myrtenal | 27.32 | 1188 | 1195 | 0.37 | - | 0.43 |
| Verbenone | 27.93 | 1200 | 1204 | 1.2 | - | - |
| Pulegone | 29.45 | 1230 | 1233 | - | 60.00 | - |
| Linalool acetate | 30.17 | 1248 | 1254 | 0.25 | - | - |
| Bornyl acetate | 31.59 | 1278 | 1284 | 5.9 | - | 0.07 |
| Lavandulyl acetate | 31.8 | 1282 | 1288 | 0.28 | - | 0.28 |
| α-Terpinyl acetate | 34.45 | 1341 | 1346 | - | - | 0.19 |
| β-Elemene | 36.35 | 1385 | 1389 | - | 0.18 | - |
| β-Caryophyllene | 37.56 | 1412 | 1417 | - | 1.2 | 0.5 |
| Alloaromadendrene | 39.31 | 1455 | 1458 | - | 0.218 | 0.11 |
| Cadina-3,5-diene | 39.38 | 1456 | - | 0.07 | - | - |
| Germacrene D | 40.13 | 1474 | 1477 | 0.22 | 12.00 | - |
| β-Selinene | 40.36 | 1480 | 1489 | 0.05 | - | - |
| 1,11-Oxidocalamenene | 40.51 | 1483 | 1490 * | 0.42 | - | - |
| Bicyclogermacrene | 40.77 | 1490 | 1500 | - | 1.3 | 0.26 |
| γ-Cadinene | 41.46 | 1507 | 1513 | 0.12 | - | - |
| δ-Cadinene | 41.81 | 1516 | 1522 | 0.38 | 0.6 | - |
| Elemol | 42.84 | 1541 | 1548 | - | - | 0.9 |
| Globulol | 44.57 | 1585 | 1590 | - | 2.00 | 1.5 |
| Ledol | 45.02 | 1597 | 1602 | - | - | 0.22 |
| 1,10-Di-epi-cubenol | 45.43 | 1609 | 1618 | 0.37 | - | - |
| γ-Eudesmol | 46.07 | 1625 | 1630 | - | 0.3 | 0.24 |
| Monoterpene hydrocarbons | 14.7 | 10.9 | 19,00 | |||
| Oxygen-containing monoterpenes | 73.00 | 70.00 | 74.7 | |||
| Sesquiterpene hydrocarbons | 0.8 | 15.00 | 0.9 | |||
| Oxygen-containing sesquiterpenes | 7.3 | 2.7 | 2.8 | |||
| Total identified | 95.80 | 98.60 | 97.40 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, M.P.; Neves, B.G.; Varela, C.; Zuzarte, M.; Gonçalves, A.C.; Dias, M.I.; Amaral, J.S.; Barros, L.; Magalhães, M.; Cabral, C. Essential Oils from Côa Valley Lamiaceae Species: Cytotoxicity and Antiproliferative Effect on Glioblastoma Cells. Pharmaceutics 2023, 15, 341. https://doi.org/10.3390/pharmaceutics15020341
Marques MP, Neves BG, Varela C, Zuzarte M, Gonçalves AC, Dias MI, Amaral JS, Barros L, Magalhães M, Cabral C. Essential Oils from Côa Valley Lamiaceae Species: Cytotoxicity and Antiproliferative Effect on Glioblastoma Cells. Pharmaceutics. 2023; 15(2):341. https://doi.org/10.3390/pharmaceutics15020341
Chicago/Turabian StyleMarques, Mário Pedro, Beatriz Guapo Neves, Carla Varela, Mónica Zuzarte, Ana Cristina Gonçalves, Maria Inês Dias, Joana S. Amaral, Lillian Barros, Mariana Magalhães, and Célia Cabral. 2023. "Essential Oils from Côa Valley Lamiaceae Species: Cytotoxicity and Antiproliferative Effect on Glioblastoma Cells" Pharmaceutics 15, no. 2: 341. https://doi.org/10.3390/pharmaceutics15020341