

Polymeric Particle BAM15 Targeting Macrophages Attenuates the Severity of LPS-Induced Sepsis: A Proof of Concept for Specific Immune Cell-Targeted Therapy
 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Animal Model
2.2. Preparation of Poly Lactic-Co-Glycolic Acid Particles (PLGA)
2.3. In Vivo Uptake of Fluorescein Isothiocyanate (FITC)-Tagged Particles (PLGA-FITC)
2.4. Human Cell Line Experiments
2.5. Mouse Cell Line Experiments
2.6. Quantitative Real-Time PCR
2.7. Analysis of Cell Energy Metabolism
2.8. Statistical Analysis
3. Results
3.1. BAM15 Interfered with the Polarization of M1 but Not M2 Macrophages in a Human Macrophage Cell Line
3.2. BAM15 Altered Mitochondrial Energy Metabolism in M1 Macrophages
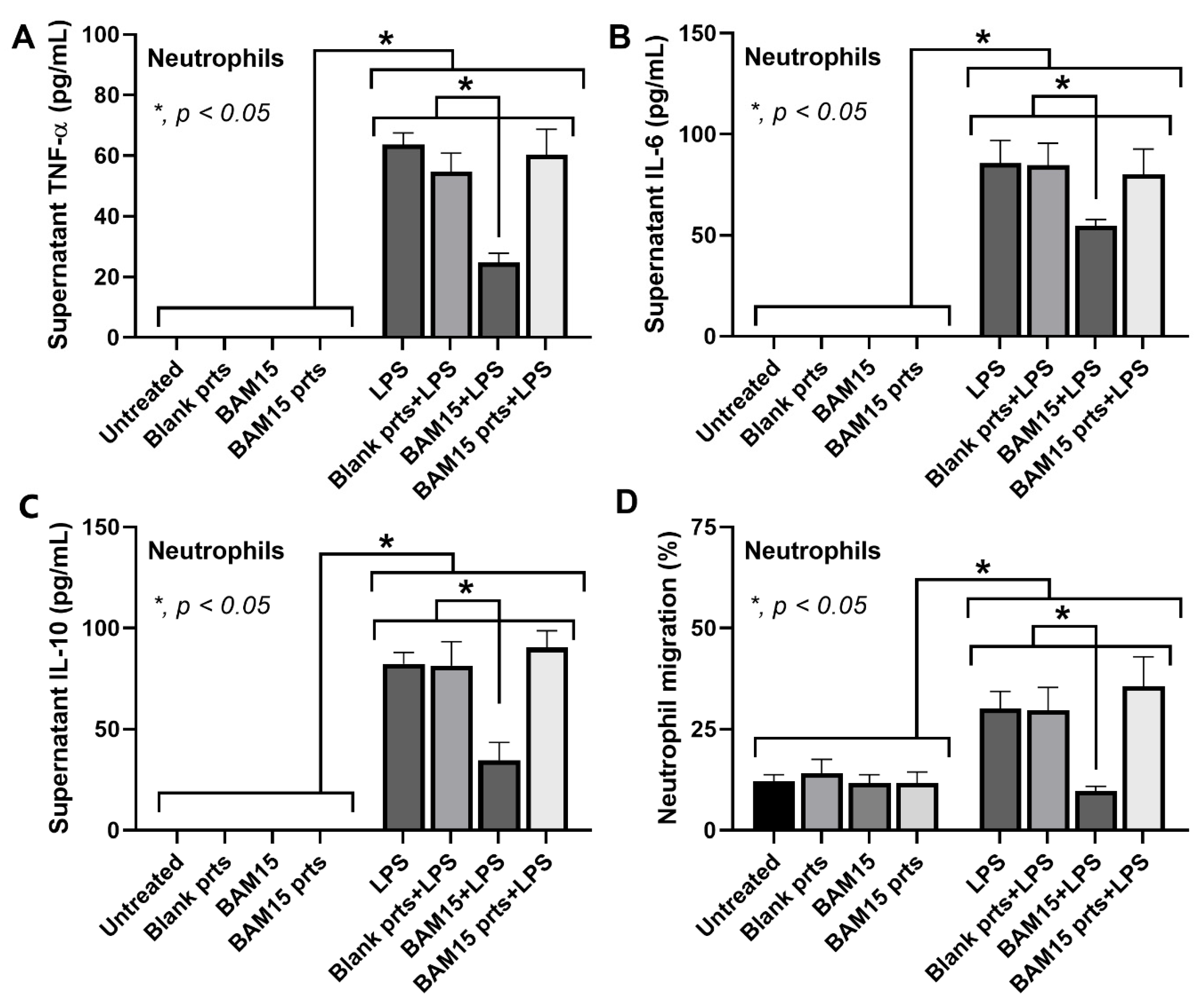
3.3. BAM15 and BAM15-PLGA Particles (BAM15 Particles) Attenuated Inflammatory Responses in LPS-Activated Macrophages but Not Neutrophils
3.4. In Vivo Uptake of PLGA-FITC Particles by Macrophages
3.5. BAM15 Particles Attenuated Inflammation in LPS-Induced Sepsis Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, C.; Ma, C.; Gong, L.; Guo, Y.; Fu, K.; Zhang, Y.; Zhou, H.; Li, Y. Macrophage Polarization and Its Role in Liver Disease. Front. Immunol. 2021, 12, 803037. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.; Gao, Y.; Shou, S.; Chai, Y. The roles of macrophage polarization in the host immune response to sepsis. Int. Immunopharmacol. 2021, 96, 107791. [Google Scholar] [CrossRef]
- Kolliniati, O.; Ieronymaki, E.; Vergadi, E.; Tsatsanis, C. Metabolic Regulation of Macrophage Activation. J. Innate Immun. 2022, 14, 51–68. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z. The role of macrophages polarization in sepsis-induced acute lung injury. Front. Immunol. 2023, 14, 1209438. [Google Scholar] [CrossRef]
- Gao, J.; Liang, Y.; Wang, L. Shaping Polarization of Tumor-Associated Macrophages in Cancer Immunotherapy. Front. Immunol. 2022, 13, 888713. [Google Scholar] [CrossRef]
- Mantovani, A.; Allavena, P.; Marchesi, F.; Garlanda, C. Macrophages as tools and targets in cancer therapy. Nat. Rev. Drug Discov. 2022, 21, 799–820. [Google Scholar] [CrossRef]
- Van den Bossche, J.; O’Neill, L.A.; Menon, D. Macrophage Immunometabolism: Where Are We (Going)? Trends Immunol. 2017, 38, 395–406. [Google Scholar] [CrossRef]
- Wang, Y.; Li, N.; Zhang, X.; Horng, T. Mitochondrial metabolism regulates macrophage biology. J. Biol. Chem. 2021, 297, 100904. [Google Scholar] [CrossRef]
- Xu, L.; Yan, X.; Zhao, Y.; Wang, J.; Liu, B.; Yu, S.; Fu, J.; Liu, Y.; Su, J. Macrophage Polarization Mediated by Mitochondrial Dysfunction Induces Adipose Tissue Inflammation in Obesity. Int. J. Mol. Sci. 2022, 23, 9252. [Google Scholar] [CrossRef]
- Luo, R.; Li, X.; Wang, D. Reprogramming Macrophage Metabolism and its Effect on NLRP3 Inflammasome Activation in Sepsis. Front. Mol. Biosci. 2022, 9, 917818. [Google Scholar] [CrossRef] [PubMed]
- Ramond, E.; Jamet, A.; Coureuil, M.; Charbit, A. Pivotal Role of Mitochondria in Macrophage Response to Bacterial Pathogens. Front. Immunol. 2019, 10, 2461. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulos, S.J.; Chen, S.Y.; Brandon, A.E.; Salamoun, J.M.; Byrne, F.L.; Garcia, C.J.; Beretta, M.; Olzomer, E.M.; Shah, D.P.; Philp, A.M.; et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nat. Commun. 2020, 11, 2397. [Google Scholar] [CrossRef] [PubMed]
- Zunica, E.R.M.; Axelrod, C.L.; Cho, E.; Spielmann, G.; Davuluri, G.; Alexopoulos, S.J.; Beretta, M.; Hoehn, K.L.; Dantas, W.S.; Stadler, K.; et al. Breast cancer growth and proliferation is suppressed by the mitochondrial targeted furazano[3,4-b]pyrazine BAM15. Cancer Metab. 2021, 9, 36. [Google Scholar] [CrossRef]
- Dang, C.P.; Issara-Amphorn, J.; Charoensappakit, A.; Udompornpitak, K.; Bhunyakarnjanarat, T.; Saisorn, W.; Sae-Khow, K.; Leelahavanichkul, A. BAM15, a Mitochondrial Uncoupling Agent, Attenuates Inflammation in the LPS Injection Mouse Model: An Adjunctive Anti-Inflammation on Macrophages and Hepatocytes. J. Innate Immun. 2021, 13, 359–375. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Kapate, N.; Shields IV, C.W.; Mitragotri, S. Drug delivery to macrophages: A review of targeting drugs and drug carriers to macrophages for inflammatory diseases. Adv. Drug Deliv. Rev. 2020, 165, 15–40. [Google Scholar] [CrossRef]
- Mukhtar, M.; Ali, H.; Ahmed, N.; Munir, R.; Talib, S.; Khan, A.S.; Ambrus, R. Drug delivery to macrophages: A review of nano-therapeutics targeted approach for inflammatory disorders and cancer. Expert Opin. Drug Deliv. 2020, 17, 1239–1257. [Google Scholar] [CrossRef]
- Horvath, D.; Basler, M. PLGA Particles in Immunotherapy. Pharmaceutics 2023, 15, 615. [Google Scholar] [CrossRef]
- Loureiro, J.A.; Pereira, M.C. PLGA Based Drug Carrier and Pharmaceutical Applications: The Most Recent Advances. Pharmaceutics 2020, 12, 903. [Google Scholar] [CrossRef]
- Khiewkamrop, P.; Kaewraemruaen, C.; Manipuntee, C.; Saengruengrit, C.; Insin, N.; Leelahavanichkul, A.; Kaewduangduen, W.; Sonpoung, O.; Ariya-Anandech, K.; Hirankarn, N.; et al. Immunosuppressive Polymeric Nanoparticles Targeting Dendritic Cells Alleviate Lupus Disease in Fcgr2b(-/-) Mice by Mediating Antigen-Specific Immune Tolerance. Int. J. Mol. Sci. 2023, 24, 8313. [Google Scholar] [CrossRef]
- Saengruengrit, C.; Ritprajak, P.; Wanichwecharungruang, S.; Sharma, A.; Salvan, G.; Zahn, D.R.; Insin, N. The combined magnetic field and iron oxide-PLGA composite particles: Effective protein antigen delivery and immune stimulation in dendritic cells. J. Colloid Interface Sci. 2018, 520, 101–111. [Google Scholar] [CrossRef]
- Mantzarlis, K.; Tsolaki, V.; Zakynthinos, E. Role of Oxidative Stress and Mitochondrial Dysfunction in Sepsis and Potential Therapies. Oxidative Med. Cell. Longev. 2017, 2017, 5985209. [Google Scholar] [CrossRef]
- Nagar, H.; Piao, S.; Kim, C.-S. Role of Mitochondrial Oxidative Stress in Sepsis. Acute Crit. Care 2018, 33, 65–72. [Google Scholar] [CrossRef]
- Ross, E.A.; Devitt, A.; Johnson, J.R. Macrophages: The Good, the Bad, and the Gluttony. Front. Immunol. 2021, 12, 708186. [Google Scholar] [CrossRef]
- Saisorn, W.; Phuengmaung, P.; Issara-Amphorn, J.; Makjaroen, J.; Visitchanakun, P.; Sae-Khow, K.; Boonmee, A.; Benjaskulluecha, S.; Nita-Lazar, A.; Palaga, T.; et al. Less Severe Lipopolysaccharide-Induced Inflammation in Conditional mgmt-Deleted Mice with LysM-Cre System: The Loss of DNA Repair in Macrophages. Int. J. Mol. Sci. 2023, 24, 10139. [Google Scholar] [CrossRef]
- Song, C.; Xu, J.; Gao, C.; Zhang, W.; Fang, X.; Shang, Y. Nanomaterials targeting macrophages in sepsis: A promising approach for sepsis management. Front. Immunol. 2022, 13, 1026173. [Google Scholar] [CrossRef]
- Yang, L.; Lou, X.; Hao, S.; Zhou, L.; Hou, Y. Macrophage-targeted Nanomedicine for Sepsis: Diagnosis and Therapy. Curr. Pharm. Des. 2023, 29, 2036–2049. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.P.; Leelahavanichkul, A. Over-expression of miR-223 induces M2 macrophage through glycolysis alteration and attenuates LPS-induced sepsis mouse model, the cell-based therapy in sepsis. PLoS ONE 2020, 15, e0236038. [Google Scholar] [CrossRef] [PubMed]
- Dhole, S.M.; Amnerkar, N.D.; Khedekar, P.B. Comparison of UV spectrophotometry and high performance liquid chromatography methods for the determination of repaglinide in tablets. Pharm. Methods 2012, 3, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Visitchanakun, P.; Saisorn, W.; Wongphoom, J.; Chatthanathon, P.; Somboonna, N.; Svasti, S.; Fucharoen, S.; Leelahavanichkul, A. Gut leakage enhances sepsis susceptibility in iron-overloaded β-thalassemia mice through macrophage hyperinflammatory responses. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G966–G979. [Google Scholar] [CrossRef]
- Lund, M.E.; To, J.; O’Brien, B.A.; Donnelly, S. The choice of phorbol 12-myristate 13-acetate differentiation protocol influences the response of THP-1 macrophages to a pro-inflammatory stimulus. J. Immunol. Methods 2016, 430, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Genin, M.; Clement, F.; Fattaccioli, A.; Raes, M.; Michiels, C. M1 and M2 macrophages derived from THP-1 cells differentially modulate the response of cancer cells to etoposide. BMC Cancer 2015, 15, 577. [Google Scholar] [CrossRef] [PubMed]
- Sae-Khow, K.; Charoensappakit, A.; Chiewchengchol, D.; Leelahavanichkul, A. High-Dose Intravenous Ascorbate in Sepsis, a Pro-Oxidant Enhanced Microbicidal Activity and the Effect on Neutrophil Functions. Biomedicines 2022, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Baranov, M.V.; Kumar, M.; Sacanna, S.; Thutupalli, S.; Bogaart, G.v.D. Modulation of Immune Responses by Particle Size and Shape. Front. Immunol. 2021, 11, 607945. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, L.; Boudard, D.; Pourchez, J.; Forest, V.; Marmuse, L.; Louis, C.; Bin, V.; Palle, S.; Grosseau, P.; Bernache-Assollant, D.; et al. Quantitative cellular uptake of double fluorescent core-shelled model submicronic particles. J. Nanoparticle Res. 2012, 14, 1221. [Google Scholar] [CrossRef]
- Durai, V.; Murphy, K.M. Functions of Murine Dendritic Cells. Immunity 2016, 45, 719–736. [Google Scholar] [CrossRef] [PubMed]
- Plitas, G.; Burt, B.M.; Stableford, J.A.; Nguyen, H.M.; Welles, A.P.; DeMatteo, R.P. Dendritic cells are required for effective cross-presentation in the murine liver. Hepatology 2008, 47, 1343–1351. [Google Scholar] [CrossRef]
- Wang, J.; Cao, X.; Zhao, J.; Zhao, H.; Wei, J.; Li, Q.; Qi, X.; Yang, Z.; Wang, L.; Zhang, H.; et al. Critical roles of conventional dendritic cells in promoting T cell-dependent hepatitis through regulating natural killer T cells. Clin. Exp. Immunol. 2017, 188, 127–137. [Google Scholar] [CrossRef]
- Li, J.; Shen, L.; Qian, K. Global, regional, and national incidence and mortality of neonatal sepsis and other neonatal infections, 1990–2019. Front. Public Health 2023, 11, 1139832. [Google Scholar] [CrossRef]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef]
- Busiello, R.A.; Savarese, S.; Lombardi, A. Mitochondrial uncoupling proteins and energy metabolism. Front. Physiol. 2015, 6, 36. [Google Scholar] [CrossRef]
- Dang, B.; Gao, Q.; Zhang, L.; Zhang, J.; Cai, H.; Zhu, Y.; Zhong, Q.; Liu, J.; Niu, Y.; Mao, K.; et al. The glycolysis/HIF-1α axis defines the inflammatory role of IL-4-primed macrophages. Cell Rep. 2023, 42, 112471. [Google Scholar] [CrossRef]
- Kelly, B.; O’Neill, L.A. Metabolic reprogramming in macrophages and dendritic cells in innate immunity. Cell Res. 2015, 25, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Marrocco, A.; Ortiz, L.A. Role of metabolic reprogramming in pro-inflammatory cytokine secretion from LPS or silica-activated macrophages. Front. Immunol. 2022, 13, 936167. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, R.; Gu, H.; Zhang, E.; Qu, J.; Cao, W.; Huang, X.; Yan, H.; He, J.; Cai, Z. Metabolic reprogramming in macrophage responses. Biomark. Res. 2021, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, N.; Tsuji, T.; Yamashita, T.; Hayase, N.; Hu, X.; Yuen, P.S.; Star, R.A. BAM15 treats mouse sepsis and kidney injury, linking mortality, mitochondrial DNA, tubule damage, and neutrophils. J. Clin. Investig. 2023, 133, e152401. [Google Scholar] [CrossRef]
- Sanjabi, S.; Zenewicz, L.A.; Kamanaka, M.; Flavell, R.A. Anti-inflammatory and pro-inflammatory roles of TGF-beta, IL-10, and IL-22 in immunity and autoimmunity. Curr. Opin. Pharmacol. 2009, 9, 447–453. [Google Scholar] [CrossRef]
- Mohammad, A.K.; Reineke, J.J. Quantitative Detection of PLGA Nanoparticle Degradation in Tissues following Intravenous Administration. Mol. Pharm. 2013, 10, 2183–2189. [Google Scholar] [CrossRef]
- Haan, J.M.D.; Kraal, G. Innate Immune Functions of Macrophage Subpopulations in the Spleen. J. Innate Immun. 2012, 4, 437–445. [Google Scholar] [CrossRef]
- Lau, A.H.; Thomson, A.W. Dendritic cells and immune regulation in the liver. Gut 2003, 52, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Doroudian, M.; Zanganeh, S.; Abbasgholinejad, E.; Donnelly, S.C. Nanomedicine in Lung Cancer Immunotherapy. Front. Bioeng. Biotechnol. 2023, 11, 1144653. [Google Scholar] [CrossRef] [PubMed]
- Gharavi, A.T.; Hanjani, N.A.; Movahed, E.; Doroudian, M. The role of macrophage subtypes and exosomes in immunomodulation. Cell. Mol. Biol. Lett. 2022, 27, 83. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [PubMed]
- Lam, R.S.; O’Brien-Simpson, N.M.; Holden, J.A.; Lenzo, J.C.; Fong, S.B.; Reynolds, E.C. Unprimed, M1 and M2 Macrophages Differentially Interact with Porphyromonas gingivalis. PLoS ONE 2016, 11, e0158629. [Google Scholar] [CrossRef]
- Hiengrach, P.; Visitchanakun, P.; Tongchairawewat, P.; Tangsirisatian, P.; Jungteerapanich, T.; Ritprajak, P.; Wannigama, D.L.; Tangtanatakul, P.; Leelahavanichkul, A. Sepsis Encephalopathy Is Partly Mediated by miR370-3p-Induced Mitochondrial Injury but Attenuated by BAM15 in Cecal Ligation and Puncture Sepsis Male Mice. Int. J. Mol. Sci. 2022, 23, 5445. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|
| Arginase-1 (Arg1) (mouse) | CTTGGCTTGCTTCGGAACTC | GGAGAAGGCGTTTGCTTAGTT |
| Inducible nitric oxide synthase (Nos2) (mouse) | ACCCACATCTGGCAGAATGAG | AGCCATGACCTTTCGCATTAG |
| Interleukin-1β (Il1b) (mouse) | GAAATGCCACCTTTTGACAGTG | TGGATGCTCTCATCAGGACAG |
| Found in inflammatory zone 1 (Fizz1) (mouse) | GCCAGGTCCTGGAACCTTTC | GGAGCAGGGAGATGCAGATGA |
| Transforming growth factor-β (Tgfb) (mouse) | CAGAGCTGCGCTTGCAGAG | GTCAGCAGCCGGTTACCAAG |
| β-Actin (Actb) (mouse) | CCTGGCACCCAGCACAAT | GCCGATCCACACGGAGTACT |
| Arginase-1 (ARG1) (human) | CTTGGCTTGCTTCGGAACTC | GGAGAAGGCGTTTGCTTAGTTC |
| Inducible nitric oxide synthase (NOS2) (human) | ACCCACATCTGGCAGAATGAG | AGCCATGACCTTTCGCATTAG |
| Interleukin-1β (IL-1β) (human) | CCACAGACCTTCCAGGAGAATG | GTGCAGTTCAGTGATCGTACAGG |
| Found in inflammatory zone 1 (FIZZ1) (human) | GCAAGAAGCTCTCGTGTGCTAG | AACATCCCACGAACCACAGCCA |
| Transforming growth factor-β (TGFB) (human) | CAGAGCTGCGCTTGCAGAG | GTCAGCAGCCGGTTACCAAG |
| β-Actin (ACTB) (human) | CCTGGCACCCAGCACAAT | GCCGATCCACACGGAGTACT |
| Particles | Hydrodynamic Size (nm) | PDI | Zeta Potential (mV) |
|---|---|---|---|
| PLGA | 527 ± 6 | 0.227 ± 0.009 | −36.1 ± 0.3 |
| PLGA-BAM15 | 559 ± 17 | 0.441 ± 0.147 | −30.1 ± 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Udompornpitak, K.; Bhunyakarnjanarat, T.; Saisorn, W.; Manipuntee, C.; Plengplang, K.; Sittichaitaweekul, S.; Jenphatanapong, P.; Udomkarnjananun, S.; Kaewduangduen, W.; Ariya-anandech, K.; et al. Polymeric Particle BAM15 Targeting Macrophages Attenuates the Severity of LPS-Induced Sepsis: A Proof of Concept for Specific Immune Cell-Targeted Therapy. Pharmaceutics 2023, 15, 2695. https://doi.org/10.3390/pharmaceutics15122695
Udompornpitak K, Bhunyakarnjanarat T, Saisorn W, Manipuntee C, Plengplang K, Sittichaitaweekul S, Jenphatanapong P, Udomkarnjananun S, Kaewduangduen W, Ariya-anandech K, et al. Polymeric Particle BAM15 Targeting Macrophages Attenuates the Severity of LPS-Induced Sepsis: A Proof of Concept for Specific Immune Cell-Targeted Therapy. Pharmaceutics. 2023; 15(12):2695. https://doi.org/10.3390/pharmaceutics15122695
Chicago/Turabian StyleUdompornpitak, Kanyarat, Thansita Bhunyakarnjanarat, Wilasinee Saisorn, Chonnavee Manipuntee, Kittawat Plengplang, Samarch Sittichaitaweekul, Panisa Jenphatanapong, Suwasin Udomkarnjananun, Warerat Kaewduangduen, Kasirapat Ariya-anandech, and et al. 2023. "Polymeric Particle BAM15 Targeting Macrophages Attenuates the Severity of LPS-Induced Sepsis: A Proof of Concept for Specific Immune Cell-Targeted Therapy" Pharmaceutics 15, no. 12: 2695. https://doi.org/10.3390/pharmaceutics15122695