
Enhanced Immunomodulation, Anti-Apoptosis, and Improved Tear Dynamics of (PEG)-BHD1028, a Novel Adiponectin Receptor Agonist Peptide, for Treating Dry Eye Disease

 , , and
, , and 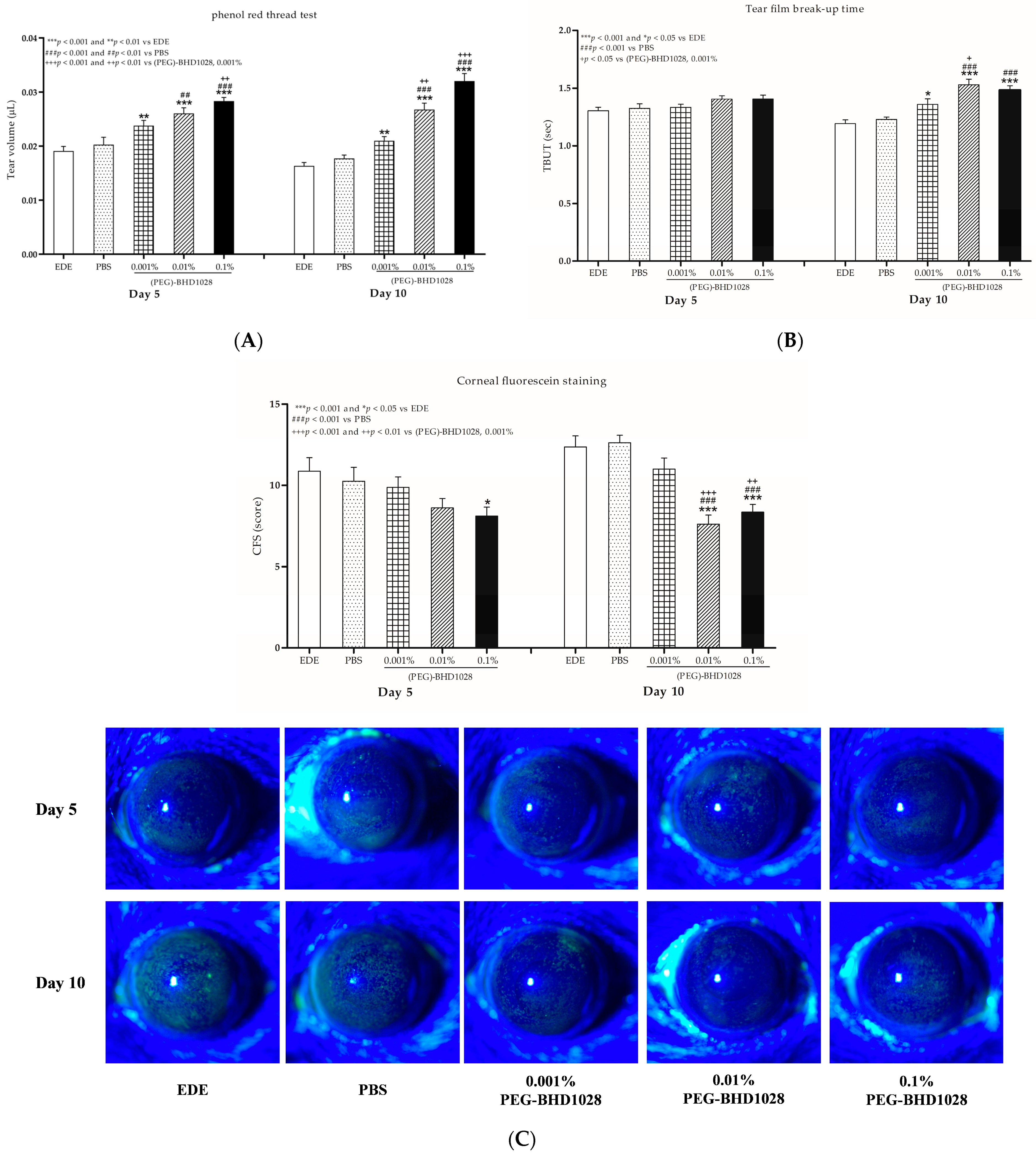{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Baseline Ophthalmic Examinations in Experimentally Induced Dry Eye (EDE) on Day 0
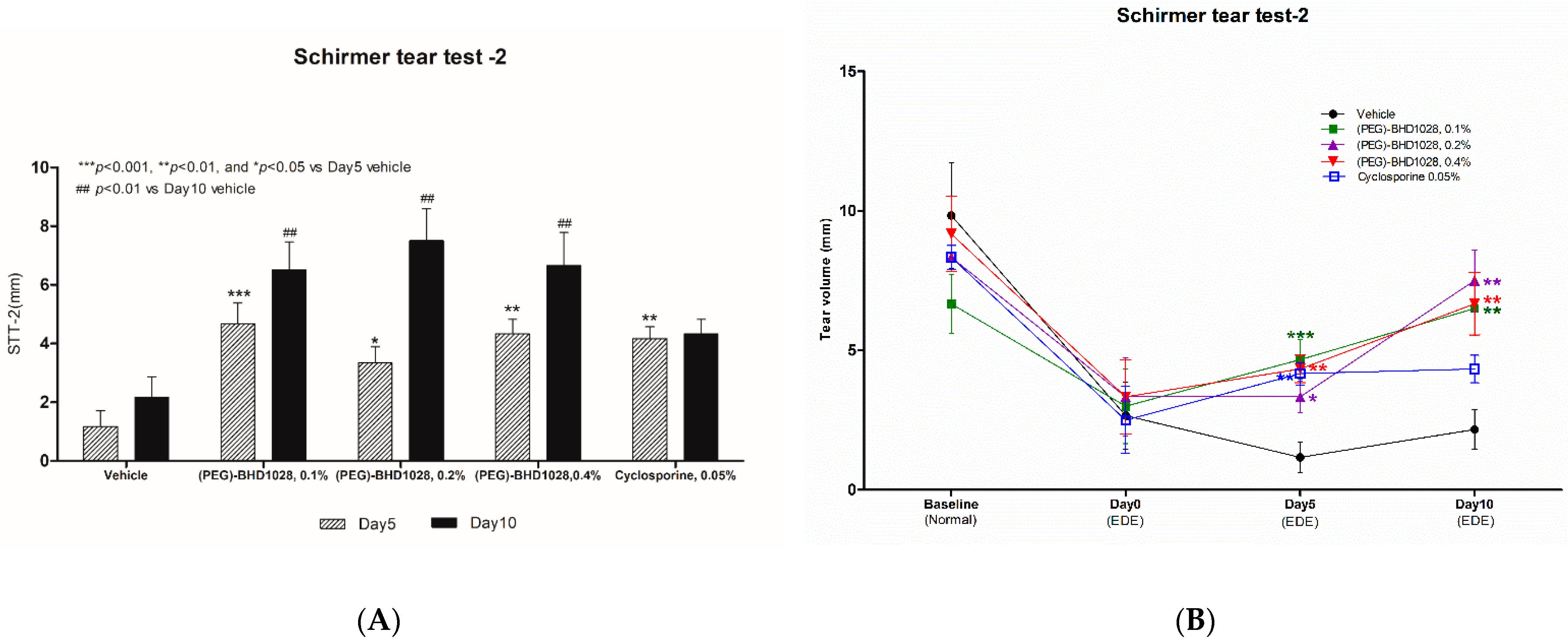
2.2. Evaluation of the Tear Production in Each EDE Animal Model
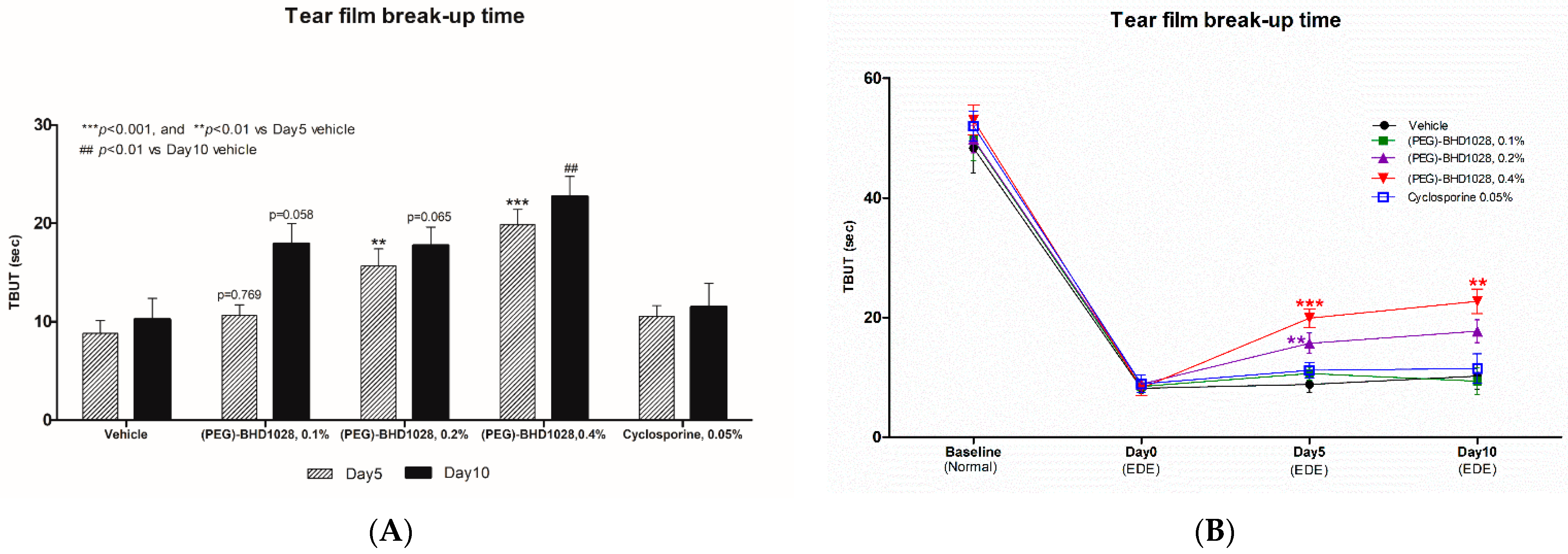
2.3. Evaluation of TBUT
2.4. Evaluation of the CFS Score
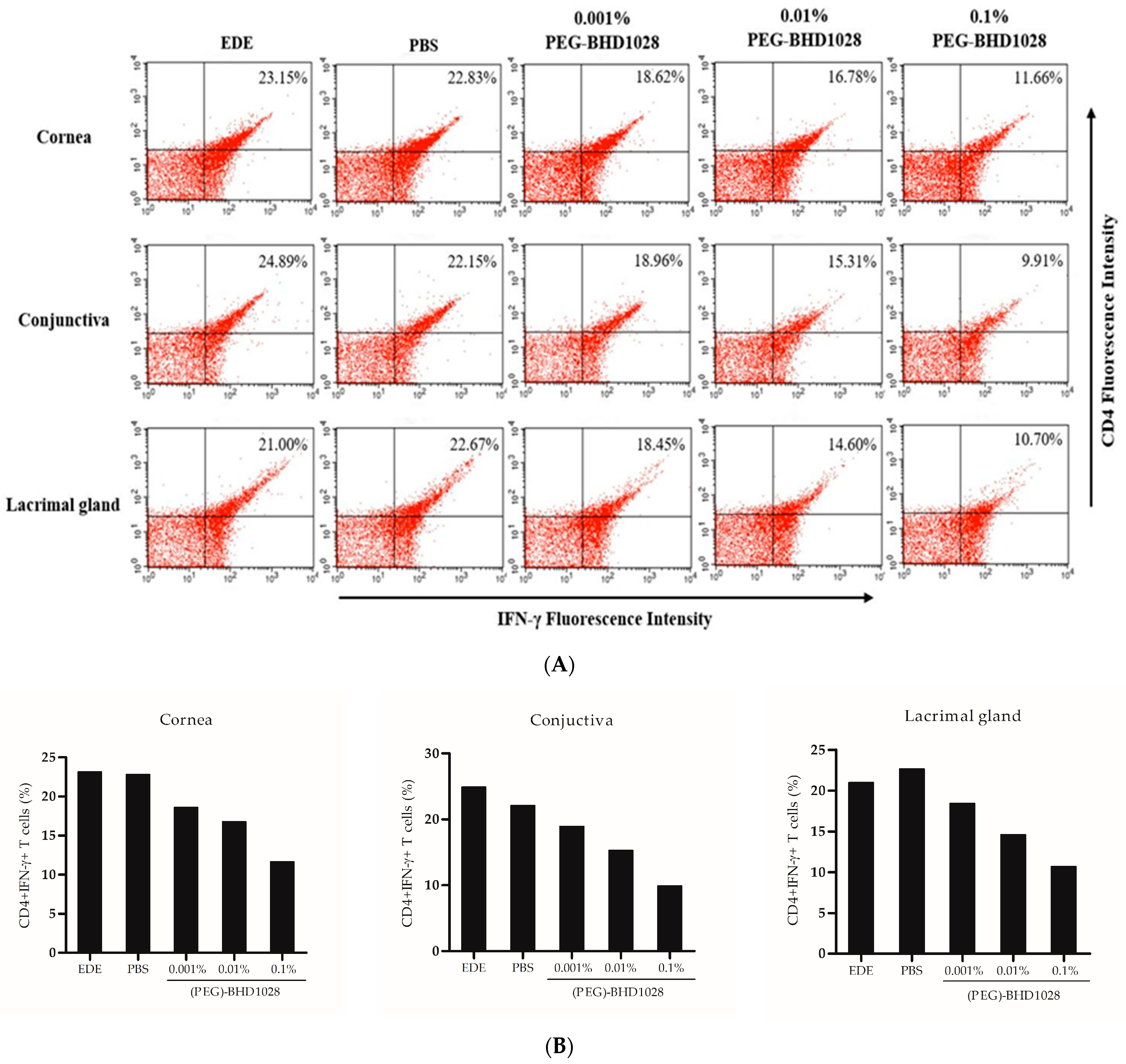
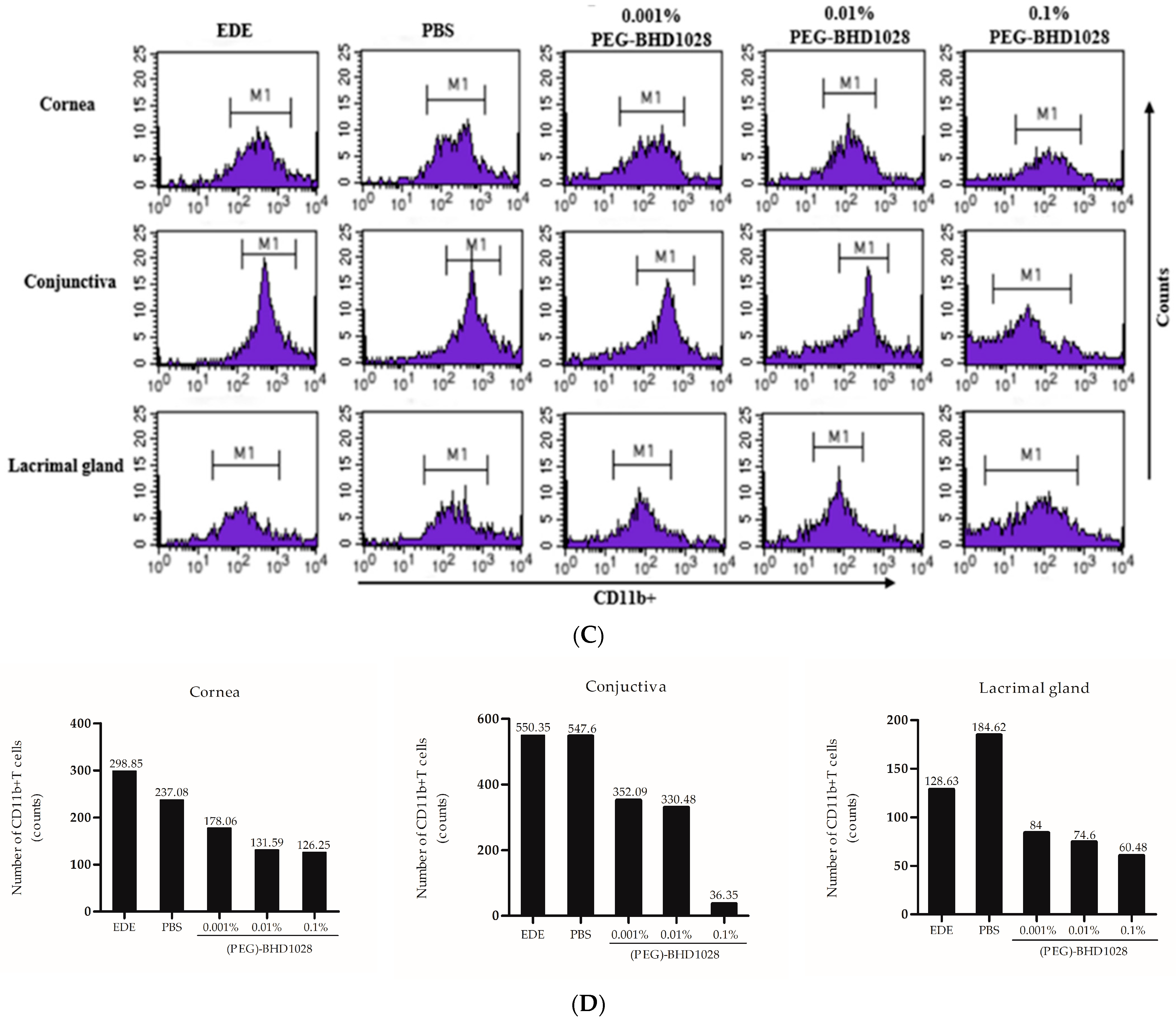
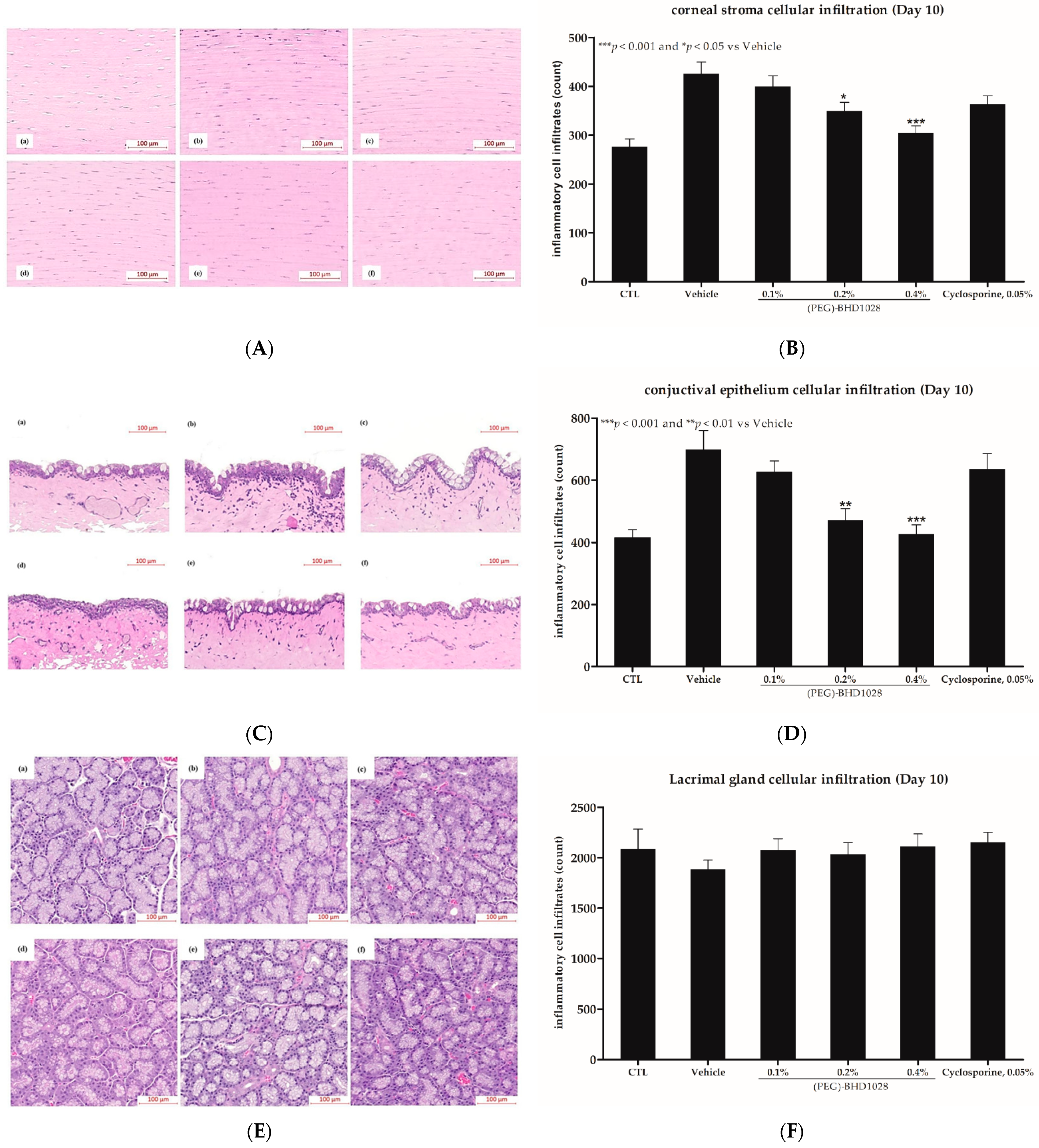
2.5. Anti-Inflammatory Effect
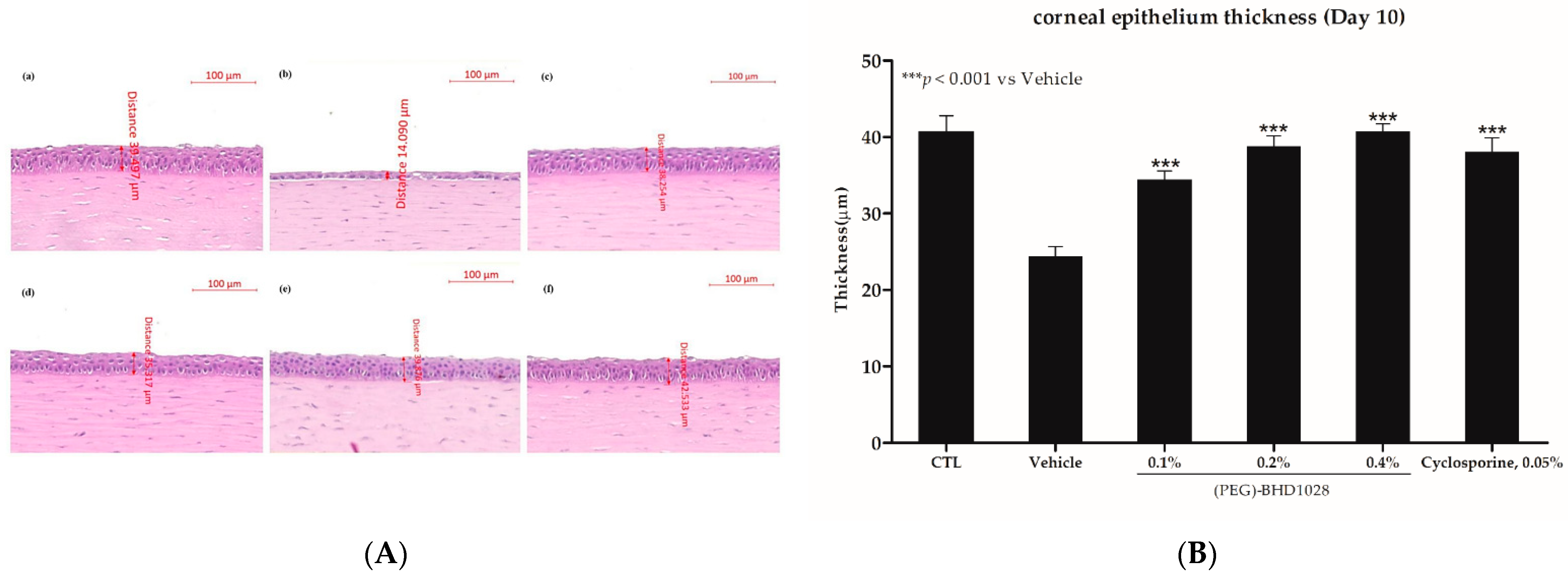
2.6. Corneal Epithelium Thickness Evaluation in the EDE Rabbit Model
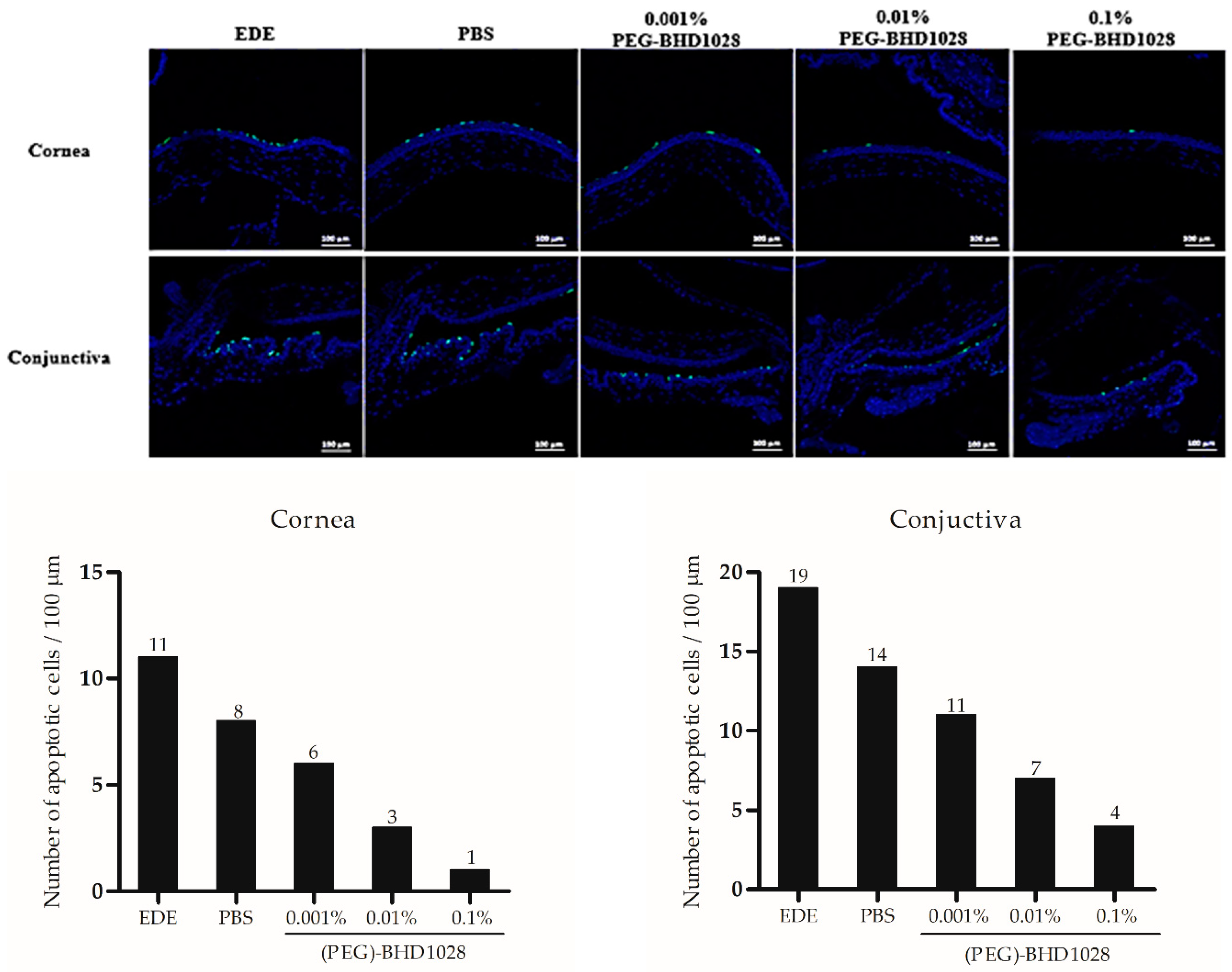
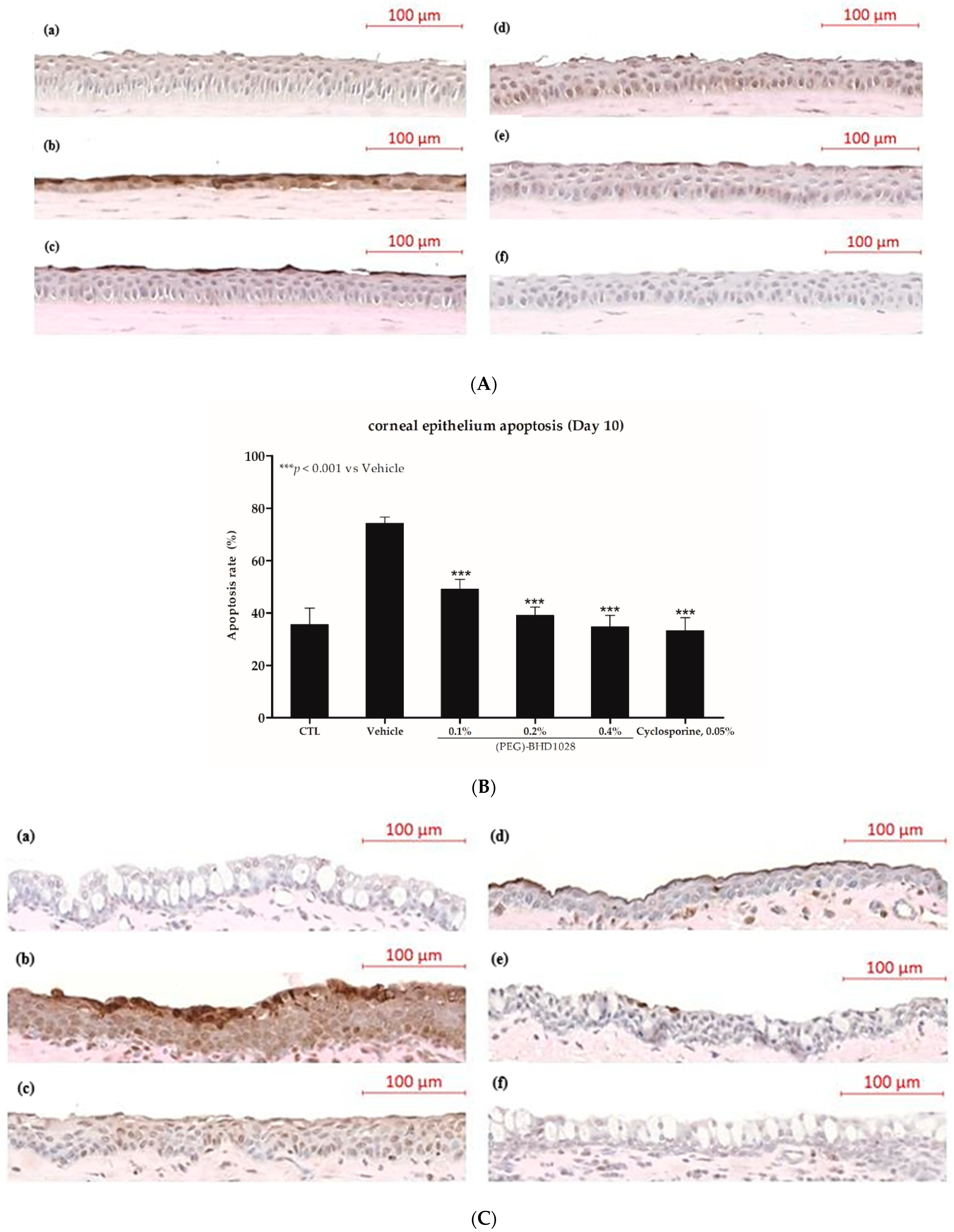
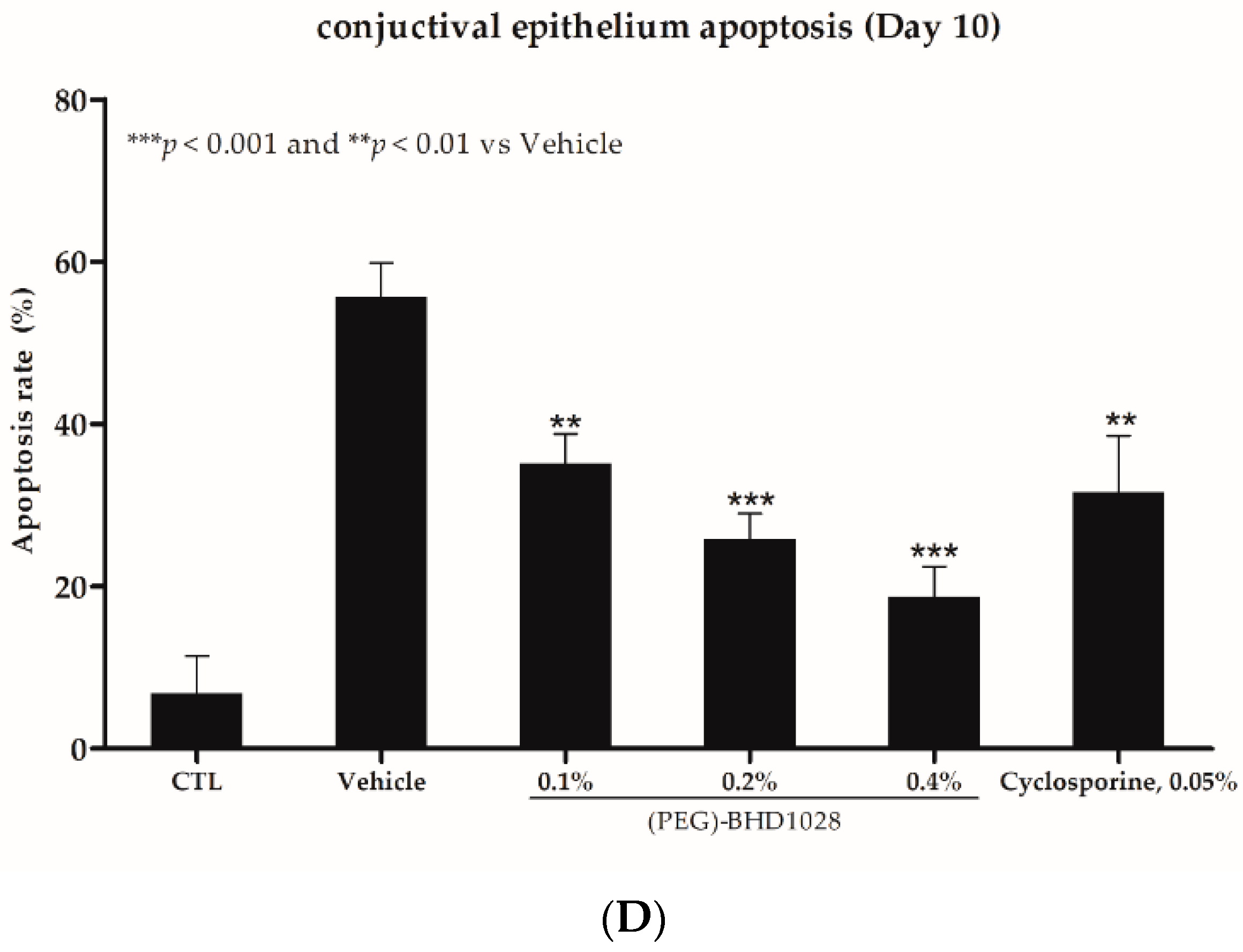
2.7. Anti-Apoptotic Effect of (PEG)-BHD1028 in EDE Mouse and Rabbit Models
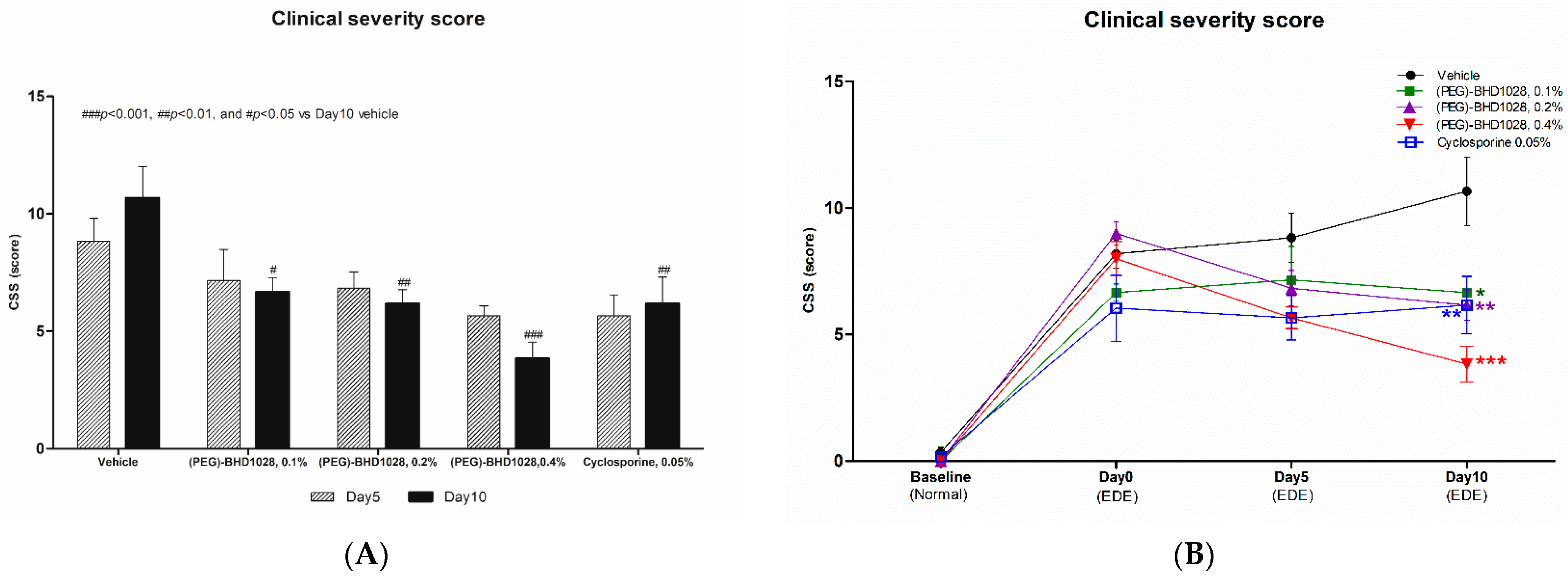
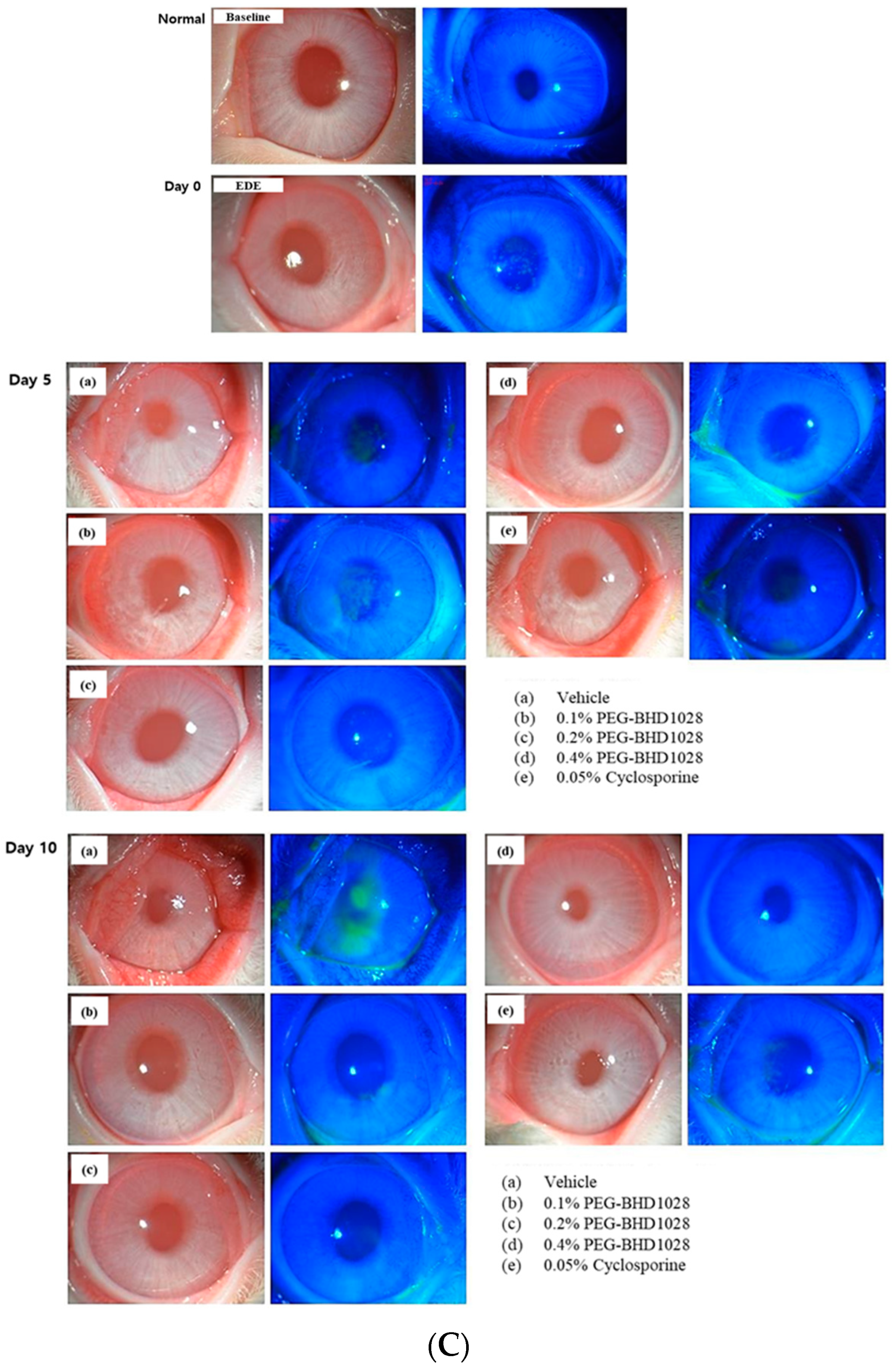
2.8. Comparative Analysis between 0.4% (PEG)-BHD1028 and 0.05% Cyclosporine in the Rabbit EDE Model on Day 10
3. Discussion
4. Materials and Methods
4.1. Synthesis of (PEG)-BHD1028
4.2. Animals
4.3. Study Design and DED Modeling
4.4. Tear Volume Determination
4.5. Tear Break-Up Time
4.6. Evaluating Corneal Fluorescent Staining
4.7. Evaluation of the Ocular Surface Inflammation in Mice
4.8. Histopathological Evaluation of the Inflammatory Cell Infiltration on the Ocular Surface in Rabbits
4.9. Apoptosis Evaluation
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, E.C. Diagnosis and Treatment of Dry Eye Syndrome. JKMA 2018, 61, 352–364. [Google Scholar] [CrossRef]
- Ortiz, G.; Chao, C.; Jamali, A.; Seyed-Razavi, Y.; Kenyon, B.; Harris, D.L.; Zoukhri, D.; Hamrah, P. Effect of Dry Eye Disease on the Kinetics of Lacrimal Gland Dendritic Cells as Visualized by Intravital Multi-Photon Microscopy. Front. Immunol. 2020, 11, 1713. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, F.; Alves, M.; Bunya, V.Y.; Jalbert, I.; Lekhanont, K.; Malet, F.; Na, K.S.; Schaumberg, D.; Uchino, M.; Vehof, J.; et al. TFOS DEWS II Epidemiology Report. Ocul. Surf. 2017, 15, 334–365. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.M.D. Adiponectin: Role in Physiology and Pathophysiology. Int. J. Prev. Med. 2020, 11, 136. [Google Scholar] [CrossRef]
- Li, H.Y.; Hong, X.; Cao, Q.Q.; So, K.F. Adiponectin, exercise and eye diseases. Int. Rev. Neurobiol. 2019, 147, 281–294. [Google Scholar] [CrossRef]
- Li, Z.; Woo, J.M.; Chung, S.W.; Kwon, M.Y.; Choi, J.S.; Oh, H.J.; Yoon, K.C. Therapeutic effect of topical adiponectin in a mouse model of desiccating stress-induced dry eye. Investig. Ophthalmol. Vis. Sci. 2013, 54, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Nishinaka, A.; Nakamura, S.; Tanaka, M.; Masuda, T.; Inoue, Y.; Yamamoto, T.; Imai, T.; Hidaka, Y.; Shimazawa, M.; Hara, H. Excess adiponectin in eyes with progressive ocular vascular diseases. FASEB J. 2021, 35, e21313. [Google Scholar] [CrossRef]
- Sluch, V.M.; Banks, A.; Li, H.; Crowley, M.A.; Davis, V.; Xiang, C.; Yang, J.; Demirs, J.T.; Vrouvlianis, J.; Leehy, B.; et al. ADIPOR1 is essential for vision and its RPE expression is lost in the Mfrp(rd6) mouse. Sci. Rep. 2018, 8, 14339. [Google Scholar] [CrossRef] [Green Version]
- Bora, P.S.; Kaliappan, S.; Lyzogubov, V.V.; Tytarenko, R.G.; Thotakura, S.; Viswanathan, T.; Bora, N.S. Expression of adiponectin in choroidal tissue and inhibition of laser induced choroidal neovascularization by adiponectin. FEBS Lett. 2007, 581, 1977–1982. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.K.; Kim, G.; Kim, D.H.; Kim, B.B. PEG-BHD1028 Peptide Regulates Insulin Resistance and Fatty Acid Beta-Oxidation, and Mitochondrial Biogenesis by Binding to Two Heterogeneous Binding Sites of Adiponectin Receptors, AdipoR1 and AdipoR2. Int. J. Mol. Sci. 2021, 22, 884. [Google Scholar] [CrossRef]
- Doughty, M.J. Tear Film Stability and Tear Break Up Time (TBUT) in Laboratory Rabbits—A Systematic Review. Curr. Eye Res. 2018, 43, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Perez, V.L.; Stern, M.E.; Pflugfelder, S.C. Inflammatory basis for dry eye disease flares. Exp. Eye Res. 2020, 201, 108294. [Google Scholar] [CrossRef] [PubMed]
- Lio, C.T.; Dhanda, S.K.; Bose, T. Cluster Analysis of Dry Eye Disease Models Based on Immune Cell Parameters—New Insight into Therapeutic Perspective. Front. Immunol. 2020, 11, 1930. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Prins, J.; Venkatesh, B. Clinical review: Adiponectin biology and its role in inflammation and critical illness. Crit. Care 2011, 15, 221. [Google Scholar] [CrossRef] [Green Version]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A novel serum protein similar to C1q, produced exclusively in adipocytes. J. Biol. Chem. 1995, 270, 26746–26749. [Google Scholar] [CrossRef] [Green Version]
- Ouchi, N.; Walsh, K. Adiponectin as an anti-inflammatory factor. Clin. Chim. Acta 2007, 380, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Ajuwon, K.M.; Spurlock, M.E. Adiponectin inhibits LPS-induced NF-kappaB activation and IL-6 production and increases PPARgamma2 expression in adipocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1220–R1225. [Google Scholar] [CrossRef]
- Mandal, P.; Park, P.H.; McMullen, M.R.; Pratt, B.T.; Nagy, L.E. The anti-inflammatory effects of adiponectin are mediated via a heme oxygenase-1-dependent pathway in rat Kupffer cells. Hepatology 2010, 51, 1420–1429. [Google Scholar] [CrossRef] [Green Version]
- Neumeier, M.; Weigert, J.; Schaffler, A.; Wehrwein, G.; Muller-Ladner, U.; Scholmerich, J.; Wrede, C.; Buechler, C. Different effects of adiponectin isoforms in human monocytic cells. J. Leukoc. Biol. 2006, 79, 803–808. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, K.; Parker, J.L.; Ouchi, N.; Higuchi, A.; Vita, J.A.; Gokce, N.; Pedersen, A.A.; Kalthoff, C.; Tullin, S.; Sams, A.; et al. Adiponectin promotes macrophage polarization toward an anti-inflammatory phenotype. J. Biol. Chem. 2010, 285, 6153–6160. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.M.; Wolf, D.; Rumpold, H.; Enrich, B.; Tilg, H. Adiponectin induces the anti-inflammatory cytokines IL-10 and IL-1RA in human leukocytes. Biochem. Biophys. Res. Commun. 2004, 323, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Wulster-Radcliffe, M.C.; Ajuwon, K.M.; Wang, J.; Christian, J.A.; Spurlock, M.E. Adiponectin differentially regulates cytokines in porcine macrophages. Biochem. Biophys. Res. Commun. 2004, 316, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, Y.; Liu, Z.; He, L. Adiponectin promotes repair of renal tubular epithelial cells by regulating mitochondrial biogenesis and function. Metabolism 2022, 128, 154959. [Google Scholar] [CrossRef]
- Kraus, D.; Winter, J.; Jepsen, S.; Jager, A.; Meyer, R.; Deschner, J. Interactions of adiponectin and lipopolysaccharide from Porphyromonas gingivalis on human oral epithelial cells. PLoS ONE 2012, 7, e30716. [Google Scholar] [CrossRef] [Green Version]
- Nigro, E.; Scudiero, O.; Sarnataro, D.; Mazzarella, G.; Sofia, M.; Bianco, A.; Daniele, A. Adiponectin affects lung epithelial A549 cell viability counteracting TNFalpha and IL-1ss toxicity through AdipoR1. Int. J. Biochem. Cell Biol. 2013, 45, 1145–1153. [Google Scholar] [CrossRef]
- Yang, Q.; Fu, C.; Zhang, X.; Zhang, Z.; Zou, J.; Xiao, J.; Ye, Z. Adiponectin protects against uric acidinduced renal tubular epithelial inflammatory responses via the AdipoR1/AMPK signaling pathway. Int. J. Mol. Med. 2019, 43, 1542–1552. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Liu, Y.; Tan, L.; Yan, L.; Zuo, X. Retraction Note: Adiponectin administration alleviates DSS-induced colonic inflammation in Caco-2 cells and mice. Inflamm. Res. 2022, 71, 727. [Google Scholar] [CrossRef]
- Lin, T.; Qiu, Y.; Liu, Y.; Mohan, R.; Li, Q.; Lei, B. Expression of adiponectin and its receptors in type 1 diabetes mellitus in human and mouse retinas. Mol. Vis. 2013, 19, 1769–1778. [Google Scholar]
- Kim, S.; Lee, Y.; Kim, J.W.; Son, Y.J.; Ma, M.J.; Um, J.H.; Kim, N.D.; Min, S.H.; Kim, D.I.; Kim, B.B. Discovery of a novel potent peptide agonist to adiponectin receptor 1. PLoS ONE 2018, 13, e0199256. [Google Scholar] [CrossRef]
- Kurata, M.; Atsumi, I.; Yamagiwa, Y.; Sakaki, H. Ocular Instillation Toxicity Study: Current Status and Points to Consider on Study Design and Evaluation. Fundam. Toxicol. Sci. 2016, 3, 217–232. [Google Scholar] [CrossRef]
- Alam, J.; de Paiva, C.S.; Pflugfelder, S.C. Immune—Goblet cell interaction in the conjunctiva. Ocul. Surf. 2020, 18, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Pflugfelder, S.C.; Bian, F.; Gumus, K.; Farley, W.; Stern, M.E.; De Paiva, C.S. Severity of Sjogren’s Syndrome Keratoconjunctivitis Sicca Increases with Increased Percentage of Conjunctival Antigen-Presenting Cells. Int. J. Mol. Sci. 2018, 19, 2760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuazo, F.; Lopez-Ponce, D.; Salinas-Toro, D.; Valenzuela, F.; Sans-Puroja, J.; Srur, M.; Lopez-Solis, R.O.; Traipe-Castro, L. Conjunctival impression cytology in patients with normal and impaired OSDI scores. Arch. Soc. Esp. Oftalmol. 2014, 89, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Pflugfelder, S.C.; De Paiva, C.S.; Moore, Q.L.; Volpe, E.A.; Li, D.Q.; Gumus, K.; Zaheer, M.L.; Corrales, R.M. Aqueous Tear Deficiency Increases Conjunctival Interferon-gamma (IFN-gamma) Expression and Goblet Cell Loss. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7545–7550. [Google Scholar] [CrossRef] [Green Version]
- Villareal, A.L.; Farley, W.; Pflugfelder, S.C. Effect of topical ophthalmic epinastine and olopatadine on tear volume in mice. Eye Contact Lens 2006, 32, 272–276. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, I.-K.; Yoon, K.-C.; Kang, S.-S.; Seon, S.-K.; Lee, K.; Kim, B.B. Enhanced Immunomodulation, Anti-Apoptosis, and Improved Tear Dynamics of (PEG)-BHD1028, a Novel Adiponectin Receptor Agonist Peptide, for Treating Dry Eye Disease. Pharmaceutics 2023, 15, 78. https://doi.org/10.3390/pharmaceutics15010078
Lee I-K, Yoon K-C, Kang S-S, Seon S-K, Lee K, Kim BB. Enhanced Immunomodulation, Anti-Apoptosis, and Improved Tear Dynamics of (PEG)-BHD1028, a Novel Adiponectin Receptor Agonist Peptide, for Treating Dry Eye Disease. Pharmaceutics. 2023; 15(1):78. https://doi.org/10.3390/pharmaceutics15010078
Chicago/Turabian StyleLee, In-Kyung, Kyung-Chul Yoon, Seong-Soo Kang, Su-Kyung Seon, Kwanghyun Lee, and Brian B. Kim. 2023. "Enhanced Immunomodulation, Anti-Apoptosis, and Improved Tear Dynamics of (PEG)-BHD1028, a Novel Adiponectin Receptor Agonist Peptide, for Treating Dry Eye Disease" Pharmaceutics 15, no. 1: 78. https://doi.org/10.3390/pharmaceutics15010078