
Combination of a Chaperone Synthesis Inducer and an Inhibitor of GAPDH Aggregation for Rehabilitation after Traumatic Brain Injury: A Pilot Study

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
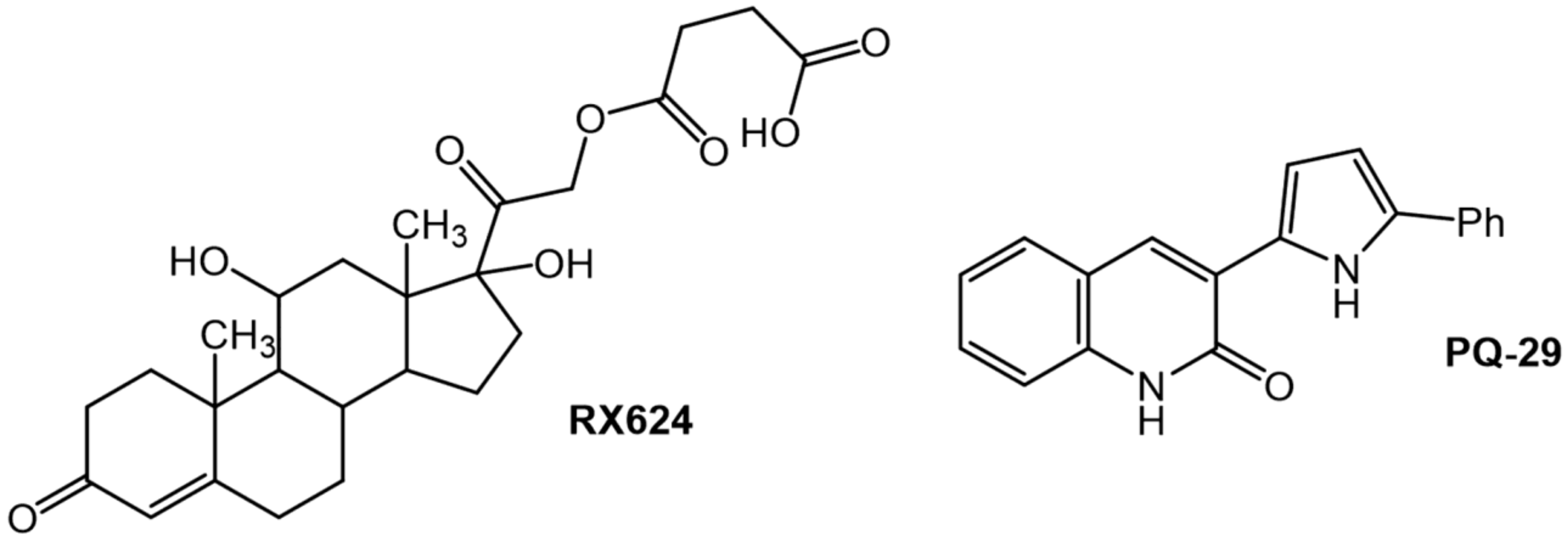
2.2. Chemical Compounds
2.3. Modeling of Traumatic Brain Injury
2.4. Cell Proliferation Assay
2.5. Apoptotic Cell Analysis
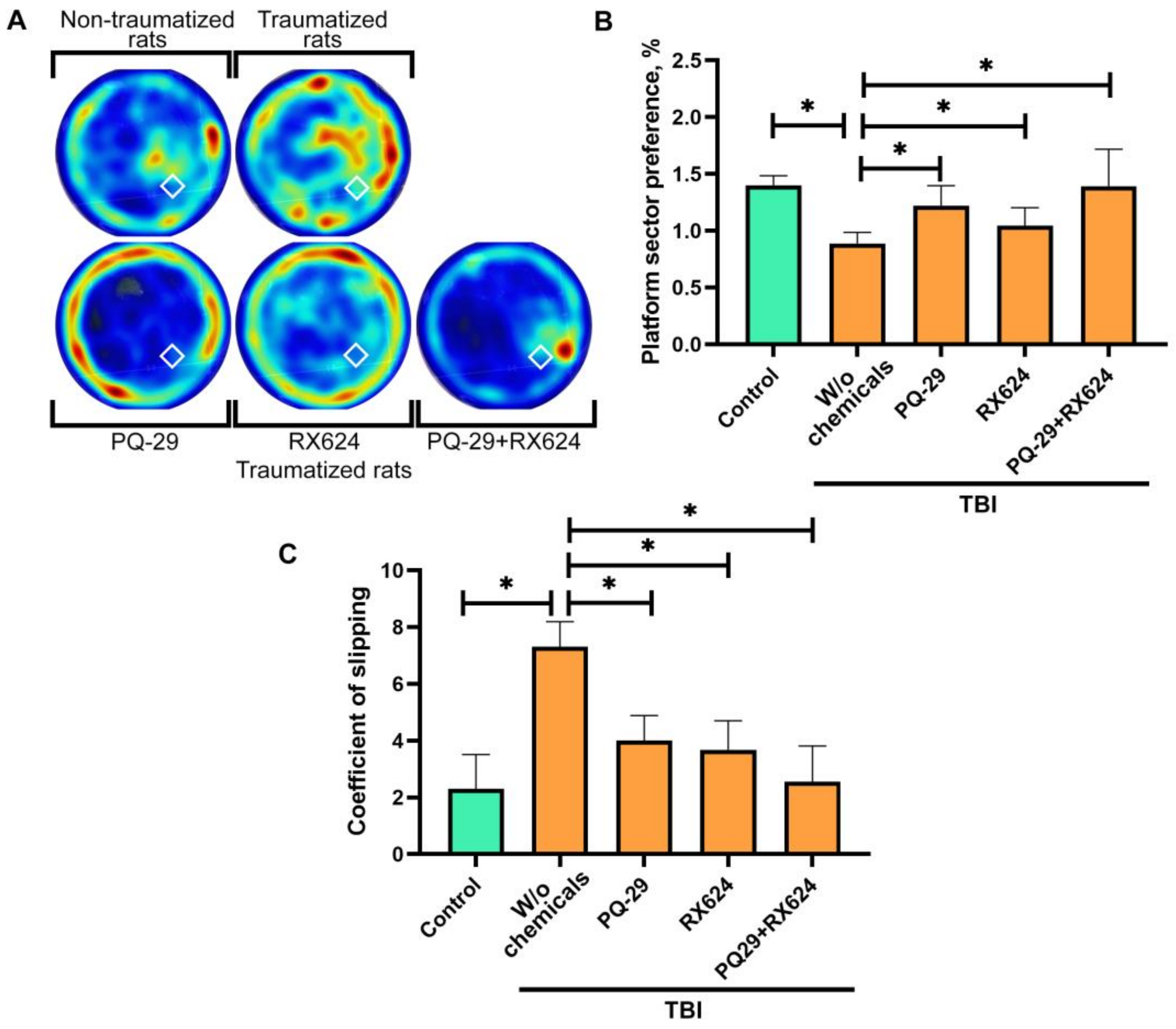
2.6. Morris Water Test
2.7. Analysis of the Locomotor Function of Rats
2.8. Immunohistochemistry
2.9. Aggregate Detection
2.10. Statistical Analysis
3. Results
3.1. Effect of the Combined Use of RX624 and PQ-29 Preparations on the Physiological Parameters of C6 Cells in an In Vitro TBI Model
3.2. Effect of Combination Therapy Using RX624 and PQ-29 on the Behavioral, Hystological and Physiological Characteristics of Traumatized Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dewan, M.C.; Rattani, A.; Gupta, S.; Baticulon, R.E.; Hung, Y.C.; Punchak, M.; Agrawal, A.; Adeleye, A.O.; Shrime, M.G.; Rubiano, A.M.; et al. Estimating the global incidence of traumatic brain injury. J. Neurosurg. 2018, 130, 1080–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andriessen, T.M.J.C.; Jacobs, B.; Vos, P.E. Clinical characteristics and pathophysiological mechanisms of focal and diffuse traumatic brain injury. J. Cell. Mol. Med. 2010, 14, 2381–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabet, N.; Soltani, Z.; Khaksari, M. Multipotential and systemic effects of traumatic brain injury. J. Neuroimmunol. 2021, 357, 577619. [Google Scholar] [CrossRef] [PubMed]
- Hakiminia, B.; Alikiaii, B.; Khorvash, F.; Mousavi, S. Oxidative stress and mitochondrial dysfunction following traumatic brain injury: From mechanistic view to targeted therapeutic opportunities. Fundam. Clin. Pharmacol. 2022, 36, 612–662. [Google Scholar] [CrossRef] [PubMed]
- Edwards, G.; Moreno-Gonzalez, I.; Soto, C. Amyloid-beta and tau pathology following repetitive mild traumatic brain injury. Biochem. Biophys. Res. Commun. 2017, 483, 1137–1142. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Campolo, M.; Bruschetta, G.; Crupi, R.; Cordaro, M.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Traumatic Brain Injury Leads to Development of Parkinson’s Disease Related Pathology in Mice. Front. Neurosci. 2016, 10, 458. [Google Scholar] [CrossRef] [Green Version]
- Wright, D.K.; Liu, S.; van der Poel, C.; McDonald, S.J.; Brady, R.D.; Taylor, L.; Yang, L.; Gardner, A.J.; Ordidge, R.; O’Brien, T.J.; et al. Traumatic Brain Injury Results in Cellular, Structural and Functional Changes Resembling Motor Neuron Disease. Cereb. Cortex 2016, 27, 4503–4515. [Google Scholar] [CrossRef] [Green Version]
- Thapa, K.; Khan, H.; Singh, T.G.; Kaur, A. Traumatic Brain Injury: Mechanistic Insight on Pathophysiology and Potential Therapeutic Targets. J. Mol. Neurosci. 2021, 71, 1725–1742. [Google Scholar] [CrossRef]
- Cossu, G. Therapeutic options to enhance coma arousal after traumatic brain injury: State of the art of current treatments to improve coma recovery. Br. J. Neurosurg. 2014, 28, 187–198. [Google Scholar] [CrossRef]
- Kondrikov, D.; Fulton, D.; Dong, Z.; Su, Y. Heat Shock Protein 70 Prevents Hyperoxia-Induced Disruption of Lung Endothelial Barrier via Caspase-Dependent and AIF-Dependent Pathways. PLoS ONE 2015, 10, e0129343. [Google Scholar] [CrossRef]
- Mosser, D.D.; Caron, A.W.; Bourget, L.; Denis-Larose, C.; Massie, B. Role of the human heat shock protein hsp70 in protection against stress-induced apoptosis. Mol. Cell. Biol. 1997, 17, 5317–5327. [Google Scholar] [CrossRef] [Green Version]
- Lackie, R.E.; Maciejewski, A.; Ostapchenko, V.G.; Marques-Lopes, J.; Choy, W.Y.; Duennwald, M.L.; Prado, V.F.; Prado, M.A.M. The Hsp70/Hsp90 chaperone machinery in neurodegenerative diseases. Front. Neurosci. 2017, 11, 254. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Zhu, B.; Murshid, A.; Adachi, H.; Song, B.; Lee, A.; Liu, C.; Calderwood, S.K. T cell activation by heat shock protein 70 vaccine requires TLR signaling and scavenger receptor expressed by endothelial cells-1. J. Immunol. 2009, 183, 3092–3098. [Google Scholar] [CrossRef] [Green Version]
- Duncan, E.J.; Cheetham, M.E.; Chapple, J.P.; van der Spuy, J. The Role of HSP70 and Its Co-chaperones in Protein Misfolding, Aggregation and Disease. Subcell. Biochem. 2015, 78, 243–273. [Google Scholar] [CrossRef]
- Dutysheva, E.A.; Mikeladze, M.A.; Trestsova, M.A.; Aksenov, N.D.; Utepova, I.A.; Mikhaylova, E.R.; Suezov, R.V.; Charushin, V.N.; Chupakhin, O.N.; Guzhova, I.V.; et al. Pyrrolylquinoxaline-2-one derivative as a potent therapeutic factor for brain trauma rehabilitation. Pharmaceutics 2020, 12, 414. [Google Scholar] [CrossRef]
- Dutysheva, E.A.; Utepova, I.A.; Trestsova, M.A.; Anisimov, A.S.; Charushin, V.N.; Chupakhin, O.N.; Margulis, B.A.; Guzhova, I.V.; Lazarev, V.F. Synthesis and approbation of new neuroprotective chemicals of pyrrolyl- and indolylazine classes in a cell model of Alzheimer’s disease. Eur. J. Med. Chem. 2021, 222, 113577. [Google Scholar] [CrossRef]
- Gerszon, J.; Rodacka, A. Oxidatively modified glyceraldehyde-3-phosphate dehydrogenase in neurodegenerative processes and the role of low molecular weight compounds in counteracting its aggregation and nuclear translocation. Ageing Res. Rev. 2018, 48, 21–31. [Google Scholar] [CrossRef]
- Tristan, C.; Shahani, N.; Sedlak, T.W.; Sawa, A. The diverse functions of GAPDH: Views from different subcellular compartments. Cell. Signal. 2011, 23, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Naletova, I.; Schmalhausen, E.; Kharitonov, A.; Katrukha, A.; Saso, L.; Caprioli, A.; Muronetz, V. Non-native glyceraldehyde-3-phosphate dehydrogenase can be an intrinsic component of amyloid structures. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2008, 1784, 2052–2058. [Google Scholar] [CrossRef]
- Guzhova, I.V.; Lazarev, V.F.; Kaznacheeva, A.V.; Ippolitova, M.V.; Muronetz, V.I.; Kinev, A.V.; Margulis, B.A. Novel mechanism of Hsp70 chaperone-mediated prevention of polyglutamine aggregates in a cellular model of huntington disease. Hum. Mol. Genet. 2011, 20, 3953–3963. [Google Scholar] [CrossRef]
- Lazarev, V.F.; Sverchinskyi, D.V.; Ippolitova, M.V.; Stepanova, A.V.; Guzhova, I.V.; Margulis, B.A. Factors Affecting Aggregate Formation in Cell Models of Huntington’s Disease and Amyotrophic Lateral Sclerosis. Acta Nat. 2013, 5, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Lazarev, V.F.; Nikotina, A.D.; Semenyuk, P.I.; Evstafyeva, D.B.; Mikhaylova, E.R.; Muronetz, V.I.; Shevtsov, M.A.; Tolkacheva, A.V.; Dobrodumov, A.V.; Shavarda, A.L.; et al. Small molecules preventing GAPDH aggregation are therapeutically applicable in cell and rat models of oxidative stress. Free Radic. Biol. Med. 2016, 92, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Lazarev, V.F.; Dutysheva, E.A.; Komarova, E.Y.; Mikhaylova, E.R.; Guzhova, I.V.; Margulis, B.A. GAPDH-targeted therapy—A new approach for secondary damage after traumatic brain injury on rats. Biochem. Biophys. Res. Commun. 2018, 4, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Utepova, I.A.; Trestsova, M.A.; Chupakhin, O.N.; Charushin, V.N.; Rempel, A.A. Aerobic oxidative C-H/C-H coupling of azaaromatics with indoles and pyrroles in the presence of TiO2 as a photocatalyst. Green Chem. 2015, 17, 4401–4410. [Google Scholar] [CrossRef]
- Mychasiuk, R.; Farran, A.; Angoa-Perez, M.; Briggs, D.; Kuhn, D.; Esser, M.J. A Novel Model of Mild Traumatic Brain Injury for Juvenile Rats. J. Vis. Exp. 2014, 94, e51820. [Google Scholar] [CrossRef] [Green Version]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Silachev, D.N.; Shubina, M.I.; Iankauskas, S.S.; Mkrtchian, V.P.; Manskikh, V.N.; Guliaev, M.V.; Zorov, D.B. Evaluation of a long-term sensomotor deficit after neonatal rat brain ischemia/hypoxia. Zhurnal Vyss. Nervn. Deiatelnosti Im. IP Pavlov. 2013, 63, 405–416. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: Sydney, Australia, 1982; ISBN 978-0-12-547620-1. [Google Scholar]
- Baracaldo-Santamaría, D.; Ariza-Salamanca, D.F.; Corrales-Hernández, M.G.; Pachón-Londoño, M.J.; Hernandez-Duarte, I.; Calderon-Ospina, C.A. Revisiting Excitotoxicity in Traumatic Brain Injury: From Bench to Bedside. Pharmaceutics 2022, 14, 152. [Google Scholar] [CrossRef]
- Schäfer, M.K.E.; Pfeiffer, A.; Jaeckel, M.; Pouya, A.; Dolga, A.M.; Methner, A. Regulators of mitochondrial Ca2+ homeostasis in cerebral ischemia. Cell Tissue Res. 2014, 357, 395–405. [Google Scholar] [CrossRef]
- Quillinan, N.; Herson, P.S.; Traystman, R.J. Neuropathophysiology of Brain Injury. Anesthesiol. Clin. 2016, 34, 453–464. [Google Scholar] [CrossRef]
- Smrcka, M.; Vidlák, M.; Máca, K.; Smrcka, V.; Gál, R. The influence of mild hypothermia on ICP, CPP and outcome in patients with primary and secondary brain injury. Acta Neurochir. Suppl. 2005, 95, 273–275. [Google Scholar]
- Broussard, J.I.; Acion, L.; De Jesús-Cortés, H.; Yin, T.; Britt, J.K.; Salas, R.; Costa-Mattioli, M.; Robertson, C.; Pieper, A.A.; Arciniegas, D.B.; et al. Repeated mild traumatic brain injury produces neuroinflammation, anxiety-like behaviour and impaired spatial memory in mice. Brain Inj. 2018, 32, 113–122. [Google Scholar] [CrossRef]
- Hiskens, M.I. Targets of neuroprotection and review of pharmacological interventions in traumatic brain injury. J. Pharmacol. Exp. Ther. 2022, 382, 149–166. [Google Scholar] [CrossRef]
- Nakajima, H.; Kubo, T.; Ihara, H.; Hikida, T.; Danjo, T.; Nakatsuji, M.; Shahani, N.; Itakura, M.; Ono, Y.; Azuma, Y.-T.; et al. Nuclear-translocated Glyceraldehyde-3-phosphate Dehydrogenase Promotes Poly(ADP-ribose) Polymerase-1 Activation during Oxidative/Nitrosative Stress in Stroke. J. Biol. Chem. 2015, 290, 14493–14503. [Google Scholar] [CrossRef] [Green Version]
- Yenari, M.A.; Liu, J.; Zheng, Z.; Vexler, Z.S.; Lee, J.E.; Giffard, R.G. Antiapoptotic and anti-inflammatory mechanisms of heat-shock protein protection. Ann. N. Y. Acad. Sci. 2005, 1053, 74–83. [Google Scholar] [CrossRef]
- Nakajima, H.; Amano, W.; Fujita, A.; Fukuhara, A.; Azuma, Y.-T.; Hata, F.; Inui, T.; Takeuchi, T. The Active Site Cysteine of the Proapoptotic Protein Glyceraldehyde-3-phosphate Dehydrogenase Is Essential in Oxidative Stress-induced Aggregation and Cell Death. J. Biol. Chem. 2007, 282, 26562–26574. [Google Scholar] [CrossRef]
- Lazarev, V.F.; Mikhaylova, E.R.; Guzhova, I.V.; Margulis, B.A. Possible function of molecular chaperones in diseases caused by propagating amyloid aggregates. Front. Neurosci. 2017, 11, 277. [Google Scholar] [CrossRef] [Green Version]
- Lazarev, V.F.; Guzhova, I.V.; Margulis, B.A. Glyceraldehyde-3-phosphate dehydrogenase is a multifaceted therapeutic target. Pharmaceutics 2020, 12, 416. [Google Scholar] [CrossRef]
- Itakura, M.; Nakajima, H.; Semi, Y.; Higashida, S.; Azuma, Y.-T.; Takeuchi, T. Glyceraldehyde-3-phosphate dehydrogenase aggregation inhibitor peptide: A potential therapeutic strategy against oxidative stress-induced cell death. Biochem. Biophys. Res. Commun. 2015, 467, 373–376. [Google Scholar] [CrossRef] [Green Version]
- Sunaga, K.; Takahashi, H.; Chuang, D.M.; Ishitani, R. Glyceraldehyde-3-phosphate dehydrogenase is over-expressed during apoptotic death of neuronal cultures and is recognized by a monoclonal antibody against amyloid plaques from Alzheimer’s brain. Neurosci. Lett. 1995, 200, 133–136. [Google Scholar] [CrossRef]
- Katsube, N.; Sunaga, K.; Aishita, H.; Chuang, D.M.; Ishitani, R. ONO-1603, a potential antidementia drug, delays age-induced apoptosis and suppresses overexpression of glyceraldehyde-3-phosphate dehydrogenase in cultured central nervous system neurons. J. Pharmacol. Exp. Ther. 1999, 288, 6–13. [Google Scholar] [PubMed]
- Zhao, Z.; Faden, A.I.; Loane, D.J.; Lipinski, M.M.; Sabirzhanov, B.; Stoica, B.A. Neuroprotective effects of geranylgeranylacetone in experimental traumatic brain injury. J. Cereb. Blood Flow Metab. 2013, 33, 1897–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitarska, D.; Tylki-Szymańska, A.; Ługowska, A. Treatment trials in Niemann-Pick type C disease. Metab. Brain Dis. 2021, 36, 2215–2221. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, W.; Qu, Y.; Feng, C.; Wang, D.; Yin, H.; Li, C.; Sun, Z.; Sun, D. Development and Application of Medicine-Engineering Integration in the Rehabilitation of Traumatic Brain Injury. BioMed Res. Int. 2021, 2021, 9962905. [Google Scholar] [CrossRef]
- Krainer, M. Efficacy of Combination Therapy Versus Monotherapy. Breast Cancer Res. Treat. 2003, 81, S11–S15. [Google Scholar] [CrossRef]
- Lazarev, V.F.; Nikotina, A.D.; Semenyuk, P.I.; Evstafyeva, D.B.; Mikhaylova, E.R.; Muronetz, V.I.; Shevtsov, M.A.; Tolkacheva, A.V.; Dobrodumov, A.V.; Shavarda, A.L.; et al. Properties of substances inhibiting aggregation of oxidized GAPDH: Data on the interaction with the enzyme and the impact on its intracellular content. Data Br. 2016, 7, 524–528. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dutysheva, E.A.; Mikhaylova, E.R.; Trestsova, M.A.; Andreev, A.I.; Apushkin, D.Y.; Utepova, I.A.; Serebrennikova, P.O.; Akhremenko, E.A.; Aksenov, N.D.; Bon’, E.I.; et al. Combination of a Chaperone Synthesis Inducer and an Inhibitor of GAPDH Aggregation for Rehabilitation after Traumatic Brain Injury: A Pilot Study. Pharmaceutics 2023, 15, 7. https://doi.org/10.3390/pharmaceutics15010007
Dutysheva EA, Mikhaylova ER, Trestsova MA, Andreev AI, Apushkin DY, Utepova IA, Serebrennikova PO, Akhremenko EA, Aksenov ND, Bon’ EI, et al. Combination of a Chaperone Synthesis Inducer and an Inhibitor of GAPDH Aggregation for Rehabilitation after Traumatic Brain Injury: A Pilot Study. Pharmaceutics. 2023; 15(1):7. https://doi.org/10.3390/pharmaceutics15010007
Chicago/Turabian StyleDutysheva, Elizaveta A., Elena R. Mikhaylova, Maria A. Trestsova, Alexander I. Andreev, Danila Yu. Apushkin, Irina A. Utepova, Polina O. Serebrennikova, Eugenia A. Akhremenko, Nikolay D. Aksenov, Elizaveta I. Bon’, and et al. 2023. "Combination of a Chaperone Synthesis Inducer and an Inhibitor of GAPDH Aggregation for Rehabilitation after Traumatic Brain Injury: A Pilot Study" Pharmaceutics 15, no. 1: 7. https://doi.org/10.3390/pharmaceutics15010007