
A Self-Emulsified Adjuvant System Containing the Immune Potentiator Alpha Tocopherol Induces Higher Neutralizing Antibody Responses than a Squalene-Only Emulsion When Evaluated with a Recombinant Cytomegalovirus (CMV) Pentamer Antigen in Mice
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Formulation Materials
2.2. Sterile Filtration of Emulsions
2.3. Percent Content of Squalene and α-Tocopherol in Emulsions
2.4. Optimization of the SE-AS
2.5. Cryo-Electron Microscopy (Cryo-EM) Imaging
2.6. In Vitro Innate Immune Response Using Human Whole Blood
2.7. In Vivo Evaluations Using CMV Pentamer Antigen
3. Results
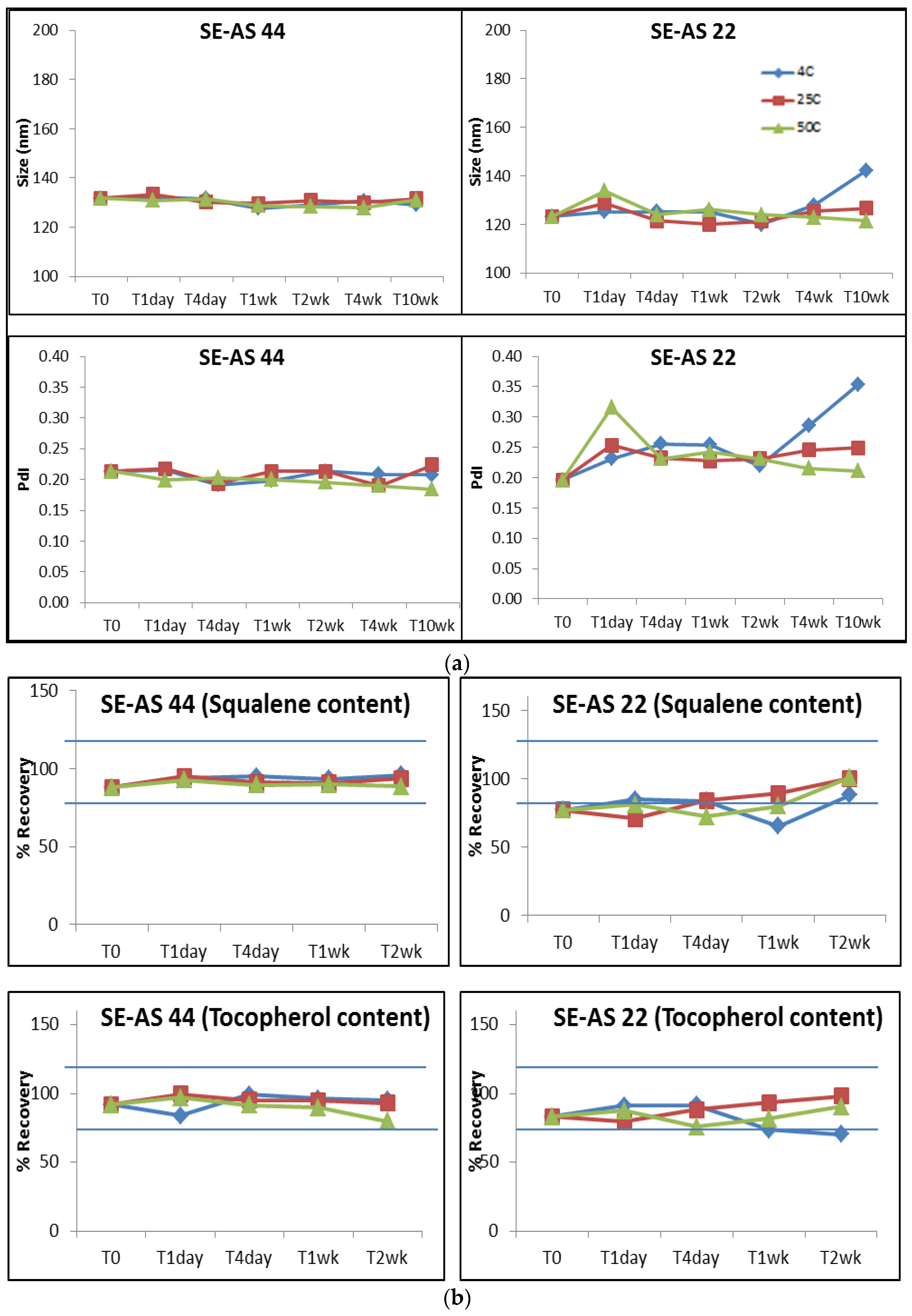
3.1. Optimization of the SE-AS Emulsions
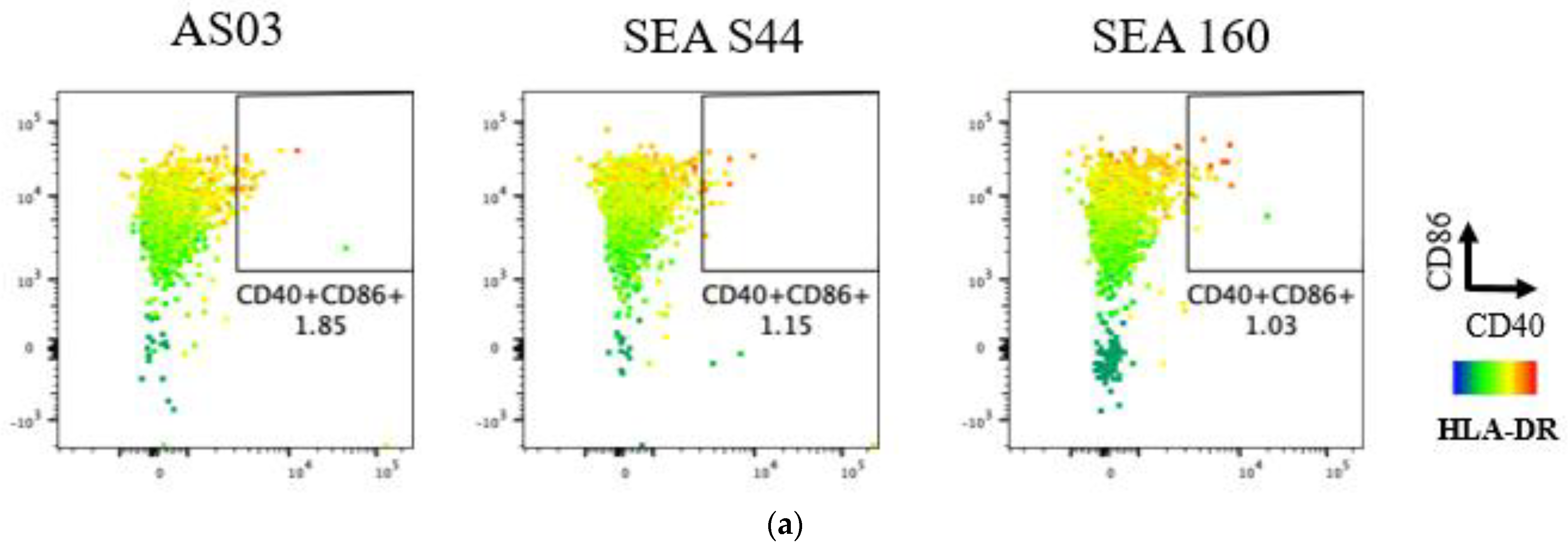
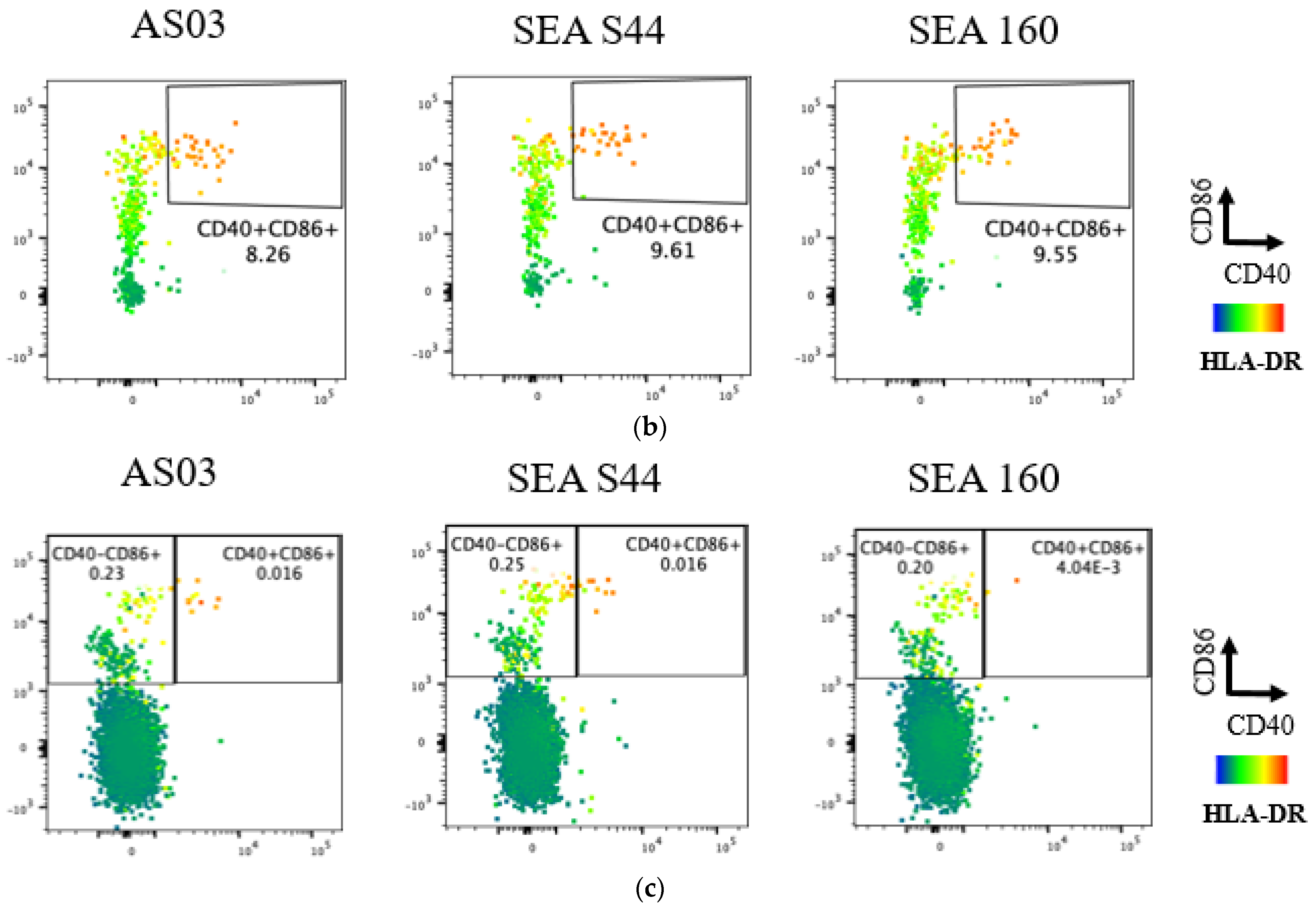
3.2. In Vitro Evaluation of Novel Emulsion Adjuvants Using Human Innate Signaling in Whole Blood Cells
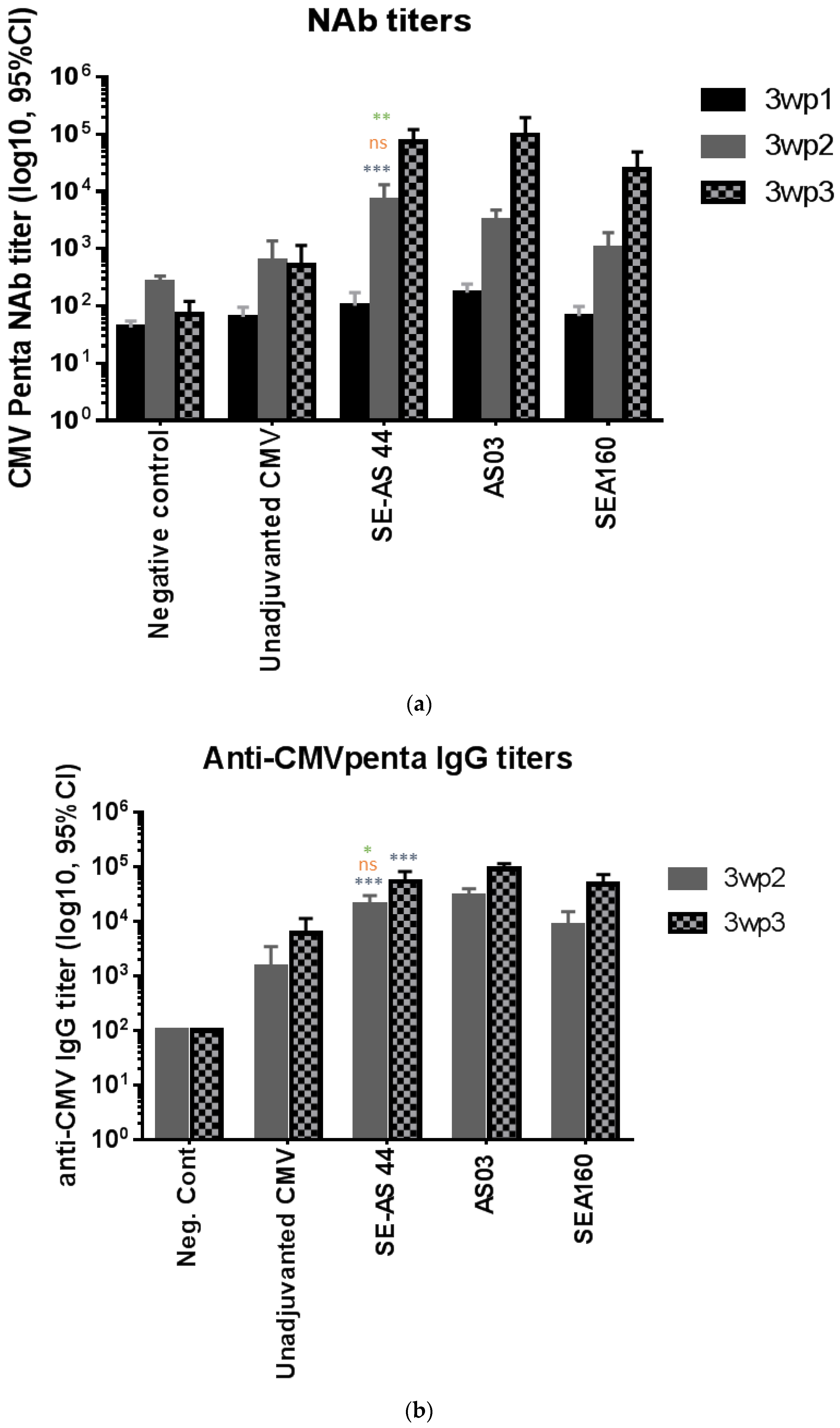
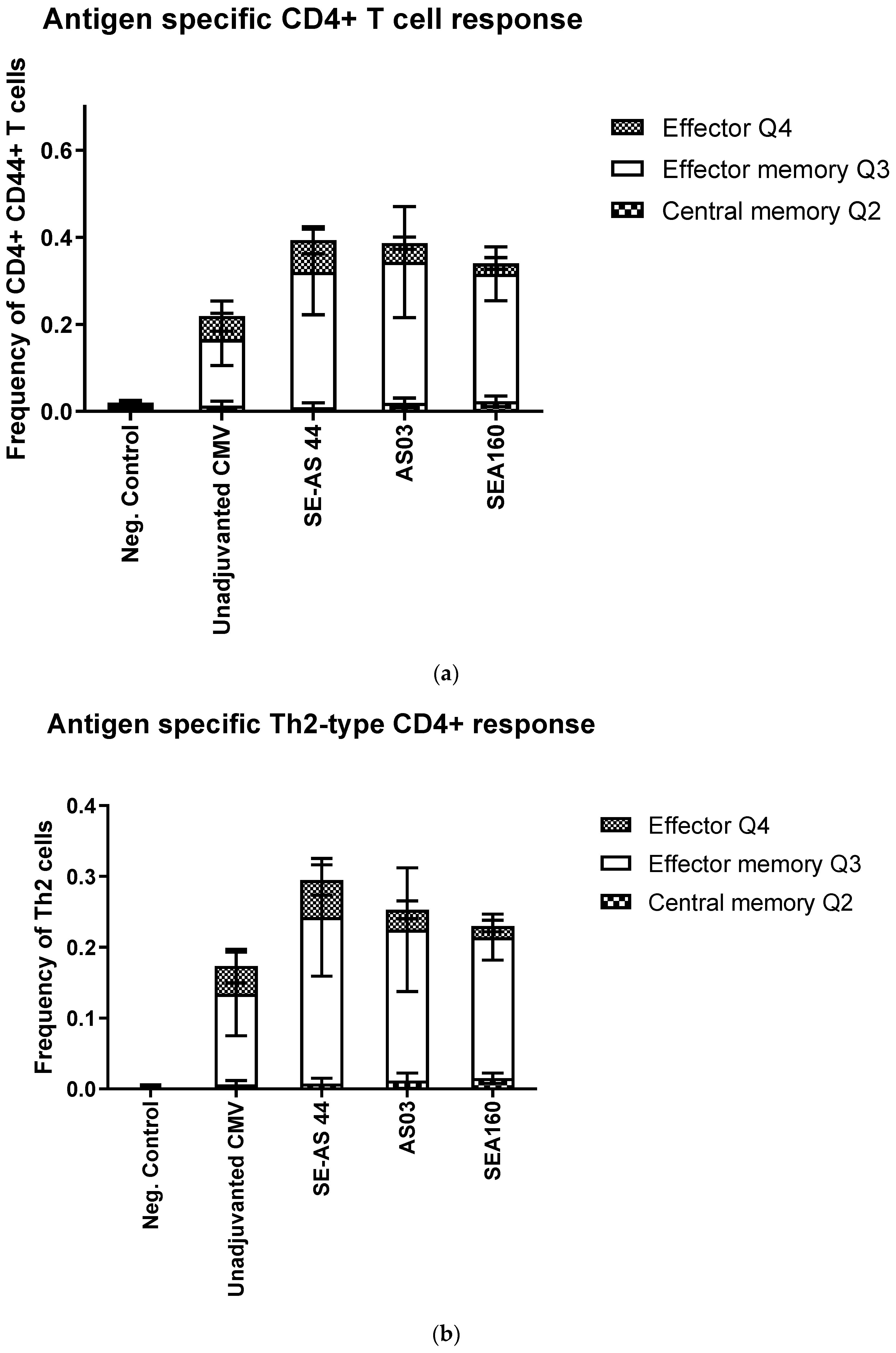
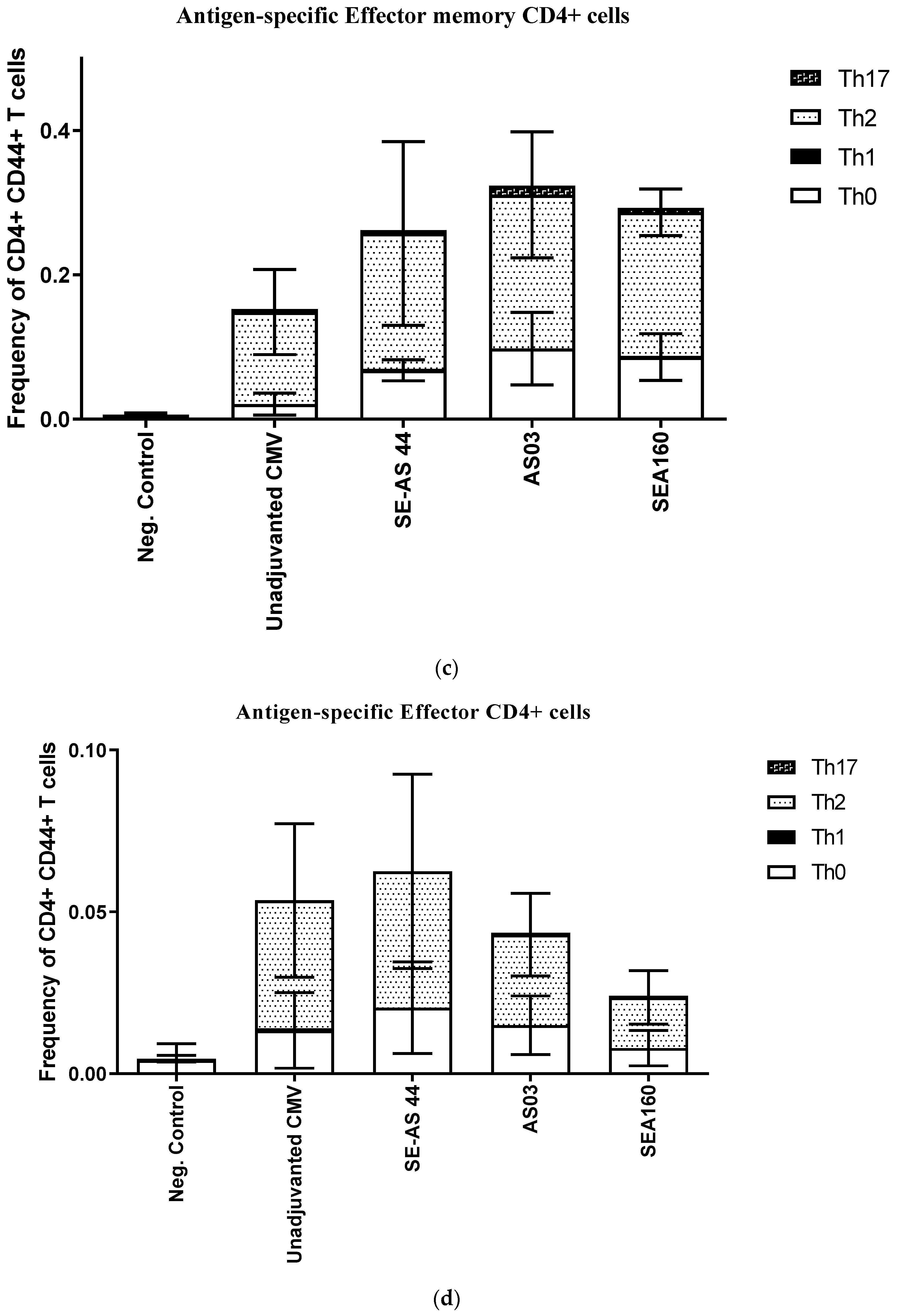
3.3. In Vivo Potency of Emulsion Adjuvants in Mice Using CMV Pentamer Antigen
4. Discussion
5. Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fox, C.B.; Haensler, J. An update on safety and immunogenicity of vaccines containing emulsion-based adjuvants. Expert Rev. Vaccines 2013, 12, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Vogel, F.R.; Caillet, C.; Kusters, I.C.; Haensler, J. Emulsion-based adjuvants for influenza vaccines. Expert Rev. Vaccines 2009, 8, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.R.; Brito, L.A.; O’Hagan, D.T.; Amiji, M.M. Emulsions as Vaccine Adjuvants. In Subunit Vaccine Delivery; Foged, C., Rades, T., Perrie, Y., Hook, S., Eds.; Springer: New York, NY, USA, 2015; pp. 59–76. [Google Scholar]
- Rappuoli, R.; Dormitzer, P.R. Influenza: Options to improve pandemic preparation. Science 2012, 336, 1531–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughn, D.W.; Seifert, H.; Hepburn, A.; Dewe, W.; Li, P.; Drame, M.; Cohet, C.; Innis, B.L.; Fries, L.F. Safety of AS03-adjuvanted inactivated split virion A (H1N1) pdm09 and H5N1 influenza virus vaccines administered to adults: Pooled analysis of 28 clinical trials. Hum. Vaccines Immunother. 2014, 10, 2942–2957. [Google Scholar] [CrossRef] [Green Version]
- Carmona, A.; Omeñaca, F.; Tejedor, J.C.; Merino, J.M.; Vaman, T.; Dieussaert, I.; Gillard, P.; Arístegui, J. Immunogenicity and safety of AS03-adjuvanted 2009 influenza A H1N1 vaccine in children 6–35 months. Vaccine 2010, 28, 5837–5844. [Google Scholar] [CrossRef]
- Hager, K.J.; Pérez Marc, G.; Gobeil, P.; Diaz, R.S.; Heizer, G.; Llapur, C.; Makarkov, A.I.; Vasconcellos, E.; Pillet, S.; Riera, F.; et al. Efficacy and Safety of a Recombinant Plant-Based Adjuvanted COVID-19 Vaccine. N. Engl. J. Med. 2022, 386, 2084–2096. [Google Scholar] [CrossRef] [PubMed]
- Ward, B.J.; Gobeil, P.; Séguin, A.; Atkins, J.; Boulay, I.; Charbonneau, P.-Y.; Couture, M.; D’Aoust, M.-A.; Dhaliwall, J.; Finkle, C. Phase 1 randomized trial of a plant-derived virus-like particle vaccine for COVID-19. Nature medicine 2021, 27, 1071–1078. [Google Scholar] [CrossRef]
- Chappell, K.J.; Mordant, F.L.; Li, Z.; Wijesundara, D.K.; Ellenberg, P.; Lackenby, J.A.; Cheung, S.T.; Modhiran, N.; Avumegah, M.S.; Henderson, C.L. Safety and immunogenicity of an MF59-adjuvanted spike glycoprotein-clamp vaccine for SARS-CoV-2: A randomised, double-blind, placebo-controlled, phase 1 trial. Lancet Infect. Dis. 2021, 21, 1383–1394. [Google Scholar] [CrossRef]
- O’Hagan, D.T.; van der Most, R.; Lodaya, R.N.; Coccia, M.; Lofano, G. “World in motion”–emulsion adjuvants rising to meet the pandemic challenges. npj Vaccines 2021, 6, 158. [Google Scholar] [CrossRef]
- Bishai, D.; McQuestion, M.; Chaudhry, R.; Wigton, A. The costs of scaling up vaccination in the world’s poorest countries. Health Aff. 2006, 25, 348–356. [Google Scholar] [CrossRef]
- Stavaru, C.; Onu, A.; Lupulescu, E.; Tucureanu, C.; Rasid, O.; Vlase, E.; Coman, C.; Caras, I.; Ghiorghisor, A.; Berbecila, L.; et al. Technology transfer of oil-in-water emulsion adjuvant manufacturing for pandemic influenza vaccine production in Romania: Preclinical evaluation of split virion inactivated H5N1 vaccine with adjuvant. Hum. Vaccines Immunother. 2016, 12, 1009–1026. [Google Scholar] [CrossRef] [Green Version]
- Fox, C.B.; Huynh, C.; O’Hara, M.K.; Onu, A. Technology transfer of oil-in-water emulsion adjuvant manufacturing for pandemic influenza vaccine production in Romania. Vaccine 2013, 31, 1633–1640. [Google Scholar] [CrossRef] [Green Version]
- Lodaya, R.N.; Kanitkar, A.P.; Friedrich, K.; Henson, D.; Yamagata, R.; Nuti, S.; Mallett, C.P.; Bertholet, S.; Amiji, M.M.; O’Hagan, D.T. Formulation Design, Optimization and In Vivo Evaluations of an α-Tocopherol-Containing Self-Emulsified Adjuvant System using Inactivated Influenza Vaccine. J. Control. Release 2019, 316, 12–21. [Google Scholar] [CrossRef]
- Shah, R.R.; Dodd, S.; Schaefer, M.; Ugozzoli, M.; Singh, M.; Otten, G.R.; Amiji, M.M.; O’Hagan, D.T.; Brito, L.A. The Development of Self-Emulsifying Oil-in-Water Emulsion Adjuvant and an Evaluation of the Impact of Droplet Size on Performance. J. Pharm. Sci. 2015, 104, 1352–1361. [Google Scholar] [CrossRef] [PubMed]
- Morel, S.; Didierlaurent, A.; Bourguignon, P.; Delhaye, S.; Baras, B.; Jacob, V.; Planty, C.; Elouahabi, A.; Harvengt, P.; Carlsen, H.; et al. Adjuvant System AS03 containing alpha-tocopherol modulates innate immune response and leads to improved adaptive immunity. Vaccine 2011, 29, 2461–2473. [Google Scholar] [CrossRef]
- Jackson, L.A.; Campbell, J.D.; Frey, S.E.; Edwards, K.M.; Keitel, W.A.; Kotloff, K.L.; Berry, A.A.; Graham, I.; Atmar, R.L.; Creech, C.B.; et al. Effect of varying doses of a monovalent h7n9 influenza vaccine with and without as03 and mf59 adjuvants on immune response: A randomized clinical trial. JAMA 2015, 314, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillet, S.; Arunachalam, P.S.; Andreani, G.; Golden, N.; Fontenot, J.; Aye, P.P.; Röltgen, K.; Lehmicke, G.; Gobeil, P.; Dubé, C. Safety, immunogenicity, and protection provided by unadjuvanted and adjuvanted formulations of a recombinant plant-derived virus-like particle vaccine candidate for COVID-19 in nonhuman primates. Cell. Mol. Immunol. 2022, 19, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, P.S.; Walls, A.C.; Golden, N.; Atyeo, C.; Fischinger, S.; Li, C.; Aye, P.; Navarro, M.J.; Lai, L.; Edara, V.V. Adjuvanting a subunit COVID-19 vaccine to induce protective immunity. Nature 2021, 594, 253–258. [Google Scholar] [CrossRef]
- Shah, R.R.; Taccone, M.; Monaci, E.; Brito, L.A.; Bonci, A.; O’Hagan, D.T.; Amiji, M.M.; Seubert, A. The droplet size of emulsion adjuvants has significant impact on their potency, due to differences in immune cell-recruitment and-activation. Sci. Rep. 2019, 9, 11520. [Google Scholar] [CrossRef] [Green Version]
- O’Hagan, D.T.; Friedland, L.R.; Hanon, E.; Didierlaurent, A.M. Towards an evidence based approach for the development of adjuvanted vaccines. Curr. Opin. Immunol. 2017, 47, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Garcon, N.; Vaughn, D.W.; Didierlaurent, A.M. Development and evaluation of AS03, an Adjuvant System containing alpha-tocopherol and squalene in an oil-in-water emulsion. Expert. Rev. Vaccines 2012, 11, 349–366. [Google Scholar] [CrossRef]
- Graaf, H.d.; Faust, S.N. Fluarix quadrivalent vaccine for influenza. Expert. Rev. Vaccines 2015, 14, 1055–1063. [Google Scholar] [CrossRef]
- Lidgate, D.M.; Trattner, T.; Shultz, R.M.; Maskiewicz, R. Sterile Filtration of a Parenteral Emulsion. Pharm. Res. 1992, 9, 860–863. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.B. Squalene emulsions for parenteral vaccine and drug delivery. Molecules 2009, 14, 3286–3312. [Google Scholar] [CrossRef] [PubMed]
- Kolhe, P.; Shah, M.; Rathore, N. Sterile Product Development: Formulation, Process, Quality and Regulatory Considerations; Springer: Berlin/Heidelberg, Germany, 2013; Volume 6. [Google Scholar]
- Howard, L.M.; Hoek, K.L.; Goll, J.B.; Samir, P.; Galassie, A.; Allos, T.M.; Niu, X.; Gordy, L.E.; Creech, C.B.; Prasad, N.; et al. Cell-Based Systems Biology Analysis of Human AS03-Adjuvanted H5N1 Avian Influenza Vaccine Responses: A Phase I Randomized Controlled Trial. PLoS ONE 2017, 12, e0167488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esser, M.T.; Marchese, R.D.; Kierstead, L.S.; Tussey, L.G.; Wang, F.; Chirmule, N.; Washabaugh, M.W. Memory T cells and vaccines. Vaccine 2003, 21, 419–430. [Google Scholar] [CrossRef]
- Sallusto, F.; Lanzavecchia, A.; Araki, K.; Ahmed, R. From Vaccines to Memory and Back. Immunity 2010, 33, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, S.L.; Shaverdian, N.; Goto, Y.; Duthie, M.S.; Raman, V.S.; Evers, T.; Mompoint, F.; Vedvick, T.S.; Bertholet, S.; Coler, R.N.; et al. Enhanced humoral and Type 1 cellular immune responses with Fluzone® adjuvanted with a synthetic TLR4 agonist formulated in an emulsion. Vaccine 2009, 27, 5956–5963. [Google Scholar] [CrossRef]
- Baudner, B.C.; Ronconi, V.; Casini, D.; Tortoli, M.; Kazzaz, J.; Singh, M.; Hawkins, L.D.; Wack, A.; O’Hagan, D.T. MF59 Emulsion Is an Effective Delivery System for a Synthetic TLR4 Agonist (E6020). Pharm. Res. 2009, 26, 1477–1485. [Google Scholar] [CrossRef]
- Chandramouli, S.; Malito, E.; Nguyen, T.; Luisi, K.; Donnarumma, D.; Xing, Y.; Norais, N.; Yu, D.; Carfi, A. Structural basis for potent antibody-mediated neutralization of human cytomegalovirus. Sci. Immunol. 2017, 2, eaan1457. [Google Scholar] [CrossRef]
- Sridhar, S.; Joaquin, A.; Bonaparte, M.I.; Bueso, A.; Chabanon, A.-L.; Chen, A.; Chicz, R.M.; Diemert, D.; Essink, B.J.; Fu, B.; et al. Safety and immunogenicity of an AS03-adjuvanted SARS-CoV-2 recombinant protein vaccine (CoV2 preS dTM) in healthy adults: Interim findings from a phase 2, randomised, dose-finding, multicentre study. Lancet Infect. Dis. 2022, 22, 636–648. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SE-AS | Tween80 (%v/v) | Squalene (%v/v) | α-Tocopherol (%v/v) | Avg Size (nm) | PdI |
|---|---|---|---|---|---|
| 14 | 35 | 55 | 10 | 79.76 | 0.113 |
| 41 | 33 | 55 | 12 | 94.13 | 0.166 |
| 42 | 30 | 55 | 15 | 120.73 | 0.219 |
| 43 | 30 | 52 | 18 | 134.20 | 0.360 |
| 44 | 25 | 60 | 15 | 131.73 | 0.192 |
| 45 | 20 | 65 | 15 | 139.10 | 0.215 |
| 22 | 15 | 70 | 15 | 129.07 | 0.212 |
| AS03 | 16 | 42 | 42 | 155 | ˂0.2 |
| SE-AS | Before Filtration | After Filtration | % Content Loss | |||
|---|---|---|---|---|---|---|
| Size | PdI | Size | PdI | Squalene | α-Tocopherol | |
| 44 | 133.7 | 0.184 | 134.9 | 0.203 | 16.3 | 12.64 |
| 45 | 141.5 | 0.201 | 177.1667 | 0.329 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lodaya, R.N.; Kanitkar, A.P.; Ashraf, A.; Bamba, D.; Amiji, M.M.; O’Hagan, D.T. A Self-Emulsified Adjuvant System Containing the Immune Potentiator Alpha Tocopherol Induces Higher Neutralizing Antibody Responses than a Squalene-Only Emulsion When Evaluated with a Recombinant Cytomegalovirus (CMV) Pentamer Antigen in Mice. Pharmaceutics 2023, 15, 238. https://doi.org/10.3390/pharmaceutics15010238
Lodaya RN, Kanitkar AP, Ashraf A, Bamba D, Amiji MM, O’Hagan DT. A Self-Emulsified Adjuvant System Containing the Immune Potentiator Alpha Tocopherol Induces Higher Neutralizing Antibody Responses than a Squalene-Only Emulsion When Evaluated with a Recombinant Cytomegalovirus (CMV) Pentamer Antigen in Mice. Pharmaceutics. 2023; 15(1):238. https://doi.org/10.3390/pharmaceutics15010238
Chicago/Turabian StyleLodaya, Rushit N., Amey P. Kanitkar, Asma Ashraf, Douty Bamba, Mansoor M. Amiji, and Derek T. O’Hagan. 2023. "A Self-Emulsified Adjuvant System Containing the Immune Potentiator Alpha Tocopherol Induces Higher Neutralizing Antibody Responses than a Squalene-Only Emulsion When Evaluated with a Recombinant Cytomegalovirus (CMV) Pentamer Antigen in Mice" Pharmaceutics 15, no. 1: 238. https://doi.org/10.3390/pharmaceutics15010238