
Identification of Novel Loci Involved in Adalimumab Response in Crohn’s Disease Patients Using Integration of Genome Profiling and Isoform-Level Immune-Cell Deconvoluted Transcriptome Profiling of Colon Tissue

Abstract
:1. Introduction
- Perform isoform-level transcriptome profiling of colon tissue.
- Combine transcriptome profiling with refined information concerning immune cell landscapes in colon tissue.
- Stringently select promising gene candidates based on gene ontology analysis.
- Integrate and functionally annotate the findings from genome profiling data.
2. Materials and Methods
2.1. Enrolled Subjects
2.2. Extraction of Nucleic Acids
2.3. Transcript Specific Tissue RNA-Seq Analysis
2.4. Gene Ontology Analysis
2.5. Genome-Wide Association Analysis
2.6. Functional Annotation and eQTL Estimation
2.7. Construction of Gene Interaction Network and Visualization
2.8. Machine Learning Validation
3. Results
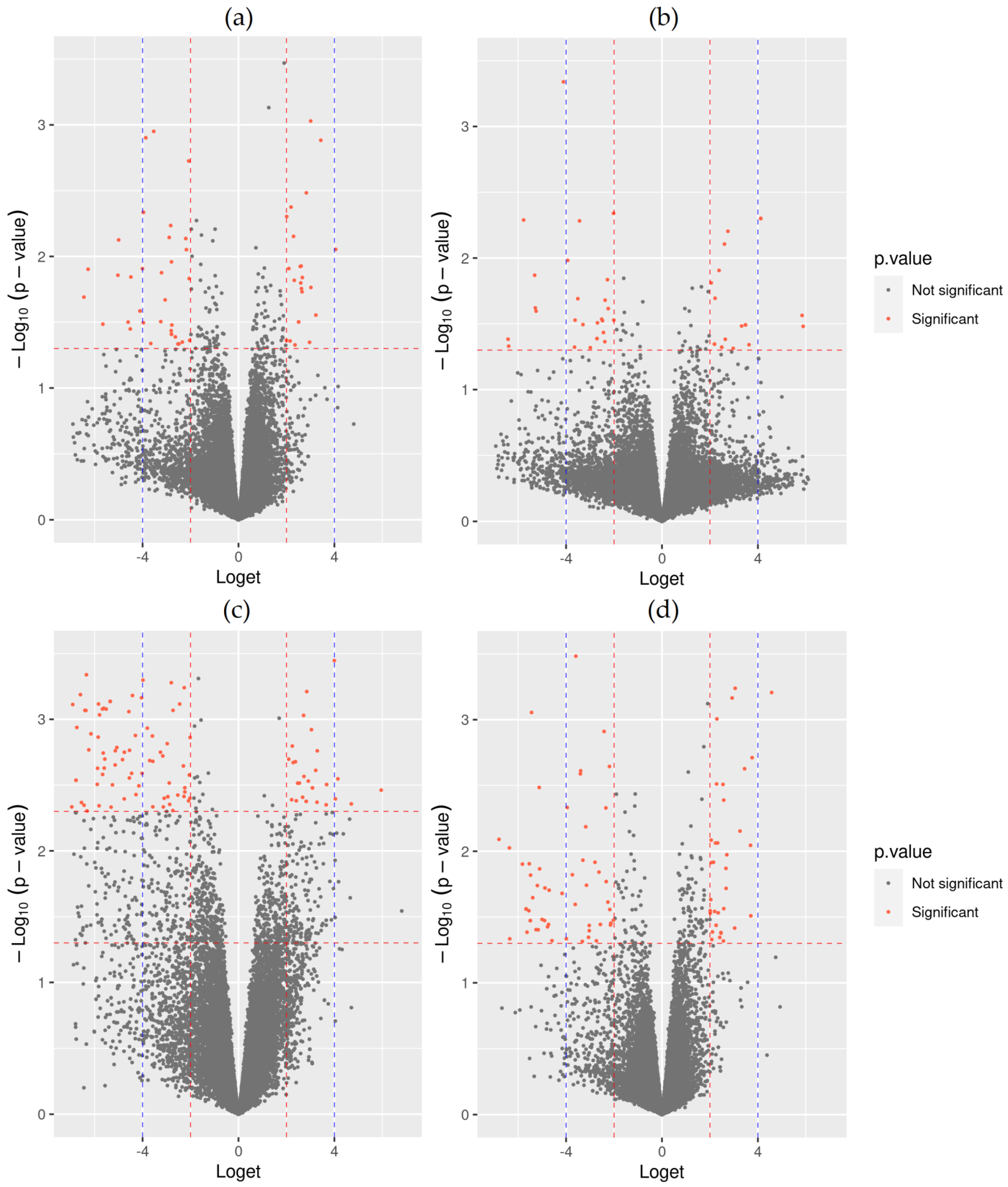
3.1. Transcript Specific RNA-Seq Analysis
3.2. Gene Ontology Enrichment Analysis
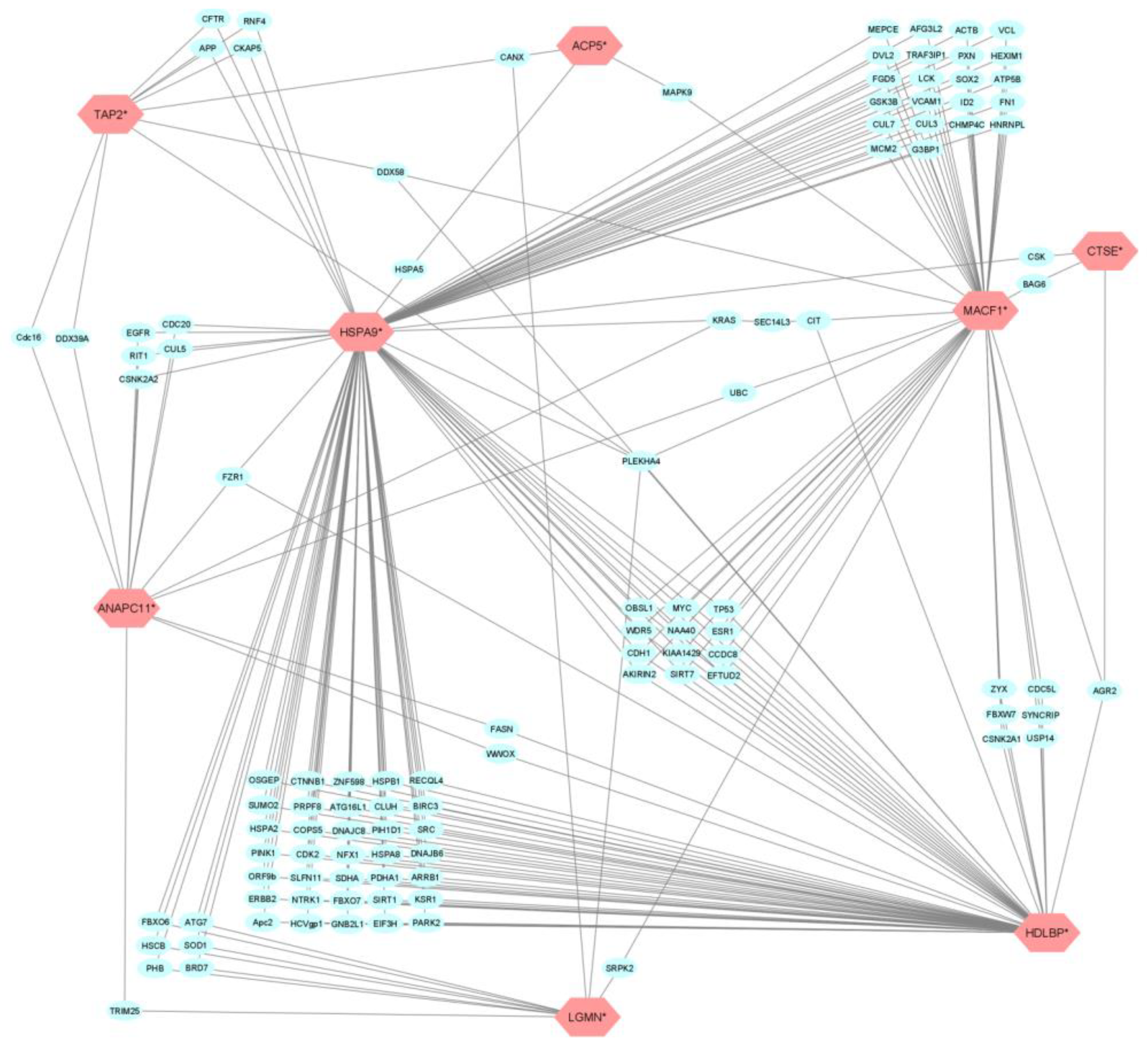
3.3. GWAs Integration, Gene Selection and Functional Annotation
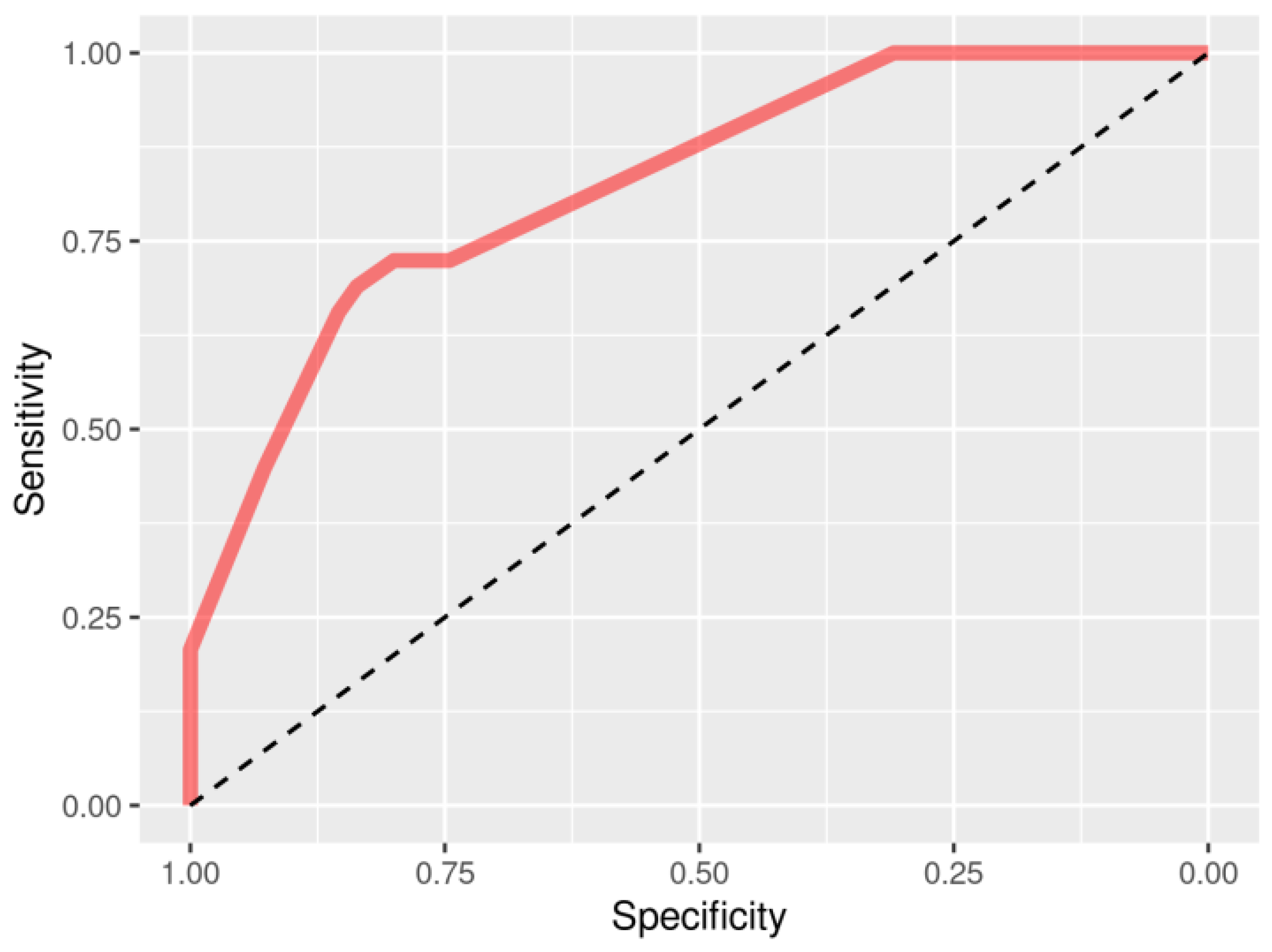
3.4. Interactions and Machine Learning Validation of Selected Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abraham, C.; Cho, J.H. Inflammatory bowel disease. N. Engl. J. Med. 2009, 361, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Ordas, I.; Eckmann, L.; Talamini, M.; Baumgart, D.C.; Sandborn, W.J. Ulcerative colitis. Lancet 2012, 380, 1606–1619. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Sandborn, W.J. Crohn’s disease. Lancet 2012, 380, 1590–1605. [Google Scholar] [CrossRef]
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012, 142, 46–54.e42. [Google Scholar] [CrossRef] [PubMed]
- Rosenstiel, P.; Sina, C.; Franke, A.; Schreiber, S. Towards a molecular risk map--recent advances on the etiology of inflammatory bowel disease. Semin. Immunol. 2009, 21, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Wehkamp, J.; Gotz, M.; Herrlinger, K.; Steurer, W.; Stange, E.F. Inflammatory Bowel Disease. Dtsch. Arztebl. Int. 2016, 113, 72–82. [Google Scholar] [CrossRef]
- Duchmann, R.; May, E.; Heike, M.; Knolle, P.; Neurath, M.; Meyer zum Buschenfelde, K.H. T cell specificity and cross reactivity towards enterobacteria, bacteroides, bifidobacterium, and antigens from resident intestinal flora in humans. Gut 1999, 44, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Moussata, D.; Goetz, M.; Gloeckner, A.; Kerner, M.; Campbell, B.; Hoffman, A.; Biesterfeld, S.; Flourie, B.; Saurin, J.C.; Galle, P.R.; et al. Confocal laser endomicroscopy is a new imaging modality for recognition of intramucosal bacteria in inflammatory bowel disease in vivo. Gut 2011, 60, 26–33. [Google Scholar] [CrossRef]
- Wehkamp, J.; Fellermann, K.; Herrlinger, K.R.; Bevins, C.L.; Stange, E.F. Mechanisms of disease: Defensins in gastrointestinal diseases. Nat. Clin. Pract. Gastroenterol. Hepatol. 2005, 2, 406–415. [Google Scholar] [CrossRef]
- Van Dussen, K.L.; Liu, T.C.; Li, D.; Towfic, F.; Modiano, N.; Winter, R.; Haritunians, T.; Taylor, K.D.; Dhall, D.; Targan, S.R.; et al. Genetic variants synthesize to produce paneth cell phenotypes that define subtypes of Crohn’s disease. Gastroenterology 2014, 146, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Zundler, S.; Neurath, M.F. Immunopathogenesis of inflammatory bowel diseases: Functional role of T cells and T cell homing. Clin. Exp. Rheumatol. 2015, 33, S19–S28. [Google Scholar] [PubMed]
- Danese, S.; Fiorino, G.; Reinisch, W. Review article: Causative factors and the clinical management of patients with Crohn’s disease who lose response to anti-TNF-alpha therapy. Aliment. Pharmacol. Ther. 2011, 34, 1–10. [Google Scholar] [CrossRef]
- Van Deventer, S.J. Tumour necrosis factor and Crohn’s disease. Gut 1997, 40, 443–448. [Google Scholar] [CrossRef]
- Colombel, J.F.; Sandborn, W.J.; Rutgeerts, P.; Enns, R.; Hanauer, S.B.; Panaccione, R.; Schreiber, S.; Byczkowski, D.; Li, J.; Kent, J.D.; et al. Adalimumab for maintenance of clinical response and remission in patients with Crohn’s disease: The CHARM trial. Gastroenterology 2007, 132, 52–65. [Google Scholar] [CrossRef]
- Hanauer, S.B.; Feagan, B.G.; Lichtenstein, G.R.; Mayer, L.F.; Schreiber, S.; Colombel, J.F.; Rachmilewitz, D.; Wolf, D.C.; Olson, A.; Bao, W.; et al. Maintenance infliximab for Crohn’s disease: The ACCENT I randomised trial. Lancet 2002, 359, 1541–1549. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Hanauer, S.B.; Rutgeerts, P.; Fedorak, R.N.; Lukas, M.; MacIntosh, D.G.; Panaccione, R.; Wolf, D.; Kent, J.D.; Bittle, B.; et al. Adalimumab for maintenance treatment of Crohn’s disease: Results of the CLASSIC II trial. Gut 2007, 56, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Sandborn, W.J.; Feagan, B.G.; Rutgeerts, P.; Hanauer, S.; Colombel, J.F.; Sands, B.E.; Lukas, M.; Fedorak, R.N.; Lee, S.; Bressler, B.; et al. Vedolizumab as induction and maintenance therapy for Crohn’s disease. N. Engl. J. Med. 2013, 369, 711–721. [Google Scholar] [CrossRef]
- Schoultz, I.; Keita, A.V. Cellular and Molecular Therapeutic Targets in Inflammatory Bowel Disease-Focusing on Intestinal Barrier Function. Cells 2019, 8, 193. [Google Scholar] [CrossRef]
- Feagan, B.G.; Sandborn, W.J.; Gasink, C.; Jacobstein, D.; Lang, Y.; Friedman, J.R.; Blank, M.A.; Johanns, J.; Gao, L.L.; Miao, Y.; et al. Ustekinumab as Induction and Maintenance Therapy for Crohn’s Disease. N. Engl. J. Med. 2016, 375, 1946–1960. [Google Scholar] [CrossRef]
- Feagan, B.G.; Panes, J.; Ferrante, M.; Kaser, A.; D’Haens, G.R.; Sandborn, W.J.; Louis, E.; Neurath, M.F.; Franchimont, D.; Dewit, O.; et al. Risankizumab in patients with moderate to severe Crohn’s disease: An open-label extension study. Lancet Gastroenterol. Hepatol. 2018, 3, 671–680. [Google Scholar] [CrossRef]
- Sands, B.E.; Peyrin-Biroulet, L.; Loftus, E.V., Jr.; Danese, S.; Colombel, J.F.; Toruner, M.; Jonaitis, L.; Abhyankar, B.; Chen, J.; Rogers, R.; et al. Vedolizumab versus Adalimumab for Moderate-to-Severe Ulcerative Colitis. N. Engl. J. Med. 2019, 381, 1215–1226. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Murad, M.H.; Fumery, M.; Sedano, R.; Jairath, V.; Panaccione, R.; Sandborn, W.J.; Ma, C. Comparative efficacy and safety of biologic therapies for moderate-to-severe Crohn’s disease: A systematic review and network meta-analysis. Lancet Gastroenterol. Hepatol. 2021, 6, 1002–1014. [Google Scholar] [CrossRef]
- Dezelak, M.; Repnik, K.; Koder, S.; Ferkolj, I.; Potocnik, U. A Prospective Pharmacogenomic Study of Crohn’s Disease Patients during Routine Therapy with Anti-TNF-alpha Drug Adalimumab: Contribution of ATG5, NFKB1, and CRP Genes to Pharmacodynamic Variability. OMICS A J. Integr. Biol. 2016, 20, 296–309. [Google Scholar] [CrossRef]
- Koder, S.; Repnik, K.; Ferkolj, I.; Pernat, C.; Skok, P.; Weersma, R.K.; Potocnik, U. Genetic polymorphism in ATG16L1 gene influences the response to adalimumab in Crohn’s disease patients. Pharmacogenomics 2015, 16, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Repnik, K.; Koder, S.; Skok, P.; Ferkolj, I.; Potocnik, U. Transferrin Level Before Treatment and Genetic Polymorphism in HFE Gene as Predictive Markers for Response to Adalimumab in Crohn’s Disease Patients. Biochem. Genet. 2016, 54, 476–486. [Google Scholar] [CrossRef]
- Barber, G.E.; Yajnik, V.; Khalili, H.; Giallourakis, C.; Garber, J.; Xavier, R.; Ananthakrishnan, A.N. Genetic Markers Predict Primary Non-Response and Durable Response To Anti-TNF Biologic Therapies in Crohn’s Disease. Am. J. Gastroenterol. 2016, 111, 1816–1822. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.M.; Haritunians, T.; Chhina, S.; Liu, Z.; Yang, S.; Landers, C.; Li, D.; Ye, B.D.; Shih, D.; Vasiliauskas, E.A.; et al. Colonic Phenotypes Are Associated with Poorer Response to Anti-TNF Therapies in Patients with IBD. Inflamm. Bowel Dis. 2017, 23, 1382–1393. [Google Scholar] [CrossRef] [PubMed]
- Arijs, I.; De Hertogh, G.; Lemaire, K.; Quintens, R.; Van Lommel, L.; Van Steen, K.; Leemans, P.; Cleynen, I.; Van Assche, G.; Vermeire, S.; et al. Mucosal gene expression of antimicrobial peptides in inflammatory bowel disease before and after first infliximab treatment. PLoS ONE 2009, 4, e7984. [Google Scholar] [CrossRef]
- Gorenjak, M.; Repnik, K.; Jezernik, G.; Jurgec, S.; Skok, P.; Potocnik, U. Genetic prediction profile for adalimumab response in Slovenian Crohn’s disease patients. Z. Gastroenterol. 2019, 57, 1218–1225. [Google Scholar] [CrossRef]
- Gorenjak, M.; Zupin, M.; Jezernik, G.; Skok, P.; Potocnik, U. Omics data integration identifies ELOVL7 and MMD gene regions as novel loci for adalimumab response in patients with Crohn’s disease. Sci. Rep. 2021, 11, 5449. [Google Scholar] [CrossRef]
- Schmitt, H.; Billmeier, U.; Dieterich, W.; Rath, T.; Sonnewald, S.; Reid, S.; Hirschmann, S.; Hildner, K.; Waldner, M.J.; Mudter, J.; et al. Expansion of IL-23 receptor bearing TNFR2+ T cells is associated with molecular resistance to anti-TNF therapy in Crohn’s disease. Gut 2019, 68, 814–828. [Google Scholar] [CrossRef]
- Martin, J.C.; Chang, C.; Boschetti, G.; Ungaro, R.; Giri, M.; Grout, J.A.; Gettler, K.; Chuang, L.S.; Nayar, S.; Greenstein, A.J.; et al. Single-Cell Analysis of Crohn’s Disease Lesions Identifies a Pathogenic Cellular Module Associated with Resistance to Anti-TNF Therapy. Cell 2019, 178, 1493–1508.e1420. [Google Scholar] [CrossRef]
- Gole, B.; Potocnik, U. Pre-Treatment Biomarkers of Anti-Tumour Necrosis Factor Therapy Response in Crohn’s Disease-A Systematic Review and Gene Ontology Analysis. Cells 2019, 8, 515. [Google Scholar] [CrossRef]
- Bai, X.; Liu, W.; Chen, H.; Zuo, T.; Wu, X. Immune Cell Landscaping Reveals Distinct Immune Signatures of Inflammatory Bowel Disease. Front. Immunol. 2022, 13, 861790. [Google Scholar] [CrossRef] [PubMed]
- Maaser, C.; Sturm, A.; Vavricka, S.R.; Kucharzik, T.; Fiorino, G.; Annese, V.; Calabrese, E.; Baumgart, D.C.; Bettenworth, D.; Nunes, P.B.; et al. ECCO-ESGAR Guideline for Diagnostic Assessment in IBD Part 1: Initial diagnosis, monitoring of known IBD, detection of complications. J. Crohns Colitis 2019, 13, 144–164. [Google Scholar] [CrossRef] [PubMed]
- Panaccione, R.; Loftus, E.V., Jr.; Binion, D.; McHugh, K.; Alam, S.; Chen, N.; Guerette, B.; Mulani, P.; Chao, J. Efficacy and safety of adalimumab in Canadian patients with moderate to severe Crohn’s disease: Results of the Adalimumab in Canadian SubjeCts with ModErate to Severe Crohn’s DiseaSe (ACCESS) trial. Can. J. Gastroenterol. 2011, 25, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Hlavaty, T.; Persoons, P.; Vermeire, S.; Ferrante, M.; Pierik, M.; Van Assche, G.; Rutgeerts, P. Evaluation of short-term responsiveness and cutoff values of inflammatory bowel disease questionnaire in Crohn’s disease. Inflamm. Bowel Dis. 2006, 12, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Guyatt, G.; Mitchell, A.; Irvine, E.J.; Singer, J.; Williams, N.; Goodacre, R.; Tompkins, C. A new measure of health status for clinical trials in inflammatory bowel disease. Gastroenterology 1989, 96, 804–810. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 29 April 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Hernandez-Pacheco, N.; Gorenjak, M.; Jurgec, S.; Corrales, A.; Jorgensen, A.; Karimi, L.; Vijverberg, S.J.; Berce, V.; Schieck, M.; Acosta-Herrera, M.; et al. Combined analysis of transcriptomic and genetic data for the identification of loci involved in glucocorticosteroid response in asthma. Allergy 2021, 76, 1238–1243. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.M.; Liu, C.L.; Green, M.R.; Gentles, A.J.; Feng, W.; Xu, Y.; Hoang, C.D.; Diehn, M.; Alizadeh, A.A. Robust enumeration of cell subsets from tissue expression profiles. Nat. Methods 2015, 12, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Prada, C.; Lima, D.; Nakaya, H. MetaVolcanoR: Gene Expression Meta-Analysis Visualization Tool. 2020. Available online: https://www.bioconductor.org/packages/release/bioc/html/MetaVolcanoR.html (accessed on 29 April 2022).
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Machiela, M.J.; Chanock, S.J. LDlink: A web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants. Bioinformatics 2015, 31, 3555–3557. [Google Scholar] [CrossRef] [PubMed]
- Pruim, R.J.; Welch, R.P.; Sanna, S.; Teslovich, T.M.; Chines, P.S.; Gliedt, T.P.; Boehnke, M.; Abecasis, G.R.; Willer, C.J. LocusZoom: Regional visualization of genome-wide association scan results. Bioinformatics 2010, 26, 2336–2337. [Google Scholar] [CrossRef]
- Ward, L.D.; Kellis, M. HaploReg: A resource for exploring chromatin states, conservation, and regulatory motif alterations within sets of genetically linked variants. Nucleic Acids Res. 2012, 40, D930–D934. [Google Scholar] [CrossRef]
- GTEx Consortium. Erratum: Genetic effects on gene expression across human tissues. Nature 2018, 553, 530. [Google Scholar] [CrossRef]
- Hunt, S.E.; McLaren, W.; Gil, L.; Thormann, A.; Schuilenburg, H.; Sheppard, D.; Parton, A.; Armean, I.M.; Trevanion, S.J.; Flicek, P.; et al. Ensembl variation resources. Database 2018, 2018, 119. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Oughtred, R.; Stark, C.; Breitkreutz, B.J.; Rust, J.; Boucher, L.; Chang, C.; Kolas, N.; O‘Donnell, L.; Leung, G.; McAdam, R.; et al. The BioGRID interaction database: 2019 update. Nucleic Acids Res. 2019, 47, D529–D541. [Google Scholar] [CrossRef] [PubMed]
- Stark, C.; Breitkreutz, B.J.; Reguly, T.; Boucher, L.; Breitkreutz, A.; Tyers, M. BioGRID: A general repository for interaction datasets. Nucleic Acids Res. 2006, 34, D535–D539. [Google Scholar] [CrossRef] [PubMed]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R. News 2002, 2, 18–22. [Google Scholar]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Muller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef]
- Suozzi, K.C.; Wu, X.; Fuchs, E. Spectraplakins: Master orchestrators of cytoskeletal dynamics. J. Cell Biol. 2012, 197, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yue, J.; Zhang, Y.; Shi, C.; Odenwald, M.; Liang, W.G.; Wei, Q.; Goel, A.; Gou, X.; Zhang, J.; et al. ACF7 regulates inflammatory colitis and intestinal wound response by orchestrating tight junction dynamics. Nat. Commun. 2017, 8, 15375. [Google Scholar] [CrossRef] [PubMed]
- Clark, P.M.; Dawany, N.; Dampier, W.; Byers, S.W.; Pestell, R.G.; Tozeren, A. Bioinformatics analysis reveals transcriptome and microRNA signatures and drug repositioning targets for IBD and other autoimmune diseases. Inflamm. Bowel Dis. 2012, 18, 2315–2333. [Google Scholar] [CrossRef]
- Zimmermann, P.; Baumann, R. Genes Specifically Regulated in Inflammatory Bowel Disease; Nebion: Zürich, Switzerland, 2015. [Google Scholar]
- Ehrhardt, K.; Steck, N.; Kappelhoff, R.; Stein, S.; Rieder, F.; Gordon, I.O.; Boyle, E.C.; Braubach, P.; Overall, C.M.; Finlay, B.B.; et al. Persistent Salmonella enterica Serovar Typhimurium Infection Induces Protease Expression During Intestinal Fibrosis. Inflamm. Bowel Dis. 2019, 25, 1629–1643. [Google Scholar] [CrossRef]
- Zinnall, U.; Milek, M.; Minia, I.; Vieira-Vieira, C.H.; Muller, S.; Mastrobuoni, G.; Hazapis, O.G.; Del Giudice, S.; Schwefel, D.; Bley, N.; et al. HDLBP binds ER-targeted mRNAs by multivalent interactions to promote protein synthesis of transmembrane and secreted proteins. Nat. Commun. 2022, 13, 2727. [Google Scholar] [CrossRef]
- Kosmas, K.; Filippakis, H.; Khabibullin, D.; Turkiewicz, M.; Lam, H.C.; Yu, J.; Kedersha, N.L.; Anderson, P.J.; Henske, E.P. TSC2 Interacts with HDLBP/Vigilin and Regulates Stress Granule Formation. Mol. Cancer Res. 2021, 19, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Gundamaraju, R.; Vemuri, R.; Angelucci, C.; Geraghty, D.; Gueven, N.; Eri, R. Identification of Key Pro-Survival Proteins in Isolated Colonic Goblet Cells of Winnie, a Murine Model of Spontaneous Colitis. Inflamm. Bowel Dis. 2020, 26, 80–92. [Google Scholar] [CrossRef]
- Loren, V.; Garcia-Jaraquemada, A.; Naves, J.E.; Carmona, X.; Manosa, M.; Aransay, A.M.; Lavin, J.L.; Sanchez, I.; Cabre, E.; Manye, J.; et al. ANP32E, a Protein Involved in Steroid-Refractoriness in Ulcerative Colitis, Identified by a Systems Biology Approach. J. Crohns Colitis 2019, 13, 351–361. [Google Scholar] [CrossRef]
- Yang, L.; Tang, S.; Baker, S.S.; Arijs, I.; Liu, W.; Alkhouri, R.; Lan, P.; Baker, R.D.; Tang, Z.; Ji, G.; et al. Difference in Pathomechanism Between Crohn’s Disease and Ulcerative Colitis Revealed by Colon Transcriptome. Inflamm. Bowel Dis. 2019, 25, 722–731. [Google Scholar] [CrossRef]
- Goyette, P.; Boucher, G.; Mallon, D.; Ellinghaus, E.; Jostins, L.; Huang, H.; Ripke, S.; Gusareva, E.S.; Annese, V.; Hauser, S.L.; et al. High-density mapping of the MHC identifies a shared role for HLA-DRB1*01:03 in inflammatory bowel diseases and heterozygous advantage in ulcerative colitis. Nat. Genet. 2015, 47, 172–179. [Google Scholar] [CrossRef]
- Heresbach, D.; Alizadeh, M.; Bretagne, J.F.; Dabadie, A.; Colombel, J.F.; Pagenault, M.; Heresbach-Le Berre, N.; Genetet, B.; Gosselin, M.; Semana, G. TAP gene transporter polymorphism in inflammatory bowel diseases. Scand. J. Gastroenterol. 1997, 32, 1022–1027. [Google Scholar] [CrossRef]
- Burakoff, R.; Pabby, V.; Onyewadume, L.; Odze, R.; Adackapara, C.; Wang, W.; Friedman, S.; Hamilton, M.; Korzenik, J.; Levine, J.; et al. Blood-based biomarkers used to predict disease activity in Crohn’s disease and ulcerative colitis. Inflamm. Bowel Dis. 2015, 21, 1132–1140. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Liang, X.; Wen, Y.; Li, P.; Zhang, L.; Ma, M.; Cheng, S.; Du, Y.; Liu, L.; Ding, M.; et al. Integrative analysis of transcriptome-wide association study data and messenger RNA expression profiles identified candidate genes and pathways for inflammatory bowel disease. J. Cell Biochem. 2019, 120, 14831–14837. [Google Scholar] [CrossRef]
- Taman, H.; Fenton, C.G.; Hensel, I.V.; Anderssen, E.; Florholmen, J.; Paulssen, R.H. Transcriptomic Landscape of Treatment-Naive Ulcerative Colitis. J. Crohns Colitis 2018, 12, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Anyona, S.B.; Cheng, Q.; Raballah, E.; Hurwitz, I.; Lambert, C.G.; McMahon, B.H.; Ouma, C.; Perkins, D.J. Ingestion of hemozoin by peripheral blood mononuclear cells alters temporal gene expression of ubiquitination processes. Biochem. Biophys. Rep. 2022, 29, 101207. [Google Scholar] [CrossRef] [PubMed]
- Drouet, Y.; Treilleux, I.; Viari, A.; Leon, S.; Devouassoux-Shisheboran, M.; Voirin, N.; de la Fouchardiere, C.; Manship, B.; Puisieux, A.; Lasset, C.; et al. Integrated analysis highlights APC11 protein expression as a likely new independent predictive marker for colorectal cancer. Sci. Rep. 2018, 8, 7386. [Google Scholar] [CrossRef]
- Turner, E.L.; Malo, M.E.; Pisclevich, M.G.; Dash, M.D.; Davies, G.F.; Arnason, T.G.; Harkness, T.A. The Saccharomyces cerevisiae anaphase-promoting complex interacts with multiple histone-modifying enzymes to regulate cell cycle progression. Eukaryot. Cell 2010, 9, 1418–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Approach | GO ID | Description | q Value |
|---|---|---|---|
| Biological Process | |||
| I consistent * | GO:0002474 | Antigen processing and presentation of peptide antigen via MHC class I | 9.27 × 10−6 |
| I opposite ** | NA | NA | NA |
| II consistent | GO:0048002 | Antigen processing and presentation of peptide antigen | 0.00018 |
| II opposite | NA | NA | NA |
| III consistent | GO:0002181 | Cytoplasmic translation | 4.16 × 10−17 |
| III opposite | GO:0019885 | Antigen processing and presentation of endogenous peptide antigen via MHC class I | 0.00014 |
| IV consistent | GO:0019885 | Antigen processing and presentation of endogenous peptide antigen via MHC class I | 1.13 × 10−05 |
| IV opposite | GO:0039531 | Regulation of viral-induced cytoplasmic pattern recognition receptor signaling pathway | 0.0019 |
| Cellular Component | |||
| I consistent | GO:0071556 | Integral component of lumenal side of endoplasmic reticulum membrane | 2.91 × 10−05 |
| I opposite | GO:0030139 | Endocytic vesicle | 0.0057 |
| II consistent | GO:0042611 | MHC protein complex | 0.0015 |
| II opposite | NA | NA | NA |
| III consistent | GO:0022626 | Cytosolic ribosome | 2.15 × 10−16 |
| III opposite | GO:0071556 | Integral component of lumenal side of endoplasmic reticulum membrane | 3.25 × 10−05 |
| IV consistent | GO:0071556 | Integral component of lumenal side of endoplasmic reticulum membrane | 0.00049 |
| IV opposite | NA | NA | NA |
| Molecular Function | |||
| I consistent | GO:0003823 | Antigen binding | 4.39 × 10−08 |
| I opposite | NA | NA | NA |
| II consistent | NA | NA | NA |
| II opposite | NA | NA | NA |
| III consistent | GO:0003735 | Structural constituent of ribosome | 2.24 × 10−13 |
| III opposite | GO:0042605 | Peptide antigen binding | 9.90 × 10−05 |
| IV consistent | GO:0042605 | Peptide antigen binding | 9.83 × 10−05 |
| IV opposite | GO:0045296 | Cadherin binding | 3.44 × 10−05 |
| LOCUS | dbSNP | CHR | BP | p Value | q Value | β | SE | AF.RESP | AF.NON |
|---|---|---|---|---|---|---|---|---|---|
| MACF1 | rs10888633 | 1 | 39487164 | 0.0081 | 0.0398 | 1.0167 | 0.38416 | 0.44545 | 0.22414 |
| MACF1 | rs35441329 | 1 | 39537291 | 0.0030 | 0.0148 | −1.4006 | 0.47237 | 0.12727 | 0.31034 |
| CTSE | rs28396806 | 1 | 206256869 | 0.0060 | 0.0360 | −1.3133 | 0.47774 | 0.11818 | 0.31034 |
| HDLBP | rs75590598 | 2 | 242185654 | 0.0002 | 0.0496 | −2.3973 | 0.65155 | 0.054545 | 0.24138 |
| HSPA9 | rs6891007 | 5 | 137966643 | 0.0054 | 0.0294 | −1.0835 | 0.38979 | 0.25455 | 0.46552 |
| HSPA9 | rs28386947 | 5 | 137978163 | 0.0021 | 0.0115 | −1.4205 | 0.46251 | 0.18182 | 0.41379 |
| HSPA9 | rs58914268 | 5 | 137987814 | 0.0057 | 0.0306 | −1.0578 | 0.38237 | 0.19091 | 0.39655 |
| HLA-DMB | rs7761882 | 6 | 32859137 | 0.0019 | 0.0443 | −1.7556 | 0.56605 | 0.1 | 0.2931 |
| TAP2 * | rs7761882 | 6 | 32859137 | 0.0019 | 0.0297 | −1.7556 | 0.56605 | 0.1 | 0.2931 |
| LGMN | rs58531216 | 14 | 93071325 | 0.0017 | 0.0158 | −1.6627 | 0.52838 | 0.072727 | 0.25862 |
| LGMN | rs10132165 | 14 | 93091829 | 0.0036 | 0.0347 | 1.4152 | 0.48635 | 0.88182 | 0.7069 |
| LGMN | rs11621843 | 14 | 93116124 | 0.0007 | 0.0071 | −1.9507 | 0.57841 | 0.054545 | 0.25862 |
| LGMN | rs3814830 | 14 | 93118198 | 0.0042 | 0.0399 | −1.3242 | 0.46212 | 0.1 | 0.2931 |
| LGMN | rs10137934 | 14 | 93193912 | 0.0035 | 0.0339 | 2.3774 | 0.81496 | 0.22727 | 0.034483 |
| ANAPC11 | rs144948822 | 17 | 79766520 | 0.0068 | 0.0136 | −3.2032 | 1.1837 | 0.0090909 | 0.12069 |
| ANAPC11 | rs28454947 | 17 | 79769466 | 0.0214 | 0.0428 | −1.1223 | 0.48779 | 0.10909 | 0.24138 |
| ACP5 | rs2229531 | 19 | 11687195 | 0.0011 | 0.0392 | −1.96 | 0.60067 | 0.081818 | 0.24138 |
| GENE | TRANSCRIPT | dbSNP | eQTL p Value ALL | eQTL p Value H | eQTL p Value D |
|---|---|---|---|---|---|
| MACF1 | ENST00000372925.2 | rs10888633 | 0.084 | 0.485 | 0.099 |
| MACF1 | ENST00000372925.2 | rs35441329 | 1 | 0.134 | 0.119 |
| CTSE | ENST00000360218.2 | rs28396806 | 0.315 | 0.439 | 0.142 |
| CTSE | ENST00000581049.1 | rs28396806 | 0.315 | 0.439 | 0.142 |
| HDLBP | ENST00000310931.4 | rs75590598 | 0.115 | 1 | 0.197 |
| HSPA9 | ENST00000501917.2 | rs6891007 | 0.805 | 0.319 | 0.887 |
| HSPA9 | ENST00000501917.2 | rs28386947 | 0.065 | 0.102 | 0.331 |
| HSPA9 | ENST00000501917.2 | rs58914268 | 0.422 | 0.077 | 0.187 |
| HLA-DMB | ENST00000383231.2 | rs7761882 | 0.462 | 0.299 | 0.339 |
| HLA-DMB | ENST00000428420.2 | rs7761882 | 0.462 | 0.299 | 0.339 |
| HLA-DMB | ENST00000440078.2 | rs7761882 | 0.462 | 0.299 | 0.339 |
| TAP2 | ENST00000443713.2 | rs7761882 | 0.068 | 0.235 | 0.218 |
| TAP2 | ENST00000452371.2 | rs7761882 | 0.181 | 0.325 | 0.441 |
| TAP2 | ENST00000455842.2 | rs7761882 | 0.181 | 0.325 | 0.441 |
| TAP2 | ENST00000457634.2 | rs7761882 | 0.083 | 0.325 | 0.339 |
| LGMN | ENST00000555169.1 | rs58531216 | 0.004 | 0.053 | 0.052 |
| LGMN | ENST00000555169.1 | rs10132165 | 0.345 | 0.134 | 0.769 |
| LGMN | ENST00000555169.1 | rs11621843 | 0.211 | 0.699 | 0.052 |
| LGMN | ENST00000555169.1 | rs3814830 | 0.355 | 0.699 | 0.065 |
| LGMN | ENST00000555169.1 | rs10137934 | 0.637 | 0.617 | 0.462 |
| ANAPC11 | ENST00000583839.1 | rs144948822 | 0.501 | 0.134 | 0.883 |
| ANAPC11 | ENST00000583839.1 | rs28454947 | 0.395 | 1 | 0.378 |
| ACP5 | ENST00000218758.5 | rs2229531 | 0.515 | 0.121 | 0.624 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorenjak, M.; Jezernik, G.; Krušič, M.; Skok, P.; Potočnik, U. Identification of Novel Loci Involved in Adalimumab Response in Crohn’s Disease Patients Using Integration of Genome Profiling and Isoform-Level Immune-Cell Deconvoluted Transcriptome Profiling of Colon Tissue. Pharmaceutics 2022, 14, 1893. https://doi.org/10.3390/pharmaceutics14091893
Gorenjak M, Jezernik G, Krušič M, Skok P, Potočnik U. Identification of Novel Loci Involved in Adalimumab Response in Crohn’s Disease Patients Using Integration of Genome Profiling and Isoform-Level Immune-Cell Deconvoluted Transcriptome Profiling of Colon Tissue. Pharmaceutics. 2022; 14(9):1893. https://doi.org/10.3390/pharmaceutics14091893
Chicago/Turabian StyleGorenjak, Mario, Gregor Jezernik, Martina Krušič, Pavel Skok, and Uroš Potočnik. 2022. "Identification of Novel Loci Involved in Adalimumab Response in Crohn’s Disease Patients Using Integration of Genome Profiling and Isoform-Level Immune-Cell Deconvoluted Transcriptome Profiling of Colon Tissue" Pharmaceutics 14, no. 9: 1893. https://doi.org/10.3390/pharmaceutics14091893