
In Vitro Biological Activity and In Vivo Human Study of Porcine-Placenta-Extract-Loaded Nanovesicle Formulations for Skin and Hair Rejuvenation

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Porcine Placenta Extract (PPE)
2.3. Quantification of Proteins and Growth Factors in the Extract
2.4. Determination of Cell Viability
2.5. Determination of Cell Proliferation
2.6. Determination of Cell Migration
2.7. Determination of Cell Aggregation of HFDPC
2.8. Determination of Intracellular ROS Reduction
2.9. Determination of Lipoxygenase (LOX) Inhibition
2.10. Intracellular TNF-α Determination
2.11. Preparation of PPE-Loaded Niosome Nanovesicles and Serum Formulations
2.12. Characterization of PPE-Loaded Niosome Nanovesicles
2.13. In Vivo Human Study
2.14. Statistical Analysis
3. Results
3.1. PPE Extract
3.2. Effect of PPE on Skin and Hair Cell Viability
3.3. Effect of PPE on Skin and Hair Cell Proliferation
3.4. Effect of PPE on Skin Cell Migration Activity
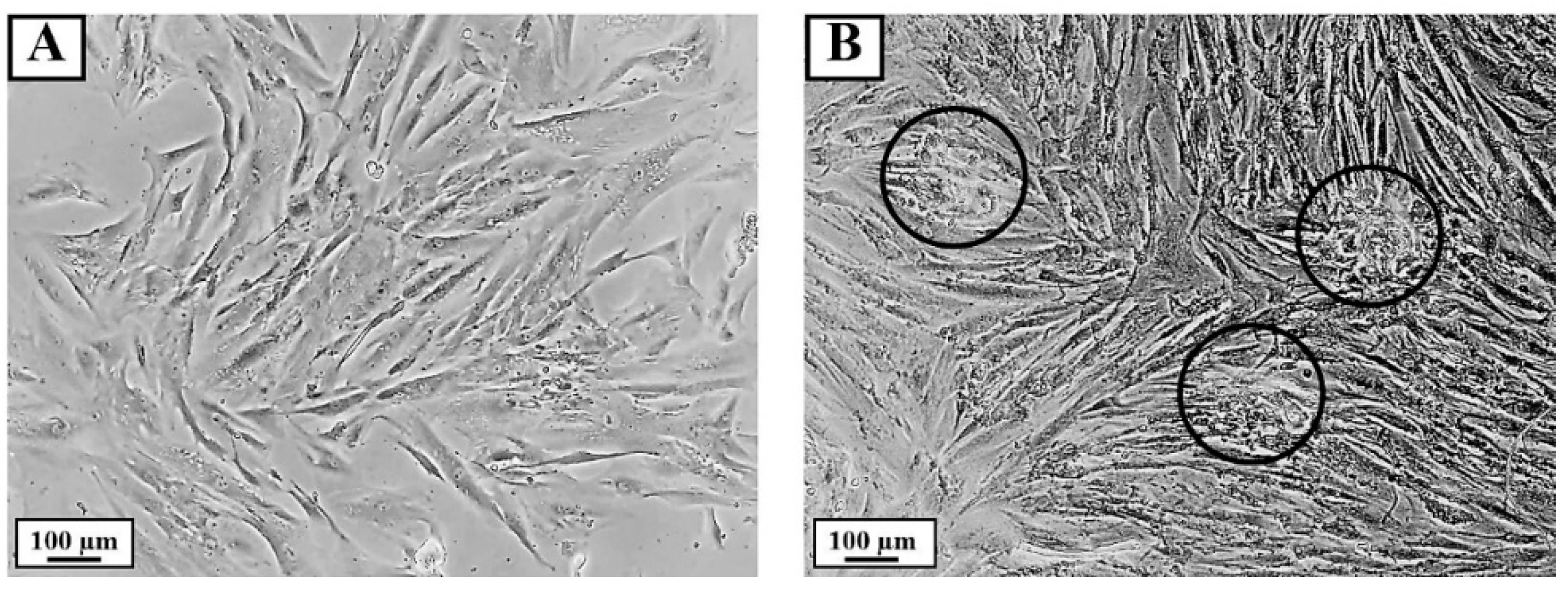
3.5. Effect of PPE on HFDPC Aggregation
3.6. Effect of PPE on Intracellular ROS Reduction
3.7. Effect of PPE on Lipoxygenase Inhibition
3.8. Effect of PPE on Intracellular TNF-α Reduction
3.9. Characterization of PPE-Loaded Niosomal Serum
3.10. In Vivo Human Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tobin, D.J. Introduction to skin aging. J. Tissue Viability 2017, 26, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Korkina, L.; Pastore, S. The role of redox regulation in normal physiology and inflammatory disease of skin. Front. Biosci. (Elite Ed.) 2009, 1, 123–141. [Google Scholar]
- Fuchs, J.; Zollner, T.M.; Kaufmann, R.; Podda, M. Redox-modulated pathways in inflammatory skin diseases. Free Radic. Biol. Med. 2001, 30, 337–353. [Google Scholar] [CrossRef]
- Hu, C.; Ma, S. Recent development of lipoxygenase inhibitors as anti-inflammatory agents. MedChemComm 2017, 9, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Kendall, A.C.; Nicolaou, A. Bioactive lipid mediators in skin inflammation and immunity. Prog. Lipid Res. 2013, 52, 141–164. [Google Scholar] [CrossRef]
- Geyfman, M.; Andersen, B. Clock genes, hair growth and aging. Aging 2010, 2, 122–128. [Google Scholar] [CrossRef]
- Tang, W.; Zhang, S.G.; Zhang, J.K.; Chen, S.; Zhu, H.; Ge, S.R. Ageing effects on the diameter, nanomechanical properties and tactile perception of human hair. Int. J. Cosmet. Sci. 2016, 38, 155–163. [Google Scholar] [CrossRef]
- Trüeb, R.M. Aging of hair. J. Cosmet. Dermatol. 2005, 4, 60–72. [Google Scholar] [CrossRef]
- Pan, S.Y.; Chan, M.; Wong, M.; Klokol, D.; Chernykh, V. Placental therapy: An insight to their biological and therapeutic properties. J. Med. Therap. 2017, 1, 12. [Google Scholar]
- Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Makrantonaki, E.; Zouboulis, C.C. Skin anti-aging strategies. Dermatoendocrinology 2012, 4, 308–319. [Google Scholar] [CrossRef]
- Seo, T.B.; Han, I.S.; Yoon, J.H.; Seol, I.C.; Kim, Y.S.; Jo, H.K.; An, J.J.; Hong, K.E.; Seo, Y.B.; Kim, D.H.; et al. Growth-promoting activity of hominis placenta extract on regenerating sciatic nerve. Acta Pharmacol. Sin. 2006, 27, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Schuetz, Y.B.; Naik, A.; Guy, R.H.; Kalia, Y.N. Emerging strategies for the transdermal delivery of peptide and protein drugs. Expert Opin. Drug Deliv. 2005, 2, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Agrahari, V.; Agrahari, V.; Mitra, A.K. Nanocarrier fabrication and macromolecule drug delivery: Challenges and opportunities. Ther. Deliv. 2016, 7, 257–278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Huilong, O.; Liu, C.; Zhang, Y.; Mitragotri, S.; Wang, D.; Chen, M. Skin delivery of hydrophilic biomacromolecules using marine sponge spicules. Mol. Pharm. 2017, 14, 3188–3200. [Google Scholar] [CrossRef]
- Tansathien, K.; Suriyaaumporn, P.; Charoenputtakhun, P.; Ngawhirunpat, T.; Opanasopit, P.; Rangsimawong, W. Development of sponge microspicule cream as a transdermal delivery system for protein and growth factors from deer antler velvet extract. Biol. Pharm. Bull. 2019, 42, 1207–1215. [Google Scholar] [CrossRef]
- Thiengpimol, P.; Tappreang, S.; Onarun, P. Reproductive performance of purebred and crossbred Landrace and Large White sows raised under Thai Commercial Swine Herd. Sci. Technol. Asia 2017, 22, 16–22. [Google Scholar]
- Wadkhien, K.; Chinpaisal, C.; Satiraphan, M.; Wetwitayaklung, P.; Pongnimitprasert, N. Anti-inflammatory effects of rhein and crude extracts from Cassia alata L. in HaCaT cells. Sci. Eng. Health Stud. 2018, 12, 19–32. [Google Scholar]
- Chung, L.Y.; Soo, W.K.; Chan, K.Y.; Mustafa, M.R.; Goh, S.H.; Imiyabir, Z. Lipoxygenase inhibiting activity of some Malaysian plants. Pharm. Biol. 2009, 47, 1142–1148. [Google Scholar] [CrossRef]
- Takehara, K. Growth regulation of skin fibroblasts. J. Dermatol. Sci. 2000, 24, S70–S77. [Google Scholar] [CrossRef]
- Schlessinger, J.; Schreiber, A.B.; Levi, A.; Lax, I.; Libermann, T.; Yarden, Y. Regulation of cell proliferation by epidermal growth factor. Crit. Rev. Biochem. Mol. Biol. 1983, 14, 93–111. [Google Scholar] [CrossRef]
- Anitua, E.; Sánchez, M.; Zalduendo, M.M.; de la Fuente, M.; Prado, R.; Orive, G.; Andía, I. Fibroblastic response to treatment with different preparations rich in growth factors. Cell Prolif. 2009, 42, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, P.; Braye, F.; Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Adv. Drug Deliv. Rev. 2019, 146, 344–365. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.W.; Kang, M.C.; Lee, K.S. TGF-β1-treated ADSCs-CM promotes expression of type I collagen and MMP-1, migration of human skin fibroblasts, and wound healing in vitro and in vivo. Int. J. Mol. Med. 2010, 26, 901–906. [Google Scholar] [PubMed]
- Bektas, A.; Schurman, S.H.; Sen, R.; Ferrucci, L. Aging, inflammation and the environment. Exp. Gerontol. 2018, 105, 10–18. [Google Scholar] [CrossRef]
- Dainichi, T.; Hanakawa, S.; Kabashima, K. Classification of inflammatory skin diseases: A proposal based on the disorders of the three-layered defense systems, barrier, innate immunity and acquired immunity. J. Dermatol. Sci. 2014, 76, 81–89. [Google Scholar] [CrossRef]
- Masaki, H. Role of antioxidants in the skin: Anti-aging effects. J. Dermatol. Sci. 2010, 58, 85–90. [Google Scholar] [CrossRef]
- Nègre-Salvayre, A.; Augé, N.; Duval, C.; Robbesyn, F.; Thiers, J.C.; Nazzal, D.; Benoist, H.; Salvayre, R. Detection of intracellular reactive oxygen species in cultured cells using fluorescent probes. Methods Enzymol. 2002, 352, 62–71. [Google Scholar]
- Czapski, G.A.; Czubowicz, K.; Strosznajder, R.P. Evaluation of the antioxidative properties of lipoxygenase inhibitors. Pharmacol. Rep. 2012, 64, 1179–1188. [Google Scholar] [CrossRef]
- Kim, S.N.; Akindehin, S.; Kwon, H.J.; Son, Y.H.; Saha, A.; Jung, Y.S.; Seong, J.K.; Lim, K.M.; Sung, J.H.; Maddipati, K.R.; et al. Anti-inflammatory role of 15-lipoxygenase contributes to the maintenance of skin integrity in mice. Sci. Rep. 2018, 8, 8856. [Google Scholar] [CrossRef]
- Binic, I.; Lazarevic, V.; Ljubenovic, M.; Mojsa, J.; Sokolovic, D. Skin ageing: Natural weapons and strategies. Evid. Based Complementary Altern. Med. 2013, 2013, 827248. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.B.; Je, J.Y.; Cho, Y.S. Antioxidant and anti-inflammatory peptide fraction from salmon byproduct protein hydrolysates by peptic hydrolysis. Food Res. Int. 2012, 49, 92–98. [Google Scholar] [CrossRef]
- Madaan, A.; Verma, R.; Singh, A.T.; Jaggi, M. Review of hair follicle dermal papilla cells as in vitro screening model for hair growth. Int. J. Cosmet. Sci. 2018, 40, 429–450. [Google Scholar] [CrossRef]
- Chi, W.; Wu, E.; Morgan, B.A. Dermal papilla cell number specifies hair size, shape and cycling and its reduction causes follicular decline. Development 2013, 140, 1676. [Google Scholar] [CrossRef] [PubMed]
- Sari, A.R.P.; Rufaut, N.W.; Jones, L.N.; Sinclair, R.D. Characterization of ovine dermal papilla cell aggregation. Int. J. Trichol. 2016, 8, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Azizi, W.; Azad, A.K.; Ahmad, N.A.; Sunzida, N.K. Clinical efficacy of aloe vera based products available in the market as skin moisturiser measured by tewl value and skin hydration level by using dermalab technology. Pharmacologyonline 2016, 2, 42–49. [Google Scholar]
- Plessis, J.D.; Stefaniak, A.; Eloff, F.; John, S.; Agner, T.; Chou, T.C.; Nixon, R.; Steiner, M.; Franken, A.; Kudla, I.; et al. International guidelines for the in vivo assessment of skin properties in non-clinical settings: Part 2. transepidermal water loss and skin hydration. Skin Res. Technol. 2013, 19, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Hadi, H.; Awadh, A.I.; Hanif, N.M.; Md Sidik, N.F.; Mohd Rani, M.R.; Suhaimi, M.S. The investigation of the skin biophysical measurements focusing on daily activities, skin care habits, and gender differences. Skin Res. Technol. 2016, 22, 247–254. [Google Scholar] [CrossRef]
- Khosrowpour, Z.; Ahmad Nasrollahi, S.; Ayatollahi, A.; Samadi, A.; Firooz, A. Effects of four soaps on skin trans-epidermal water loss and erythema index. J. Cosmet. Dermatol. 2019, 18, 857–861. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulations | Size (nm) | PDI | Zeta Potential (mV) |
|---|---|---|---|
| Niosomal solutions | 107.60 ± 0.20 | 0.274 ± 0.02 | −35.73 ± 1.50 |
| PPE-loaded niosomal solutions | 124.63 ± 2.02 ** | 0.352 ± 0.01 ** | −32.0 ± 2.02 |
| PPE-loaded niosomal serum | 171.50 ± 1.08 *,** | 0.328 ± 0.02 ** | −28.5 ± 4.02 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tansathien, K.; Ngawhirunpat, T.; Rangsimawong, W.; Patrojanasophon, P.; Opanasopit, P.; Nuntharatanapong, N. In Vitro Biological Activity and In Vivo Human Study of Porcine-Placenta-Extract-Loaded Nanovesicle Formulations for Skin and Hair Rejuvenation. Pharmaceutics 2022, 14, 1846. https://doi.org/10.3390/pharmaceutics14091846
Tansathien K, Ngawhirunpat T, Rangsimawong W, Patrojanasophon P, Opanasopit P, Nuntharatanapong N. In Vitro Biological Activity and In Vivo Human Study of Porcine-Placenta-Extract-Loaded Nanovesicle Formulations for Skin and Hair Rejuvenation. Pharmaceutics. 2022; 14(9):1846. https://doi.org/10.3390/pharmaceutics14091846
Chicago/Turabian StyleTansathien, Kritsanaporn, Tanasait Ngawhirunpat, Worranan Rangsimawong, Prasopchai Patrojanasophon, Praneet Opanasopit, and Nopparat Nuntharatanapong. 2022. "In Vitro Biological Activity and In Vivo Human Study of Porcine-Placenta-Extract-Loaded Nanovesicle Formulations for Skin and Hair Rejuvenation" Pharmaceutics 14, no. 9: 1846. https://doi.org/10.3390/pharmaceutics14091846