
Advances in Chitosan-Based CRISPR/Cas9 Delivery Systems


Abstract
:1. Introduction
2. Chitosan in Gene Drug Delivery
3. Chitosan-Based CRISPR/Cas9 Delivery Systems
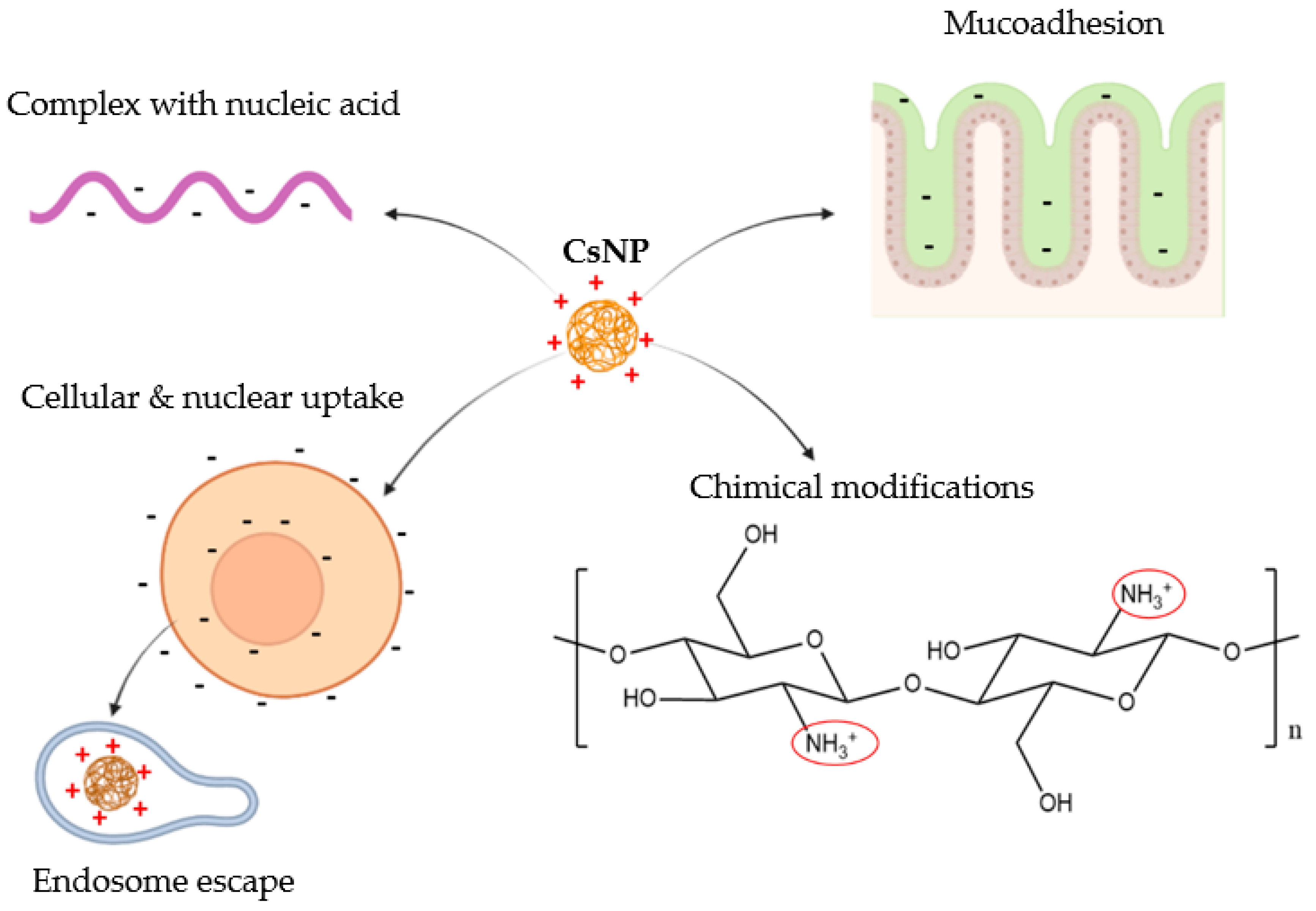
3.1. Strategies Using Pristine Chitosan Backbone
3.2. Strategies Improving the Specificity
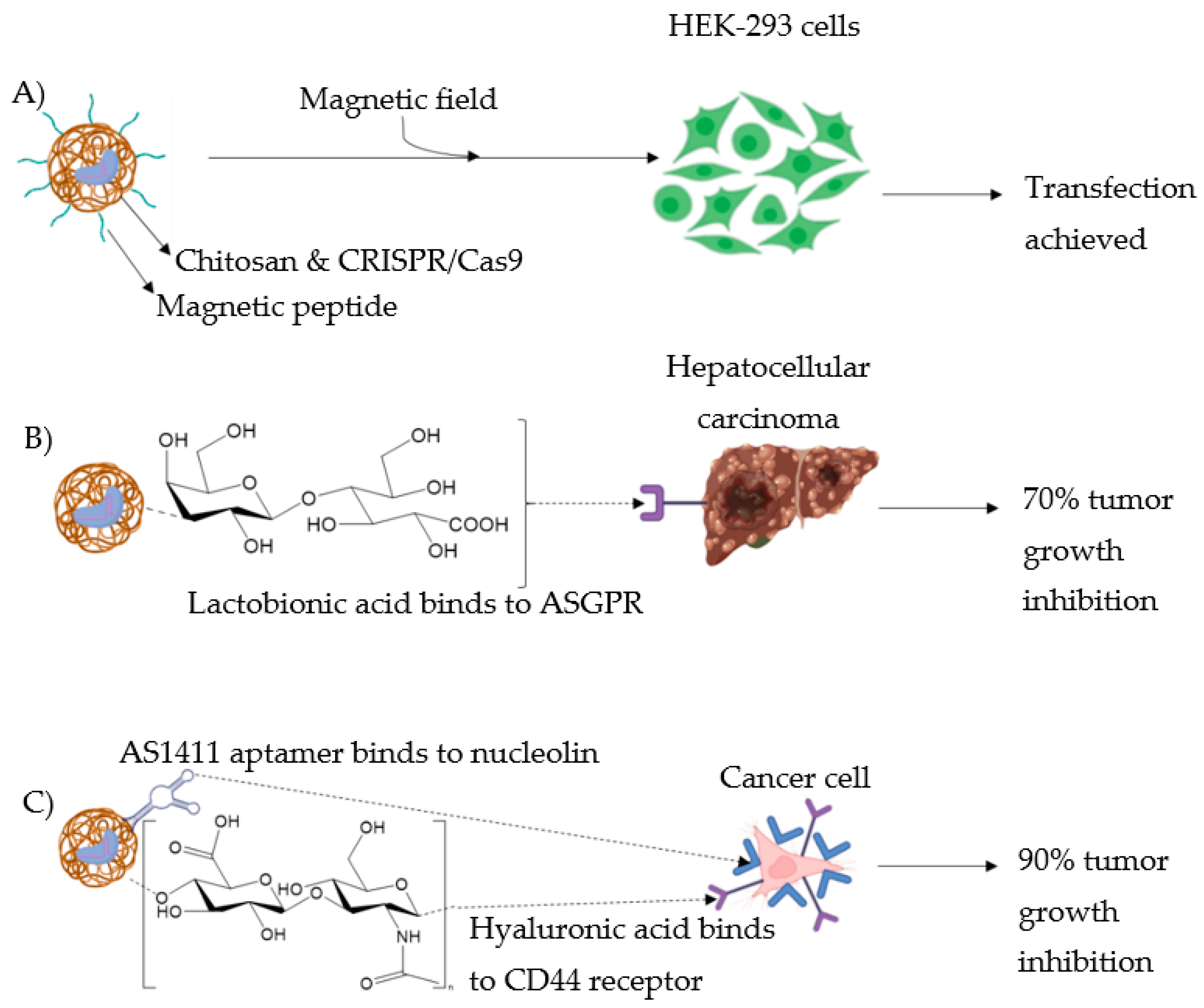
3.2.1. Magnetic Field
3.2.2. Use of Ligands
3.3. Strategies Increasing the Stability
3.3.1. Chitosan Tetrazole
3.3.2. PEGylation
3.3.3. Chitosan-Coated Red Fluorescent Protein
3.4. Strategies Combining Higher Specificity and Stability
3.4.1. Trimethyl Chitosan with Stimulus-Triggering and Targeting Responses
3.4.2. Carboxymethyl Chitosan with Cellular and Nuclear Targeting
3.5. Strategy Boosting the Buffering Ability
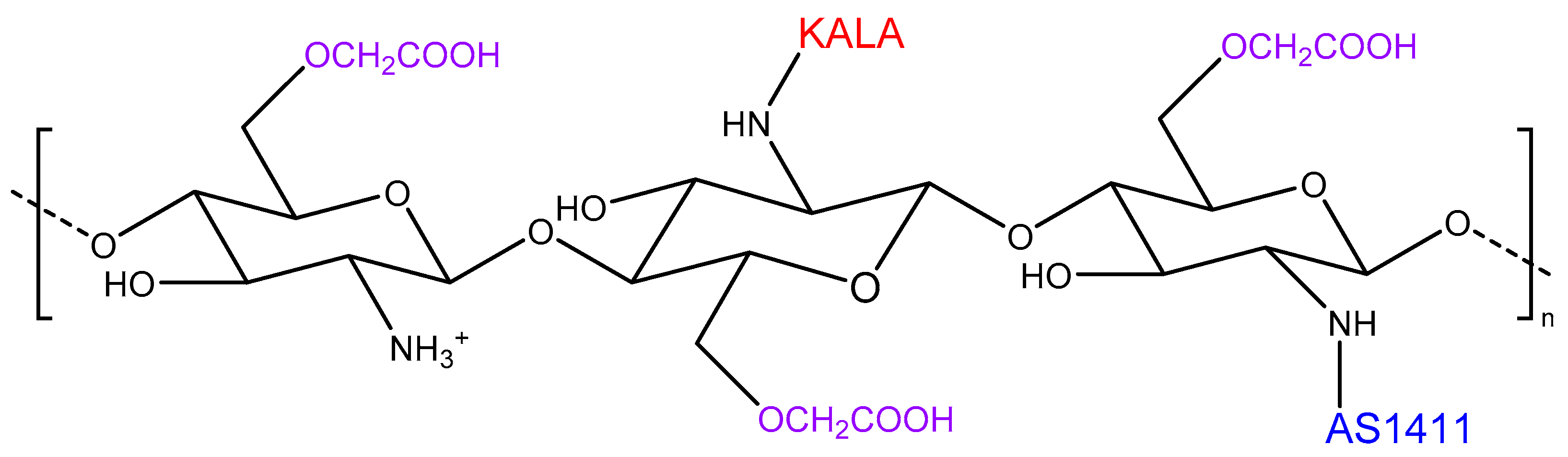
3.6. Strategy Combining Higher Specificity, Stability, and Buffering Ability
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Belete, T.M. The Current Status of Gene Therapy for the Treatment of Cancer. Biologics 2021, 15, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Li, M.; Lee, C.M.; Chakraborty, S.; Kim, H.W.; Bao, G.; Leong, K.W. CRISPR/Cas9-Based Genome Editing for Disease Modeling and Therapy: Challenges and Opportunities for Nonviral Delivery. Chem. Rev. 2017, 117, 9874–9906. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.B.T.; Platt, R.J.; Zhang, F. Therapeutic Genome Editing: Prospects and Challenges. Nat. Med. 2015, 21, 121–131. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, L.; Liu, H.; Cheng, K. Delivery Strategies of the CRISPR-Cas9 Gene-Editing System for Therapeutic Applications. J. Control. Release 2017, 266, 17–26. [Google Scholar] [CrossRef]
- Hasanzadeh, A.; Noori, H.; Jahandideh, A.; Niloofar, H.M.; Moghaddam, H.; Mahtab, S.; Mousavi, K.; Nourizadeh, H.; Saeedi, S.; Karimi, M.; et al. Smart Strategies for Precise Delivery of CRISPR/Cas9 in Genome Editing. ACS Appl. Bio. Mater. 2022, 5, 413–437. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Moyo, B.; Lee, C.M.; Leong, K.; Bao, G. Engineered Materials for in Vivo Delivery of Genome-Editing Machinery. Nat. Rev. Mater. 2019, 4, 726–737. [Google Scholar] [CrossRef]
- Yip, B.H. Recent Advances in CRISPR/Cas9 Delivery Strategies. Biomolecules 2020, 10, 839. [Google Scholar] [CrossRef] [PubMed]
- Alatorre-Meda, M.; Alvarez-Lorenzo, C.; Concheiro, A.; Taboada, P. UV and Near-IR Triggered Release from Polymeric Micelles and Nanoparticles. Smart Mater. Drug Deliv. 2013, 1, 304–348. [Google Scholar]
- Chew, W.L.; Tabebordbar, M.; Cheng, J.K.W.; Mali, P.; Wu, E.Y.; Ng, A.H.M.; Zhu, K.; Wagers, A.J.; Church, G.M. A Multifunctional AAV–CRISPR–Cas9 and Its Host Response. Nat. Methods 2016, 13, 868–874. [Google Scholar] [CrossRef]
- Givens, B.E.; Naguib, Y.W.; Geary, S.M.; Devor, E.J.; Salem, A.K. Nanoparticle-Based Delivery of CRISPR/Cas9 Genome-Editing Therapeutics. AAPS J. 2018, 20, 108. [Google Scholar] [CrossRef]
- Duan, L.; Ouyang, K.; Xu, X.; Xu, L.; Wen, C.; Zhou, X.; Qin, Z.; Xu, Z.; Sun, W.; Liang, Y. Nanoparticle Delivery of CRISPR/Cas9 for Genome Editing. Front. Genet. 2021, 12, 673286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Bahamondez-Canas, T.F.; Zhang, Y.; Leal, J.; Smyth, H.D.C. PEGylated Chitosan for Nonviral Aerosol and Mucosal Delivery of the CRISPR/Cas9 System in Vitro. Mol. Pharm. 2018, 15, 4814–4826. [Google Scholar] [CrossRef] [PubMed]
- Chuan, D.; Jin, T.; Fan, R.; Zhou, L.; Guo, G. Chitosan for Gene Delivery: Methods for Improvement and Applications. Adv. Colloid Interface Sci. 2019, 268, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Ojeda-Hernández, D.D.; Canales-Aguirre, A.A.; Matias-Guiu, J.; Gomez-Pinedo, U.; Mateos-Díaz, J.C. Potential of Chitosan and Its Derivatives for Biomedical Applications in the Central Nervous System. Front. Bioeng. Biotechnol. 2020, 8, 389. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Shen, C.; Gao, L.; Li, D.; Shang, Y.; Yin, K.; Zhao, D.; Cheng, W.; Quan, D. Anti-Inflammatory Activity of Chitosan Nanoparticles Carrying NF-ΚB/P65 Antisense Oligonucleotide in RAW264.7 Macropghage Stimulated by Lipopolysaccharide. Colloids Surf. 2016, 142, 297–306. [Google Scholar] [CrossRef]
- Garg, P.; Kumar, S.; Pandey, S.; Seonwoo, H.; Choung, P.H.; Koh, J.; Chung, J.H. Triphenylamine Coupled Chitosan with High Buffering Capacity and Low Viscosity for Enhanced Transfection in Mammalian Cells, in Vitro and in Vivo. J. Mater. Chem. B 2013, 1, 6053–6065. [Google Scholar] [CrossRef]
- Garcia, B.B.; Mertins, O.; da Silva, E.R.; Mathews, P.D.; Han, S.W. Arginine-Modified Chitosan Complexed with Liposome Systems for Plasmid DNA Delivery. Colloids Surf. B Biointerfaces 2020, 193, 111131. [Google Scholar] [CrossRef]
- Sosnik, A.; Das Neves, J.; Sarmento, B. Mucoadhesive Polymers in the Design of Nano-Drug Delivery Systems for Administration by Non-Parenteral Routes: A Review. Prog. Polym. Sci. 2014, 39, 2030–2075. [Google Scholar] [CrossRef]
- Sogias, I.A.; Williams, A.C.; Khutoryanskiy, V.V. Why Is Chitosan Mucoadhesive? Biomacromolecules 2008, 9, 1837–1842. [Google Scholar] [CrossRef]
- Puras, G.; Zarate, J.; Aceves, M.; Murua, A.; Díaz, A.R.; Avilés-Triguero, M.; Fernández, E.; Pedraz, J.L. Low Molecular Weight Oligochitosans for Non-Viral Retinal Gene Therapy. Eur. J. Pharm. Biopharm. 2013, 83, 131–140. [Google Scholar] [CrossRef]
- Liu, L.; Dong, X.; Zhu, D.; Song, L.; Zhang, H.; Leng, X.G. TAT-LHRH Conjugated Low Molecular Weight Chitosan as a Gene Carrier Specific for Hepatocellular Carcinoma Cells. Int. J. Nanomed. 2014, 9, 2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbet, C.; Ragelle, H.; Pourcelle, V.; Vanvarenberg, K.; Marchand-Brynaert, J.; Préat, V.; Feron, O. Delivery of SiRNA Targeting Tumor Metabolism Using Non-Covalent PEGylated Chitosan Nanoparticles: Identification of an Optimal Combination of Ligand Structure, Linker and Grafting Method. J. Control. Release 2016, 223, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Nugrahaningsih, D.A.A.; Purnomo, E.; Wasityastuti, W.; Martien, R.; Arfian, N.; Hartatik, T. BMPR2 Editing in Fibroblast NIH3T3 Using CRISPR/Cas9 Affecting BMPR2 MRNA Expression and Proliferation. Indones. Biomed. J. 2022, 14, 45–51. [Google Scholar] [CrossRef]
- Tuder, R.M.; Abman, S.H.; Braun, T.; Capron, F.; Stevens, T.; Thistlethwaite, P.A.; Haworth, S.G. Development and Pathology of Pulmonary Hypertension. J. Am. Coll. Cardiol. 2009, 54, S3–S9. [Google Scholar] [CrossRef] [PubMed]
- Srivastav, A.; Gupta, K.; Chakraborty, D.; Dandekar, P.; Jain, R. Efficiency of Chitosan-Coated PLGA Nanocarriers for Cellular Delivery of SiRNA and CRISPR/Cas9 Complex. J. Pharm. Innov. 2020, 17, 180–193. [Google Scholar] [CrossRef]
- Chronopoulou, L.; Massimi, M.; Giardi, M.F.; Cametti, C.; Devirgiliis, L.C.; Dentini, M.; Palocci, C. Chitosan-Coated PLGA Nanoparticles: A Sustained Drug Release Strategy for Cell Cultures. Colloids Surf. B Biointerfaces 2013, 103, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Alallam, B.; Doolaanea, A.A.; Oo, M.K.; Nasir, M.H.M.; Taher, M. Influence of Nanoparticles Surface Coating on Physicochemical Properties for CRISPR Gene Delivery. J. Drug Deliv. Sci. Technol. 2021, 66, 102910. [Google Scholar] [CrossRef]
- Lee, M.H.; Lin, C.C.; Thomas, J.L.; Li, J.A.; Lin, H.Y. Cellular Reprogramming with Multigene Activation by the Delivery of CRISPR/DCas9 Ribonucleoproteins via Magnetic Peptide-Imprinted Chitosan Nanoparticles. Mater. Today Bio 2021, 9, 100091. [Google Scholar] [CrossRef]
- Zhang, B.C.; Wu, P.Y.; Zou, J.J.; Jiang, J.L.; Zhao, R.R.; Luo, B.Y.; Liao, Y.Q.; Shao, J.W. Efficient CRISPR/Cas9 Gene-Chemo Synergistic Cancer Therapy via a Stimuli-Responsive Chitosan-Based Nanocomplex Elicits Anti-Tumorigenic Pathway Effect. Chem. Eng. J. 2020, 393, 124688. [Google Scholar] [CrossRef]
- Zheng, G.; Zhao, R.; Xu, A.; Shen, Z.; Chen, X.; Shao, J. Co-Delivery of Sorafenib and SiVEGF Based on Mesoporous Silica Nanoparticles for ASGPR Mediated Targeted HCC Therapy. Eur. J. Pharm. Sci. 2018, 111, 492–502. [Google Scholar] [CrossRef]
- Ghosh, A.; Dasgupta, D.; Ghosh, A.; Roychoudhury, S.; Kumar, D.; Gorain, M.; Butti, R.; Datta, S.; Agarwal, S.; Gupta, S.; et al. MiRNA199a-3p Suppresses Tumor Growth, Migration, Invasion and Angiogenesis in Hepatocellular Carcinoma by Targeting VEGFA, VEGFR1, VEGFR2, HGF and MMP2. Cell Death Dis. 2017, 8, e2706. [Google Scholar] [CrossRef] [PubMed]
- Khademi, Z.; Ramezani, M.; Alibolandi, M.; Zirak, M.R.; Salmasi, Z.; Abnous, K.; Taghdisi, S.M. A Novel Dual-Targeting Delivery System for Specific Delivery of CRISPR/Cas9 Using Hyaluronic Acid, Chitosan and AS1411. Carbohydr. Polym. 2022, 292, 119691. [Google Scholar] [CrossRef] [PubMed]
- Mattheolabakis, G.; Milane, L.; Singh, A.; Amiji, M.M. Hyaluronic Acid Targeting of CD44 for Cancer Therapy: From Receptor Biology to Nanomedicine. J Drug Target. 2015, 23, 605–618. [Google Scholar] [CrossRef]
- He, X.-Y.; Ren, X.-H.; Peng, Y.; Zhang, J.-P.; Ai, S.-L.; Liu, B.-Y.; Xu, C.; Cheng, S.-X.; He, X.-Y.; Ren, X.-H.; et al. Aptamer/Peptide-Functionalized Genome-Editing System for Effective Immune Restoration through Reversal of PD-L1-Mediated Cancer Immunosuppression. Adv. Mater. 2020, 32, 2000208. [Google Scholar] [CrossRef]
- Wang, Y.; Wen, L.; Zhao, S.H.; Ai, Z.H.; Guo, J.Z.; Liu, W.C. FoxM1 Expression Is Significantly Associated with Cisplatin-Based Chemotherapy Resistance and Poor Prognosis in Advanced Non-Small Cell Lung Cancer Patients. Lung Cancer 2013, 79, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Rabiee, N.; Bagherzadeh, M.; Tavakolizadeh, M.; Pourjavadi, A.; Atarod, M.; Webster, T.J. Synthesis, Characterization and Mechanistic Study of Nano Chitosan Tetrazole as a Novel and Promising Platform for CRISPR Delivery. Int. J. Polym. Mater. Polym. Biomater. 2020, 71, 116–126. [Google Scholar] [CrossRef]
- Caprifico, A.E.; Foot, P.J.S.; Polycarpou, E.; Calabrese, G. Overcoming the Protein Corona in Chitosan-Based Nanoparticles. Drug Discov. Today 2021, 26, 1825–1840. [Google Scholar] [CrossRef]
- Rastegari, A.; Mottaghitalab, F.; Dinarvand, R.; Amini, M.; Arefian, E.; Gholami, M.; Atyabi, F. Inhibiting Hepatic Gluconeogenesis by Chitosan Lactate Nanoparticles Containing CRTC2 SiRNA Targeted by Poly(Ethylene Glycol)-Glycyrrhetinic Acid. Drug Deliv. Transl. Res. 2019, 9, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Guo, T.; Zhou, D.; Hu, Y.; Zhou, H.; Wang, S.; Chen, J.; Zhang, Z. A Novel Glutathione Modified Chitosan Conjugate for Efficient Gene Delivery. J. Control. Release 2011, 154, 177–188. [Google Scholar] [CrossRef]
- Hatakeyama, H.; Akita, H.; Harashima, H. A Multifunctional Envelope Type Nano Device (MEND) for Gene Delivery to Tumours Based on the EPR Effect: A Strategy for Overcoming the PEG Dilemma. Adv. Drug Deliv. Rev. 2011, 63, 152–160. [Google Scholar] [CrossRef]
- Yang, Q.; Jacobs, T.M.; McCallen, J.D.; Moore, D.T.; Huckaby, J.T.; Edelstein, J.N.; Lai, S.K. Analysis of Pre-Existing IgG and IgM Antibodies against Polyethylene Glycol (PEG) in the General Population. Anal. Chem. 2016, 88, 11804–11812. [Google Scholar] [CrossRef] [PubMed]
- Suk, J.S.; Kim, A.J.; Trehan, K.; Schneider, C.S.; Cebotaru, L.; Woodward, O.M.; Boylan, N.J.; Boyle, M.P.; Lai, S.K.; Guggino, W.B.; et al. Lung Gene Therapy with Highly Compacted DNA Nanoparticles That Overcome the Mucus Barrier. J. Control. Release 2014, 178, 8–17. [Google Scholar] [CrossRef]
- Song, Y.; Salinas, D.; Nielson, D.W.; Verkman, A.S. Hyperacidity of Secreted Fluid from Submucosal Glands in Early Cystic Fibrosis. Am. J. Physiol.-Cell Physiol. 2006, 290, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, Y.; Williams, R.O.; Smyth, H.D.C. Development of PEGylated Chitosan/CRISPR-Cas9 Dry Powders for Pulmonary Delivery via Thin-Film Freeze-Drying. Int. J. Pharm. 2021, 605, 120831. [Google Scholar] [CrossRef] [PubMed]
- Sanchis, J.; Corrigan, C.; Levy, M.L.; Viejo, J.L. Inhaler Devices—From Theory to Practice. Respir. Med. 2013, 107, 495–502. [Google Scholar] [CrossRef]
- Qiao, J.; Sun, W.; Lin, S.; Jin, R.; Ma, L.; Liu, Y. Cytosolic Delivery of CRISPR/Cas9 Ribonucleoproteins for Genome Editing Using Chitosan-Coated Red Fluorescent Protein. Chem. Commun. 2019, 55, 4707–4710. [Google Scholar] [CrossRef]
- Mout, R.; Ray, M.; Yesilbag Tonga, G.; Lee, Y.W.; Tay, T.; Sasaki, K.; Rotello, V.M. Direct Cytosolic Delivery of CRISPR/Cas9-Ribonucleoprotein for Efficient Gene Editing. ACS Nano 2017, 11, 2452–2458. [Google Scholar] [CrossRef]
- Zhou, W.; Cui, H.; Ying, L.; Yu, X.-F. Enhanced Cytosolic Delivery and Release of CRISPR/Cas9 by Black Phosphorus Nanosheets for Genome Editing. Angew. Chem. 2018, 130, 10425–10429. [Google Scholar] [CrossRef]
- Mourya, V.K.; Inamdar, N.N. Trimethyl Chitosan and Its Applications in Drug Delivery. J. Mater. Sci. Mater. Med. 2009, 20, 1057–1079. [Google Scholar] [CrossRef]
- Li, Q.; Lv, X.; Tang, C.; Yin, C. Co-Delivery of Doxorubicin and CRISPR/Cas9 or RNAi-Expressing Plasmid by Chitosan-Based Nanoparticle for Cancer Therapy. Carbohydr. Polym. 2022, 287, 119315. [Google Scholar] [CrossRef]
- Geisberger, G.; Gyenge, E.B.; Maake, C.; Patzke, G.R. Trimethyl and Carboxymethyl Chitosan Carriers for Bio-Active Polymer-Inorganic Nanocomposites. Carbohydr. Polym. 2013, 91, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.T.; Warren, N.J.; Lewis, A.L.; Armes, S.P.; Battaglia, G. Effect of PH and Temperature on PMPC-PDPA Copolymer Self-Assembly. Macromolecules 2013, 46, 1400–1407. [Google Scholar] [CrossRef]
- Fasehee, H.; Dinarvand, R.; Ghavamzadeh, A.; Esfandyari-Manesh, M.; Moradian, H.; Faghihi, S.; Ghaffari, S.H. Delivery of Disulfiram into Breast Cancer Cells Using Folate-Receptor-Targeted PLGA-PEG Nanoparticles: In Vitro and in Vivo Investigations. J. Nanobiotechnol. 2016, 14, 32. [Google Scholar] [CrossRef]
- Kalliola, S.; Repo, E.; Srivastava, V.; Zhao, F.; Heiskanen, J.P.; Sirviö, J.A.; Liimatainen, H.; Sillanpää, M. Carboxymethyl Chitosan and Its Hydrophobically Modified Derivative as PH-Switchable Emulsifiers. Langmuir 2018, 34, 2800–2806. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Nam, J.P.; Kim, Y.M.; Kim, J.H.; Nah, J.W.; Jang, M.K. Branched Polyethylenimine-Grafted-Carboxymethyl Chitosan Copolymer Enhances the Delivery of PDNA or SiRNA in Vitro and in Vivo. Int. J. Nanomed. 2013, 8, 3663. [Google Scholar]
- Liu, B.Y.; He, X.Y.; Xu, C.; Xu, L.; Ai, S.L.; Cheng, S.X.; Zhuo, R.X. A Dual-Targeting Delivery System for Effective Genome Editing and in Situ Detecting Related Protein Expression in Edited Cells. Biomacromolecules 2018, 19, 2957–2968. [Google Scholar] [CrossRef]
- Ren, W.X.; Han, J.; Uhm, S.; Jang, Y.J.; Kang, C.; Kim, J.H.; Kim, J.S. Recent Development of Biotin Conjugation in Biological Imaging, Sensing, and Target Delivery. Chem. Commun. 2015, 51, 10403–10418. [Google Scholar] [CrossRef]
- Peyressatre, M.; Prével, C.; Pellerano, M.; Morris, M.C. Targeting Cyclin-Dependent Kinases in Human Cancers: From Small Molecules to Peptide Inhibitors. Cancers 2015, 7, 179–237. [Google Scholar] [CrossRef]
- He, X.Y.; Liu, B.Y.; Peng, Y.; Zhuo, R.X.; Cheng, S.X. Multifunctional Vector for Delivery of Genome Editing Plasmid Targeting β-Catenin to Remodulate Cancer Cell Properties. ACS Appl. Mater. Interfaces 2019, 11, 226–237. [Google Scholar] [CrossRef]
- Park, D.; Lee, J.Y.; Cho, H.K.; Hong, W.J.; Kim, J.; Seo, H.; Choi, I.; Lee, Y.; Kim, J.; Min, S.J.; et al. Cell-Penetrating Peptide-Patchy Deformable Polymeric Nanovehicles with Enhanced Cellular Uptake and Transdermal Delivery. Biomacromolecules 2018, 19, 2682–2690. [Google Scholar] [CrossRef]
- Shekhar, S.; Roy, A.; Hong, D.; Kumta, P.N. Nanostructured Silicate Substituted Calcium Phosphate (NanoSiCaPs) Nanoparticles—Efficient Calcium Phosphate Based Non-Viral Gene Delivery Systems. Mater. Sci. Eng. C 2016, 69, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Rabiee, N.; Bagherzadeh, M.; Ghadiri, A.M.; Kiani, M.; Ahmadi, S.; Jajarmi, V.; Fatahi, Y.; Aldhaher, A.; Tahriri, M.; Webster, T.J.; et al. Calcium-Based Nanomaterials and Their Interrelation with Chitosan: Optimization for PCRISPR Delivery. J. Nanostructure Chem. 2021, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.Y.; He, X.Y.; Zhuo, R.X.; Cheng, S.X. Tumor Targeted Genome Editing Mediated by a Multi-Functional Gene Vector for Regulating Cell Behaviors. J. Control. Release 2018, 291, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Miura, N.; Akita, H.; Tateshita, N.; Nakamura, T.; Harashima, H. Modifying Antigen-Encapsulating Liposomes with KALA Facilitates MHC Class I Antigen Presentation and Enhances Anti-Tumor Effects. Mol. Ther. 2017, 25, 1003–1013. [Google Scholar] [CrossRef]
- Zhou, Y.; Han, C.; Li, D.; Yu, Z.; Li, F.; Li, F.; An, Q.; Bai, H.; Zhang, X.; Duan, Z.; et al. Cyclin-Dependent Kinase 11p110 (CDK11p110) Is Crucial for Human Breast Cancer Cell Proliferation and Growth. Sci. Rep. 2015, 5, 10433. [Google Scholar] [CrossRef]
- Cyranoski, D. First Trial of CRISPR in People: Chinese Team Approved to Test Gene-Edited Cells in People with Lung Cancer. Nature 2016, 535, 476–477. [Google Scholar] [CrossRef]
- Aggarwal, P.; Hall, J.B.; McLeland, C.B.; Dobrovolskaia, M.A.; McNeil, S.E. Nanoparticle Interaction with Plasma Proteins as it Relates to Particle Biodistribution, Biocompatibility and Therapeutic Efficacy. Adv. Drug Deliv. Rev. 2009, 61, 428–437. [Google Scholar] [CrossRef]
- Barui, A.K.; Oh, J.Y.; Jana, B.; Kim, C.; Ryu, J. Cancer-Targeted Nanomedicine: Overcoming the Barrier of the Protein Corona. Adv. Ther. 2020, 3, 1900124. [Google Scholar] [CrossRef]
- Xing, L.; Fan, Y.T.; Zhou, T.J.; Gong, J.H.; Cui, L.H.; Cho, K.H.; Choi, Y.J.; Jiang, H.L.; Cho, C.S. Chemical Modification of Chitosan for Efficient Vaccine Delivery. Molecules 2018, 23, 229. [Google Scholar] [CrossRef]
- Liu, Z.; Lv, D.; Liu, S.; Gong, J.; Wang, D.; Xiong, M.; Chen, X.; Xiang, R.; Tan, X. Alginic Acid-Coated Chitosan Nanoparticles Loaded with Legumain DNA Vaccine: Effect against Breast Cancer in Mice. PLoS ONE 2013, 8, e60190. [Google Scholar] [CrossRef]
- Pandey, A.P.; Sawant, K.K. Polyethylenimine: A Versatile, Multifunctional Non-Viral Vector for Nucleic Acid Delivery. Mater. Sci. Eng. C 2016, 68, 904–918. [Google Scholar] [CrossRef] [PubMed]
- Ryu, N.; Kim, M.A.; Park, D.; Lee, B.; Kim, Y.R.; Kim, K.H.; Baek, J.I.; Kim, W.J.; Lee, K.Y.; Kim, U.K. Effective PEI-Mediated Delivery of CRISPR-Cas9 Complex for Targeted Gene Therapy. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 2095–2102. [Google Scholar] [CrossRef] [PubMed]
- Akinc, A.; Thomas, M.; Klibanov, A.M.; Langer, R. Exploring Polyethylenimine-Mediated DNA Transfection and the Proton Sponge Hypothesis. J. Gene Med. 2005, 7, 657–663. [Google Scholar] [CrossRef]
- Di Gioia, S.; Conese, M. Polyethylenimine-Mediated Gene Delivery to the Lung and Therapeutic Applications. Drug Des. Devel. Ther. 2008, 2, 163. [Google Scholar]
- Kunath, K.; Von Harpe, A.; Fischer, D.; Petersen, H.; Bickel, U.; Voigt, K.; Kissel, T. Low-Molecular-Weight Polyethylenimine as a Non-Viral Vector for DNA Delivery: Comparison of Physicochemical Properties, Transfection Efficiency and in Vivo Distribution with High-Molecular-Weight Polyethylenimine. J. Control. Release 2003, 89, 113–125. [Google Scholar] [CrossRef]
- Cardarelli, F.; Digiacomo, L.; Marchini, C.; Amici, A.; Salomone, F.; Fiume, G.; Rossetta, A.; Gratton, E.; Pozzi, D.; Caracciolo, G. The Intracellular Trafficking Mechanism of Lipofectamine-Based Transfection Reagents and Its Implication for Gene Delivery. Sci. Rep. 2016, 6, 25879. [Google Scholar] [CrossRef] [PubMed]
- Cahalane, C.; Bonezzi, J.; Shelestak, J.; Clements, R.; Boika, A.; Yun, Y.H.; Shriver, L.P. Targeted Delivery of Anti-Inflammatory and Imaging Agents to Microglial Cells with Polymeric Nanoparticles. Mol. Pharm. 2020, 17, 1816–1826. [Google Scholar] [CrossRef]
- Dos Santos Silva, M.; Cocenza, D.S.; Grillo, R.; de Melo, N.F.S.; Tonello, P.S.; de Oliveira, L.C.; Cassimiro, D.L.; Rosa, A.H.; Fraceto, L.F. Paraquat-Loaded Alginate/Chitosan Nanoparticles: Preparation, Characterization and Soil Sorption Studies. J. Hazard. Mater. 2011, 190, 366–374. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties of Chitosan | Strategies | Transfection Efficiency of CRISPR/Cas9 | Reference |
|---|---|---|---|
| Pristine properties | CsNPs | Achieved in vitro | [23] |
| Cs-coated PLGA NPs | >80% in vitro | [25] | |
| Cs-coated AG NPs | >20% in vitro | [27] | |
| Improved specificity | Magnetic peptide-imprinted CsNPs | Achieved in vitro | [28] |
| La-ligand conjugated CsNPs | 39% in vitro 33% in vivo | [29] | |
| Ha and AS1411-decorated CsNPs | 20–30% in vitro 90% in vivo | [32] | |
| Increased stability | Cs tetrazole | >25% in vitro | [36] |
| PEGylated Cs | ~15% in vitro | [12,44] | |
| Cs-coated red fluorescent protein | ~12.5% in vitro | [46] | |
| Improved specificity and stability | DPA and Fa-ligand-modified TMC | ~80% in vitro ~30% in vivo | [50] |
| Biotin- and AS1411-modified CmCs | 90% in vitro | [56] | |
| AS1411 and TAT-modified CmCs | ~30% in vitro | [59] | |
| Boosted buffering ability | Cs and calcium phosphate NPs | 25% in vitro | [62] |
| Improved specificity, stability, and buffering ability | AS1411 and KALA-modified CmCs | >75% in vitro | [63] |
| Strategies | Size (nm) | Zeta Potential (mV) | Reference |
|---|---|---|---|
| CsNPs | Not specified | Not specified | [23] |
| Cs-coated PLGA NPs | ~300 | +32 | [25] |
| Cs-coated AG NPs | ~400 | +33 | [27] |
| Magnetic-peptide-imprinted CsNPs | Not specified | Not specified | [28] |
| La-ligand-conjugated CsNPs | ~200 | +26 | [29] |
| Ha and AS1411-decorated CsNPs | ~200 | +20 | [32] |
| Cs tetrazole | ~80 | +40 | [36] |
| PEGylated Cs | ~200 | +20 | [12,44] |
| Cs-coated red fluorescent protein | ~200 | +50 | [46] |
| DPA and Fa-ligand-modified TMC | ~200 | +20 | [50] |
| Biotin and AS1411-modified CmCs | ~300 | −9 | [56] |
| AS1411 and TA-modified CmCs | ~300 | −7 | [59] |
| Cs and calcium phosphate NPs | ~150 | +27 | [62] |
| AS1411 and KALA-modified CmCs | ~240 | +17 | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caprifico, A.E.; Foot, P.J.S.; Polycarpou, E.; Calabrese, G. Advances in Chitosan-Based CRISPR/Cas9 Delivery Systems. Pharmaceutics 2022, 14, 1840. https://doi.org/10.3390/pharmaceutics14091840
Caprifico AE, Foot PJS, Polycarpou E, Calabrese G. Advances in Chitosan-Based CRISPR/Cas9 Delivery Systems. Pharmaceutics. 2022; 14(9):1840. https://doi.org/10.3390/pharmaceutics14091840
Chicago/Turabian StyleCaprifico, Anna E., Peter J. S. Foot, Elena Polycarpou, and Gianpiero Calabrese. 2022. "Advances in Chitosan-Based CRISPR/Cas9 Delivery Systems" Pharmaceutics 14, no. 9: 1840. https://doi.org/10.3390/pharmaceutics14091840