
Non-Cellular Layers of the Respiratory Tract: Protection against Pathogens and Target for Drug Delivery


Abstract
:1. Introduction
2. Organization of the Respiratory Tract
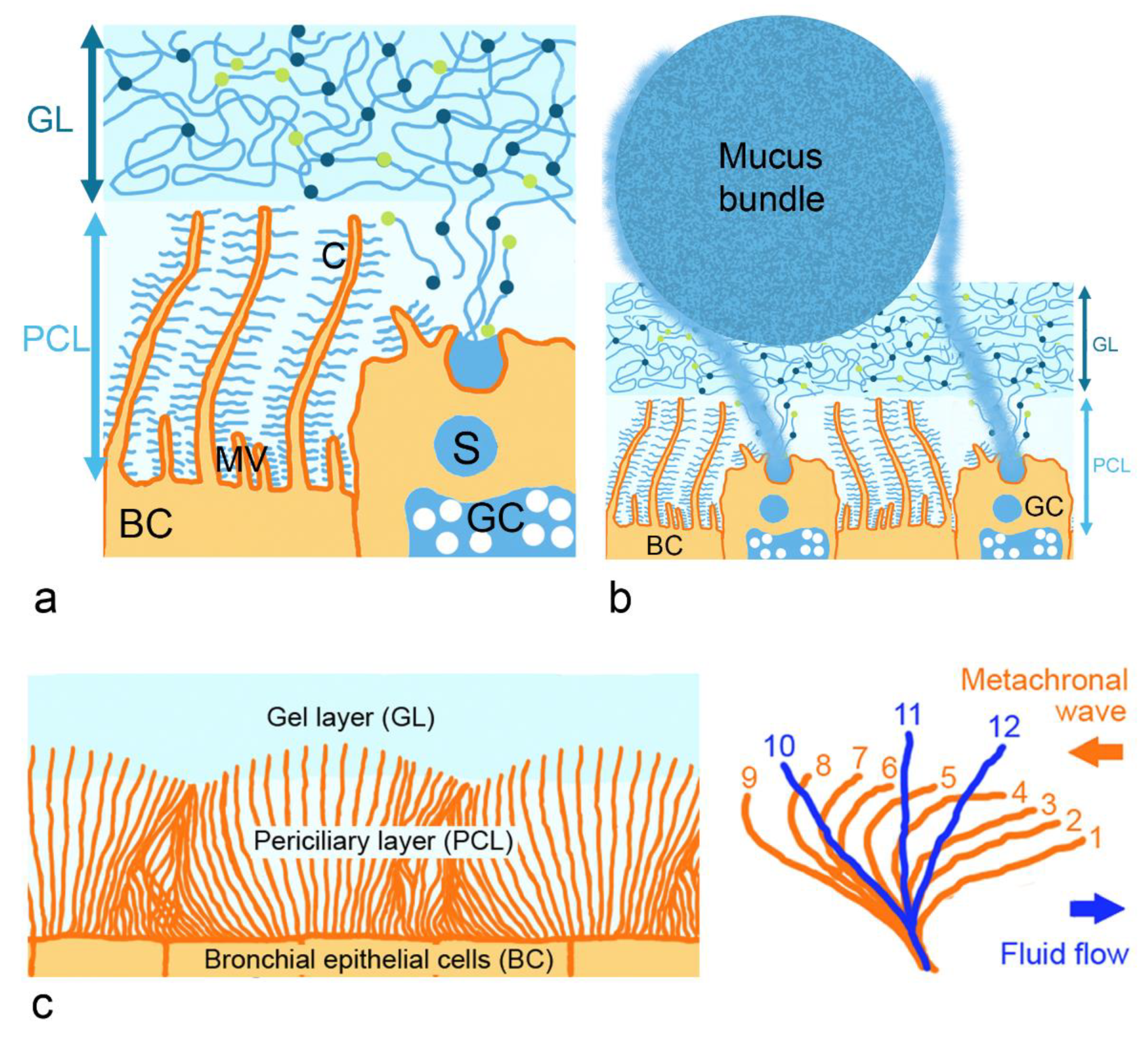
2.1. Architecture and Rheological Properties of the Mucus Layer
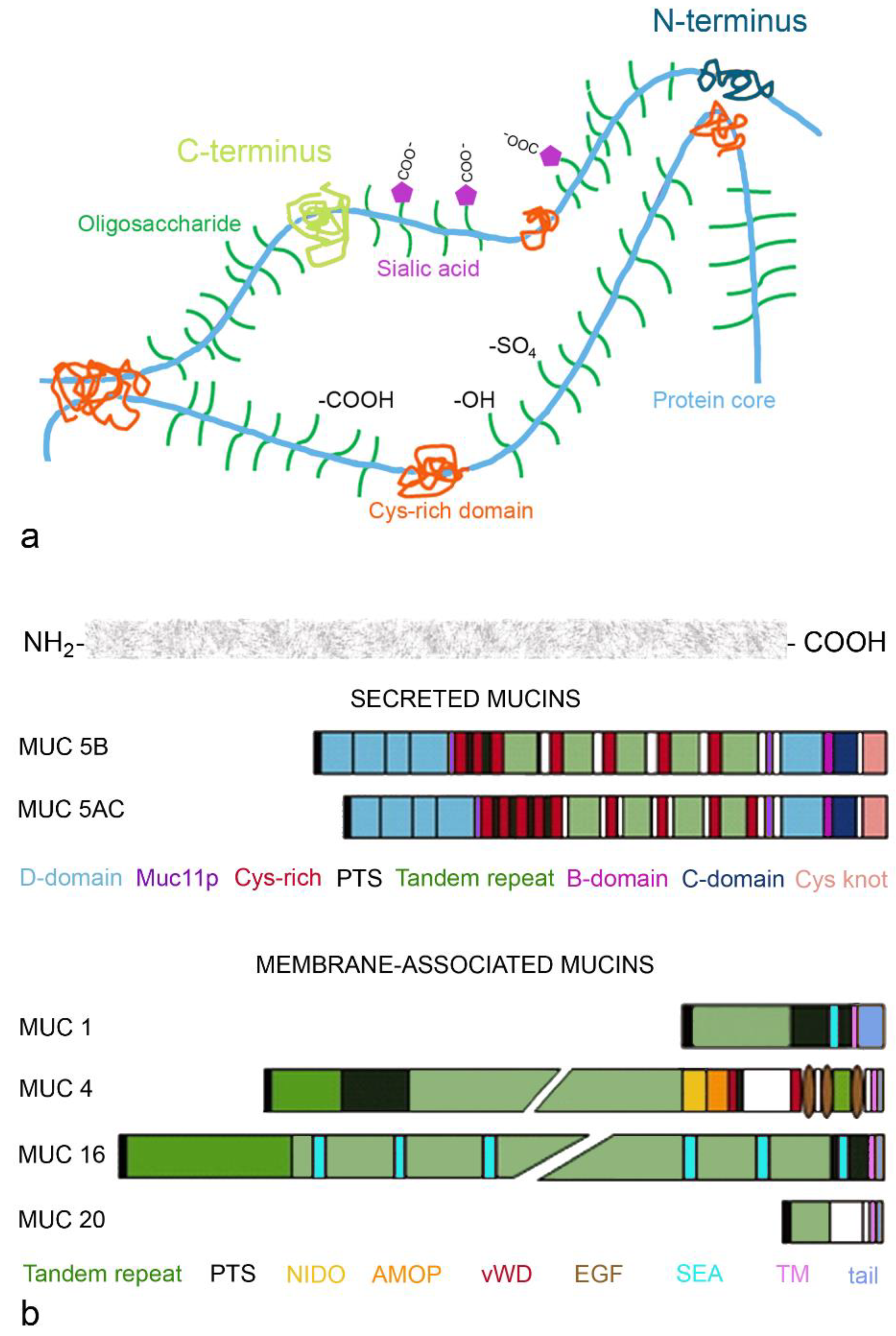
2.1.1. Gel-Forming Mucins
2.1.2. Membrane-Bound Mucins
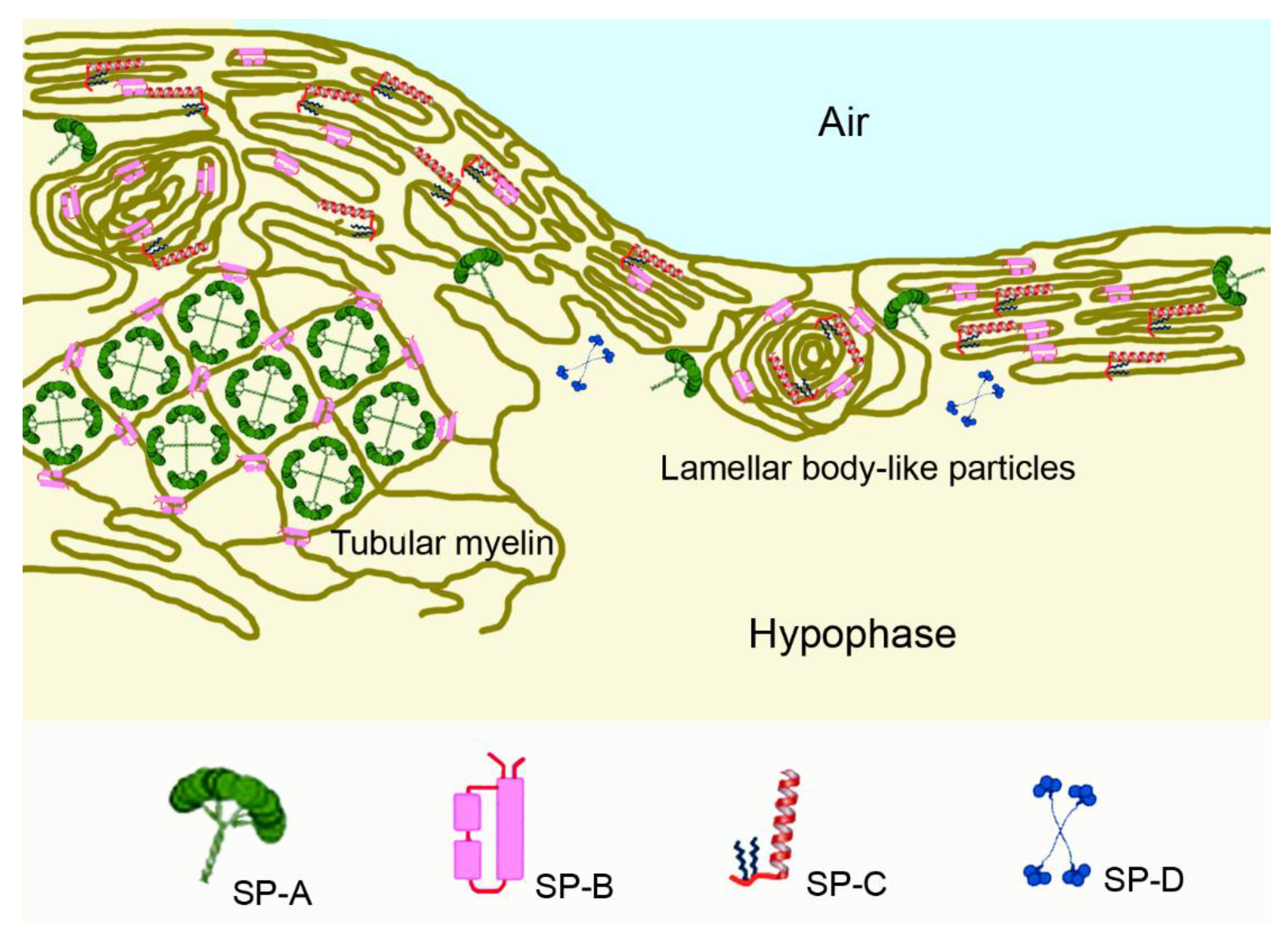
2.2. The Pulmonary Surfactant Layer
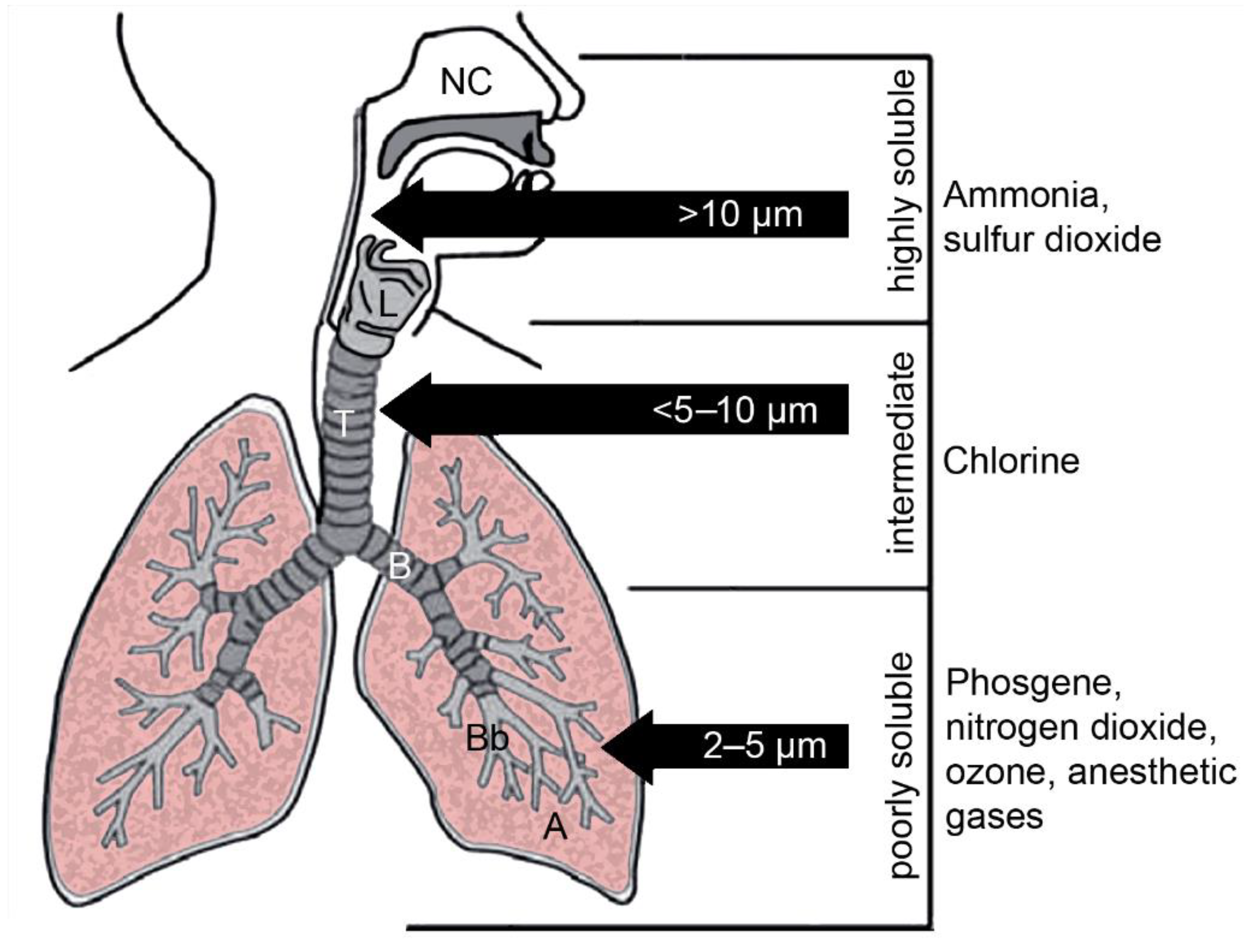
3. Contact of Gases with the Non-Cellular Surfaces of the Lung
4. Contact of Pathogens with Non-Cellular Surfaces
4.1. Infectious Aerosols
4.2. Viruses
4.2.1. Influenza Virus
4.2.2. Corona Viruses
4.2.3. Respiratory Syncytial Virus
4.3. Bacteria
4.4. Fungi
5. Overcoming Non-Cellular Barriers
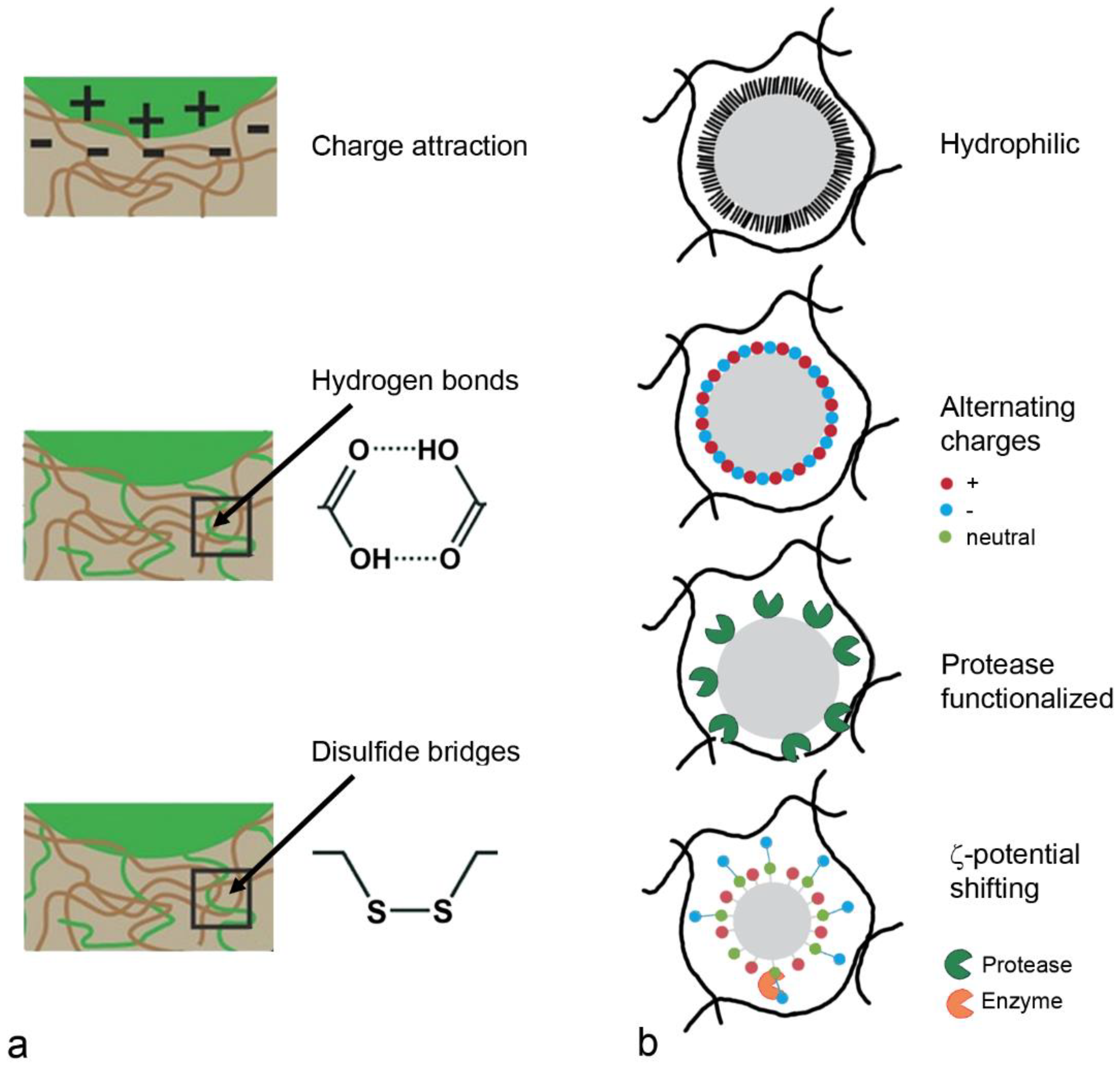
5.1. Mucus-Targeting Strategies
5.1.1. Mucoadhesion
5.1.2. Mucopermeation
5.1.3. Carriers for Nasal Drug Delivery
5.1.4. Carriers for Pulmonary Drug Delivery
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACE2 | Angiotensin converting enzyme 2 |
| AIDS | Acquired immune deficiency syndrome |
| AD | Alzheimer’s disease |
| AMOP | Adhesion-associated domain in MUC4 and other proteins |
| BACE1 | β-site amyloid precursor protein cleaving enzyme 1 |
| CFD | Computational fluid dynamic |
| COVID-19 | Coronavirus disease 2019 |
| DLPC | Dilauroylphosphatidylcholine |
| DMBT1 | Deleted in malignant brain tumor 1 |
| DPPC | Dipalmitoyl phosphatidylcholine |
| EGF | Epidermal growth factor |
| FDA | Drug administration |
| GRAS | Generally regarded as safe |
| HA | Hemagglutinin |
| LL-37 | Cathelicidins |
| MERS | Middle east respiratory syndrome coronavirus |
| NA | Neuraminidase |
| NIDO | Nidogen-like domain |
| PC | Phosphatidylcholine |
| PD | Parkinson’s disease |
| PEG | Polyethylene glycol |
| PLA | Polylactic acid |
| PLGA | Poly(lactic-co-glycolic acid) |
| PLUC | Nasal epithelium clone |
| PTS | Proline, threonine, serine rich |
| PVA | Polyvinyl acetate |
| RSV | Respiratory syncytial virus |
| SARS-CoV | Severe acute respiratory syndrome corona virus |
| SEA | Sea urchin sperm protein |
| SP | Surfactant protein |
| TPGS | Tocopherol polyethylene glycol succinate |
| vWD | von Willebrand factor D |
References
- Janeway, C., Jr.; Travers, P. Principles of innate and adaptive immunity. In Innate Immunity, Immunobiology: The Immune System in Health and Disease; Janeway, C., Jr., Travers, P., Walport, M., Shlomchik, M., Eds.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Oltulu, P.; Ince, B.; Kokbudak, N.; Findik, S.; Kilinc, F. Measurement of epidermis, dermis, and total skin thicknesses from six different body regions with a new ethical histometric technique. Turk. J. Plast. Surg. 2018, 26, 56–61. [Google Scholar] [CrossRef]
- Walton, K.D.; Freddo, A.M.; Wang, S.; Gumucio, D.L. Generation of intestinal surface: An absorbing tale. Development 2016, 143, 2261–2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widdicombe, J.H. Volume of airway surface liquid in health and disease. Am. J. Respir. Crit. Care Med. 2002, 165, 1566. [Google Scholar] [CrossRef] [PubMed]
- Atuma, C.; Strugala, V.; Allen, A.; Holm, L. The adherent gastrointestinal mucus gel layer: Thickness and physical state in vivo. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G922–G929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahy, J.V.; Dickey, B.F. Airway mucus function and dysfunction. N. Engl. J. Med. 2010, 363, 2233–2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reznik, G.K. Comparative anatomy, physiology, and function of the upper respiratory tract. Environ. Health Perspect. 1990, 85, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Lenfant, C. Discovery of Endogenous Surfactant and Overview of its Metabolism and Actions. In Lung Surfactants, Basic Science and Clinical Applications; Notter, R., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 2000; Volume 149. [Google Scholar]
- Sekerdag, E. Nasal Physiology and Drug Transport. In Nanotechnology Methods for Neurological Diseases and Brain Tumors. Drug Delivery across the Blood-Brain Barrier; Gürsoy-Özdemir, Y., Bozdağ-Pehlivan, S., Sekerdag, E., Eds.; Elsevier: London, UK, 2017; pp. 93–102. [Google Scholar] [CrossRef]
- Leal, J.; Smyth, H.D.C.; Ghosh, D. Physicochemical properties of mucus and their impact on transmucosal drug delivery. Int. J. Pharm. 2017, 532, 555–572. [Google Scholar] [CrossRef]
- Ottaviano, G.; Gerven, L. Objective Assessment of Nasal Function. In Cummings Otolaryngology: Head and Neck Surgery; Flint, P., Haughey, B., Lund, V., Robbins, K., Thomas, J., Lesperance, M., Francis, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 37, pp. 603–615.e7. [Google Scholar]
- Witten, J.; Samad, T.; Ribbeck, K. Selective permeability of mucus barriers. Curr. Opin. Biotechnol. 2018, 52, 124–133. [Google Scholar] [CrossRef]
- Wallace, L.E.; Liu, M.; van Kuppeveld, F.J.M.; de Vries, E.; de Haan, C.A.M. Respiratory mucus as a virus-host range determinant. Trends Microbiol. 2021, 29, 983–992. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, M.; Luo, Y.; Deng, L.; Hu, Z.; Song, Y. Determination of rheology and surface tension of airway surface liquid: A review of clinical relevance and measurement techniques. Respir. Res. 2019, 20, 274. [Google Scholar] [CrossRef]
- Ridley, C.; Thornton, D.J. Mucins: The frontline defence of the lung. Biochem. Soc. Trans. 2018, 46, 1099–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randell, S.H.; Boucher, R.C.; University of North Carolina Virtual Lung, G. Effective mucus clearance is essential for respiratory health. Am. J. Respir. Cell Mol. Biol. 2006, 35, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Hansson, G.C. Mucus and mucins in diseases of the intestinal and respiratory tracts. J. Intern. Med. 2019, 285, 479–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottier, M.; Blanchon, S.; Pelle, G.; Bequignon, E.; Isabey, D.; Coste, A.; Escudier, E.; Grotberg, J.B.; Papon, J.F.; Filoche, M.; et al. A new index for characterizing micro-bead motion in a flow induced by ciliary beating: Part I, experimental analysis. PLoS Comput. Biol. 2017, 13, e1005605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flint, P. Objective Assessment of Nasal Function. In Cummings Otolaryngology: Head and Neck Surgery; Flint, P., Haughey, B., Lund, V., Robbins, K., Thomas, J., Lesperance, M., Francis, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; Volume 3. [Google Scholar]
- Shang, Y.; Inthavong, K.; Qiu, D.; Singh, N.; He, F.; Tu, J. Prediction of nasal spray drug absorption influenced by mucociliary clearance. PLoS ONE 2021, 16, e0246007. [Google Scholar] [CrossRef] [PubMed]
- Chovancová, M.; Elcner, J. The pressure gradient in the human respiratory tract. EPJ Web Conf. 2014, 67, 02047. [Google Scholar] [CrossRef] [Green Version]
- Levy, R.; Hill, D.B.; Forest, M.G.; Grotberg, J.B. Pulmonary fluid flow challenges for experimental and mathematical modeling. Integr. Comp. Biol. 2014, 54, 985–1000. [Google Scholar] [CrossRef] [Green Version]
- Lillehoj, E.P.; Kato, K.; Lu, W.; Kim, K.C. Cellular and molecular biology of airway mucins. Int. Rev. Cell Mol. Biol. 2013, 303, 139–202. [Google Scholar] [CrossRef] [Green Version]
- Meldrum, O.W.; Chotirmall, S.H. Mucus, Microbiomes and Pulmonary Disease. Biomedicines 2021, 9, 675. [Google Scholar] [CrossRef]
- Nawroth, J.C.; van der Does, A.M.; Ryan Firth, A.; Kanso, E. Multiscale mechanics of mucociliary clearance in the lung. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190160. [Google Scholar] [CrossRef]
- Denneny, E.; Sahota, J.; Beatson, R.; Thornton, D.; Burchell, J.; Porter, J. Mucins and their receptors in chronic lung disease. Clin. Transl. Immunol. 2020, 9, e01120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahmasebi, F.; Nath, R.; Sokolovsky, N.; Scaffidi, J.; Boley, J.; Mehra, G.; Sayanseh, A. Incidental Finding of Raised CA125: A Cause for Concern? Crit. Care Obst. Gyne. 2018, 5, 3. [Google Scholar] [CrossRef]
- Kirch, J.; Guenther, M.; Doshi, N.; Schaefer, U.F.; Schneider, M.; Mitragotri, S.; Lehr, C.M. Mucociliary clearance of micro- and nanoparticles is independent of size, shape and charge--an ex vivo and in silico approach. J. Control. Release 2012, 159, 128–134. [Google Scholar] [CrossRef]
- Rosenberg, O.; Seiliev, A.; Zhuikov, A. Lung Surfactants: Correlation Between Biophysical Characteristics, Composition, and Therapeutic Efficacy. In Interactions of Liposomes with Biological Milieu; Gregoriadis, G., Ed.; CRC Press: Boca Raton, FL, USA, 2007; Volume III, pp. 317–345. [Google Scholar]
- Bernhard, W. Lung surfactant: Function and composition in the context of development and respiratory physiology. Ann. Anat. 2016, 208, 146–150. [Google Scholar] [CrossRef]
- Cochrane, C.G. A critical examination of the role of SP-B in alveolar expansion. Appl. Cardiopul. Pathophysiol. 2004, 13, 27–28. [Google Scholar]
- Cañadas, O.; Olmeda, B.; Alonso, A.; Pérez-Gil, J. Lipid-Protein and Protein-Protein Interactions in the Pulmonary Surfactant System and Their Role in Lung Homeostasis. Int. J. Mol. Sci. 2020, 21, 3708. [Google Scholar] [CrossRef]
- Hickman-Davis, J.; Wright, J.; Matalon, S. Surfactant. Surfactant Protein A (SP-A) In Encyclopedia of Respiratory Medicine; Laurent, G., Shapiro, S., Eds.; Elsevier Ltd.: Oxford, UK, 2006; pp. 143–147. [Google Scholar] [CrossRef]
- Mason, R.; Dobbs, L. Alveolar Epithelium and Pulmonary Surfactant. In Murray and Nadel’s Textbook of Respiratory Medicine; Broaddus, V., Mason, R., Ernst, J., King, J.T.E., Krevans, J., Lazarus, S., Murray, J., Nadel, J., Slutsky, A., Gotway, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 134–149.e5. [Google Scholar] [CrossRef]
- Adegunsoye, D.; Rafeq, S. Pulmonary Medicine. Available online: https://www.pulmonologyadvisor.com/home/decision-support-in-medicine/pulmonary-medicine/toxic-inhalational-lung-injury/ (accessed on 20 February 2022).
- Public Health England. Ammonia Toxicological Overview. In Compendium of Chemical Hazard; PHE Publication Gateway Number 2014790; Government: London, UK, 2015; pp. 1–12. [Google Scholar]
- White, C.W.; Martin, J.G. Chlorine gas inhalation: Human clinical evidence of toxicity and experience in animal models. Proc. Am. Thorac Soc. 2010, 7, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massa, C.B.; Scott, P.; Abramova, E.; Gardner, C.; Laskin, D.L.; Gow, A.J. Acute chlorine gas exposure produces transient inflammation and a progressive alteration in surfactant composition with accompanying mechanical dysfunction. Toxicol. Appl. Pharmacol. 2014, 278, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Huang, S.; Meng, X.; Zhang, J.; Yu, S.; Li, J.; Shi, M.; Fan, H.; Zhao, Y. Mechanism of Phosgene-Induced Acute Lung Injury and Treatment Strategy. Int. J. Mol. Sci. 2021, 22, 933. [Google Scholar] [CrossRef]
- Williams, R.A.; Rhoades, R.A.; Adams, W.S. The response of lung tissue and surfactant to nitrogen dioxide exposure. Arch. Intern. Med. 1971, 128, 101–108. [Google Scholar] [CrossRef]
- Zaiats, L.M. Effect of sulfur dioxide on surface activity of the pulmonary surfactants and ultrastructure of type II alveolocytes. Fiziolohichnyi Zhurnal 2004, 50, 52–56. [Google Scholar] [PubMed]
- Oda, Y.; Isohama, Y.; Kai, H.; Okano, Y.; Takahama, K.; Miyata, T. Increased production and/or secretion of pulmonary surfactant in rats by long term sulfur dioxide exposure. J. Pharmacobiodyn. 1989, 12, 726–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraswat, V. Effects of anaesthesia techniques and drugs on pulmonary function. Indian J. Anaesth. 2015, 59, 557–564. [Google Scholar] [CrossRef]
- Kopp Lugli, A.; Yost, C.S.; Kindler, C.H. Anaesthetic mechanisms: Update on the challenge of unravelling the mystery of anaesthesia. Eur. J. Anaesthesiol. 2009, 26, 807–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobin, W.R.; Kaiser, H.E.; Groeger, A.M.; Lema, M.J. The effects of volatile anesthetic agents on pulmonary surfactant function. In Vivo 2000, 14, 157–163. [Google Scholar] [PubMed]
- Stephanova, E.; Valtcheva-Sarker, R.; Topouzova-Hristova, T.; Lalchev, Z. Influence of Volatile Anaesthetics on Lung Cells and Lung Surfactant. Biotechnol. Biotechnol. Equ. 2007, 21, 393. [Google Scholar] [CrossRef] [Green Version]
- Aldemir, T.; Turan, S.; Gomceli, I.; Ayik, I.; Erdem, O.; Yagar, S.; Kul, N.; Erdemli, O. A comparison of the effects of desflurane and isoflurane on rat pulmonary parenchyme histopathology and malondialdehyde levels. J. Pak. Med. Assoc. 2012, 62, 1174–1178. [Google Scholar]
- Hawco, M.W.; Davis, P.J.; Keough, K.M. Lipid fluidity in lung surfactant: Monolayers of saturated and unsaturated lecithins. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1981, 51, 509–515. [Google Scholar] [CrossRef]
- Malacrida, L.; Reta, G.; Piriz, H.; Rocchiccioli, F.; Botti, H.; Denicola, A.; Briva, A. Sevoflurane anesthesia deteriorates pulmonary surfactant promoting alveolar collapse in male Sprague-Dawley rats. Pulm. Pharmacol. Ther. 2014, 28, 122–129. [Google Scholar] [CrossRef]
- Gaya da Costa, M.; Kalmar, A.F.; Struys, M. Inhaled Anesthetics: Environmental Role, Occupational Risk, and Clinical Use. J. Clin. Med. 2021, 10, 1306. [Google Scholar] [CrossRef]
- Tellier, R.; Li, Y.; Cowling, B.J.; Tang, J.W. Recognition of aerosol transmission of infectious agents: A commentary. BMC Infect. Dis. 2019, 19, 101. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Weng, W.; Huang, Q. Characterizations of particle size distribution of the droplets exhaled by sneeze. Interface 2013, 10, 20130560. [Google Scholar] [CrossRef] [PubMed]
- Loudon, R.G.; Roberts, R.M. Droplet expulsion from the respiratory tract. Am. Rev. Respir. Dis. 1967, 95, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.Y.H.; Wan, M.P.; Morawska, L.; Johnson, G.R.; Ristovski, Z.D.; Hargreaves, M.; Mengersen, K.; Corbett, S.; Li, Y.; Xie, X.; et al. Characterization of expiration air jets and droplet size distributions immediately at the mouth opening. J. Aerosol Sci. 2009, 40, 122–133. [Google Scholar] [CrossRef] [Green Version]
- Gorbunov, B. Aerosol Particles Generated by Coughing and Sneezing of a SARS-CoV-2 (COVID-19) Host Travel over 30 m Distance. Aerosol Air Qual. Res. 2021, 21, 200468. [Google Scholar] [CrossRef]
- Li, Y.; Tang, X.X. Abnormal Airway Mucus Secretion Induced by Virus Infection. Front. Immunol. 2021, 12, 701443. [Google Scholar] [CrossRef]
- Carlson, T.L.; Lock, J.Y.; Carrier, R.L. Engineering the Mucus Barrier. Annu. Rev. Biomed. Eng. 2018, 20, 197–220. [Google Scholar] [CrossRef]
- Scheuch, G. Breathing Is Enough: For the Spread of Influenza Virus and SARS-CoV-2 by Breathing Only. J. Aerosol Med. Pulm. Drug Deliv. 2020, 33, 230–234. [Google Scholar] [CrossRef]
- Cao, D.; Gao, Y.; Liang, B. Structural Insights into the Respiratory Syncytial Virus RNA Synthesis Complexes. Viruses 2021, 13, 834. [Google Scholar] [CrossRef]
- Dreschers, S.; Adams, C. Common Cold Viruses. In Encyclopedia of Virology; Van Regenmortel, M., Mahy, B., Eds.; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Mifsud, E.J.; Kuba, M.; Barr, I.G. Innate Immune Responses to Influenza Virus Infections in the Upper Respiratory Tract. Viruses 2021, 13, 2090. [Google Scholar] [CrossRef]
- Clementi, N.; Ghosh, S.; De Santis, M.; Castelli, M.; Criscuolo, E.; Zanoni, I.; Clementi, M.; Mancini, N. Viral respiratory pathogens and lung injury. Clin. Microbiol. Rev. 2021, 34, e00103–e00120. [Google Scholar] [CrossRef] [PubMed]
- Zanin, M.; Baviskar, P.; Webster, R.; Webby, R. The Interaction between Respiratory Pathogens and Mucus. Cell Host Microbe 2016, 19, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Numata, M.; Mitchell, J.R.; Tipper, J.L.; Brand, J.D.; Trombley, J.E.; Nagashima, Y.; Kandasamy, P.; Chu, H.W.; Harrod, K.S.; Voelker, D.R. Pulmonary surfactant lipids inhibit infections with the pandemic H1N1 influenza virus in several animal models. J. Biol. Chem. 2020, 295, 1704–1715. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Yang, H.; Ji, W.; Wu, W.; Chen, S.; Zhang, W.; Duan, G. Virology, Epidemiology, Pathogenesis, and Control of COVID-19. Viruses 2020, 12, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, W.; Ma, B.; Li, Z.; Wang, X.; Gao, X.; Li, Y.; Qin, B.; Shang, S.; Cui, S.; Tan, Z. Binding of the SARS-CoV-2 spike protein to glycans. Sci. Bull. 2021, 66, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Bose, M.; Mitra, B.; Mukherjee, P. Mucin signature as a potential tool to predict susceptibility to COVID-19. Physiol. Rep. 2021, 9, e14701. [Google Scholar] [CrossRef]
- Hsieh, M.H.; Beirag, N.; Murugaiah, V.; Chou, Y.C.; Kuo, W.S.; Kao, H.F.; Madan, T.; Kishore, U.; Wang, J.Y. Human Surfactant Protein D Binds Spike Protein and Acts as an Entry Inhibitor of SARS-CoV-2 Pseudotyped Viral Particles. Front. Immunol. 2021, 12, 641360. [Google Scholar] [CrossRef]
- Jung, H.E.; Kim, T.H.; Lee, H.K. Contribution of Dendritic Cells in Protective Immunity against Respiratory Syncytial Virus Infection. Viruses 2020, 12, 102. [Google Scholar] [CrossRef] [Green Version]
- Ascough, S.; Paterson, S.; Chiu, C. Induction and Subversion of Human Protective Immunity: Contrasting Influenza and Respiratory Syncytial Virus. Front. Immunol. 2018, 9, 323. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Priti, S.; Ankit, S. Bacteria Etiological Agents Causing Lower Respiratory Tract Infections and Their Resistance Patterns. Iran. Biomed. J. 2015, 19, 240–246. [Google Scholar] [CrossRef]
- González, C.; Rubio, M.; Romero-Vivas, J.; González, M.; Picazo, J.J. Bacteremic pneumonia due to Staphylococcus aureus: A comparison of disease caused by methicillin-resistant and methicillin-susceptible organisms. Clin. Infect. Dis. 1999, 29, 1171–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonas, M.; Cunha, B.A. Bacteremic Escherichia coli pneumonia. Arch. Intern. Med. 1982, 142, 2157–2159. [Google Scholar] [CrossRef] [PubMed]
- Morimura, A.; Hamaguchi, S.; Akeda, Y.; Tomono, K. Mechanisms Underlying Pneumococcal Transmission and Factors Influencing Host-Pneumococcus Interaction: A Review. Front. Cell. Infect. Microbiol. 2021, 11, 639450. [Google Scholar] [CrossRef] [PubMed]
- Catarrhalis, M. Information about Moraxella Catarrhalis. Available online: https://www.who.int/water_sanitation_health/dwq/gdwq3_11.pdf (accessed on 3 March 2022).
- Krekeler, C.; Ziehr, H.; Klein, J. Physical methods for characterization of microbial surfaces. Experientia 1989, 45, 1047–1055. [Google Scholar] [CrossRef]
- Wilson, W.W.; Wade, M.M.; Holman, S.C.; Champlin, F.R. Status of methods for assessing bacterial cell surface charge properties based on zeta potential measurements. J. Microbiol. Methods 2001, 43, 153–164. [Google Scholar] [CrossRef]
- Todar, K. Structure and Function of Bacterial Cells; Madison: Wisconsin, MI, USA, 2020; Available online: http://www.textbookofbacteriology.net/structure_6.html (accessed on 4 March 2022).
- Natarajan, K. Microbially Induced Mineral Beneficiation. In Biotechnology of Metals. Principles, Recovery Methods, and Environmental Concerns; Natarajan, K., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 243–304. [Google Scholar]
- Gottenbos, B.; Grijpma, D.W.; van der Mei, H.C.; Feijen, J.; Busscher, H.J. Antimicrobial effects of positively charged surfaces on adhering Gram-positive and Gram-negative bacteria. J. Antimicrob. Chemother. 2001, 48, 7–13. [Google Scholar] [CrossRef]
- Fernández-Blanco, J.A.; Fakih, D.; Arike, L.; Rodríguez-Piñeiro, A.M.; Martínez-Abad, B.; Skansebo, E.; Jackson, S.; Root, J.; Singh, D.; McCrae, C.; et al. Attached stratified mucus separates bacteria from the epithelial cells in COPD lungs. JCI Insight 2018, 3, e120994. [Google Scholar] [CrossRef]
- Bakshani, C.R.; Morales-Garcia, A.L.; Althaus, M.; Wilcox, M.D.; Pearson, J.P.; Bythell, J.C.; Burgess, J.G. Evolutionary conservation of the antimicrobial function of mucus: A first defence against infection. NPJ Biofilms Microbiomes 2018, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- García-Fojeda, B.; González-Carnicero, Z.; de Lorenzo, A.; Minutti, C.M.; de Tapia, L.; Euba, B.; Iglesias-Ceacero, A.; Castillo-Lluva, S.; Garmendia, J.; Casals, C. Lung Surfactant Lipids Provide Immune Protection Against Haemophilus influenzae Respiratory Infection. Front. Immunol. 2019, 10, 458. [Google Scholar] [CrossRef] [Green Version]
- Haagsman, H.P. Interactions of surfactant protein A with pathogens. Biochim. Biophys. Acta 1998, 1408, 264–277. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, K.M.; Huffnagle, G.B.; Dickson, R.P. The significance of Candida in the human respiratory tract: Our evolving understanding. Pathog. Dis. 2017, 75, ftx029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Lu, G.; Meng, G. Pathogenic Fungal Infection in the Lung. Front. Immunol. 2019, 10, 1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGinnis, M.; Tyring, S. Introduction to Mycology. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Stanford, F.A.; Voigt, K. Iron Assimilation during Emerging Infections Caused by Opportunistic Fungi with emphasis on Mucorales and the Development of Antifungal Resistance. Genes 2020, 11, 1296. [Google Scholar] [CrossRef] [PubMed]
- Latgé, J.-P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33, e00140-18. [Google Scholar] [CrossRef] [PubMed]
- Cowley, A.C.; Thornton, D.J.; Denning, D.W.; Horsley, A. Aspergillosis and the role of mucins in cystic fibrosis. Pediatr. Pulmonol. 2017, 52, 548–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Cissé, O.H.; Kovacs, J.A. A Molecular Window into the Biology and Epidemiology of Pneumocystis spp. Clin. Microbiol. Rev. 2018, 31, e00009-18. [Google Scholar] [CrossRef] [Green Version]
- Kaneshiro, E.S. The lipids of Pneumocystis carinii. Clin. Microbiol. Rev. 1998, 11, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Xiong, G.; Tsang, W.C.; Schätzlein, A.G.; Uchegbu, I.F. Nose-to-Brain Delivery. J. Pharmacol. Exp. Ther. 2019, 370, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Lehr, C.-M.; Poelma, F.G.J.; Junginger, H.E.; Tukker, J.J. An estimate of turnover time of intestinal mucus gel layer in the rat in situ loop. Int. J. Pharm. 1991, 70, 235–240. [Google Scholar] [CrossRef]
- Taherali, F.; Varum, F.; Basit, A.W. A slippery slope: On the origin, role and physiology of mucus. Adv. Drug Deliv. Rev. 2018, 124, 16–33. [Google Scholar] [CrossRef]
- Ali, M.S.; Pearson, J.P. Upper airway mucin gene expression: A review. Laryngoscope 2007, 117, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Ye, J.; Zhou, J.; Wang, W.; Wang, H.; Zheng, X.; Yang, Y.; Xia, X.; Liu, Y. Comparative study of mucoadhesive and mucus-penetrative nanoparticles based on phospholipid complex to overcome the mucus barrier for inhaled delivery of baicalein. Acta Pharm. Sin. 2020, 10, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Pardeshi, C.; Kulkarni, A.; Sonwane, R.; Belgamwar, V.; Chaudhari, P.; Surana, S. Mucoadhesive Nanoparticles: A Roadmap to Encounter the Challenge of Rapid Nasal Mucociliary Clearance. Indian J. Pharm. Educ. Res. 2019, 53, s17–s27. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, F.; Bruschi, M.; Evangelista, R.; Gremião, M. Mucoadhesive drug delivery systems. Braz. J. Pharm. Sci. 2010, 46, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Park, K. Appendix F—Chapter II.5.16—Drug Delivery Systems: H, Mucosal Drug Delivery. In Biomaterials Science, 3rd ed.; Ratner, B.D., Hoffman, A.S., Schoen, F.J., Lemons, J.E., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 1510–1518. [Google Scholar] [CrossRef]
- Alawdi, S.; Solanki, A. Mucoadhesive Drug Delivery Systems: A Review of Recent Developments. J. Sci. Res. Med. Biol. Sci. 2021, 2, 50–64. [Google Scholar] [CrossRef]
- Schneider, C.S.; Xu, Q.; Boylan, N.J.; Chisholm, J.; Tang, B.C.; Schuster, B.S.; Henning, A.; Ensign, L.M.; Lee, E.; Adstamongkonkul, P.; et al. Nanoparticles that do not adhere to mucus provide uniform and long-lasting drug delivery to airways following inhalation. Sci. Adv. 2017, 3, e1601556. [Google Scholar] [CrossRef] [Green Version]
- Khutoryanskiy, V.V. Beyond PEGylation: Alternative surface-modification of nanoparticles with mucus-inert biomaterials. Adv. Drug Deliv. Rev. 2018, 124, 140–149. [Google Scholar] [CrossRef]
- Schlenoff, J.B. Zwitteration: Coating surfaces with zwitterionic functionality to reduce nonspecific adsorption. Langmuir 2014, 30, 9625–9636. [Google Scholar] [CrossRef]
- Mahmood, A.; Laffleur, F.; Leonaviciute, G.; Bernkop-Schnürch, A. Protease-functionalized mucus penetrating microparticles: In-vivo evidence for their potential. Int. J. Pharm. 2017, 532, 177–184. [Google Scholar] [CrossRef]
- Samaridou, E.; Karidi, K.; Pereira de Sousa, I.; Cattoz, N.; Griffiths, P.; Kammona, O.; Bernkop-Schnürch, A.; Kiparissides, C. Enzyme-Functionalized PLGA Nanoparticles with Enhanced Mucus Permeation Rate. Nano Life 2014, 4, 1441013. [Google Scholar] [CrossRef]
- Prüfert, F.; Bonengel, S.; Köllner, S.; Griesser, J.; Wilcox, M.D.; Chater, P.I.; Pearson, J.P.; Bernkop-Schnürch, A. ζ potential changing nanoparticles as cystic fibrosis transmembrane conductance regulator gene delivery system: An in vitro evaluation. Nanomedicine 2017, 12, 2713–2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metz, J.K.; Scharnowske, L.; Hans, F.; Schnur, S.; Knoth, K.; Zimmer, H.; Limberger, M.; Groß, H.; Lehr, C.M.; Hittinger, M. Safety assessment of excipients (SAFE) for orally inhaled drug products. Altex 2020, 37, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Alp, G.; Aydogan, N. Enhancing the Spreading Behavior on Pulmonary Mucus Mimicking Subphase via Catanionic Surfactant Solutions: Toward Effective Drug Delivery through the Lungs. Mol. Pharm. 2018, 15, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Ghadiri, M.; Young, P.M.; Traini, D. Strategies to Enhance Drug Absorption via Nasal and Pulmonary Routes. Pharmaceutics 2019, 11, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, W.R.; Allansmith, M.R. Rapid, atraumatic method for obtaining nasal mucus samples. Ann. Otol. Rhinol. Laryngol. 1976, 85, 391–393. [Google Scholar] [CrossRef]
- Chaturvedi, M.; Kumar, M.; Pathak, K. A review on mucoadhesive polymer used in nasal drug delivery system. J. Adv. Pharm. Technol. Res. 2011, 2, 215–222. [Google Scholar] [CrossRef]
- Sonvico, F.; Clementino, A.; Buttini, F.; Colombo, G.; Pescina, S.; Stanisçuaski Guterres, S.; Raffin Pohlmann, A.; Nicoli, S. Surface-Modified Nanocarriers for Nose-to-Brain Delivery: From Bioadhesion to Targeting. Pharmaceutics 2018, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Samaridou, E.; Alonso, M.J. Nose-to-brain peptide delivery—The potential of nanotechnology. Bioorg. Med. Chem. 2018, 26, 2888–2905. [Google Scholar] [CrossRef]
- Sun, M.; Qin, D.; Fan, P.; Chen, X.; Liu, Y. Chitosan-centered nanosystems as sustained therapeutics for allergic rhinitis intervention: Inhibition of histamine-induced cascades. J. Control. Release 2021, 335, 422–436. [Google Scholar] [CrossRef]
- Lim, S.C.L.; Hor, C.P.; Tay, K.H.; Mat Jelani, A.; Tan, W.H.; Ker, H.B.; Chow, T.S.; Zaid, M.; Cheah, W.K.; Lim, H.H.; et al. Efficacy of Ivermectin Treatment on Disease Progression Among Adults With Mild to Moderate COVID-19 and Comorbidities: The I-TECH Randomized Clinical Trial. JAMA Intern. Med. 2022, 182, 426–435. [Google Scholar] [CrossRef]
- Aref, Z.F.; Bazeed, S.; Hassan, M.H.; Hassan, A.S.; Rashad, A.; Hassan, R.G.; Abdelmaksoud, A.A. Clinical, Biochemical and Molecular Evaluations of Ivermectin Mucoadhesive Nanosuspension Nasal Spray in Reducing Upper Respiratory Symptoms of Mild COVID-19. Int. J. Nanomed. 2021, 16, 4063–4072. [Google Scholar] [CrossRef] [PubMed]
- Porfiryeva, N.N.; Semina, I.I.; Salakhov, I.A.; Moustafine, R.I.; Khutoryanskiy, V.V. Mucoadhesive and mucus-penetrating interpolyelectrolyte complexes for nose-to-brain drug delivery. Nanomedicine 2021, 37, 102432. [Google Scholar] [CrossRef] [PubMed]
- Varga, P.; Ambrus, R.; Szabó-Révész, P.; Kókai, D.; Burián, K.; Bella, Z.; Fenyvesi, F.; Bartos, C. Physico-Chemical, In Vitro and Ex Vivo Characterization of Meloxicam Potassium-Cyclodextrin Nanospheres. Pharmaceutics 2021, 13, 1883. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Herzberg, M.; Huang, Z.; Hassenkam, T.; Nielsen, H.M. A free-floating mucin layer to investigate the effect of the local microenvironment in lungs on mucin-nanoparticle interactions. Acta Biomater. 2020, 104, 115–123. [Google Scholar] [CrossRef]
- Costabile, G.; Provenzano, R.; Azzalin, A.; Scoffone, V.C.; Chiarelli, L.R.; Rondelli, V.; Grillo, I.; Zinn, T.; Lepioshkin, A.; Savina, S.; et al. PEGylated mucus-penetrating nanocrystals for lung delivery of a new FtsZ inhibitor against Burkholderia cenocepacia infection. Nanomedicine 2020, 23, 102113. [Google Scholar] [CrossRef]
- Conte, G.; Costabile, G.; Baldassi, D.; Rondelli, V.; Bassi, R.; Colombo, D.; Linardos, G.; Fiscarelli, E.V.; Sorrentino, R.; Miro, A.; et al. Hybrid Lipid/Polymer Nanoparticles to Tackle the Cystic Fibrosis Mucus Barrier in siRNA Delivery to the Lungs: Does PEGylation Make the Difference? ACS Appl. Mater. Interfaces 2022, 14, 7565–7578. [Google Scholar] [CrossRef]
- Marslin, G.; Revina, A.M.; Khandelwal, V.K.; Balakumar, K.; Sheeba, C.J.; Franklin, G. PEGylated ofloxacin nanoparticles render strong antibacterial activity against many clinically important human pathogens. Colloids Surf. B Biointerfaces 2015, 132, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhu, W.; Jin, Q.; Pan, F.; Zhu, J.; Liu, Y.; Chen, L.; Shen, J.; Yang, Y.; Chen, Q.; et al. Inhalable nanocatchers for SARS-CoV-2 inhibition. Proc. Natl. Acad. Sci. USA 2021, 118, e2102957118. [Google Scholar] [CrossRef]
- Khaitov, M.; Nikonova, A.; Shilovskiy, I.; Kozhikhova, K.; Kofiadi, I.; Vishnyakova, L.; Nikolskii, A.; Gattinger, P.; Kovchina, V.; Barvinskaia, E.; et al. Silencing of SARS-CoV-2 with modified siRNA-peptide dendrimer formulation. Allergy 2021, 76, 2840–2854. [Google Scholar] [CrossRef]
- Huang, Z.; Kłodzińska, S.N.; Wan, F.; Nielsen, H.M. Nanoparticle-mediated pulmonary drug delivery: State of the art towards efficient treatment of recalcitrant respiratory tract bacterial infections. Drug Deliv. Transl. Res. 2021, 11, 1634–1654. [Google Scholar] [CrossRef]
- Rasul, R.M.; Tamilarasi Muniandy, M.; Zakaria, Z.; Shah, K.; Chee, C.F.; Dabbagh, A.; Rahman, N.A.; Wong, T.W. A review on chitosan and its development as pulmonary particulate anti-infective and anti-cancer drug carriers. Carbohydr. Polym. 2020, 250, 116800. [Google Scholar] [CrossRef] [PubMed]
- Dhayanandamoorthy, Y.; Antoniraj, M.G.; Kandregula, C.A.B.; Kandasamy, R. Aerosolized hyaluronic acid decorated, ferulic acid loaded chitosan nanoparticle: A promising asthma control strategy. Int. J. Pharm. 2020, 591, 119958. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, N.A.N.; El-Kemary, M.A. Silymarin/curcumin loaded albumin nanoparticles coated by chitosan as muco-inhalable delivery system observing anti-inflammatory and anti COVID-19 characterizations in oleic acid triggered lung injury and in vitro COVID-19 experiment. Int. J. Biol. Macromol. 2022, 198, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.S.; Rodrigues, M.T.; Domingues, R.M.A.; Torrado, E.; Reis, R.L.; Pedrosa, J.; Gomes, M.E. Exploring inhalable polymeric dry powders for anti-tuberculosis drug delivery. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 93, 1090–1103. [Google Scholar] [CrossRef] [PubMed]
- Drago, S.E.; Craparo, E.F.; Luxenhofer, R.; Cavallaro, G. Development of polymer-based nanoparticles for zileuton delivery to the lung: PMeOx and PMeOzi surface chemistry reduces interactions with mucins. Nanomedicine 2021, 37, 102451. [Google Scholar] [CrossRef]
- Zhao, J.; Su, J.; Qin, L.; Zhang, X.; Mao, S. Exploring the influence of inhaled liposome membrane fluidity on its interaction with pulmonary physiological barriers. Biomater. Sci. 2020, 8, 6786–6797. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Vaghasiya, K.; Gupta, P.; Singh, A.K.; Gupta, U.D.; Verma, R.K. Dynamic mucus penetrating microspheres for efficient pulmonary delivery and enhanced efficacy of host defence peptide (HDP) in experimental tuberculosis. J. Control. Release 2020, 324, 17–33. [Google Scholar] [CrossRef]
- Kim, Y.C.; Hsueh, H.T.; Kim, N.; Rodriguez, J.; Leo, K.T.; Rao, D.; West, N.E.; Hanes, J.; Suk, J.S. Strategy to enhance dendritic cell-mediated DNA vaccination in the lung. Adv. Ther. 2021, 4, 2000228. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungus | Composition | Reference |
|---|---|---|
| Aspergillus, trophic form | Chitin, β-1,3-glucan and β-1,6-glucan, glucomannan, galactosaminoglycan, glycosylated proteins | [89] |
| Aspergillus conidia | Chitin, β-1,3-glucan and β-1,6-glucan, melanin, rodlet layer | [88] |
| Cryptococcus | Chitin, β-1,3-glucan and β-1,6-glucan, mannans, glucuroxylomannan, galactoxylomannan | [88] |
| Mucorales | Chitin, β-1,3-glucan and β-1,6-glucan, melanin, extracellular polysaccharide, protein | [88] |
| Pneumocystis cysts | β-1,3-glucan and β-1,6-glucan, protein, mannans | [91] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fröhlich, E. Non-Cellular Layers of the Respiratory Tract: Protection against Pathogens and Target for Drug Delivery. Pharmaceutics 2022, 14, 992. https://doi.org/10.3390/pharmaceutics14050992
Fröhlich E. Non-Cellular Layers of the Respiratory Tract: Protection against Pathogens and Target for Drug Delivery. Pharmaceutics. 2022; 14(5):992. https://doi.org/10.3390/pharmaceutics14050992
Chicago/Turabian StyleFröhlich, Eleonore. 2022. "Non-Cellular Layers of the Respiratory Tract: Protection against Pathogens and Target for Drug Delivery" Pharmaceutics 14, no. 5: 992. https://doi.org/10.3390/pharmaceutics14050992