
SRC Kinase-Mediated Tyrosine Phosphorylation of TUBB3 Regulates Its Stability and Mitotic Spindle Dynamics in Prostate Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Cloning and Constructs
2.3. Transient Transfections
2.4. Cell Viability Assays
2.5. Immunoprecipitation (IP), Western Blots, and Antibodies
2.6. Subcellular Fractionation
2.7. Immunofluorescence (IF) and Quantification
2.8. Protein Stability Assays
2.9. Phosphorylation Predictive Software, and 3-D Molecular Modeling
3. Results
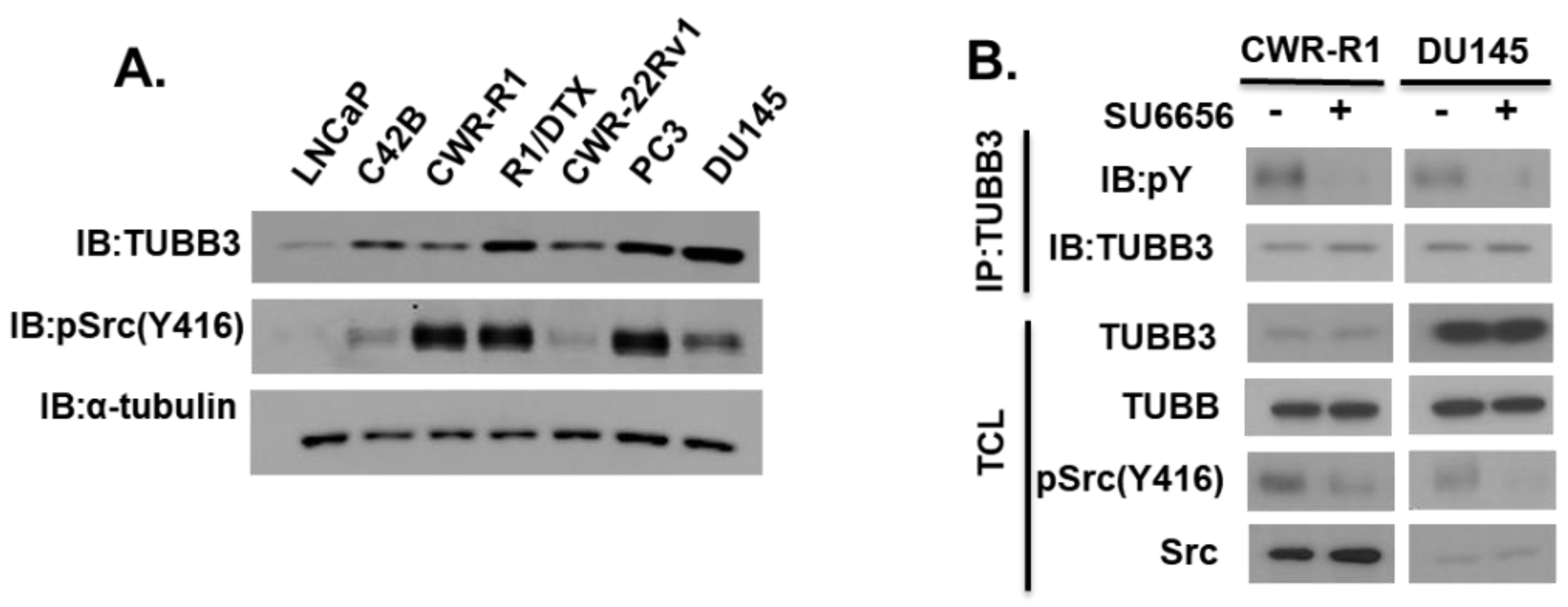
3.1. Tyrosine Phosphorylation of TUBB3 Mediated by SRC Kinase in PCa Cells
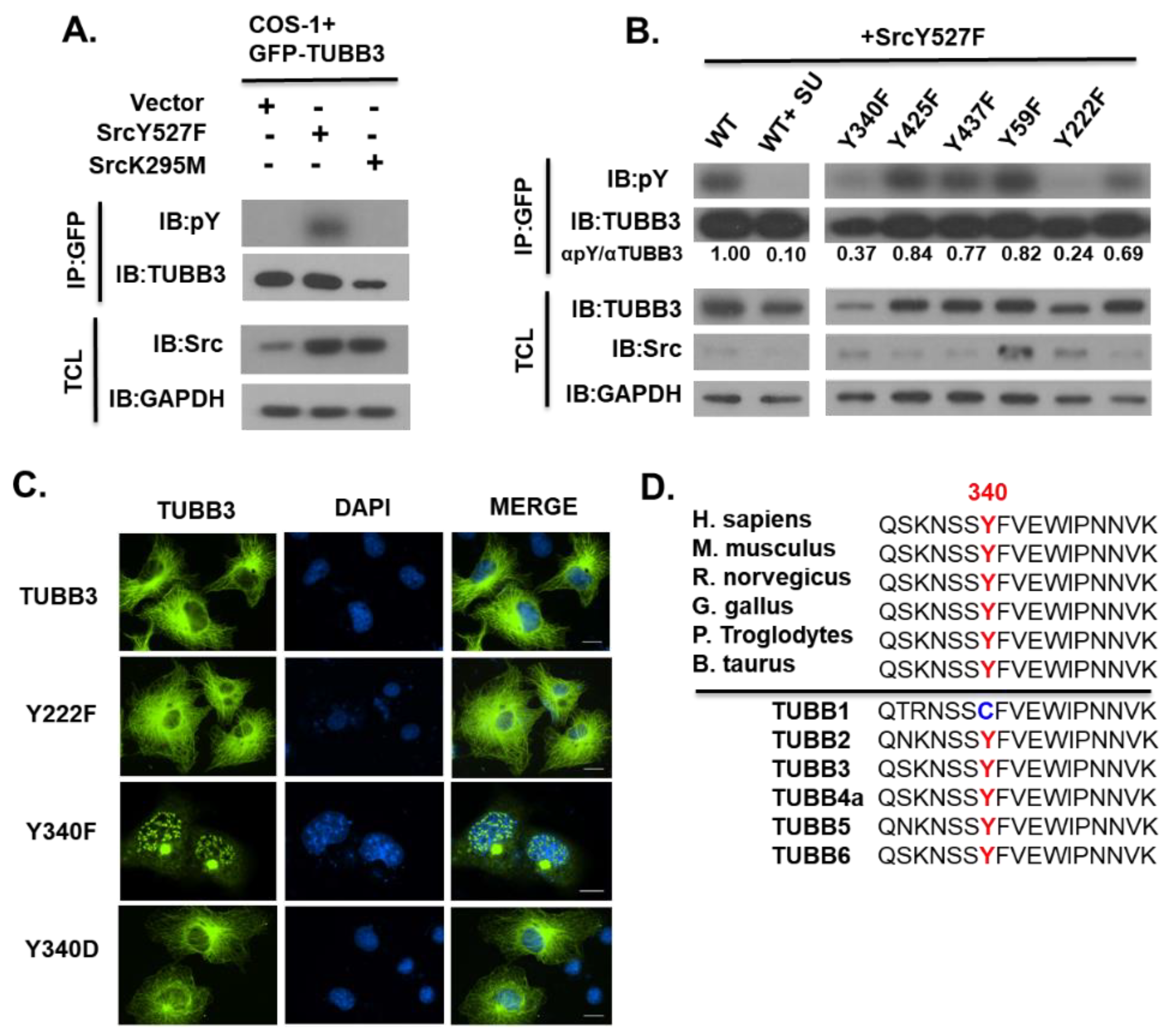
3.2. Identification of TUBB3 Phosphorylation Site
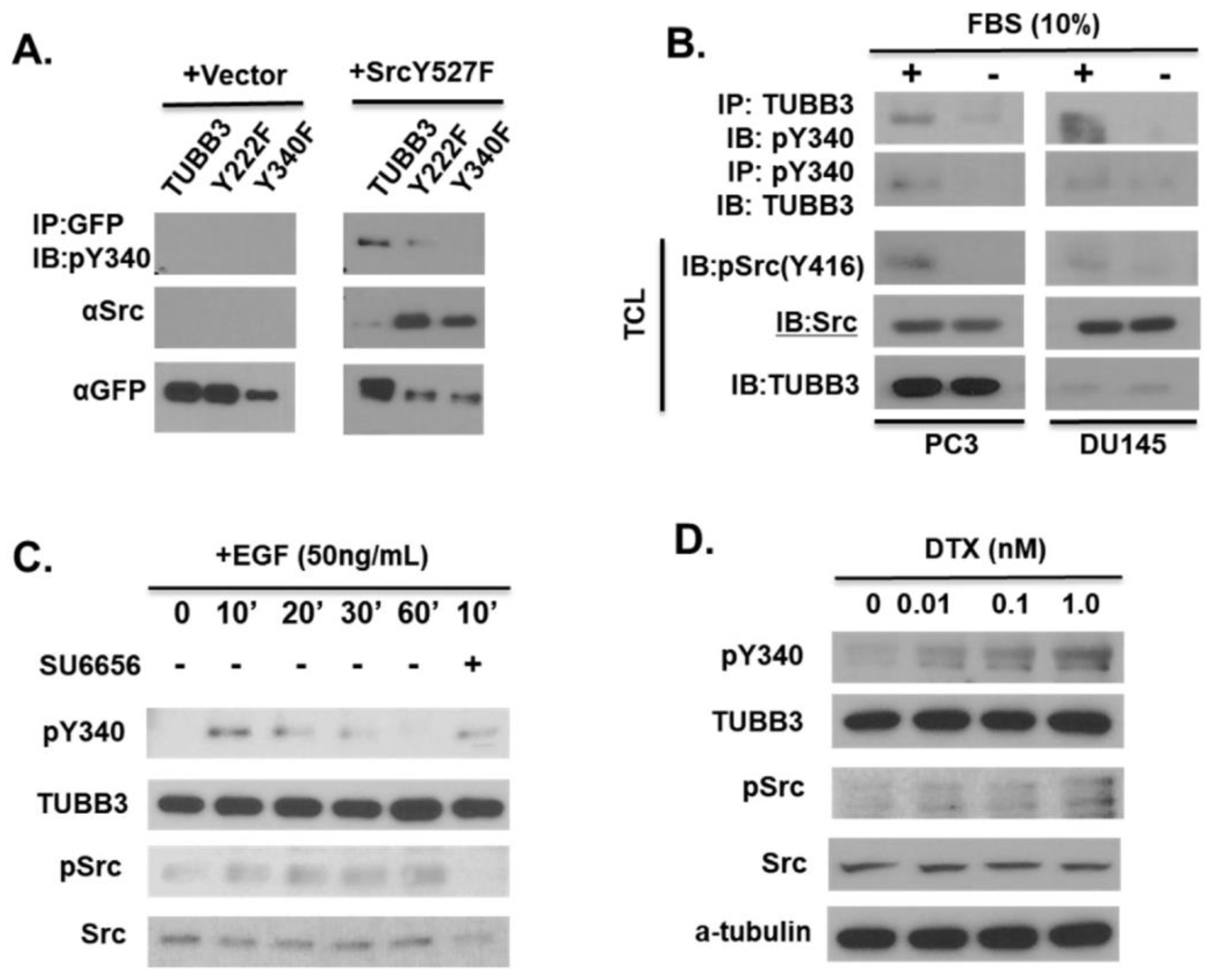
3.3. Modulation of TUBB3 Y340 Phosphorylation by SRC Kinase in Response to EGF and Docetaxel
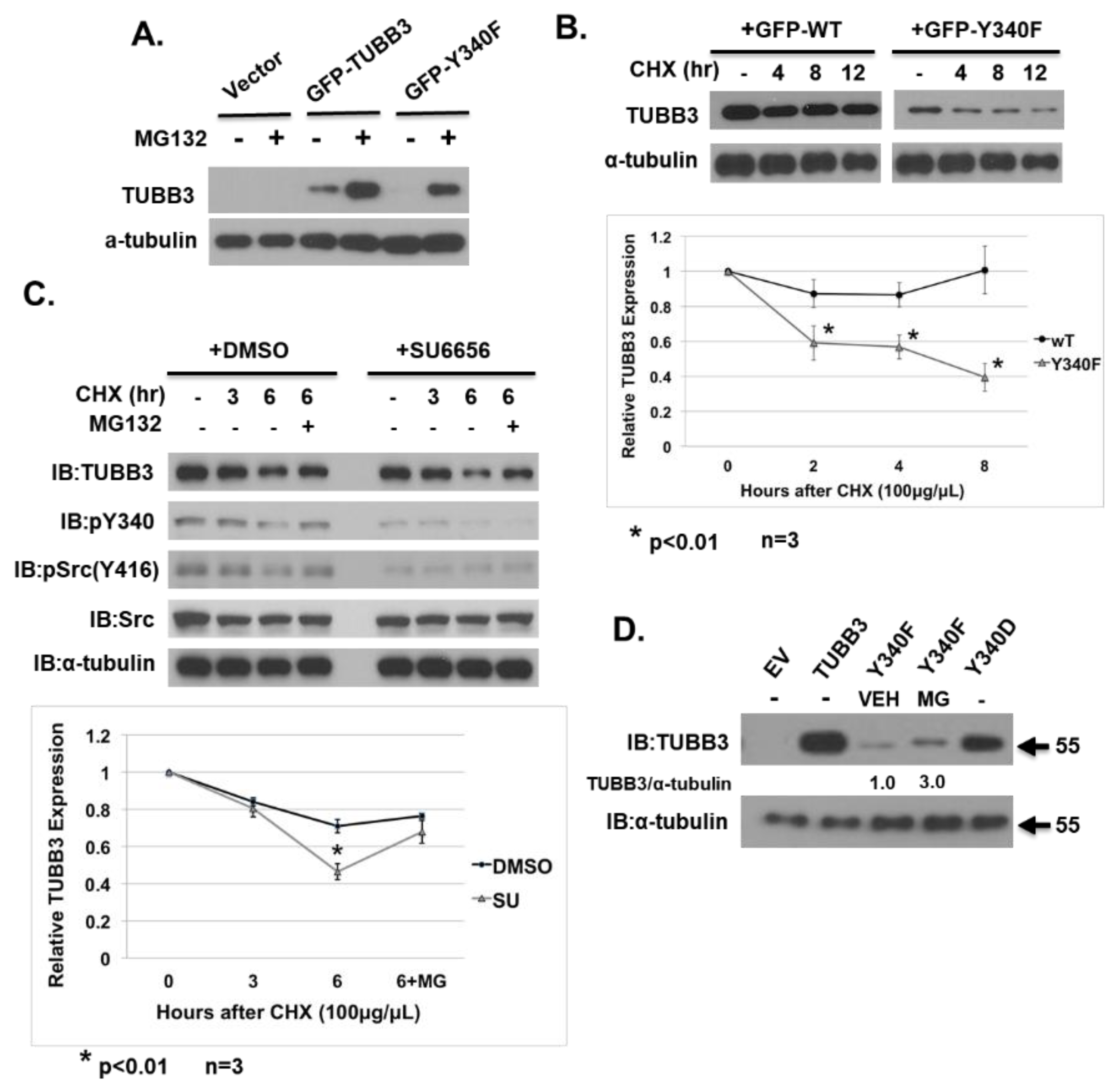
3.4. Phosphorylation of Y340 Stabilizes TUBB3 Protein
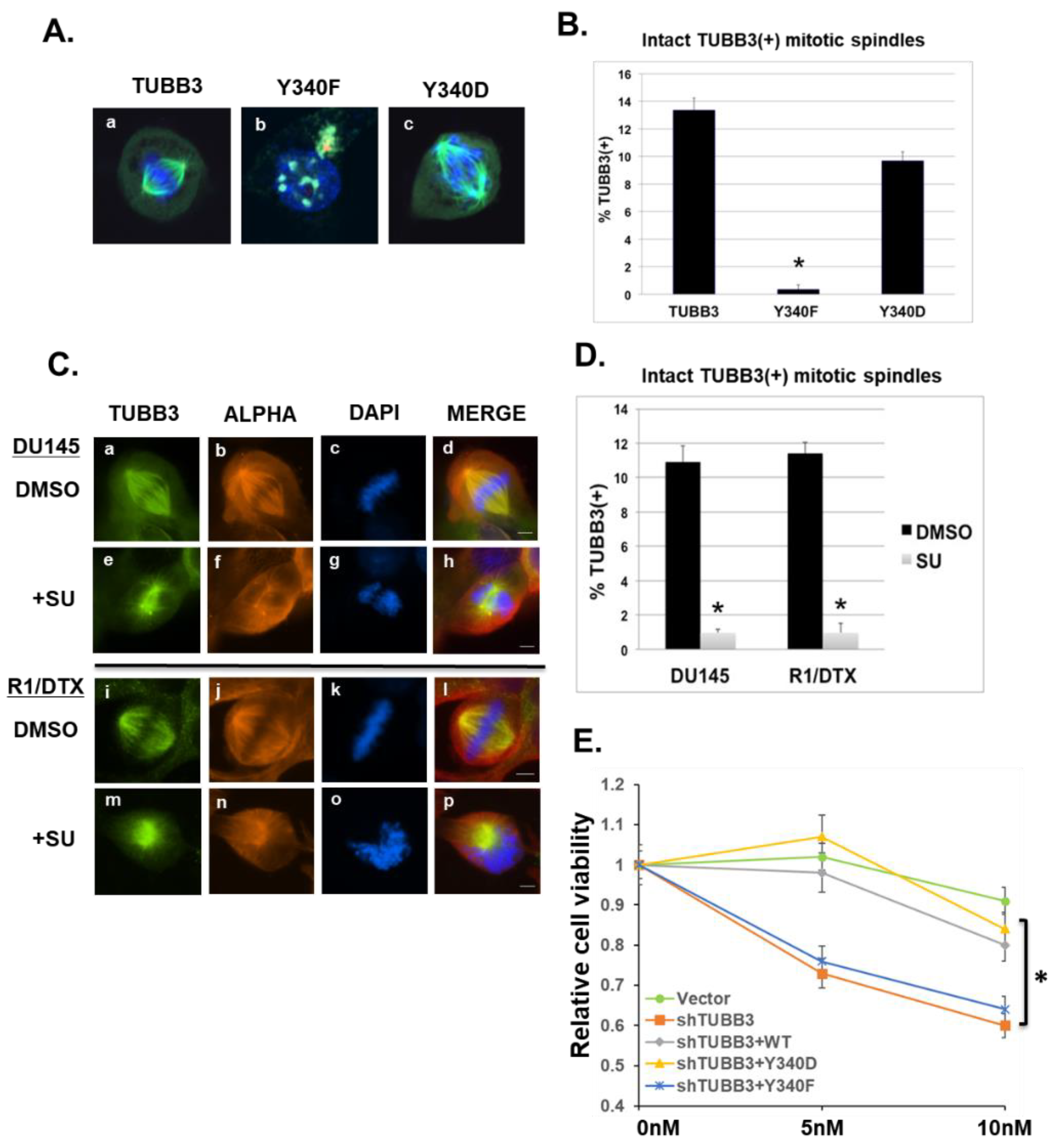
3.5. Effects of Y340 Phosphorylation on TUBB3 Subcellular Localization and Mitotic Spindle Stability
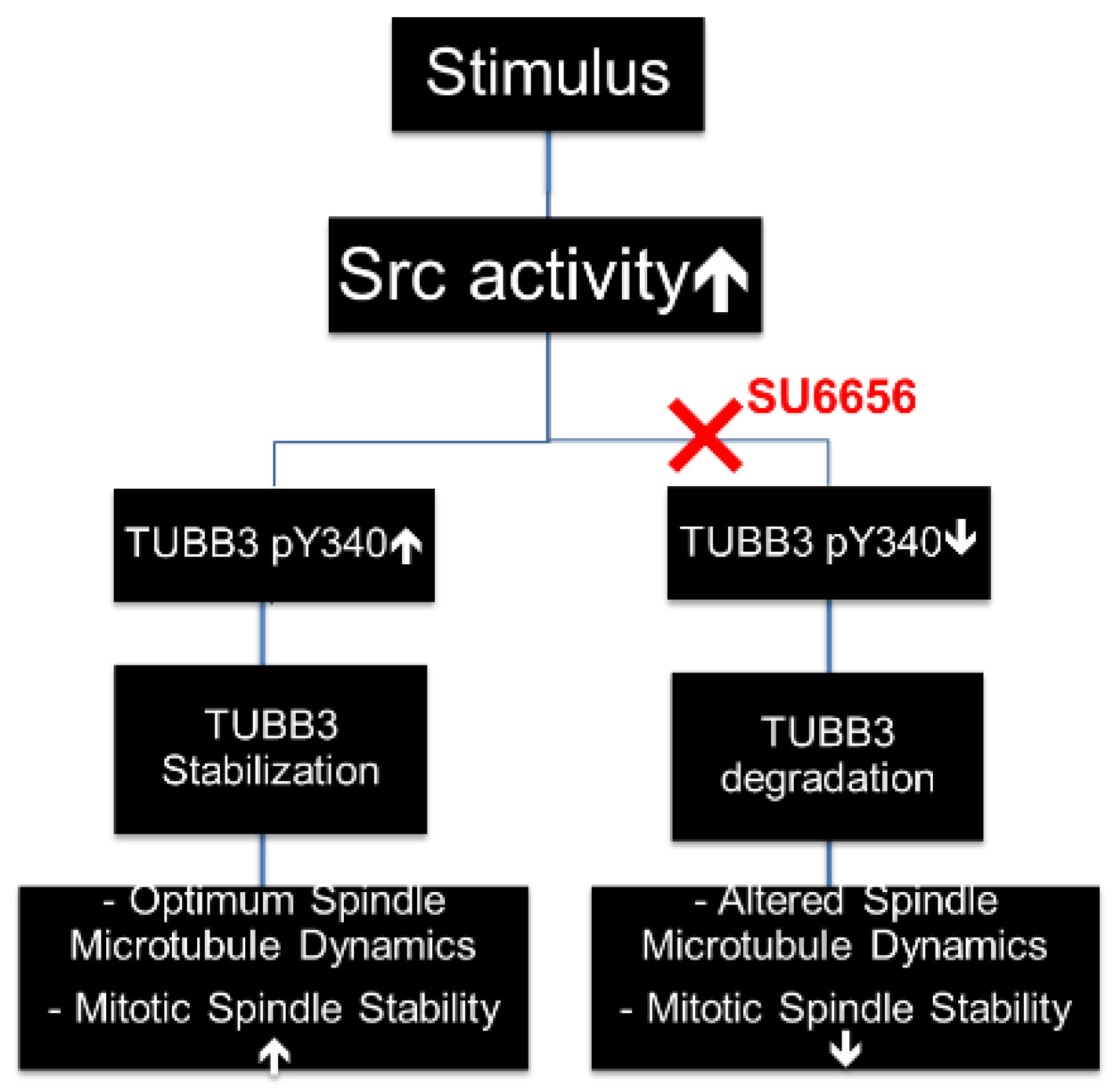
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cancer Facts & Figures 2013. American Cancer Society Website. 2013. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2013.html (accessed on 15 August 2015).
- Shen, M.; Shen, C. Molecular Genetics of Prostate Cancer. Genes Dev. 2010, 24, 1967–2000. [Google Scholar] [CrossRef] [Green Version]
- George, D.; Moul, J. Emerging Treatment Options for Patients with Castration-Resistant Prostate Cancer. Prostate 2012, 72, 338–349. [Google Scholar] [CrossRef]
- Shapiro, D.; Tareen, B. Current and Emerging Treatments in the Management of Castration-Resistant Prostate Cancer. Expert Rev. Anticancer Ther. 2012, 12, 951–964. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.C.; Song, L.; Haura, E.B. Src kinases as therapeutic targets for cancer. Nat. Rev. Clin. Oncol. 2009, 6, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Stehelin, D.; Varmus, H.E.; Bishop, J.M.; Vogt, P.K. DNA related to the transforming gene(s) of avian sarcoma viruses is present in normal avian DNA. Nature 1976, 260, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Yeatman, T.J. A Renaissance for SRC. Nat. Rev. Cancer 2004, 4, 470–480. [Google Scholar] [CrossRef]
- Chang, Y.M.; Kung, H.J.; Evans, C.P. Nonreceptor Tyrosine Kinases in Prostate Cancer. Neoplasia 2007, 9, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Fizazi, K. The role of Src in prostate cancer. Ann. Oncol. 2007, 18, 1765–1773. [Google Scholar] [CrossRef]
- Guarino, M. Src Signaing in Cancer Invasion. J. Cell. Physiol. 2010, 223, 14–26. [Google Scholar]
- Roche, S.; Fumagalli, S.; Courtneidge, S.A. Requirement for Src family protein-tyrosine kinases in G2 for fibroblast cell division. Science 1995, 269, 1567–1569. [Google Scholar] [CrossRef]
- Chackalaparampil, I.; Shalloway, D. Altered phosphorylation and activation of pp60c-Src during fibroblast mitosis. Cell 1988, 52, 695–701. [Google Scholar] [CrossRef]
- Moasser, M.M.; Srethapakdi, M.; Sachar, K.S.; Kraker, A.J.; Rosen, N. Inhibition of Src kinases by a selective inhibitor causes mitotic arrest. Cancer Res. 1999, 59, 6145–6152. [Google Scholar] [PubMed]
- Ng, M.M.; Chang, F.; Burgess, D.R. Movement of membrane domains and requirement of membrane signaling molecules for cytokinesis. Dev. Cell 2005, 9, 781–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasahara, K.; Nakayama, Y.; Nakazato, Y.; Ikeda, K.; Kuga, T.; Yamaguchi, N. Src signaling regulates completion of abscission in cytokinesis through ERK/MAPK activation at the midbody. J. Biol. Chem. 2007, 282, 5327–5339. [Google Scholar] [CrossRef] [Green Version]
- David-Pfeuty, T.; Bagrodia, S.; Shalloway, D. Differential localization patterns of myristoylated and non-myristoylated c-Src proteins in interphase and mitotic c-Src overexpressor cells. J. Cell Biol. 1993, 125, 639–649. [Google Scholar]
- Nakayama, Y.; Matsui, Y.; Takeda, Y.; Okamoto, M.; Abe, K.; Fukumoto, Y.; Yamaguchi, N. c-Src but not Fyn Promotes Proper Spindle Orientation in Early Prometaphase. J. Biol. Chem. 2012, 287, 24905–24915. [Google Scholar] [CrossRef] [Green Version]
- Kilpinen, S.; Autio, R.; Ojala, K.; Iljin, K.; Bucher, E.; Sara, H.; Pisto, T.; Saarela, M.; Skotheim, R.I.; Björkman, M.; et al. Systematic bioinformatics analysis of expression levels of 17,330 human genes across 9,783 samples from 175 types of healthy and pathological tissues. Genome Biol. 2008, 9, R139. [Google Scholar] [CrossRef] [Green Version]
- Jouhilahti, E.M.; Peltonen, S.; Peltonen, J. Class III β-tubulin Is a Component of the Mitotic Spindle in Multiple Cell Types. J. Histochem. Cytochem. 2008, 56, 1113–1119. [Google Scholar] [CrossRef] [Green Version]
- Tommasi, S.; Mangia, A.; Lacalamita, R.; Bellizzi, A.; Fedele, V.; Chiriatti, A.; Thomssen, C.; Kendzierski, N.; Latorre, A.; Lorusso, V.; et al. Cytoskeleton and paclitaxel sensitivity in breast cancer: The role of β-tubulins. Int. J. Cancer 2007, 120, 2078–2085. [Google Scholar] [CrossRef]
- Hetland, T.E.; Hellesylt, E.; Flørenes, V.A.; Tropé, C.; Davidson, B.; Kærn, J. Class III β-tubulin expression in advanced-stage serous ovarian carcinoma effusions is associated with poor survival and primary chemoresistance. Hum. Pathol. 2010, 42, 1019–1026. [Google Scholar] [CrossRef]
- Vilmar, A.C.; Sontoni-Rugiu, E.; Sørensen, J.B. Class III β-tubulin in Advanced NSCLC of Adenocarcinoma Subtype Predicts Superior Outcome in a Randomized Trial. Clin. Cancer Res. 2011, 17, 5205–5214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, Y.; Kuriyama, H.; Umezu, H.; Tanaka, J.; Yoshimasu, T.; Furukawa, T.; Tanaka, H.; Kagamu, H.; Gejyo, F.; Yoshizawa, H. Class III β-tubulin expression in Tumor Cells is Correlated with Resistance to Docetaxel in Patients with Completely Resected NSCLC. Intern. Med. 2009, 48, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seve, P.; Mackey, J.; Isaac, S.; Trédan, O.; Souquet, P.J.; Pérol, M.; Lai, R.; Voloch, A.; Dumontet, C. Class III β-tubulin expression in tumor cells predicts response and outcome in patients with NSCLC receiving paclitaxel. Mol. Cancer Ther. 2005, 4, 2001–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, S.; Ploussard, G.; Allory, Y.; Nicolaiew, N.; Boissiere-Michot, F.; Maille, P.; Kheuang, L.; Coppolani, E.; Ali, A.; Bibeau, F.; et al. Increased expression of class III β-tubulin in castration-resistant human prostate cancer. Br. J. Cancer 2009, 101, 951–956. [Google Scholar] [CrossRef] [Green Version]
- Ploussard, G.; Terry, S.; Maillé, P.; Allory, Y.; Sirab, N.; Kheuang, L.; Soyeux, P.; Nicolaiew, N.; Coppolani, E.; Paule, B.; et al. Class III β-tubulin Expression Predicts Prostate Tumor Aggressiveness and Patient Response to Docetaxel-Based Chemotherapy. Cancer Res. 2010, 70, 9253–9264. [Google Scholar] [CrossRef] [Green Version]
- Tsourlakis, M.C.; Weigand, P.; Grupp, K.; Kluth, M.; Steurer, S.; Schlomm, T.; Graefen, M.; Huland, H.; Salomon, G.; Steuber, T.; et al. β(III)-tubulin is an independent predictor of prostate cancer progression tightly linked to ERG fusion status and PTEN deletion. Am. J. Pathol. 2014, 184, 609–617. [Google Scholar] [CrossRef]
- Katsetos, C.D.; Dráberová, E.; Šmejkalová, B.; Reddy, G.; Bertrand, L.; de Chadarévian, J.P.; Legido, A.; Nissanov, J.; Baas, P.W.; Dráber, P. Class III β-tubulin and gamma-tubulin are co-expressed and form complexes in human glioblastoma cells. Neurochem. Res. 2007, 32, 1387–1398. [Google Scholar] [CrossRef]
- Oakley, B.R. An abundance of tubulins. Trends Cell Biol. 2000, 10, 537–542. [Google Scholar] [CrossRef]
- Shibazaki, M.; Maesawa, C.; Akasaka, K.; Kasai, S.; Yasuhira, S.; Kanno, K.; Nakayama, I.; Sugiyama, T.; Wakabayasi, G.; Masuda, T.; et al. Transcriptional and post-transcriptional regulation of β(III)-tubulin protein expression in relation with cell cycle-dependent regulation of tumor cells. Int. J. Oncol. 2012, 40, 695–702. [Google Scholar]
- Narvi, E.; Jaakkola, K.; Winsel, S.; Oetken-Lindholm, C.; Halonen, P.; Kallio, L.; Kallio, M.J. Altered TUBB3 expression contributes to the epothilone response of mitotic cells. Br. J. Cancer 2013, 108, 82–90. [Google Scholar] [CrossRef] [Green Version]
- McCarroll, J.A.; Gan, P.P.; Liu, M.; Kavallaris, M. β(III)-tubulin is a Multifunctional Protein Involved in Drug Sensitivity and Tumorigenesis in Non-Small Cell Lung Cancer. Cancer Res. 2010, 70, 4995–5003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavallaris, M.; Kuo, D.Y.; Burkhart, C.A.; Regl, D.L.; Norris, M.D.; Haber, M.; Horwitz, S.B. Taxol-resistant epithelial ovarian tumors are associated with altered expression of specific β-tubulin isotypes. J. Clin. Investig. 1997, 100, 1282–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, P.P.; Pasquier, E.; Kavallaris, M. Class III β-tubulin Mediates Sensitivity to Chemotherapeutic Drugs in Non-Small Cell Lung Cancer. Cancer Res. 2007, 67, 9356–9363. [Google Scholar] [CrossRef] [Green Version]
- Gan, P.P.; McCarroll, J.A.; Po’uha, S.T.; Kamath, K.; Jordan, M.A.; Kavallaris, M. Microtubule Dynamics, Mitotic Arrest, and Apoptosis: Drug-Induced Differential Effects of β(III)-tubulin. Mol. Cancer Ther. 2010, 9, 1339–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdier-Pinard, P.; Pasquier, E.; Xiao, H.; Burd, B.; Villard, C.; Lafitte, D.; Miller, L.M.; Angeletti, R.H.; Horwitz, S.B.; Braguer, D. Tubulin proteomics: Towards breaking the code. Anal. Biochem. 2009, 384, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavallaris, M. Microtubules and resistance to tubulin-binding agents. Nat. Rev. Cancer 2010, 10, 194–204. [Google Scholar] [CrossRef]
- Verhey, K.; Gaertig, J. The Tubulin Code. Cell Cycle 2007, 6, 2152–2160. [Google Scholar] [CrossRef]
- Wloga, D.; Gaertig, J. Post-translational modifications of microtubules. J. Cell Sci. 2010, 123, 3447–3455. [Google Scholar] [CrossRef] [Green Version]
- Janke, C. The tubulin code: Molecular components, readout mechanisms, and functions. J. Cell Biol. 2014, 206, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Matten, W.T.; Aubry, M.; West, J. Tubulin is phosphorylated at tyrosine by pp60c-Src in nerve growth cone membranes. J. Cell Biol. 1990, 111, 1959–1979. [Google Scholar] [CrossRef]
- Katagiri, K.; Katagiri, T.; Kajiyama, K.; Yamamoto, T.; Yoshida, T. Tyrosine-Phosphorylation of Tubulin during Monocytic Differentiation of HL-60 cells. J. Immunol. 1993, 150, 585–593. [Google Scholar] [PubMed]
- Parker, A.L.; Kavallaris, M.; McCarroll, J.A. Microtubules and their role in cellular stress in cancer. Front. Oncol. 2014, 4, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Pengetnze, Y.; Taylor, C.C. Src inhibition enhances paclitaxel cytotoxicity in ovarian cancer cells by caspase-9-independent activation of caspase-3. Mol. Cancer Ther. 2005, 4, 217–224. [Google Scholar] [PubMed]
- Le, X.F.; Bast, R.C. Src family kinases and paclitaxel sensitivity. Cancer Biol. Ther. 2011, 12, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Levi, M.; Maro, B.; Shalgi, R. The involvement of Fyn kinase in resumption of the first meiotic division in mouse oocytes. Cell Cycle 2010, 9, 1577–1589. [Google Scholar] [CrossRef] [Green Version]
- Levi, M.; Shalgi, R. The role of Fyn kinase in the release from metaphase in mammalian oocytes. Mol. Cell. Endocrinol. 2010, 314, 228–233. [Google Scholar] [CrossRef]
- Linn, D.E.; Yang, X.; Sun, F.; Xie, Y.; Chen, H.; Jiang, R.; Chen, H.; Chumsri, S.; Burger, A.M.; Qiu, Y. A Role for OCT4 in Tumor Initiation of Drug-Resistant Prostate Cancer Cells. Genes Cancer 2010, 1, 908–916. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Xu, K.; Dai, B.; Guo, Z.; Jiang, T.; Chen, H.; Qiu, Y. The 44kDa Pim-1 kinase directly interacts with tyrosine kinase Etk/BMX and protects human prostate cancer cells from apoptosis induced by chemotherapeutic drugs. Oncogene 2006, 25, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Xu, K.; Linn, D.E.; Yang, X.; Guo, Z.; Shimelis, H.; Nakanishi, T.; Ross, D.D.; Chen, H.; Fazli, L.; et al. The 44-kDa Pim-1 Kinase Phosphorylates BCRP/ABCG2 and Thereby Promotes Its Multimerization and Drug-resistant Activity. J. Biol. Chem. 2008, 283, 3349–3356. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Guo, Z.; Sun, F.; Li, W.; Alfano, A.; Shimelis, H.; Chen, M.; Brodie, A.M.; Chen, H.; Xiao, Z.; et al. Novel Membrane-associated Androgen Receptor Splice Variant Potentiates Proliferative and Survival Responses in Prosate Cancer Cells. J. Biol. Chem. 2011, 286, 36152–36160. [Google Scholar] [CrossRef] [Green Version]
- Ertych, N.; Stolz, A.; Stenzinger, A.; Weichert, W.; Kaulfuß, S.; Burfeind, P.; Aigner, A.; Wordeman, L.; Bastians, H. Increased microtubule assembly rates influence chromosomal instability in colorectal cancer cells. Nat. Cell Biol. 2014, 16, 779–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfano, A.; Xu, J.; Yang, X.; Deshmukh, D.; Qiu, Y. SRC Kinase-Mediated Tyrosine Phosphorylation of TUBB3 Regulates Its Stability and Mitotic Spindle Dynamics in Prostate Cancer Cells. Pharmaceutics 2022, 14, 932. https://doi.org/10.3390/pharmaceutics14050932
Alfano A, Xu J, Yang X, Deshmukh D, Qiu Y. SRC Kinase-Mediated Tyrosine Phosphorylation of TUBB3 Regulates Its Stability and Mitotic Spindle Dynamics in Prostate Cancer Cells. Pharmaceutics. 2022; 14(5):932. https://doi.org/10.3390/pharmaceutics14050932
Chicago/Turabian StyleAlfano, Alan, Jin Xu, Xi Yang, Dhanraj Deshmukh, and Yun Qiu. 2022. "SRC Kinase-Mediated Tyrosine Phosphorylation of TUBB3 Regulates Its Stability and Mitotic Spindle Dynamics in Prostate Cancer Cells" Pharmaceutics 14, no. 5: 932. https://doi.org/10.3390/pharmaceutics14050932