
Therapeutic Effects of Synthetic Triblock Amphiphilic Short Antimicrobial Peptides on Human Lung Adenocarcinoma

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. CCK-8 Analysis
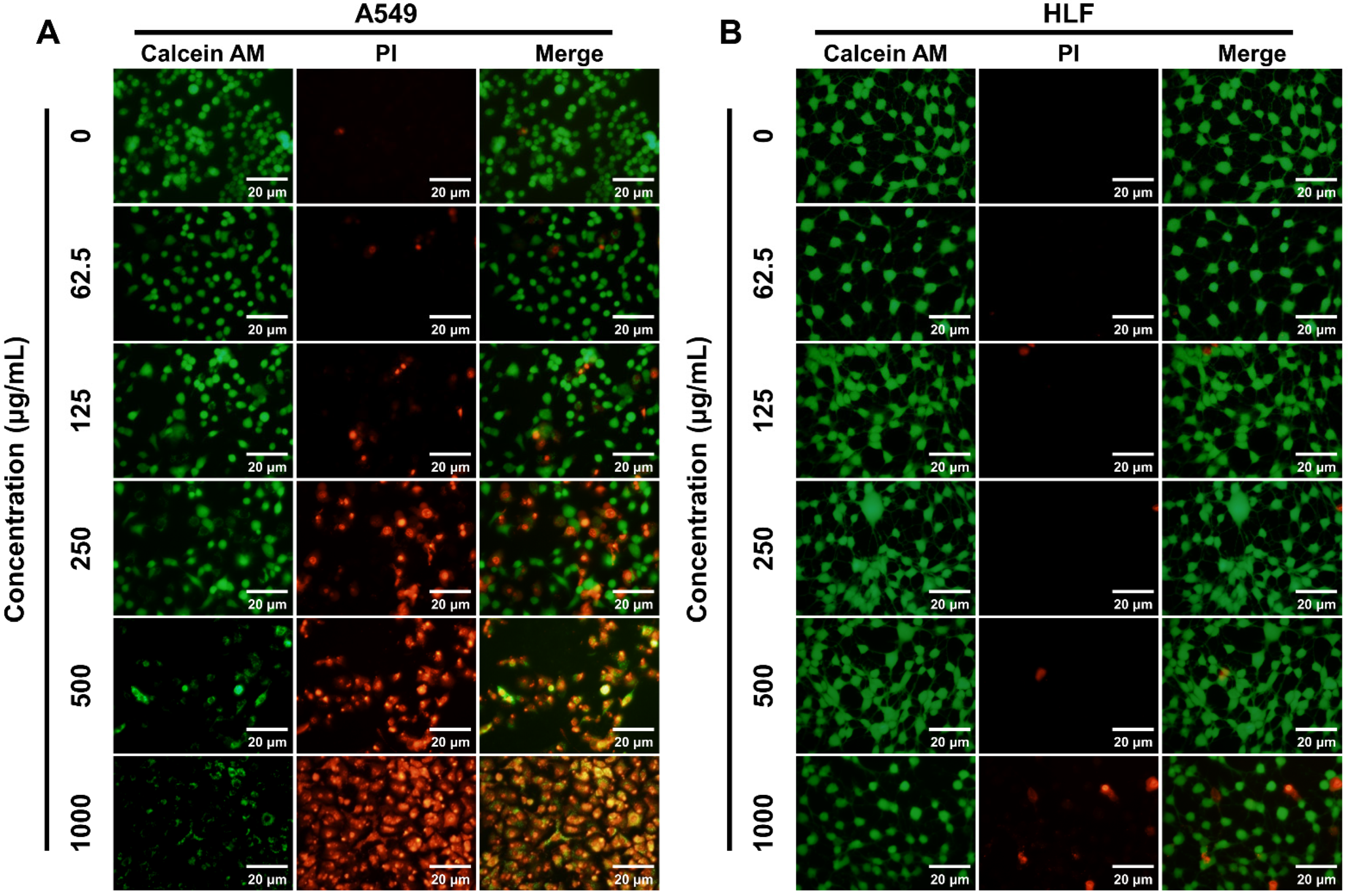
2.4. Live/Dead Staining of Cells
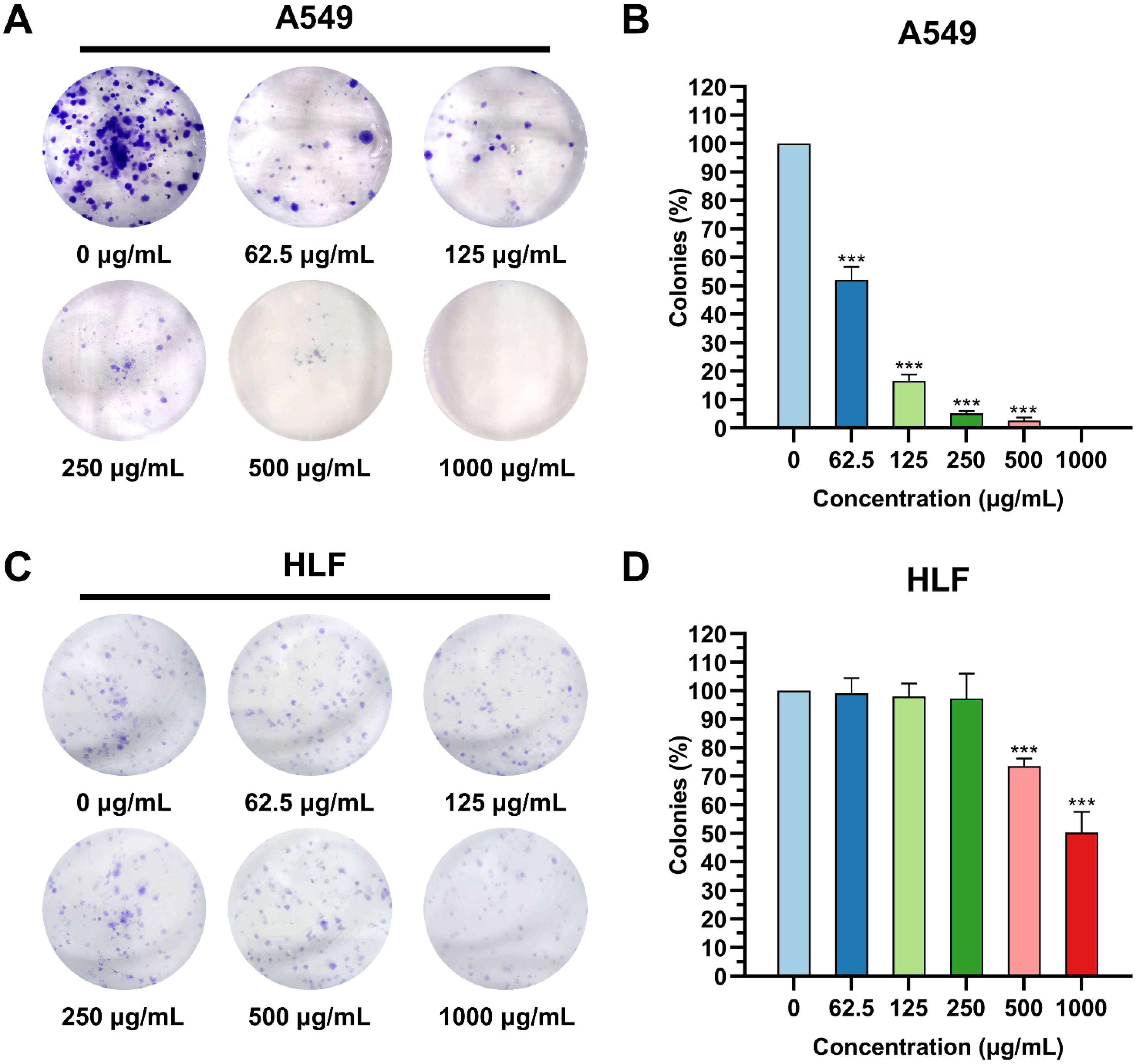
2.5. Colony Formation Assay
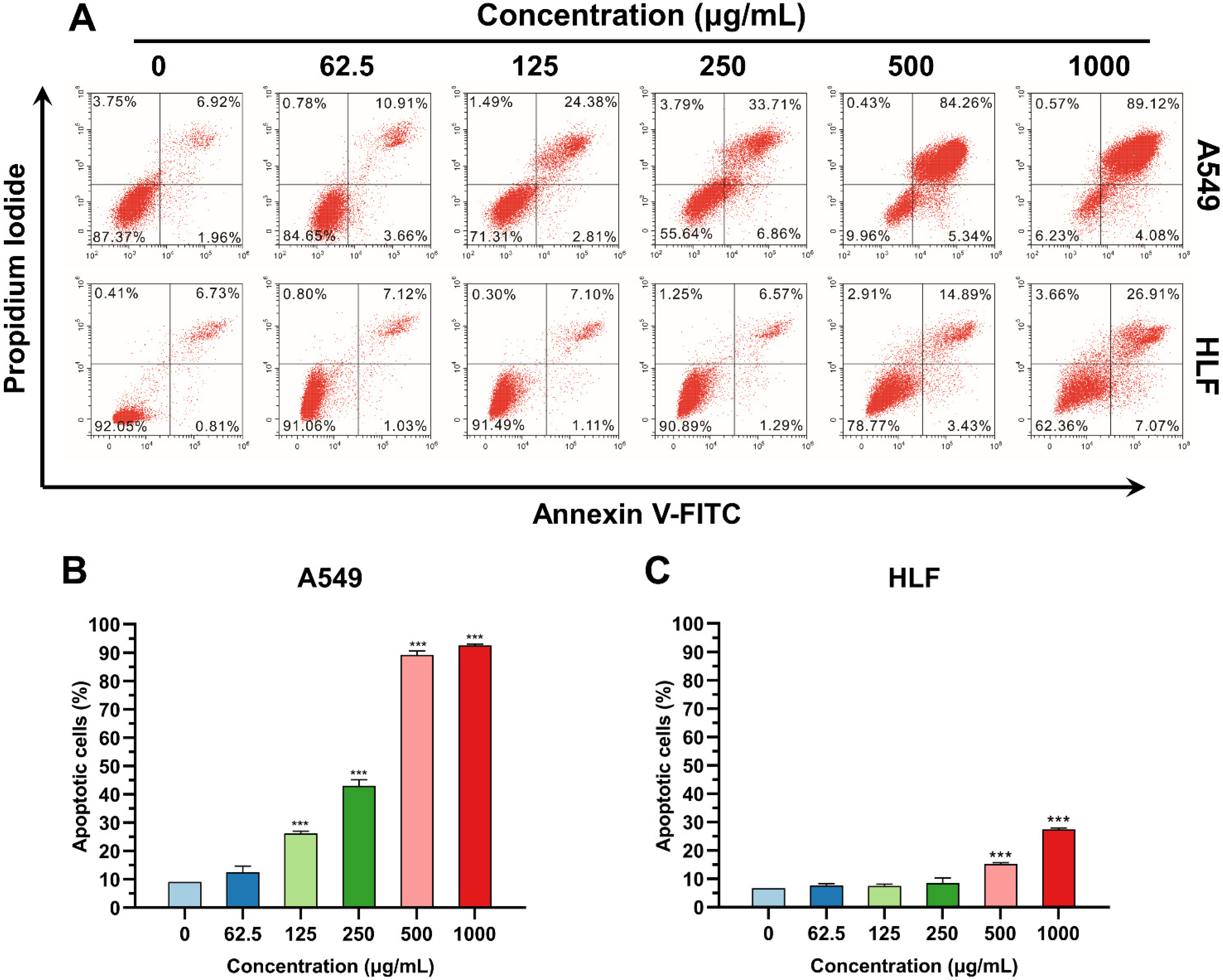
2.6. Apoptosis Analysis
2.7. Membrane Staining and Confocal Imaging
2.8. Scanning Electron Microscopy (SEM)
2.9. Xenograft Tumor Model
2.10. Statistical Analysis
3. Results
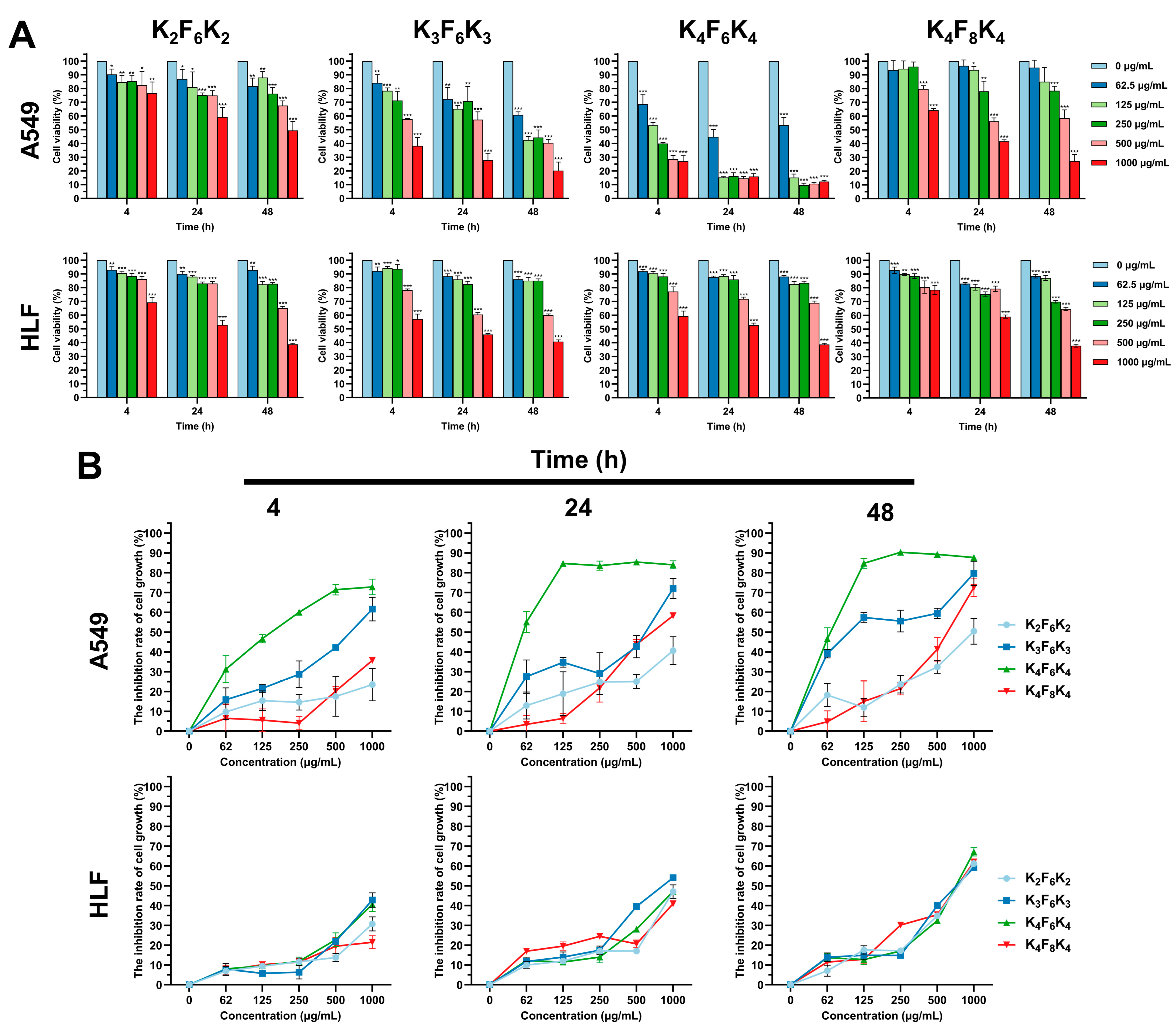
3.1. Preferential Cytotoxicity of K4F6K4 against Tumor Cells by CCK-8 and Live/Dead Assays
3.2. K4F6K4 Inhibited the Colony Formation of Lung Cancer Cells
3.3. K4F6K4 Induced Lung Cancer Cell Apoptosis
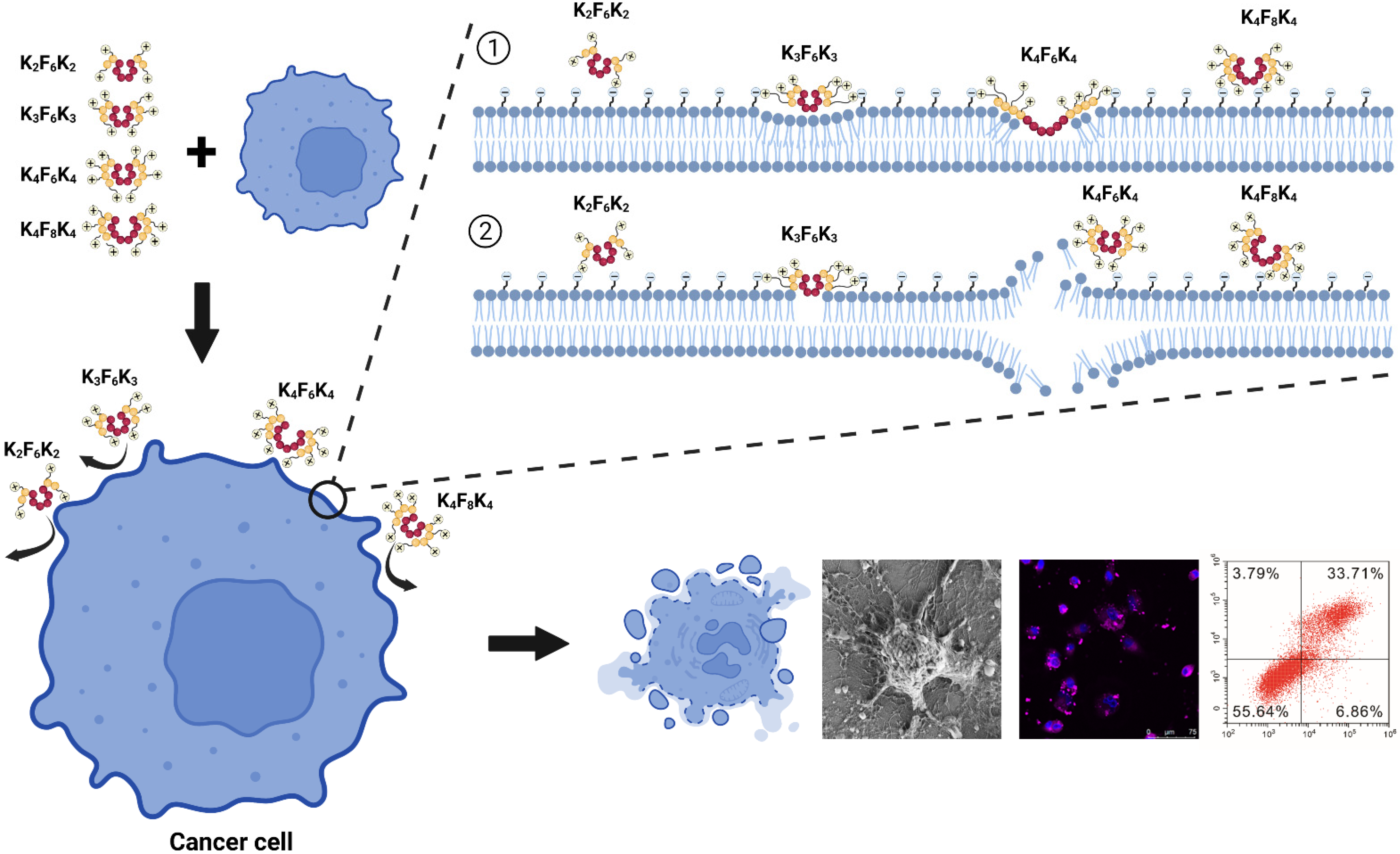
3.4. K4F6K4 Noticeably Disrupted A549 Cell Membrane Integrity and Induced Morphological Modifications
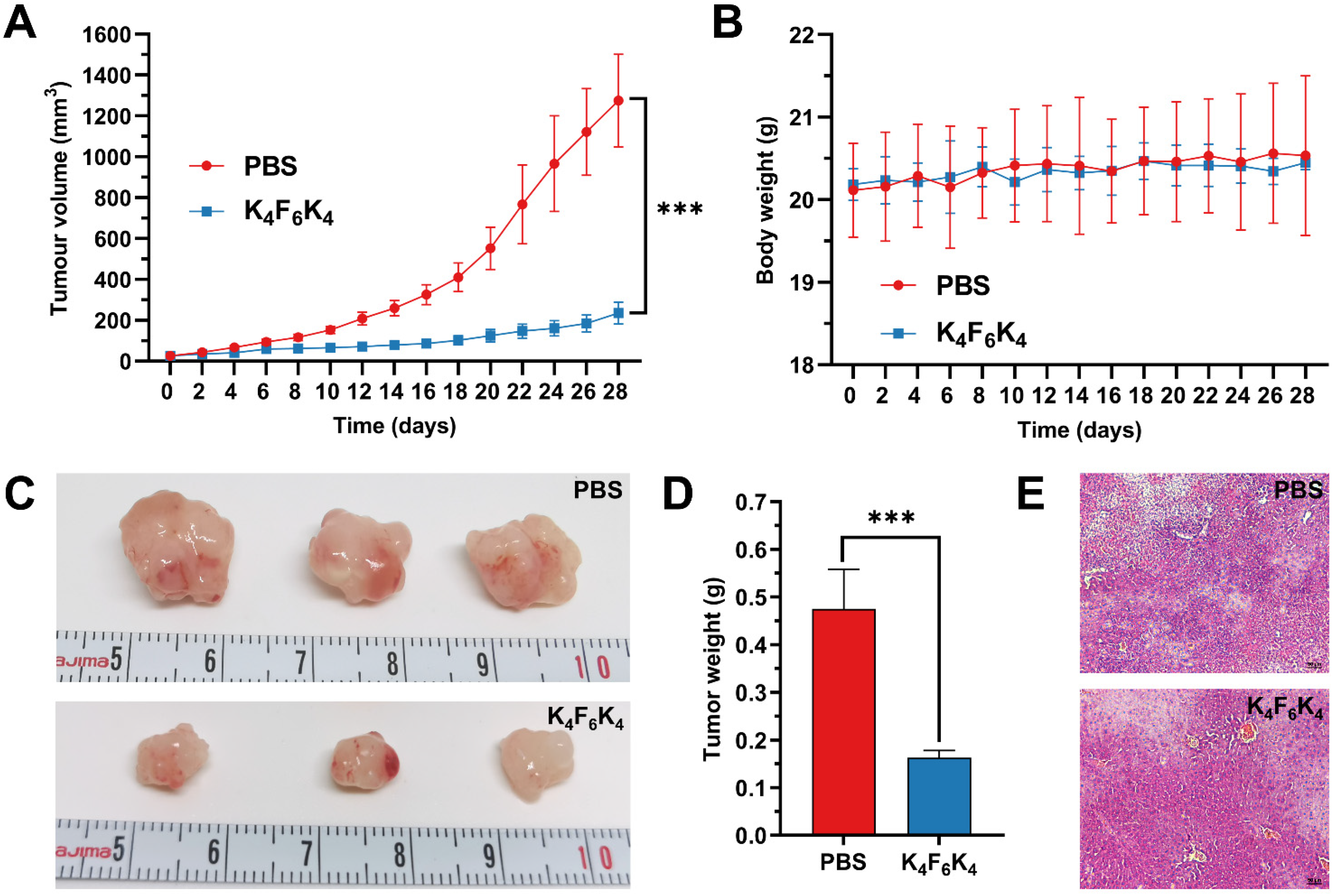
3.5. K4F6K4 Significantly Inhibited Tumor Growth in Xenografted Mice without Measurable Side Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; DeSantis, C.; Virgo, K.; Stein, K.; Mariotto, A.; Smith, T.; Cooper, D.; Gansler, T.; Lerro, C.; Fedewa, S.; et al. Cancer treatment and survivorship statistics, 2012. CA Cancer J. Clin. 2012, 62, 220–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Qian, L.; Yang, J.; Huang, H.; Feng, J.; Li, X.; Bian, T.; Ke, H.; Liu, J.; Zhang, J. The expression level and prognostic value of HIPK3 among non-small-cell lung cancer patients in China. OncoTargets Ther. 2018, 11, 7459–7469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, G.; Wang, W.; Chai, K.; Yang, S.; Li, F.; Jiang, K. Combination treatment with triptolide and hydroxycamptothecin synergistically enhances apoptosis in A549 lung adenocarcinoma cells through PP2A-regulated ERK, p38 MAPKs and Akt signaling pathways. Int. J. Oncol. 2015, 46, 1007–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doroshow, D.B.; Sanmamed, M.F.; Hastings, K.; Politi, K.; Rimm, D.L.; Chen, L.; Melero, I.; Schalper, K.A.; Herbst, R.S. Immunotherapy in Non-Small Cell Lung Cancer: Facts and Hopes. Clin. Cancer Res. 2019, 25, 4592–4602. [Google Scholar] [CrossRef] [Green Version]
- Peng, D.H.; Rodriguez, B.L.; Diao, L.; Gaudreau, P.O.; Padhye, A.; Konen, J.M.; Ochieng, J.K.; Class, C.A.; Fradette, J.J.; Gibson, L.; et al. Th17 cells contribute to combination MEK inhibitor and anti-PD-L1 therapy resistance in KRAS/p53 mutant lung cancers. Nat. Commun. 2021, 12, 2606. [Google Scholar] [CrossRef]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The biology and management of non-small cell lung cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef]
- Hastings, K.; Yu, H.A.; Wei, W.; Sanchez-Vega, F.; DeVeaux, M.; Choi, J.; Rizvi, H.; Lisberg, A.; Truini, A.; Lydon, C.A.; et al. EGFR mutation subtypes and response to immune checkpoint blockade treatment in non-small-cell lung cancer. Ann. Oncol. 2019, 30, 1311–1320. [Google Scholar] [CrossRef] [Green Version]
- Thress, K.S.; Paweletz, C.P.; Felip, E.; Cho, B.C.; Stetson, D.; Dougherty, B.; Lai, Z.; Markovets, A.; Vivancos, A.; Kuang, Y.; et al. Acquired EGFR C797S mutation mediates resistance to AZD9291 in non-small cell lung cancer harboring EGFR T790M. Nat. Med. 2015, 21, 560–562. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Landis, R.F.; Li, C.H.; Schnurr, M.; Das, R.; Lee, Y.W.; Yazdani, M.; Liu, Y.; Kozlova, A.; Rotello, V.M. Engineered Polymer Nanoparticles with Unprecedented Antimicrobial Efficacy and Therapeutic Indices against Multidrug-Resistant Bacteria and Biofilms. J. Am. Chem. Soc. 2018, 140, 12137–12143. [Google Scholar] [CrossRef]
- Nicola, A.M.; Albuquerque, P.; Paes, H.C.; Fernandes, L.; Costa, F.F.; Kioshima, E.S.; Abadio, A.K.R.; Bocca, A.L.; Felipe, M.S. Antifungal drugs: New insights in research & development. Pharmacol. Ther. 2019, 195, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, A.; Skrygan, M.; Gambichler, T.; Brockmeyer, N.H.; Stucker, M.; Herzler, C.; Potthoff, A.; Altmeyer, P.; Pfister, H.; Wieland, U. Human papillomavirus-associated induction of human beta-defensins in anal intraepithelial neoplasia. Br. J. Dermatol. 2009, 160, 1197–1205. [Google Scholar] [CrossRef] [PubMed]
- Wessely-Szponder, J.; Zdziennicka, J.; Junkuszew, A.; Latalski, M.; Swieca, M.; Szponder, T. Prospects and Applications of Natural Blood-Derived Products in Regenerative Medicine. Int. J. Mol. Sci. 2021, 23, 472. [Google Scholar] [CrossRef]
- Prats-Ejarque, G.; Li, J.; Ait-Ichou, F.; Lorente, H.; Boix, E. Testing a Human Antimicrobial RNase Chimera against Bacterial Resistance. Front. Microbiol. 2019, 10, 1357. [Google Scholar] [CrossRef]
- Tang, Q.; Wang, Q.; Sun, Z.; Kang, S.; Fan, Y.; Hao, Z. Bergenin Monohydrate Attenuates Inflammatory Response via MAPK and NF-kappaB Pathways against Klebsiella pneumonia Infection. Front. Pharmacol. 2021, 12, 651664. [Google Scholar] [CrossRef]
- Bao, P.; Paterson, D.A.; Harrison, P.L.; Miller, K.; Peyman, S.; Jones, J.C.; Sandoe, J.; Evans, S.D.; Bushby, R.J.; Gleeson, H.F. Lipid coated liquid crystal droplets for the on-chip detection of antimicrobial peptides. Lab Chip 2019, 19, 1082–1089. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.W.; Lee, E.Y.; Ferguson, A.L.; Wong, G.C.L. Machine learning antimicrobial peptide sequences: Some surprising variations on the theme of amphiphilic assembly. Curr. Opin. Colloid Interface Sci. 2018, 38, 204–213. [Google Scholar] [CrossRef]
- Li, D.; Beisswenger, C.; Herr, C.; Schmid, R.M.; Gallo, R.L.; Han, G.; Zakharkina, T.; Bals, R. Expression of the antimicrobial peptide cathelicidin in myeloid cells is required for lung tumor growth. Oncogene 2014, 33, 2709–2716. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Bandari, R.P.; Lee, L.; Li, R.; Yu, P.; Smith, C.J.; Ma, L. Design, Synthesis, and In Vitro and In Vivo Evaluation of High Affinity and Specificity Near-Infrared Fluorescent Bombesin Antagonists for Tumor Imaging. J. Med. Chem. 2018, 61, 7657–7670. [Google Scholar] [CrossRef]
- Tanhaieian, A.; Sekhavati, M.H.; Ahmadi, F.S.; Mamarabadi, M. Heterologous expression of a broad-spectrum chimeric antimicrobial peptide in Lactococcus lactis: Its safety and molecular modeling evaluation. Microb. Pathog. 2018, 125, 51–59. [Google Scholar] [CrossRef]
- Guilhelmelli, F.; Vilela, N.; Albuquerque, P.; Derengowski Lda, S.; Silva-Pereira, I.; Kyaw, C.M. Antibiotic development challenges: The various mechanisms of action of antimicrobial peptides and of bacterial resistance. Front. Microbiol. 2013, 4, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, J.C.; Lai, R.W.; Woodhead, J.S.T.; Joly, J.H.; Mitchell, C.J.; Cameron-Smith, D.; Lu, R.; Cohen, P.; Graham, N.A.; Benayoun, B.A.; et al. MOTS-c is an exercise-induced mitochondrial-encoded regulator of age-dependent physical decline and muscle homeostasis. Nat. Commun. 2021, 12, 470. [Google Scholar] [CrossRef] [PubMed]
- Grafskaia, E.; Pavlova, E.; Babenko, V.V.; Latsis, I.; Malakhova, M.; Lavrenova, V.; Bashkirov, P.; Belousov, D.; Klinov, D.; Lazarev, V. The Hirudo Medicinalis Microbiome Is a Source of New Antimicrobial Peptides. Int J. Mol. Sci. 2020, 21, 7141. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.Q.; Qian, Y.S.; Fu, X.J.; Huang, S.T.; Li, Y.; Zhou, C.C. De Novo Design of Triblock Amphiphilic Short Antimicrobial Peptides. ACS Appl. Polym. Mater. 2020, 2, 3988–3992. [Google Scholar] [CrossRef]
- Lin, L.; Chi, J.; Yan, Y.; Luo, R.; Feng, X.; Zheng, Y.; Xian, D.; Li, X.; Quan, G.; Liu, D.; et al. Membrane-disruptive peptides/peptidomimetics-based therapeutics: Promising systems to combat bacteria and cancer in the drug-resistant era. Acta Pharm. Sin. B 2021, 11, 2609–2644. [Google Scholar] [CrossRef]
- Martin, S.J.; Reutelingsperger, C.P.; McGahon, A.J.; Rader, J.A.; van Schie, R.C.; LaFace, D.M.; Green, D.R. Early redistribution of plasma membrane phosphatidylserine is a general feature of apoptosis regardless of the initiating stimulus: Inhibition by overexpression of Bcl-2 and Abl. J. Exp. Med. 1995, 182, 1545–1556. [Google Scholar] [CrossRef] [Green Version]
- Fadok, V.A.; Voelker, D.R.; Campbell, P.A.; Cohen, J.J.; Bratton, D.L.; Henson, P.M. Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages. J. Immunol. 1992, 148, 2207–2216. [Google Scholar]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: Prospect for anticancer applications. Oncotarget 2017, 8, 46635–46651. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Lai, Y.; Wang, Y.; Sun, X.; Mo, C.; Wang, J.; Wu, Y.; Li, J.; Chang, S.; Che, G. Comprehensive Pulmonary Rehabilitation is an Effective Way for Better Postoperative Outcomes in Surgical Lung Cancer Patients with Risk Factors: A Propensity Score-Matched Retrospective Cohort Study. Cancer Manag. Res. 2020, 12, 8903–8912. [Google Scholar] [CrossRef]
- Fani, S.; Kamalidehghan, B.; Lo, K.M.; Hashim, N.M.; Chow, K.M.; Ahmadipour, F. Synthesis, structural characterization, and anticancer activity of a monobenzyltin compound against MCF-7 breast cancer cells. Drug Des. Dev. Ther. 2015, 9, 6191–6201. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Shi, D.; Zhong, R.; Ye, Z.; Ma, C.; Zhou, M.; Xi, X.; Wang, L.; Chen, T.; Kwok, H.F. Bioevaluation of Ranatuerin-2Pb from the Frog Skin Secretion of Rana pipiens and its Truncated Analogues. Biomolecules 2019, 9, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mader, J.S.; Hoskin, D.W. Cationic antimicrobial peptides as novel cytotoxic agents for cancer treatment. Expert Opin. Investig. Drugs 2006, 15, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Lv, D.; Wang, X.; Wang, Y.; Hou, C.; Gao, K.; Guo, X. Inhibitory effects of Bombyx mori antimicrobial peptide cecropins on esophageal cancer cells. Eur. J. Pharmacol. 2020, 887, 173434. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Tay, J.; Hedrick, J.; Yang, Y.Y. Synthetic macromolecules as therapeutics that overcome resistance in cancer and microbial infection. Biomaterials 2020, 252, 120078. [Google Scholar] [CrossRef]
- Gaspar, D.; Veiga, A.S.; Castanho, M.A. From antimicrobial to anticancer peptides. A review. Front. Microbiol. 2013, 4, 294. [Google Scholar] [CrossRef] [Green Version]
- Di Bonaventura, I.; Jin, X.; Visini, R.; Probst, D.; Javor, S.; Gan, B.H.; Michaud, G.; Natalello, A.; Doglia, S.M.; Kohler, T.; et al. Chemical space guided discovery of antimicrobial bridged bicyclic peptides against Pseudomonas aeruginosa and its biofilms. Chem. Sci. 2017, 8, 6784–6798. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.M.; Edwards, M.A.; Li, J.; Yip, C.M.; Deber, C.M. Roles of hydrophobicity and charge distribution of cationic antimicrobial peptides in peptide-membrane interactions. J. Biol. Chem. 2012, 287, 7738–7745. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Huang, Y.; Feng, Q.; Chen, Y. Tryptophan as a probe to study the anticancer mechanism of action and specificity of alpha-helical anticancer peptides. Molecules 2014, 19, 12224–12241. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.; Wong, S.W.; Ge, L.; Shaw, C.; Siu, S.W.I.; Kwok, H.F. In Vitro and MD Simulation Study to Explore Physicochemical Parameters for Antibacterial Peptide to Become Potent Anticancer Peptide. Mol. Ther. Oncolytics 2020, 16, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, A.; Luo, H.; Ramakrishna, S. Antimicrobial Peptides and Their Applications in Biomedical Sector. Antibiotics 2021, 10, 1094. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMPs | Sequence | Molecular Weights | Proportion of the Hydrophobicity |
|---|---|---|---|
| K2F6K2 | Lys-Lys-Phe-Phe-Phe-Phe-Phe-Phe-Lys-Lys | 1431.9 | 60% |
| K3F6K3 | Lys-Lys-Lys-Phe-Phe-Phe-Phe-Phe-Phe-Lys-Lys-Lys | 1688.26 | 50% |
| K4F6K4 | Lys-Lys-Lys-Lys-Phe-Phe-Phe-Phe-Phe-Phe-Lys-Lys-Lys-Lys | 1944.66 | 43% |
| K4F8K4 | Lys-Lys-Lys-Lys-Phe-Phe-Phe-Phe-Phe-Phe-Phe-Phe-Lys-Lys-Lys-Lys | 2239.04 | 50% |
| AMPs | A549 | HLF |
|---|---|---|
| K2F6K2 | 1146.23 ± 346.83 | 758.66 ± 82.02 |
| K3F6K3 | 123.10 ± 35.19 | 758.77 ± 105.3 |
| K4F6K4 | 62.64 ± 9.55 | 808.82 ± 125.7 |
| K4F8K4 | 571.87 ± 66.23 | 704.19 ± 78.34 |
| Concentration (μg/mL) | A549 | HLF |
|---|---|---|
| 0 | 9.59 ± 1.00% | 7.87 ± 0.99% |
| 62 | 12.51 ± 2.23% | 8.85 ± 0.74% |
| 125 | 26.19 ± 0.89% | 8.67 ± 0.92% |
| 250 | 43.03 ± 2.13% | 9.85 ± 1.75% |
| 500 | 89.30 ± 1.41% | 18.82 ± 0.51% |
| 1000 | 92.66 ± 0.46% | 34.28 ± 0.27% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.; Zhu, L.; Lin, X.; Zhu, J.; Qian, Y.; Liu, W.; Chen, J.; Zhou, C.; He, J. Therapeutic Effects of Synthetic Triblock Amphiphilic Short Antimicrobial Peptides on Human Lung Adenocarcinoma. Pharmaceutics 2022, 14, 929. https://doi.org/10.3390/pharmaceutics14050929
Yang D, Zhu L, Lin X, Zhu J, Qian Y, Liu W, Chen J, Zhou C, He J. Therapeutic Effects of Synthetic Triblock Amphiphilic Short Antimicrobial Peptides on Human Lung Adenocarcinoma. Pharmaceutics. 2022; 14(5):929. https://doi.org/10.3390/pharmaceutics14050929
Chicago/Turabian StyleYang, Danjing, Liang Zhu, Xiangyu Lin, Jiaming Zhu, Yusheng Qian, Wenhui Liu, Jianjun Chen, Chuncai Zhou, and Jing He. 2022. "Therapeutic Effects of Synthetic Triblock Amphiphilic Short Antimicrobial Peptides on Human Lung Adenocarcinoma" Pharmaceutics 14, no. 5: 929. https://doi.org/10.3390/pharmaceutics14050929