
The Surface Amine Group of Ultrasmall Magnetic Iron Oxide Nanoparticles Produce Analgesia in the Spinal Cord and Decrease Long-Term Potentiation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs and Administration
2.2. The Particle Size, Zeta Potential, and Surface Group Measurement of Iron Oxide Nanoparticles
2.3. Animal
2.4. CFA Inflammatory Pain Model and Behavior Tests
2.5. Electrophysiological Recordings
2.5.1. Spinal Cord Slice
2.5.2. Hippocampal Slice
2.6. ROS Levels
2.7. Assay of mRNA Expression
2.8. Statistical Analysis
3. Results
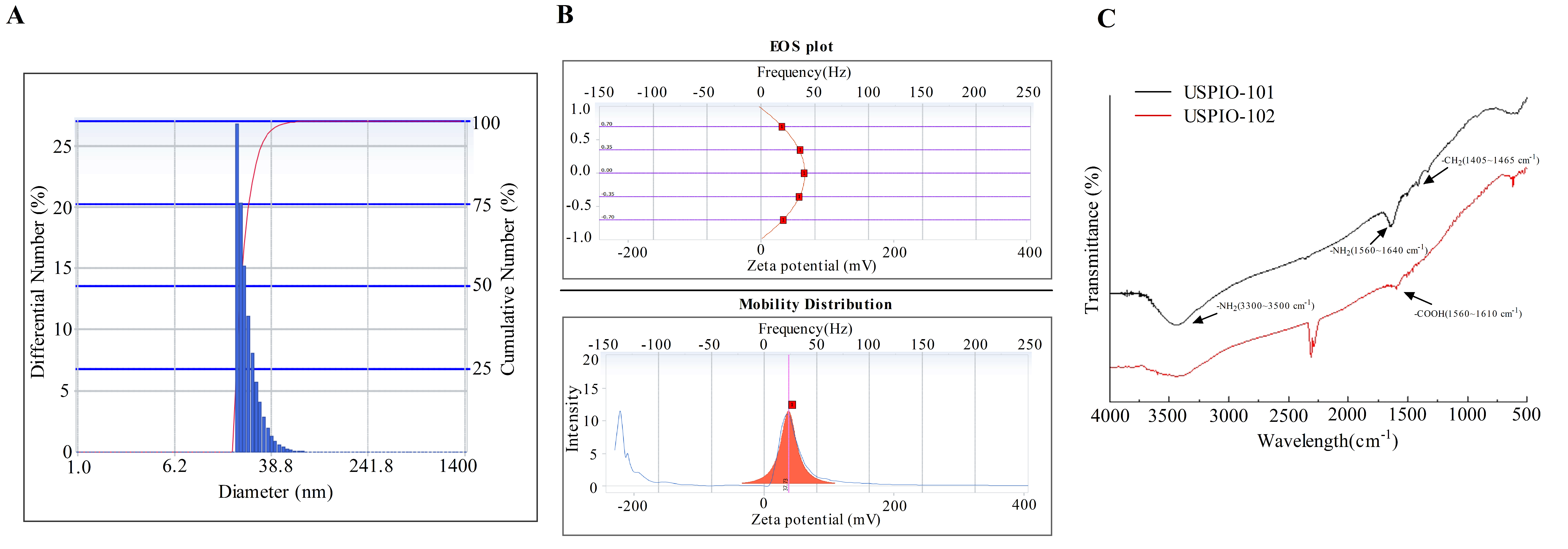
3.1. The Particle Size, Zeta Potential, and Surface Group Analysis of USPIO-101
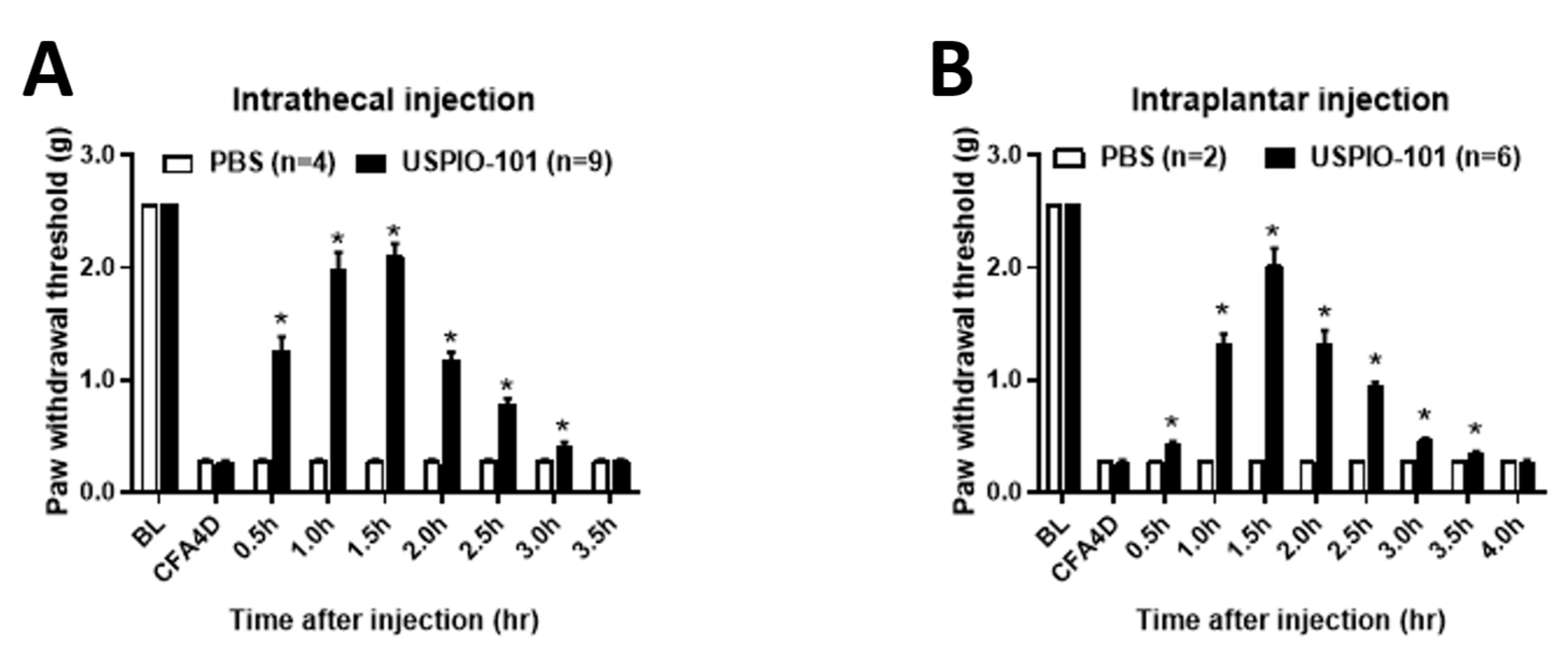
3.2. USPIO-101 Alleviated the Allodynia Behavior via Intrathecal or Intraplantar Injection in Mice with Chronic Inflammatory Pain
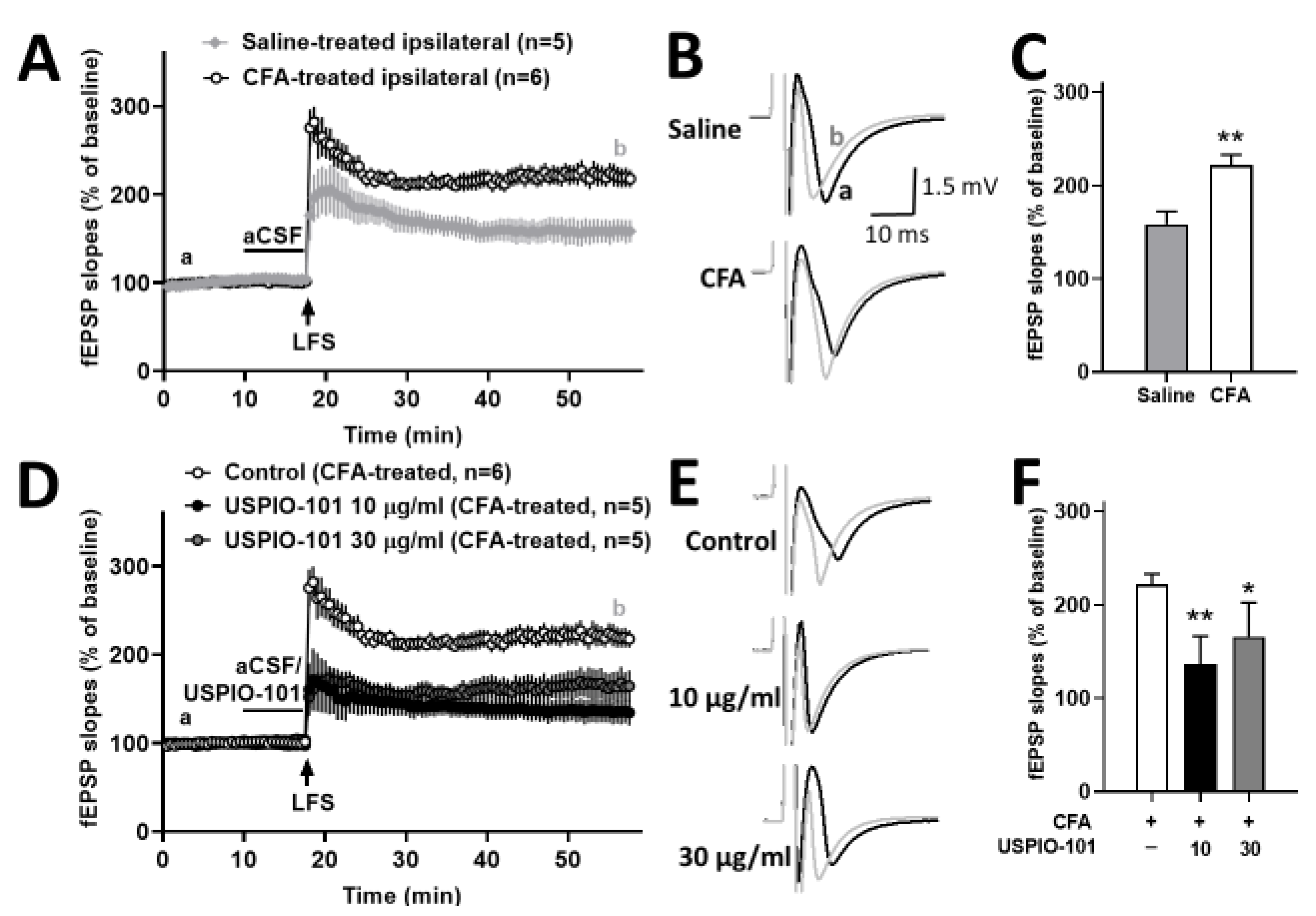
3.3. USPIO-101 Decreased the Spinal Cord LTP at Spinal Cord Superficial Dorsal Horn Synapses in Mice with Chronic Inflammatory Pain and Naïve Mice
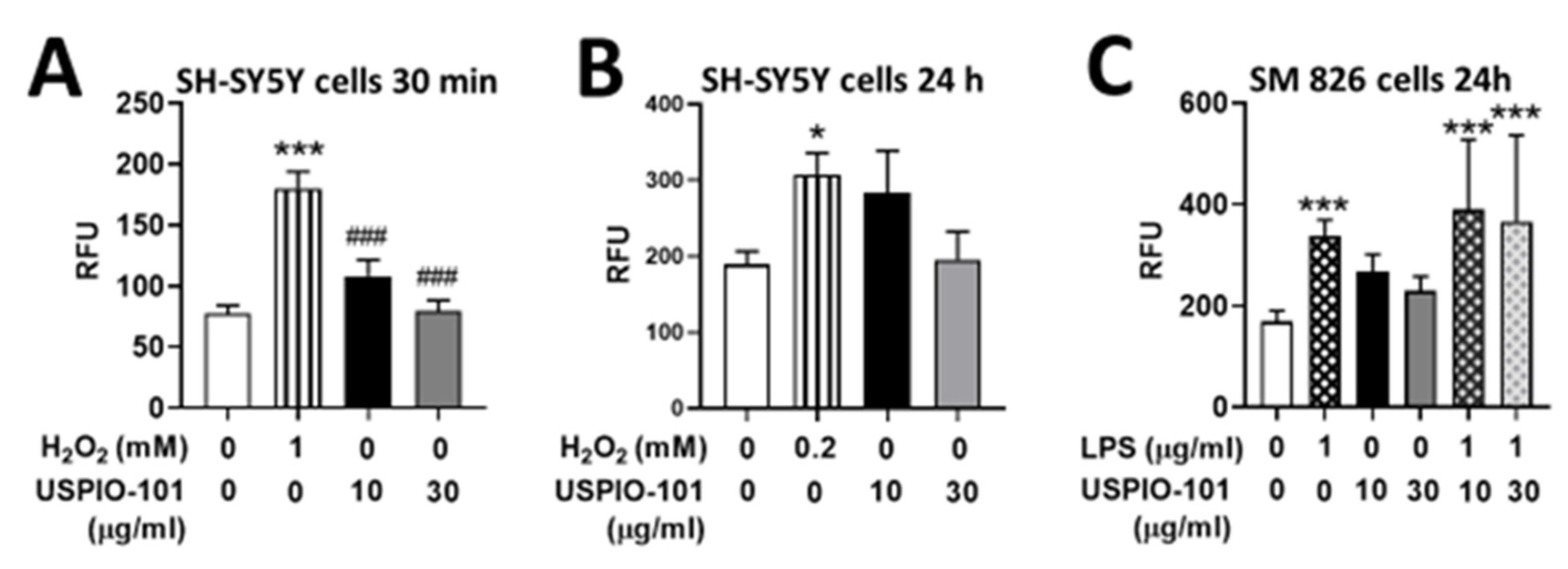
3.4. Effects of USPIO-101 on Intracellular ROS Levels
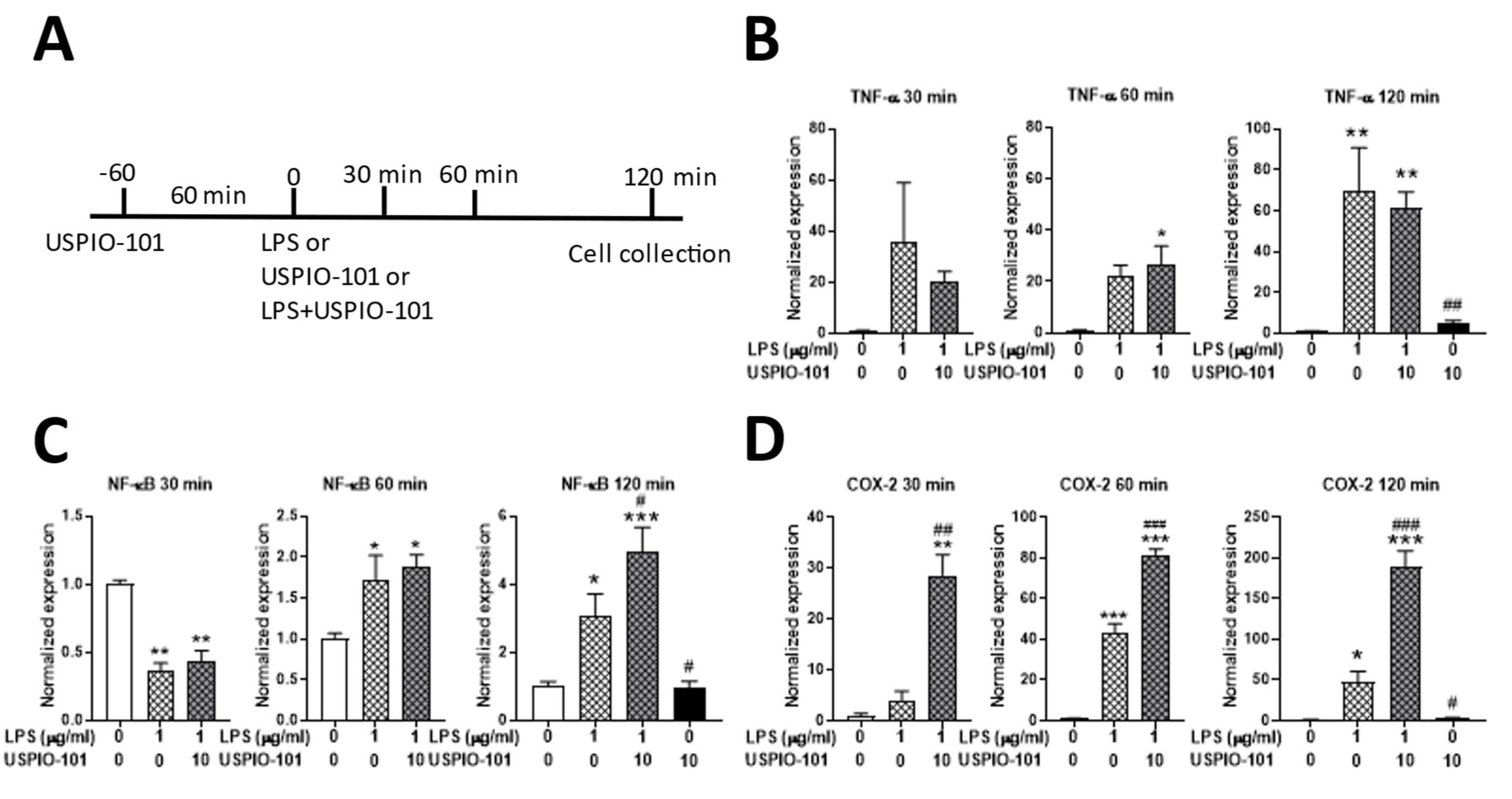
3.5. Effects of USPIO-101 on mRNA Expression with LPS Stimulation
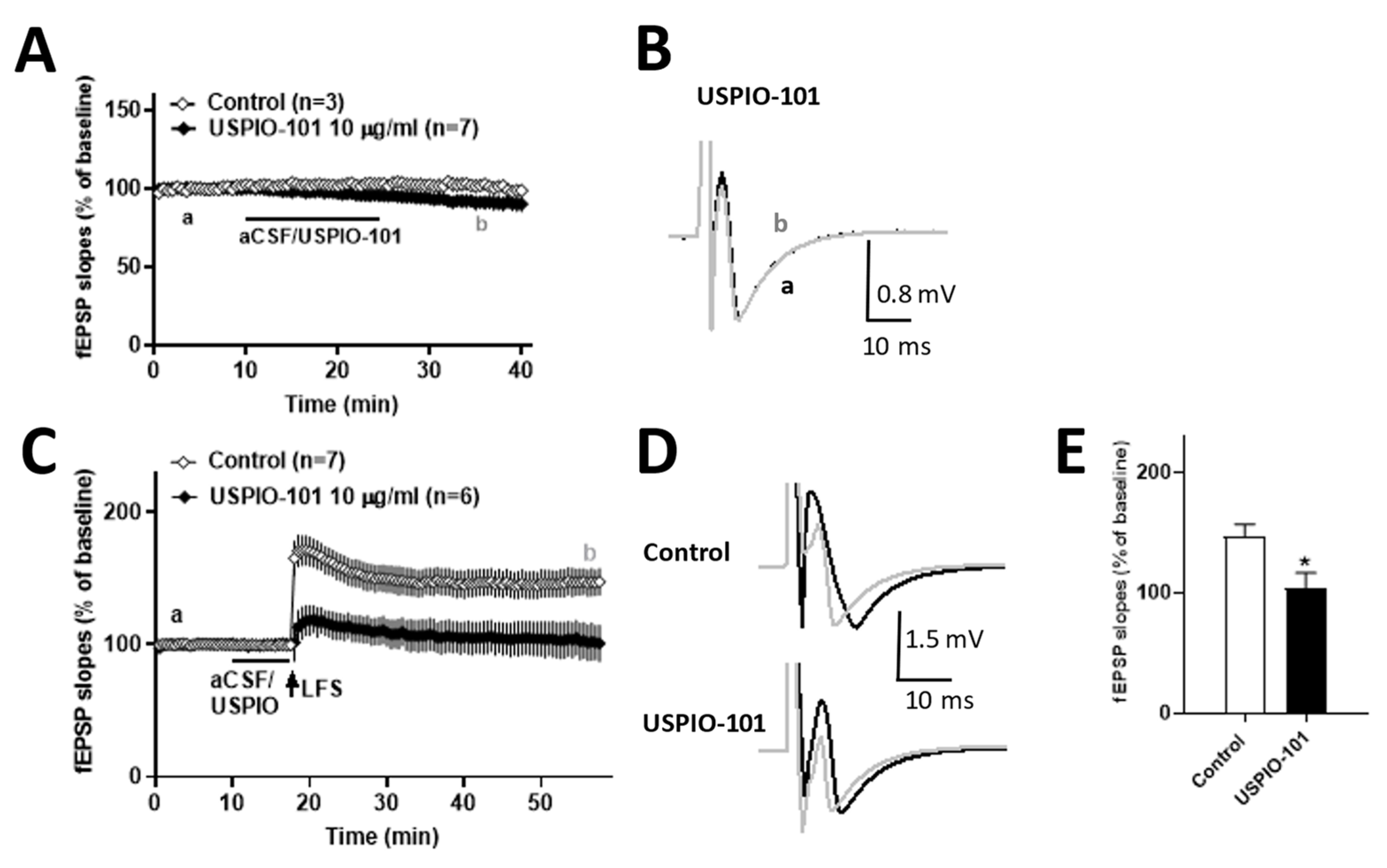
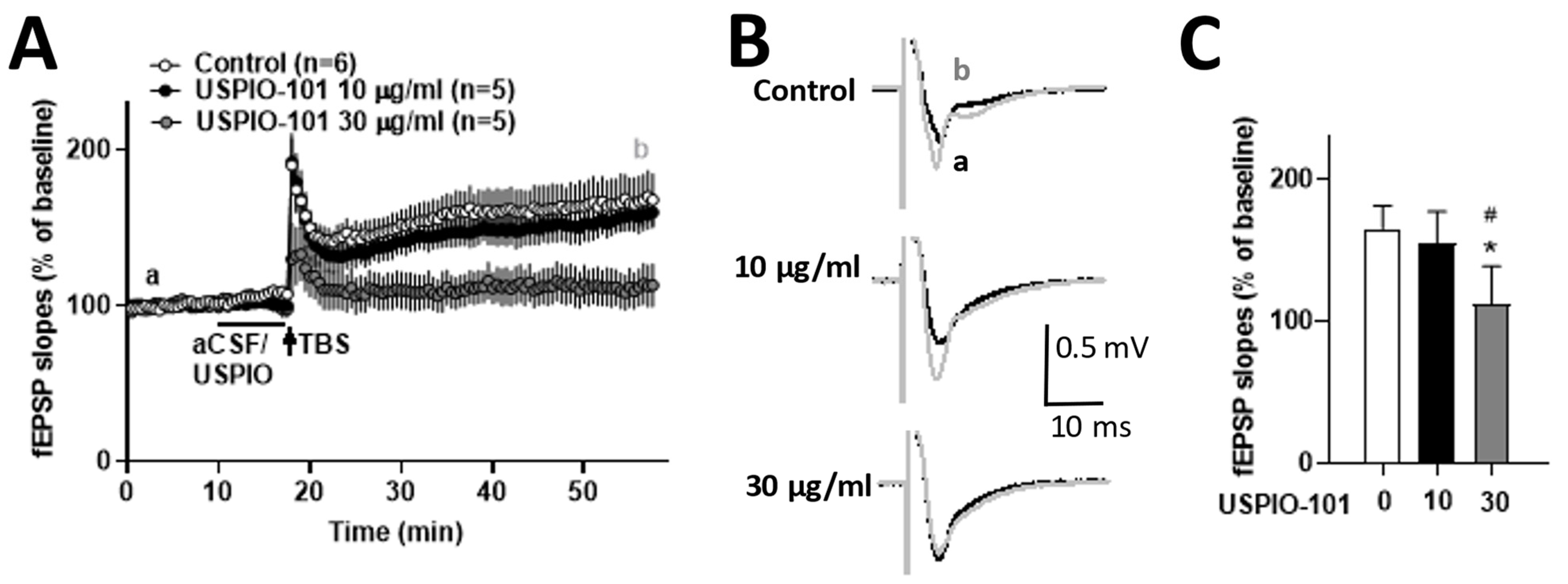
3.6. USPIO-101 Impaired the Hippocampal LTP at the Schaffer Collateral-CA1 Synapses
4. Discussions
4.1. The Analgesic Effect of USPIO-101
4.2. Cell Toxicity of USPIO-101
4.3. Hippocampal LTP Was Impaired by USPIO-101 at a Higher Concentration
4.4. The Effect of Surface Group
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alphandery, E. Iron oxide nanoparticles for therapeutic applications. Drug Discov. Today 2020, 25, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Dadfar, S.M.; Roemhild, K.; Drude, N.I.; von Stillfried, S.; Knuchel, R.; Kiessling, F.; Lammers, T. Iron oxide nanoparticles: Diagnostic, therapeutic and theranostic applications. Adv. Drug Deliv. Rev. 2019, 138, 302–325. [Google Scholar] [CrossRef] [PubMed]
- Andreu, V.; Arruebo, M. Current progress and challenges of nanoparticle-based therapeutics in pain management. J. Control. Release Off. J. Control. Release Soc. 2018, 269, 189–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Jin, T.; Zhang, H. Nanotechnology in Chronic Pain Relief. Front. Bioeng. Biotechnol. 2020, 8, 682. [Google Scholar] [CrossRef]
- Wu, P.C.; Hsiao, H.T.; Lin, Y.C.; Shieh, D.B.; Liu, Y.C. The analgesia efficiency of ultrasmall magnetic iron oxide nanoparticles in mice chronic inflammatory pain model. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 1975–1981. [Google Scholar] [CrossRef]
- Woolf, C.J. Evidence for a central component of post-injury pain hypersensitivity. Nature 1983, 306, 686–688. [Google Scholar] [CrossRef]
- Ruscheweyh, R.; Wilder-Smith, O.; Drdla, R.; Liu, X.G.; Sandkuhler, J. Long-term potentiation in spinal nociceptive pathways as a novel target for pain therapy. Mol. Pain 2011, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Patil, R.M.; Thorat, N.D.; Shete, P.B.; Bedge, P.A.; Gavde, S.; Joshi, M.G.; Tofail, S.A.M.; Bohara, R.A. Comprehensive cytotoxicity studies of superparamagnetic iron oxide nanoparticles. Biochem. Biophys. Rep. 2018, 13, 63–72. [Google Scholar] [CrossRef]
- Nemmar, A.; Beegam, S.; Yuvaraju, P.; Yasin, J.; Tariq, S.; Attoub, S.; Ali, B.H. Ultrasmall superparamagnetic iron oxide nanoparticles acutely promote thrombosis and cardiac oxidative stress and DNA damage in mice. Part. Fibre Toxicol. 2016, 13, 22. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Jiang, S.; Yao, H.; Zhang, L.; Yang, C.; Zhan, D.; Lin, G.; Zeng, Y.; Xia, Y.; Lin, Z.; et al. Endoplasmic reticulum stress mediates inflammatory response triggered by ultra-small superparamagnetic iron oxide nanoparticles in hepatocytes. Nanotoxicology 2018, 12, 1198–1214. [Google Scholar] [CrossRef]
- Che, L.; Yao, H.; Yang, C.L.; Guo, N.J.; Huang, J.; Wu, Z.L.; Zhang, L.Y.; Chen, Y.Y.; Liu, G.; Lin, Z.N.; et al. Cyclooxygenase-2 modulates ER-mitochondria crosstalk to mediate superparamagnetic iron oxide nanoparticles induced hepatotoxicity: An in vitro and in vivo study. Nanotoxicology 2020, 14, 162–180. [Google Scholar] [CrossRef]
- Vermeij, E.A.; Koenders, M.I.; Bennink, M.B.; Crowe, L.A.; Maurizi, L.; Vallee, J.P.; Hofmann, H.; van den Berg, W.B.; van Lent, P.L.; van de Loo, F.A. The in-vivo use of superparamagnetic iron oxide nanoparticles to detect inflammation elicits a cytokine response but does not aggravate experimental arthritis. PLoS ONE 2015, 10, e0126687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, R.R.; Kohno, T.; Moore, K.A.; Woolf, C.J. Central sensitization and LTP: Do pain and memory share similar mechanisms? Trends Neurosci. 2003, 26, 696–705. [Google Scholar] [CrossRef]
- Bliss, T.V.; Lomo, T. Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol. 1973, 232, 331–356. [Google Scholar] [CrossRef] [PubMed]
- Deuis, J.R.; Dvorakova, L.S.; Vetter, I. Methods Used to Evaluate Pain Behaviors in Rodents. Front. Mol. Neurosci. 2017, 10, 284. [Google Scholar] [CrossRef] [Green Version]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Zhu, M.; Zhang, D.; Peng, S.; Liu, N.; Wu, J.; Kuang, H.; Liu, T. Preparation of Acute Spinal Cord Slices for Whole-cell Patch-clamp Recording in Substantia Gelatinosa Neurons. J. Vis. Exp. JoVE 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.Y.; Ho, Y.C.; Hsieh, M.C.; Wang, H.H.; Cheng, J.K.; Chau, Y.P.; Peng, H.Y. Spinal Fbxo3-Dependent Fbxl2 Ubiquitination of Active Zone Protein RIM1alpha Mediates Neuropathic Allodynia through CaV2.2 Activation. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 9722–9738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terman, G.W.; Eastman, C.L.; Chavkin, C. Mu opiates inhibit long-term potentiation induction in the spinal cord slice. J. Neurophysiol. 2001, 85, 485–494. [Google Scholar] [CrossRef]
- Lu, G.L.; Lee, C.H.; Chiou, L.C. Orexin A induces bidirectional modulation of synaptic plasticity: Inhibiting long-term potentiation and preventing depotentiation. Neuropharmacology 2016, 107, 168–180. [Google Scholar] [CrossRef]
- Sandkuhler, J.; Gruber-Schoffnegger, D. Hyperalgesia by synaptic long-term potentiation (LTP): An update. Curr. Opin. Pharmacol. 2012, 12, 18–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdal Dayem, A.; Hossain, M.K.; Lee, S.B.; Kim, K.; Saha, S.K.; Yang, G.M.; Choi, H.Y.; Cho, S.G. The Role of Reactive Oxygen Species (ROS) in the Biological Activities of Metallic Nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, D.; Du, Q.; Ran, B.; Liu, X.; Wang, X.; Ma, X.; Cheng, F.; Sun, B. The neurotoxicity induced by engineered nanomaterials. Int. J. Nanomed. 2019, 14, 4167–4186. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, H.; Stark, J.; Fischer, H.; Wagner, M.; Drdla, R.; Jager, T.; Sandkuhler, J. Synaptic amplifier of inflammatory pain in the spinal dorsal horn. Science 2006, 312, 1659–1662. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.C.; Shieh, D.B.; Hsiao, H.T.; Wang, J.C.; Lin, Y.C.; Liu, Y.C. Magnetic field distribution modulation of intrathecal delivered ketorolac iron-oxide nanoparticle conjugates produce excellent analgesia for chronic inflammatory pain. J. Nanobiotechnology 2018, 16, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Jenkins, G.J.; Asadi, R.; Doak, S.H. Potential toxicity of superparamagnetic iron oxide nanoparticles (SPION). Nano Rev. 2010, 1, 5358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdiglesias, V.; Fernandez-Bertolez, N.; Kilic, G.; Costa, C.; Costa, S.; Fraga, S.; Bessa, M.J.; Pasaro, E.; Teixeira, J.P.; Laffon, B. Are iron oxide nanoparticles safe? Current knowledge and future perspectives. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. 2016, 38, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Vanegas, H.; Schaible, H.G. Prostaglandins and cyclooxygenases [correction of cycloxygenases] in the spinal cord. Prog. Neurobiol. 2001, 64, 327–363. [Google Scholar] [CrossRef]
- Xuan, F.L.; Chithanathan, K.; Lillevali, K.; Yuan, X.; Tian, L. Differences of Microglia in the Brain and the Spinal Cord. Front. Cell. Neurosci. 2019, 13, 504. [Google Scholar] [CrossRef]
- Lee, Y.H.; Lin, C.H.; Hsu, P.C.; Sun, Y.Y.; Huang, Y.J.; Zhuo, J.H.; Wang, C.Y.; Gan, Y.L.; Hung, C.C.; Kuan, C.Y.; et al. Aryl hydrocarbon receptor mediates both proinflammatory and anti-inflammatory effects in lipopolysaccharide-activated microglia. Glia 2015, 63, 1138–1154. [Google Scholar] [CrossRef]
- Frohlich, E. The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int. J. Nanomed. 2012, 7, 5577–5591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Li, J.; Xu, K.; Gu, J.; Huang, L.; Zhang, L.; Liu, N.; Kong, J.; Xing, M.; Zhang, L.; et al. Characterization of superparamagnetic iron oxide nanoparticle-induced apoptosis in PC12 cells and mouse hippocampus and striatum. Toxicol. Lett. 2018, 292, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ding, T.; Sun, J. Neurotoxic potential of iron oxide nanoparticles in the rat brain striatum and hippocampus. Neurotoxicology 2013, 34, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, P.; Mena, N.; Tapia, V.; Arredondo, M.; Nunez, M.T. Iron homeostasis in neuronal cells: A role for IREG1. BMC Neurosci. 2005, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Incontro, S.; Diaz-Alonso, J.; Iafrati, J.; Vieira, M.; Asensio, C.S.; Sohal, V.S.; Roche, K.W.; Bender, K.J.; Nicoll, R.A. The CaMKII/NMDA receptor complex controls hippocampal synaptic transmission by kinase-dependent and independent mechanisms. Nat. Commun. 2018, 9, 2069. [Google Scholar] [CrossRef]
- Penn, A.C.; Zhang, C.L.; Georges, F.; Royer, L.; Breillat, C.; Hosy, E.; Petersen, J.D.; Humeau, Y.; Choquet, D. Hippocampal LTP and contextual learning require surface diffusion of AMPA receptors. Nature 2017, 549, 384–388. [Google Scholar] [CrossRef]
- Shen, L.; Li, B.; Qiao, Y. Fe(3)O(4) Nanoparticles in Targeted Drug/Gene Delivery Systems. Materials 2018, 11, 324. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; He, Q.; Jiang, C. Magnetic iron oxide nanoparticles: Synthesis and surface functionalization strategies. Nanoscale Res. Lett. 2008, 3, 397–415. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, G.-L.; Lin, Y.-C.; Wu, P.-C.; Liu, Y.-C. The Surface Amine Group of Ultrasmall Magnetic Iron Oxide Nanoparticles Produce Analgesia in the Spinal Cord and Decrease Long-Term Potentiation. Pharmaceutics 2022, 14, 366. https://doi.org/10.3390/pharmaceutics14020366
Lu G-L, Lin Y-C, Wu P-C, Liu Y-C. The Surface Amine Group of Ultrasmall Magnetic Iron Oxide Nanoparticles Produce Analgesia in the Spinal Cord and Decrease Long-Term Potentiation. Pharmaceutics. 2022; 14(2):366. https://doi.org/10.3390/pharmaceutics14020366
Chicago/Turabian StyleLu, Guan-Ling, Ya-Chi Lin, Ping-Ching Wu, and Yen-Chin Liu. 2022. "The Surface Amine Group of Ultrasmall Magnetic Iron Oxide Nanoparticles Produce Analgesia in the Spinal Cord and Decrease Long-Term Potentiation" Pharmaceutics 14, no. 2: 366. https://doi.org/10.3390/pharmaceutics14020366