
The Antidiabetic Effects and Modes of Action of the Balanites aegyptiaca Fruit and Seed Aqueous Extracts in NA/STZ-Induced Diabetic Rats

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Animals
2.3. Induction of DM
2.4. Preparation of the B. aegyptiaca Fruit and Seed Aqueous Extracts
2.5. Gas Chromatography–Mass Spectrometry (GC-MS) Analysis
2.6. Experimental Design and Blood and Tissue Sampling
2.7. Blood Sampling and Tissue Sampling
2.8. Biochemical Analysis
2.9. RNA Isolation and RT-PCR
2.10. Determination of Oxidative Stress and Anti-Oxidant Defense Parameters
2.11. Histological Investigation
2.12. Statistical Analysis
3. Results
3.1. GC-MS Analysis
3.2. Effects on OGT
3.3. Effects on Serum Insulin and C-Peptide Levels
3.4. Effect on HOMA-IR Cell Function, HOMA-IS, and HOMA-β Cell Function
3.5. Effects on Liver Glycogen Content and Glucose-6-phospatase and Glycogen Phosphorylase Activities
3.6. Effects on Serum Lipid Profile
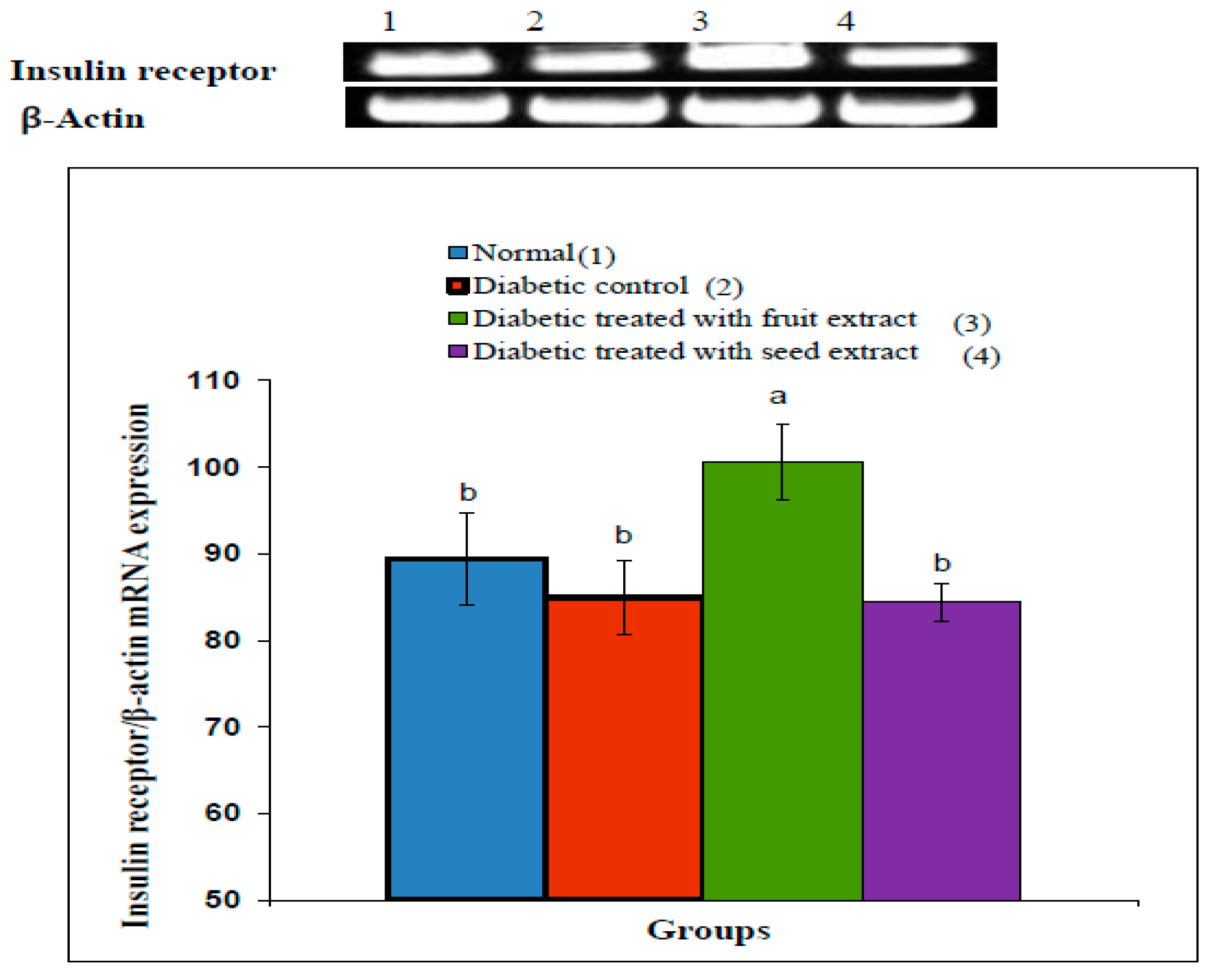
3.7. Effect on Insulin Receptor β-Subunit
3.8. Effect on Oxidative Stress and Anti-Oxidant Defense Parameters
3.9. Histological Changes in the Pancreas
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- He, J.H.; Chen, L.X.; Li, H. Progress in the discovery of naturally occurring anti-diabetic drugs and in the identification of their molecular targets. Fitoterapia 2019, 134, 270–289. [Google Scholar] [CrossRef] [PubMed]
- Reach, G.; Pechtner, V.; Gentilella, R.; Corcos, A.; Ceriello, A. Clinical inertia and its impact on treatment intensification in people with type 2 diabetes mellitus. Diabetes Metab. 2017, 43, 501–511. [Google Scholar] [CrossRef]
- Diabetes Atlas, International Diabetes Federation (IDF), 9th ed.; International Diabetes Federation: Brussels, Belgium, 2019; Available online: https://www.diabetesatlas.org/en/ (accessed on 16 January 2019).
- Hirst, M. Diabetes in 2013, The new figures. Diabetes Res. Clin. Pract. 2013, 102, 265. [Google Scholar] [CrossRef]
- Guariguata, L.; Whiting, D.R.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. Pract. 2014, 103, 137–149. [Google Scholar] [CrossRef]
- Osadebe, P.O.; Odoh, E.U.; Uzor, P.F. Natural Products as Potential Sources of Antidiabetic Drugs. Br. J. Pharmaceut. Res. 2014, 4, 2075–2095. [Google Scholar] [CrossRef]
- Abd El Mgeed, A.; Bstawi, M.; Mohamed, U.; Gabbar, M.A. Histopathological and biochemical effects of green tea and/or licorice aqueous extracts on thyroid functions in male albino rats intoxicated with dimethylnitrosamine. Nutr. Metab. 2009, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, O.M.; Hassan, M.A.; Saleh, A.S. Combinatory effect of hesperetin and mesenchymal stem cells on the deteriorated lipid profile, heart and kidney functions band antioxidant activity in STZ-induced diabetic rats. Biocell 2020, 44, 27–29. [Google Scholar] [CrossRef]
- Verma, S.; Gupta, M.; Popli, H.; Aggarwal, G. Diabetes Mellitus Treatment Using Herbal Drugs. Int. J. Phytomed. 2018, 10, 1–10. [Google Scholar] [CrossRef]
- Chang, C.L.; Lin, Y.; Bartolome, A.P.; Chen, Y.C.; Chiu, S.C.; Yang, W.C. Herbal therapies for type 2 diabetes mellitus: Chemistry, biology, and potential application of selected plants and compounds. Evid. Based Complement. Altern. Med. 2013, 2013, 378657. [Google Scholar] [CrossRef] [PubMed]
- Eskander, E.F.; Jun, H.W. Hypoglycaemic and hyperinsulinemic effects of some egyptian herbs used forthe treatment of diabetes mellitus (type II) in rats. Egypt. J. Pharm. Sci. 1995, 36, 331–342. [Google Scholar]
- Koko, W.S.; Galal, M.; Khalid, H.S. Fasciolicidal efficacy of Albizia anthelmintica and Balanites aegyptiaca compared with albendazole. J. Ethnopharmacol. 2000, 71, 247–252. [Google Scholar] [CrossRef]
- Sarker, S.D.; Bartholomew, B.; Nash, R.J. Alkaloids from Balanites aegyptiaca. Fitoterapia 2000, 71, 328–330. [Google Scholar] [CrossRef]
- Kamel, M.S.; Ohtani, K.; Kurokawa, T.; Assaf, M.H.; El-Shanawany, M.A.; Ali, A.A.; Tanaka, O. Studies on Balanites Aegyptiaca fruits, an antidiabetic Egyptian folk medicine. Chem. Pharm. Bull. 1991, 39, 1229–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gad, M.Z.; El-Sawalhi, M.M.; Ismail, M.F.; El-Tanbouly, N.D. Biochemical study of the anti-diabetic action of the Egyptian plants Fenugreek and Balanites. Mol. Cell. Biochem. 2006, 281, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Hosny, M.; Khalifa, T.; Caliġ, I.; Wright, A.D.; Sticher, O. Balanitoside, a furostanol glycoside, and 6-methyldiosgenin from Balanites aegyptiaca. Phytochemistry 1992, 31, 3565–3569. [Google Scholar] [CrossRef]
- Staerk, D.; Chapagain, B.P.; Lindin, T.; Wiesman, Z.; Jaroszewski, J.W. Structural analysis of complex saponins of Balanites aegyptiaca by 800 MHz 1H NMR spectroscopy. Magn. Reson. Chem. 2006, 44, 923–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elfeel, A.A.; Warrag, E.I. Uses and Conservation Status of Balanites aegyptiaca (L.) Del. (Hegleig Tree) in Sudan: Local People Perspective. Asian J. Agric. Sci. 2011, 3, 286–290. [Google Scholar]
- Aboonabi, A.; Rahmat, A.; Othman, F. Antioxidant effect of pomegranate against streptozotocin-nicotinamide generated oxidative stress induced diabetic rats. Toxicol. Rep. 2014, 1, 915–922. [Google Scholar] [CrossRef] [Green Version]
- Aziz, S.M.A.; Ahmed, O.M.; EL-Twab, S.M.A.; Al-Muzafar, H.M.; Amin, K.A.; Abdel-Gabbar, M. Antihyperglycemic effects and mode of actions of Musa paradisiaca leaf and fruit peel hydroethanolic extracts in nicotinamide/streptozotocin-induced diabetic rats. Evid. Based Complement. Altern. Med. 2020, 2020, 9276343. [Google Scholar]
- Trinder, P. Determination of glucose in blood using glucose oxidase with an alternative oxygen acceptor. Ann. Clin. Biochem. 1969, 6, 24–27. [Google Scholar] [CrossRef]
- Mishra, J.S.; More, A.S.; Kumar, S. Elevated androgen levels induce hyperinsulinemia through increase in Ins1 transcription in pancreatic beta cells in female rats. Biol. Reprod. 2018, 98, 520–531. [Google Scholar] [CrossRef] [Green Version]
- Kuang, L.; Huang, Z.; Hong, Z.; Chen, A.; Li, Y. Predictability of 1-h postload plasma glucose concentration: A 10-year retrospective cohort study. J. Diabetes Investig. 2015, 6, 648–654. [Google Scholar] [CrossRef]
- Seifer, S.; Dayton, S.; Novic, B.; Muntwyler, E. The estimation of glycogen with anthrone reagent. Arch. Biochem. 1950, 25, 191–200. [Google Scholar]
- Begum, N.; Moses, S.G.; Shanmugasundaram, K.R. Serum enzymes in human and experimental diabetes mellitus. Indian J. Med. Res. 1978, 68, 774–784. [Google Scholar]
- Stallman, W.; Hers, H.G. The stimulation of liver phosphorylase b by AMP, fluoride and sulfate. A technical note of the specific determination of the a and b forms of liver glycogen phosphorylase. Eur. J. Biochem. 1975, 54, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Allain, C.C.; Poon, L.S.; Chan, C.S.; Richmond, W.; Fu, P.C. Enzymatic determination of total serum cholesterol. Clin. Chem. 1974, 20, 470–475. [Google Scholar] [CrossRef]
- Fossati, P.; Prenicpe, L. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem. 1982, 28, 2077–2080. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Norbert, W.T. Clinical Guide to Laboratory Tests, 3rd ed.; W.B. Saunders Company: Philadilphia, PA, USA, 1995. [Google Scholar]
- Duncombe, W.G. The coloremitric Micro-determination of long chain fatty acids. Biochem. J. 1964, 88, 7–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomzynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Ann. Rev. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Boom, R.C.J.A.; Sol, C.J.; Salimans, M.M.; Jansen, C.L.; Dillen, P.M.W.-V.; Van der Noordaa, J.P.M.E. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Russell, D.W. Molecular Cloning Laboratory Manual, 5th ed.; Chemical Industry Press: Beijing, China, 2008. [Google Scholar]
- Ahmed, O.M.; Hassan, M.A.; Abdel-Twab, S.M.; Azeem, M.N.A. Navel orange peel hydroethanolic extract, naringin and naringenin have anti-diabetic potentials in type 2 diabetic rats. Biomed. Pharmacother. 2017, 94, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Limin, T.; Hou, X.; Liu, J.; Zhang, X.; Sun, N.; Gao, L.; Zhao, J. Chronic ethanol consumption resulting in the downregulation of insulin receptor-β subunit, insulin receptor substrate-1, and glucose transporter 4 expression in rat cardiac muscles. Alcohol 2009, 43, 51–58. [Google Scholar] [CrossRef]
- Shaker, O.G.; Sadik, N.A.H. Vaspin gene in rat adipose tissue: Relation to obesity-induced insulin resistance. Mol. Cell. Biochem. 2013, 373, 229–239. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Ahmed, A.A.; Fahim, H.I.; Zaky, M.Y. Quercetin and naringenin abate diethylnitrosamine/acetylaminofluorene-induced hepatocarcinogenesis in Wistar rats: The roles of oxidative stress, inflammation and cell apoptosis. Drug Chem. Toxicol. 2019, 42, 1–12, Erratum in Drug Chem. Toxicol. 2020, 43, 112. [Google Scholar] [CrossRef]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Mannervik, B.; Guthenberg, C. Glutathione transferase (human placenta). Methods Enzymol. 1981, 77, 231–235. [Google Scholar]
- MacDougald, O.A.; Burant, C.F. The rapidly expanding family of adipokines. Cell Metab. 2007, 6, 159–161. [Google Scholar] [CrossRef] [Green Version]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Preuss, H.G.; Jarrell, S.T.; Scheckenbach, R.; Lieberman, S.; Anderson, R.A. Comparative effect of chromium vanadium and Gymnema sylvestre on sugar-induced blood pressure elevation in SHR. J. Am. Coll. Nutr. 1998, 17, 116–123. [Google Scholar] [CrossRef]
- Bancroft, J.; Gamble, M. Throry and Practice of Histological Techniques; Churchill Livingstone: London, UK, 2002; pp. 172–175. [Google Scholar]
- Suvarna, S.K.; Layton, C.; Bancroft, J.D. Bancroft Theory and Practice of Histological Techniques, 7th ed.; Elsevier, Churchilli Livingstone: London, UK, 2013. [Google Scholar]
- Roa, M.; Blane, K.; Zonneberg, M. One Way Analysis of Variance, Version IA (C), PC-STAT, Program Coded by University of Georgia; University of Georgia: Athens, GA, USA, 1985. [Google Scholar]
- Trikkalinou, A.; Papazafropoulou, A.K.; Melidonis, A. Type 2 diabetes and quality of life. World J. Diabetes 2017, 8, 120–129. [Google Scholar] [CrossRef]
- Masiello, P. Animal models of type 2 diabetes with reduced pancreatic β-cell mass. Int. J. Biochem. Cell Biol. 2006, 38, 873–893. [Google Scholar] [CrossRef] [PubMed]
- Punitha, I.R.; Rajendran, K.; Shirwaikar, A.; Shirwaikar, A. Alcoholic stem extract of Coscinium fenestratum regulates carbohydrate metabolism and improves antioxidant status in streptozotocin–nicotinamide induced diabetic rats. Evid. Based Complement. Altern. Med. 2005, 2, 375–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veerapur, V.P.; Prabhakar, K.R.; Kandadi, M.R.; Srinivasan, K.K.; Unnikrishnan, M.K. Antidiabetic effect of Dodonaea viscosa aerial parts in high fat diet and low dose streptozotocin-induced type 2 diabetic rats: A mechanistic approach. Pharm. Biol. 2010, 48, 1137–1148. [Google Scholar] [CrossRef] [PubMed]
- Ashok-Kumar, B.S.; Lakshman, K.; Jayaveea, K.N.; SheshadriShekar, D.; Saleemulla Khan, B.S.; Veeresh, T.; Veerapur, P. Antidiabetic, antihyperlipidemic and antioxidant activities of methanolic extract of Amaran thus viridis Linn in alloxan induced diabetic rats. J. Exp. Toxicol. Pathol. 2012, 64, 75–79. [Google Scholar] [CrossRef]
- Salem, M.Z.; Zayed, M.Z.; Ali, H.M.; Abd El-Kareem, M.S. Chemical composition, antioxidant and antibacterial activities of extracts from Schinus molle wood branch growing in Egypt. J. Wood Sci. 2016, 62, 548–561. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, O.M.; Fahim, H.I.; Ahmed, H.Y.; Al-Muzafar, H.M.; Ahmed, R.R.; Amin, K.A.; Abdelazeem, W.H. The preventive effects and the mechanisms of action of navel orange peel hydroethanolic extract, naringin, and naringenin in N-acetyl-p-aminophenol-induced liver injury in Wistar rats. Oxid. Med. Cell. Longev. 2019, 2019, 2745352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burčul, F.; Mekinić, I.G.; Radan, M.; Rollin, P.; Blažević, I. Isothiocyanates: Cholinesterase inhibiting, antioxidant, and anti-inflammatory activity. J. Enzym. Inhib. Med. Chem. 2018, 33, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Alberti, K.G.; Zimmet, P.Z. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: Diagnosis and classification of diabetes mellitus. Provisional report of a WHO Consultation. Diabetes Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Katulanda, G.W.; Katulanda, P.; Dematapitiya, C.; Dissanayake, H.A.; Wijeratne, S.; Sheriff, M.H.R.; Matthews, D.R. Plasma glucose in screening for diabetes and pre-diabetes: How much is too much? Analysis of fasting plasma glucose and oral glucose tolerance test in Sri Lankans. BMC Endocr. Disord. 2019, 19, 11. [Google Scholar] [CrossRef] [Green Version]
- Akhani, S.P.; Vishwakarma, S.L.; Goyal, R.K. Anti-diabetic activity of Zingiber officinale in streptozotocin-induced type I diabetic rats. J. Pharm. Pharmacol. 2004, 56, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, O.M. Evaluation of the antihyperglycemic, antihyperlipidemic and myocardial enhancing properties of pioglitazone in diabetic and hyperthyroid rats. J. Egypt. Ger. Soc. Zool. 2006, 51, 253–278. [Google Scholar]
- Schaalan, M.; El-Abhar, H.S.; Barakat, M.; El-Denshary, E.S. Westernized-like-diet-fed rats: Effect on glucose homeostasis, lipid profile, and adipocyte hormones and their modulation by rosiglitazone and glimepiride. J. Diabetes Complicat. 2009, 23, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, O.M.; Moneim, A.A.; Yazid, I.A.; Mahmoud, A.M. Antihyperglycemic, antihyperlipidemic and antioxidant effects and the probable mechanisms of action of Ruta graveolens infusion and rutin in nicotinamide-streptozotocin-induced diabetic rats. Diabetol. Croat. 2010, 39, 15–32. [Google Scholar]
- Ahmed, O.M.; Hozayen, W.G.M.; Bastawy, M.; Hamed, M.Z. Biochemical effects of Cichorium intybus and Sonchus oleraceus infusions and esculetin on streptozotocin-induced diabetic albino rats. J. Am. Sci. 2011, 7, 1124–1138. [Google Scholar]
- Ali, A.M.; Gabbar, M.A.; Abdel-Twab, S.M.; Fahmy, E.M.; Ebaid, H.; Alhazza, I.M.; Ahmed, O.M. Antidiabetic Potency, Antioxidant Effects, and Mode of Actions of Citrus reticulata Fruit Peel Hydroethanolic Extract, Hesperidin, and Quercetin in Nicotinamide/Streptozotocin-Induced Wistar Diabetic Rats. Oxid. Med. Cell. Longev. 2020, 2020, 1730492. [Google Scholar] [CrossRef]
- Beck-Nielsen, H. Insulin resistance: Organ manifestations and cellular mechanisms. Ugeskr. Laeg. 2002, 164, 2130–2135. [Google Scholar]
- Gold, A.H. The effect of diabetes and insulin on liver glycogen synthetase activation. J. Biol. Chem. 1970, 245, 903–905. [Google Scholar] [CrossRef]
- Raju, J.; Gupta, D.; Rao, A.R.; Yadava, P.K.; Baquer, N.Z. Trigonella foenum graecum (fenugreek) seed powder improves glucose homeostasis in alloxan diabetic rat tissues by reversing the altered glycolytic, gluconeogenic and lipogenic enzymes. Mol. Cell. Biochem. 2001, 224, 45–51. [Google Scholar] [CrossRef]
- Powers, A. Diabetes Mellitus. In Harrison’s Principles of Internal Medicine; Kasper, D.L., Fauci, A.S., Longo, D., Braunwald, E., Hauser, S., Jameson, J.L., Eds.; Mcgraw-Hill: New York, NY, USA, 2005. [Google Scholar]
- Zaahkouk, S.A.; Rashid, S.Z.; Mattar, A.F. Anti-diabetic properties of water and ethanolic extracts of Balanites aegyptiaca fruits flesh in senile diabetic rats. Egypt. J. Hosp. Med. 2003, 10, 90–108. [Google Scholar] [CrossRef]
- Helal, E.G.; El-Wahab, A.; Samia, M.; El Refaey, H.; Mohammad, A.A. Antidiabetic and antihyperlipidemic effect of Balanites aegyptiaca seeds (aqueous extract) on diabetic rats. Egypt. J. Hosp. Med. 2013, 52, 725–739. [Google Scholar] [CrossRef]
- Al-Malki, A.L.; Barbour, E.K.; Abulnaja, K.O.; Moselhy, S.S. Management of hyperglycaemia by ethyl acetate extract of Balanites aegyptiaca (desert date). Molecules 2015, 20, 14425–14434. [Google Scholar] [CrossRef] [Green Version]
- Motaal, A.A.; Shaker, S.; Haddad, P.S. Antidiabetic activity of standardized extracts of Balanites aegyptiaca fruits using cell-based bioassays. Pharmacol. J. 2012, 4, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Moneim, A. Effect of some medicinal plants and gliciazide on insulin release in vitro. J. Egypt. Ger. Soc. Zool. 1998, 25, 423–445. [Google Scholar]
- Nour, M.E.; Khalifa, E.L.; Massimo, K.; Hassen, B. Preliminary study on seed pregermination treatments and vegetative propagation of Balanites aegyptiaca (L). Physiology 1991, 4, 413–415. [Google Scholar]
- Baragob, A.E.A.; AlMalki, W.H.; Shahid, I.; Bakhdhar, F.A.; Bafhaid, H.S.; Eldeen, O.M.I. The hypoglycemic effects of the aqueous extract of the fruits of Balanites aegypticea in Alloxan-induced diabetic rats. Pharmacogn. Res. 2014, 6, 1–5. [Google Scholar]
- Khalil, N.S.A.; Abou-Elhamd, A.S.; Wasfy, S.I.; El Mileegy, I.M.; Hamed, M.Y.; Ageely, H.M. Antidiabetic and antioxidant impacts of desert date (Balanites aegyptiaca) and parsley (Petroselinum sativum) aqueous extracts: Lessons from experimental rats. J. Diabetes Res. 2016, 2016, 8408326. [Google Scholar]
- El-Bayomy, M.M.; El-Mously, M.; El-Stoohy, F.; Mehanna, S.S. Effect of oral administration of the aqueous extract of Balanites aegyptiaca dates on blood glucose, serum insulin and lipids in streptozotocin-induced diabetes in rats. J. Biomed. Ther. Sci. 1992, 8, 82–89. [Google Scholar]
- Hassan, M. Physiological and Herpbiochemical Studies on the Effects of Balanites Aegyptiaca in Albino Ratápdez; Department of Botany, Faculty of Science, Assiut University: Asyut, Egypt, 2000. [Google Scholar]
- Chapagain, B.; Wiesman, Z. Variation in diosgenin level in seed kernels among different provenances of Balanites aegyptiaca Del (Zygophyllaceae) and its correlation with oil content. Afr. J. Biotechnol. 2005, 4, 1209–1213. [Google Scholar]
- Ulbricht, C.; Basch, E.; Burke, D.; Cheung, L.; Ernst, E.; Giese, N.; Weissner, W. Fenugreek (Trigonella foenum-graecum L. Leguminosae):anevidence-based systematic review by the natural standard research collaboration. J. Herb. Pharmacother. 2008, 7, 143–177. [Google Scholar] [CrossRef]
- Wahren, J.; Ekberg, K.; Samnegard, B.; Johansson, B.L. C peptide: A new potential in the treatment of diabetic nephro pathy. Curr. Diabetese Rep. 2001, 1, 261–266. [Google Scholar] [CrossRef]
- Grover, J.K.; Vats, V.; Rathi, S.S. Antihyperglycemic effect of Eugenia jambolana and Tinospora cordifolia in experimental diabetes and their effects on key metabolic enzymes involved in carbohydrate metabolism. J. Ethnopharmacol. 2000, 73, 461–470. [Google Scholar] [CrossRef]
- Sundaram, R.; Shanthi, P.; Sachdanandam, P. Effect of tangeretin, a polymethoxylated flavone on glucose metabolism in streptozotocin-induced diabetic rats. Phytomedicine 2014, 21, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Ahmed, O.M.; Ashour, M.B.; Abdel-Moneim, A. In vivo and in vitro antidiabetic effects of citrus flavonoids; a study on the mechanism of action. Int. J. Diabetes Dev. Ctries. 2015, 35, 250–263. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.; Al-Zayat, E.; Mahmoud, S. Effect of some antioxidants on streptozotocin diabetic rat’s comparative physiology. J. Egypt. Ger. Soc. Zool. 2002, 38, 213–245. [Google Scholar]
- Minnich, A.; Zilversmit, D.B. Impaired triacylglycerol catabolism in hypertriglyceridemia of the diabetic, cholesterol fed rabbit: A possible mechanism for protection from atherosclerosis. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1989, 1002, 324–332. [Google Scholar] [CrossRef]
- Goodman, L.S. Goodman Gilman’s the Pharmacological Basis of Therapeutics, 11th ed.; Brunton, L.L., Lazo, J.S., Parker, K.L., Eds.; Mcgraw-Hill: New York, NY, USA, 2006; pp. 1613–1644. [Google Scholar]
- Harvey, R.A.; Ferrier, D.R. Lippincott’s Illustrated Reviews. In Biochemistry, 5th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 307, 2011. [Google Scholar]
- Budin, S.B.; Othman, F.; Louis, S.R.; Bakar, M.A.; Radzi, M.; Osman, K.; Das, S.; Mohamed, J. Effect of alpha lipoic acid on oxidative stress and vascular wall of diabetic rats. Rom. J. Morphol. Embryol. 2009, 50, 23–30. [Google Scholar]
- Pottathil, S.; Nain, P.; Morsy, M.A.; Kaur, J.; Al-Dhubiab, B.E.; Jaiswal, S.; Nair, A.B. Mechanisms of Antidiabetic Activity of Methanolic Extract of Punica granatum Leaves in Nicotinamide/Streptozotocin-Induced Type 2 Diabetes in Rats. Plants 2020, 9, 1609. [Google Scholar] [CrossRef] [PubMed]
- Morakinyo, A.O.; Samuel, T.A.; Adekunbi, D.A. Magnesium upregulates insulin receptor and glucose transporter-4 in streptozotocin-nicotinamide-induced type-2 diabetic rats. Endocr. Regul. 2018, 52, 6–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omolola, S.O.; Bukoye, O.H. Improvement of Oral Glucose Tolerance and Total Lipid Profile of Diabetic Rats Treated with Ficus exasperata Leaf-Based Diet. Covenant J. Phys. Life Sci. 2019, 7, 982–993. [Google Scholar]
- Stanfield, C.L. Principles of Human Physiology, 4th ed.; Publishing as Benjamin Cummings; Manufactured in the United States of America; Pearson Education, Inc.: London, UK, 2011. [Google Scholar]
- Rawi, S.M.; Abdel-Moneim, A.; Ahmed, O.M. Studies on the effect of garlic oil and glibenclamide on alloxan diabetic rats. Egypt J. Zool. 1998, 30, 211–228. [Google Scholar]
- Mathe, D. Dyslipidemia and diabetes: Animal models. Diabetes Metab. 1995, 21, 106–111. [Google Scholar]
- Matter, F.E.; Helal, E.G.E. Effect of Blanitis aegyptiaca seed extract on factors affecting serum glucose level in hyperglycemic senile albino rats. Al-Azhar Bull. Sci. 2001, 12, 231–245. [Google Scholar]
- El-Rahman, S.N.A.; Al-ahmari, H. Evaluation of fertility potential of Balanites aegyptiaca sapogenin extract in male rats. Int. J. Sudan Res. 2013, 3, 15–33. [Google Scholar] [CrossRef]
- Roman, I.D.; Thewles, A.; Coleman, R. Fractionation of livers following diosgenin treatment to elevate biliary cholesterol. Biochim. Biophys. Acta 1995, 1255, 77–81. [Google Scholar] [CrossRef]
- Abdin, A.A.; Baalash, A.A.; Hamooda, H.E. Effects of rosiglitazone and aspirin on experimental model of induced type 2 diabetes in rats: Focus on insulin resistance and inflammatory markers. J. Diab. Complicat. 2010, 24, 168–178. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Mahmoud, A.M.; Abdel-Moneim, A.; Ashour, M.B. Antidiabetic effects of hesperidin and naringin in type 2 diabetic rats. Diabetol. Croat. 2012, 41, 53–67. [Google Scholar]
- Lewis, G.F.; Carpentier, A.; Adeli, K.; Giacca, A. Disordered fat storage and mobilization in the pathogenesis of insulin resistance and type 2 diabetes. Endocr. Rev. 2002, 23, 201–229. [Google Scholar] [CrossRef]
- Na, L.X.; Zhang, Y.L.; Li, Y.; Liu, L.Y.; Li, R.; Kong, T.; Sun, C.H. Curcumin improves insulin resistance in skeletal muscle of rats. Nutr. Metab. Cardiovasc. Diseases. 2011, 21, 526–533. [Google Scholar] [CrossRef]
- Qi, Y.; Nie, Z.; Lee, Y.S.; Singhal, N.S.; Scherer, P.E.; Lazar, M.A.; Ahima, R.S. Loss of resistin improves glucose homeostasis in leptin deficiency. Diabetes 2006, 55, 3083–3090. [Google Scholar] [CrossRef] [Green Version]
- Way., J.M.; Görgün, C.Z.; Tong, Q.; Uysal, K.T.; Brown, K.K.; Harrington, W.W.; Oliver, W.R.; Willson, T.M.; Kliewer, S.A.; Hotamisligil, G.S. Adipose tissue resistin expression is severely suppressed in obesity and stimulated by peroxisome proliferator-activated receptor gamma agonists. J. Biol. Chem. 2001, 276, 25651–25653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karthikesan, K.; Pari, L.; Menon, V.P. Combined treatment of tetra hyd rocu cumin and chlorogenic acid exerts potential antihyperglycemic effect on streptozotocin -nicotinamide-induced diabetic rats. Gen. Physiol. Biophys. 2010, 29, 23–30. [Google Scholar] [CrossRef]
- Randle, P.J.; Garland, C.N.; Hales, E.A. Newsholme.The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1963, 281, 785–789. [Google Scholar] [CrossRef]
- Brøns, C.; Grunnet, L.G. Mechanisms in endocrinology: Skeletal muscle lipotoxicity in insulin resistance and type 2 diabetes: A causal mechanism or an innocent bystander. Eur. J. Endocr. 2017, 176, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Belfort, R.; Mandarino, L.; Kashyap, S.; Wirfel, K.; Pratipanawatr, T.; Berria, R.; Defronzo, R.A.; Cusi, K. Dose-response effect of elevated plasma free fatty acid on insulin signaling. Diabetes 2005, 54, 1640–1648. [Google Scholar] [CrossRef] [Green Version]
- Iranloye, B. Anti-diabetic and antioxidant effects of virgin coconut oil in alloxan induced diabetic male Sprague Dawley rats. J. Diabetes Mellit. 2013, 3, 221. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, O.M.; El-Twab, S.M.A.; Al-Muzafar, H.M.; Amin, K.A.; Aziz, S.M.A.; Abdel-Gabbar, M. Musa paradisiaca L. leaf and fruit peel hydroethanolic extracts improved the lipid profile, glycemic index and oxidative stress in nicotinamide/streptozotocin-induced diabetic rats. Vet. Med. Sci. 2021, 7, 500–511. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 9, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Çelik, N.; Vurmaz, A.; Kahraman, A. Protective effect of quercetin on homocysteine-induced oxidative stress. Nutrition 2017, 33, 291–296. [Google Scholar] [CrossRef]
- Hassanin, K.M.; Mahmoud, M.O.; Hassan, H.M.; Abdel-Razik, A.R.H.; Aziz, L.N.; Rateb, M.E. Balanites aegyptiaca ameliorates insulin secretion and decreases pancreatic apoptosis in diabetic rats: Role of SAPK/JNK pathway. Biomed. Pharmacother. 2018, 102, 1084–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | (RT) Retention Time | Compound Name | Area% (Higher than 1%) |
|---|---|---|---|
| 1 | 2.123 | Hydrazine, 1,1-dimethyl- | 4.31 |
| 2 | 3.008 | Butanal, 2-methyl- | 3.04 |
| 3 | 3.043 | Butanal, 2-methyl- | 1.89 |
| 4 | 3.363 | Propanoic acid, propyl ester | 3.41 |
| 5 | 3.907 | Glyceraldehyde | 5.39 |
| 6 | 4.250 | 2-Furanmethanol | 2.55 |
| 7 | 4.645 | Isopropyl isothiocyanate | 2.64 |
| 8 | 4.889 | Triethylphosphine | 9.31 |
| 9 | 9.454 | 2,4(3H,5H)-Furandione | 1.46 |
| 10 | 9.744 | Pyridine, 4-chloro-2,6-dimethyl- | 0.96 |
| 11 | 18.561 | 3,4,6-Tri-O-methyl-d-glucose | 52.55 |
| 12 | 21.667 | Perhydrohistrionicotoxin-2-thione, 2-depentyl- | 2.07 |
| 13 | 23.758 | Propyl 11,12-methylene-octadecanoate | 3.06 |
| Number | Retention Time | Compound Name | Area% (Higher than 1%) |
|---|---|---|---|
| 1 | 4.263 | Butane, 2-methyl- | 1.67 |
| 2 | 20.538 | -3,4,6Tri-O-methyl-d-glucose | 2.59 |
| 3 | 24.018 | H-Cyclopenta [b]quinoxaline-1,2,3trione | 11.39 |
| 4 | 25.505 | -9,12Octadecadienoic acid (Z,Z)- | 2.47 |
| 5 | 25.568 | 13-Octadecenoic acid, methyl ester | 1.62 |
| 6 | 26.152 | 9,12-Octadecadienoic acid (Z,Z)- | 38.27 |
| 7 | 26.192 | 8-Dodecen-1-ol, (Z)- | 15.09 |
| 8 | 26.281 | 9,12-Octadecadienoic acid (Z,Z)-, 2,3-dihydroxypropyl ester | 11.47 |
| 9 | 26.339 | Octadec-9-enoic acid | 6.06 |
| 10 | 26.371 | Benzoic acid, 4-(4-hydroxybenzylidenamino)-, propyl ester | 6.79 |
| Parameter | Insulin (μIU/mL) | C-Peptide (pg/mL) | |
|---|---|---|---|
| Group | |||
| Normal | 2.56 ± 0.51 a | 4.7 ± 0.13 a | |
| Diabetic control | 1.00 ± 0.78 c | 1.19 ± 0.13 c | |
| Diabetic treated with fruit extract | 1.71 ± 0.11 b | 3.47 ± 0.44 b | |
| Diabetic treated with seed extract | 1.56 ± 0.11 b | 3.50 ± 0.22 b | |
| Parameter | HOMA-IR | HOMA-IS | HOMA-β Cell Function | |
|---|---|---|---|---|
| Group | ||||
| Normal | 0.37 ± 0.03 c | 55.48 ± 2.23 a | 55.50 ± 1.42 a | |
| Diabetic control | 0.51 ± 0.04 a | 45.50 ± 0.66 b | 2.71 ± 0.17 d | |
| Diabetic treated with fruit extract | 0.40 ± 0.03 b | 55.00 ± 1.23 a | 11.20 ± 0.90 b | |
| Diabetic treated with seed extract | 0.45 ± 0.03 b | 53.33 ± 1.03 a | 7.80 ± 0.39 c | |
| Parameter | Liver Glycogen (mg P liberated/g Tissue/h) | Glucose-6-phosphatase (mg Pi liberated/g Tissue/h) | Glycogen Phosphorylase (mg Pi liberated/g Tissue/h) | |
|---|---|---|---|---|
| Group | ||||
| Normal | 54.55 ± 3.60 a | 25.79 ± 1.53 b | 22.78 ± 3.09 b | |
| Diabetic control | 8.10 ± 1.66 b | 67.42 ± 3.38 a | 41.85 ± 6.08 a | |
| Diabetic treated with fruit extract | 42.75 ± 2.91 a | 28.02 ± 0.41 b | 25.45 ± 4.95 b | |
| Diabetic treated with seed extract | 50.51 ± 3.60 a | 31.45 ± 2.28 b | 15.22 ± 2.20 b | |
| Parameter | Total Cholesterol (mg/dL) | Triglycerides (mg/dL) | LDL-Cholesterol (mg/dL) | HDL- Cholesterol (mg/dL) | vLDL- Cholesterol (mg/dL) | FFAs (mg/dL) | |
|---|---|---|---|---|---|---|---|
| Group | |||||||
| Normal | 42.70 ± 1.34 c | 35.66 ± 2.25 c | 5.10 ± 2.10 c | 31.30 ± 2.30 a | 7.11 ± 0.33 c | 5.10 ± 0.86 b | |
| Diabetic control | 79.40 ± 9.48 a | 80.25 ± 15.30 a | 37.48 ± 6.03 a | 24.80 ± 2.80 b | 16.08 ± 3.00 a | 18.70 ± 2.7 a | |
| Diabetic treated with fruit extract | 59.70 ± 9.30 b | 53.70 ± 2.83 b | 13.18 ± 4.76 b | 31.00 ± 2.10 a | 8.11 ± 1.05 c | 5.10 ± 0.70 b | |
| Diabetic treated with seed extract | 51.80 ± 9.70 bc | 41.25 ± 5.34 c | 15.94 ± 7.90 b | 27.10 ± 0.80 b | 10.71 ± 0.056 b | 6.75 ± 1.80 b | |
| Parameter | LPO (nmole MDA/100 mg Tissue/h) | GSH (nmole/100 mg Tissue) | GPx (mU/100 mg Tissue) | GST (U/100mg Tissue) | SOD (U/g Tissue) | |
|---|---|---|---|---|---|---|
| Group | ||||||
| Normal | 67.17 ± 1.25 bc | 13.30 ± 1.63 a | 42.60 ± 5.1 a | 12.65 ± 3.6 b | 16.78 ± 1.59 a | |
| Diabetic control | 111.39 ± 7.57 a | 6.10 ± 1.60 c | 17.20 ± 5.1 c | 6.57 ± 1.65 c | 10.31 ± 1.58 c | |
| Diabetic treated with fruit extract | 68.75 ± 3.80 b | 12.10 ± 1.54 ab | 30.80 ± 4.6 b | 16.54 ± 0.86 a | 13.46 ± 2.55 b | |
| Diabetic treated with seed extract | 52.95 ± 5.821 c | 11.20 ± 1.5 b | 31.00 ± 2.5 b | 16.43 ± 0.85 a | 13.26 ± 2.08 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaky, A.S.; Kandeil, M.; Abdel-Gabbar, M.; Fahmy, E.M.; Almehmadi, M.M.; Ali, T.M.; Ahmed, O.M. The Antidiabetic Effects and Modes of Action of the Balanites aegyptiaca Fruit and Seed Aqueous Extracts in NA/STZ-Induced Diabetic Rats. Pharmaceutics 2022, 14, 263. https://doi.org/10.3390/pharmaceutics14020263
Zaky AS, Kandeil M, Abdel-Gabbar M, Fahmy EM, Almehmadi MM, Ali TM, Ahmed OM. The Antidiabetic Effects and Modes of Action of the Balanites aegyptiaca Fruit and Seed Aqueous Extracts in NA/STZ-Induced Diabetic Rats. Pharmaceutics. 2022; 14(2):263. https://doi.org/10.3390/pharmaceutics14020263
Chicago/Turabian StyleZaky, Asmaa S., Mohamed Kandeil, Mohamed Abdel-Gabbar, Eman M. Fahmy, Mazen M. Almehmadi, Tarek M. Ali, and Osama M. Ahmed. 2022. "The Antidiabetic Effects and Modes of Action of the Balanites aegyptiaca Fruit and Seed Aqueous Extracts in NA/STZ-Induced Diabetic Rats" Pharmaceutics 14, no. 2: 263. https://doi.org/10.3390/pharmaceutics14020263