
Standardization and Key Aspects of the Development of Whole Yeast Cell Vaccines

Abstract
:1. Introduction
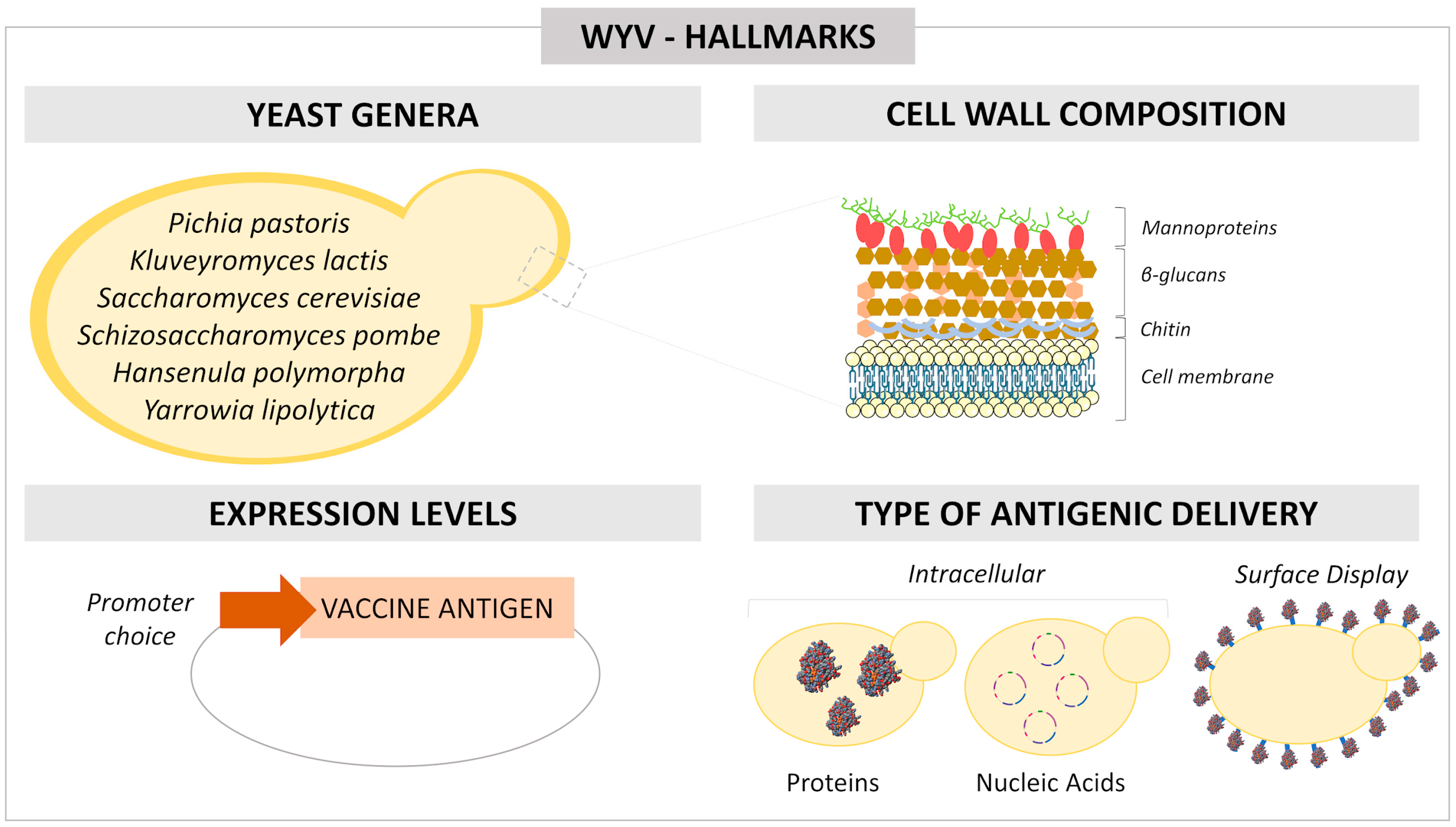
2. Yeast Genera
3. Yeast Cell Inactivation
4. Storage and Stability of Vaccine Preparations

{kind=link}
{kind=link}
| Yeast Genera | Infectious Agent | Inactivation | Route | [Dose] | Animal Models | Ref. |
|---|---|---|---|---|---|---|
| K. lactis | Infectious bursal disease virus | 90 °C/2 h | SC and oral | SC: 100 µg/200 µL (mouse) and 1 mg/500 µL (chicken). Oral: dried yeast nuggets mixed with feed (end concent. of 5% w/w). | Mouse and Chicken | [32] |
| K. lactis | Influenza A virus | 90 °C/2 h | SC | 1, 2 or 5 mg/100 µL | Mouse | [41] |
| H. polymorpha | Hepatitis B | 60 °C/2 h | IM | 2 × 108 yeast cells/100 µL | Mouse | [16] |
| P. pastoris | Plasmodium berghei | 60 °C/45 min | SC | 30 YU/100 µL | Mouse | [30] |
| P. pastoris | Human Papillomavirus | 60 °C/2 h | SC | 2.5, 5 and 10 mg | Mouse | [42] |
| P. pastoris | Human Papillomavirus | 56 °C/15 min | SC (multipoint injection) | 5 µg (2 × 102 yeast cells) | Mouse | [43] |
| P. pastoris | Highly pathogenic avian influenza | - | Oral | 6.7 × 109 cells/mL in 2.5 mL | Chicken | [44] |
| S. cerevisiae | Dengue | - | Oral | 1.6 g (fresh) in 2.4 mL | Mouse | [45] |
| S. cerevisiae | Vibrio harveyi | - | IP | 5 × 109 cells/mL in 200 µL | Marine fish (flounder turbot) | [46] |
| S. cerevisiae | Actinobacillus pleuropneumoniae | - | Oral | 1.5 × 109 cells/day | Mouse | [11] |
| S. cerevisiae | Porcine circovirus type 2 | (Freeze-dried) | Oral | 7 g (freeze dried yeast) in 20 mL of sterile | Pig | [35] |
| S. cerevisiae | H7N9 virus | 60 °C/1 h | Oral | 150 OD600 | Mouse | [17] |
| S. cerevisiae | Coccidioides immitis | 70–75 °C/3 h | Oral, IM | 1.2 × 108 cells (5 mg per dose) * 6 × 107 cells (2.5 mg per animal) | Mouse | [34] |
| S. cerevisiae | Eimeria tenella | 56 °C/1 h and 95 °C/2 min | Oral | 1.7 YU in 100 µL and 1.5 × 107 cells/mL (200 µL per animal) | Chicken | [29] |
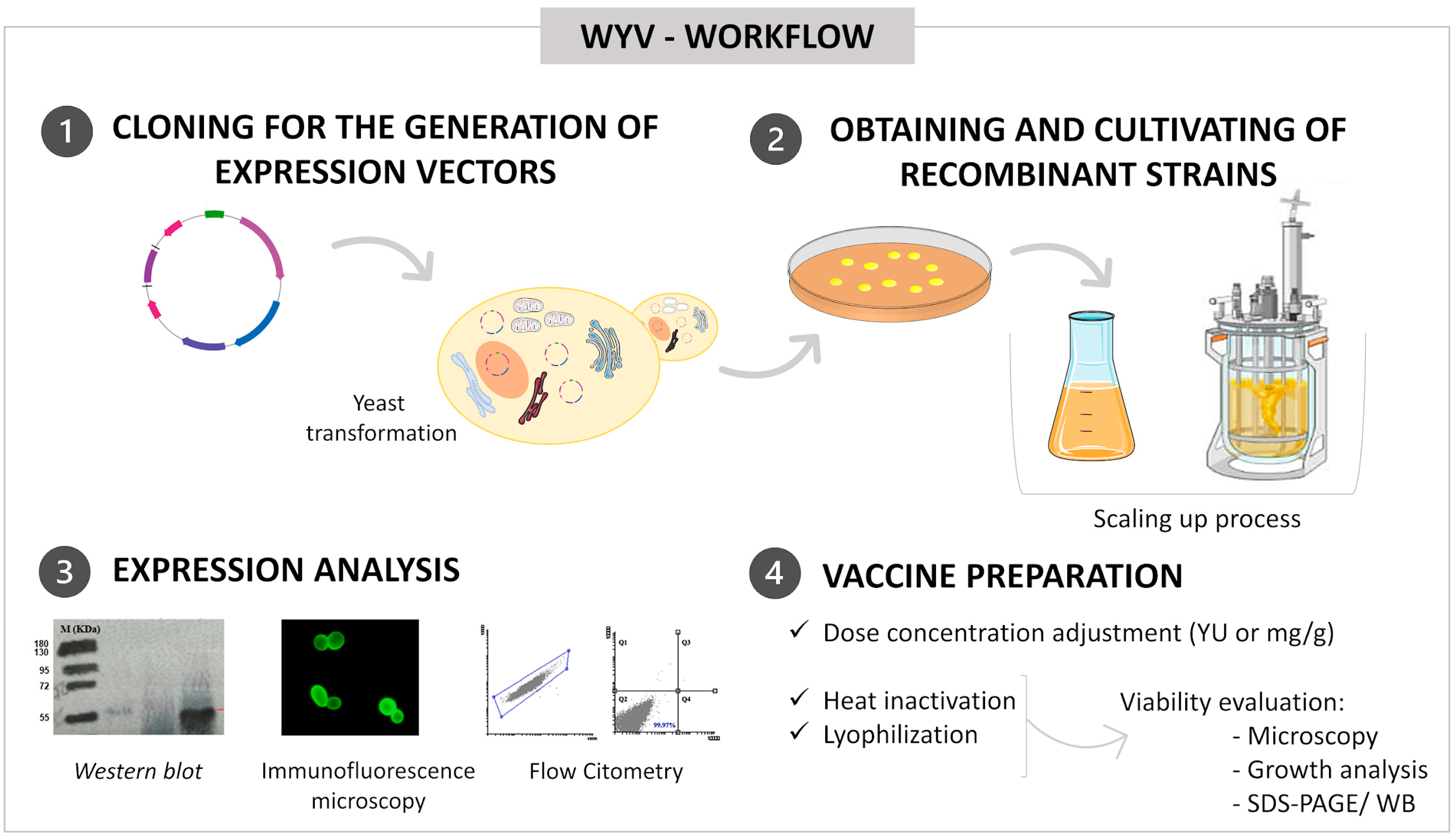
5. Definition of Doses and Quantification
6. Culture Scaling-Up
7. WYVs as a Promising Low-Cost Vaccine Platform
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, R.; Kumar, P. Yeast-based vaccines: New perspective in vaccine development and application. FEMS Yeast Res. 2019, 19, foz007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.-K.; Kim, H.S.; Kim, S.-H.; Oh, J.-M.; Han, J.Y.; Lim, J.M.; Juhnn, Y.-S.; Song, Y.-S. Human papillomavirus type 16 E5 oncoprotein as a new target for cervical cancer treatment. Biochem. Pharmacol. 2010, 80, 1930–1935. [Google Scholar] [CrossRef] [PubMed]
- Roohvand, F.; Shokri, M.; Abdollahpour-Alitappeh, M.; Ehsani, P. Biomedical applications of yeast- a patent view, part one: Yeasts as workhorses for the production of therapeutics and vaccines. Expert Opin. Ther. Pat. 2017, 27, 929–951. [Google Scholar] [CrossRef] [PubMed]
- Lünsdorf, H.; Gurramkonda, C.; Adnan, A.; Khanna, N.; Rinas, U. Virus-like particle production with yeast: Ultrastructural and immunocytochemical insights into Pichia pastoris producing high levels of the Hepatitis B surface antigen. Microb. Cell Factories 2011, 10, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bill, R.M. Recombinant protein subunit vaccine synthesis in microbes: A role for yeast? J. Pharm. Pharmacol. 2015, 67, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Bredell, H.; Smith, J.J.; Görgens, J.F.; van Zyl, W.H. Expression of unique chimeric human papilloma virus type 16 (HPV-16) L1-L2 proteins in Pichia pastoris and Hansenula polymorpha. Yeast 2018, 35, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Pino, M.; Abid, T.; Ribeiro, S.P.; Edara, V.V.; Floyd, K.; Smith, J.C.; Latif, M.B.; Pacheco-Sanchez, G.; Dutta, D.; Wang, S.; et al. A yeast-expressed RBD-based SARS-CoV-2 vaccine formulated with 3M-052-alum adjuvant promotes protective efficacy in non-human primates. Sci. Immunol. 2021, 6, abh3634. [Google Scholar] [CrossRef]
- Gomes, A.M.V.; Carmo, T.S.; Carvalho, L.S.; Bahia, F.M.; Parachin, N.S. Comparison of Yeasts as Hosts for Recombinant Protein Production. Microorganisms 2018, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Yoo, S.J.; Kang, H.A. Yeast synthetic biology for the production of recombinant therapeutic proteins. FEMS Yeast Res. 2014, 15, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ardiani, A.; Higgins, J.P.; Hodge, J.W. Vaccines based on whole recombinant Saccharomyces cerevisiae cells. FEMS Yeast Res. 2010, 10, 1060–1069. [Google Scholar] [CrossRef]
- Shin, M.-K.; Yoo, H.S. Animal vaccines based on orally presented yeast recombinants. Vaccine 2013, 31, 4287–4292. [Google Scholar] [CrossRef] [PubMed]
- Vogl, T.; Hartner, F.S.; Glieder, A. New opportunities by synthetic biology for biopharmaceutical production in Pichia pastoris. Curr. Opin. Biotechnol. 2013, 24, 1094–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlean, P. Architecture and Biosynthesis of the Saccharomyces cerevisiae Cell Wall. Genetics 2012, 192, 775–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukhithasri, V.; Nisha, N.; Biswas, L.; Anil Kumar, V.; Biswas, R. Innate immune recognition of microbial cell wall components and microbial strategies to evade such recognitions. Microbiol. Res. 2013, 168, 396–406. [Google Scholar] [CrossRef]
- Bazan, S.B.; Breinig, T.; Schmitt, M.J.; Breinig, F. Heat treatment improves antigen-specific T cell activation after protein delivery by several but not all yeast genera. Vaccine 2014, 32, 2591–2598. [Google Scholar] [CrossRef]
- Bian, G.; Cheng, Y.; Wang, Z.; Hu, Y.; Zhang, X.; Wu, M.; Chen, Z.; Shi, B.; Sun, S.; Shen, Y.; et al. Whole recombinant Hansenula polymorpha expressing hepatitis B virus surface antigen (yeast-HBsAg) induces potent HBsAg-specific Th1 and Th2 immune responses. Vaccine 2009, 28, 187–194. [Google Scholar] [CrossRef]
- Lei, H.; Xie, B.; Gao, T.; Cen, Q.; Ren, Y. Yeast display platform technology to prepare oral vaccine against lethal H7N9 virus challenge in mice. Microb. Cell Factories 2020, 19, 53. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.J.D.; Jesus, A.L.S.; Leal, L.R.S.; Silva, G.A.S.; Melo, C.M.L.; Freitas, A.C. Pichia pastoris displaying ZIKV protein epitopes from the Envelope and NS1 induce in vitro immune activation. Vaccine 2021, 39, 2545–2554. [Google Scholar] [CrossRef]
- Walch, B.; Breinig, T.; Schmitt, M.J.; Breinig, F. Delivery of functional DNA and messenger RNA to mammalian phagocytic cells by recombinant yeast. Gene Ther. 2012, 19, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Seif, M.; Hoppstädter, J.; Breinig, F.; Kiemer, A.K. Yeast-mediated mRNA delivery polarizes immuno-suppressive macrophages towards an immuno-stimulatory phenotype. Eur. J. Pharm. Biopharm. 2017, 117, 1–13. [Google Scholar] [CrossRef]
- Silva, A.J.D.; de Macêdo, L.S.; Leal, L.R.S.; de Jesus, A.L.S.; Freitas, A.C. Yeasts as a promising delivery platform for DNA and RNA vaccines. FEMS Yeast Res. 2021, 21, foab018. [Google Scholar] [CrossRef] [PubMed]
- Bilusic, M.; Heery, C.R.; Arlen, P.M.; Rauckhorst, M.; Apelian, D.; Tsang, K.Y.; Tucker, J.A.; Jochems, C.; Schlom, J.; Gulley, J.L.; et al. Phase I trial of a recombinant yeast-CEA vaccine (GI-6207) in adults with metastatic CEA-expressing carcinoma. Cancer Immunol. Immunother. 2014, 63, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lok, A.S.; Pan, C.Q.; Han, S.-H.B.; Trinh, H.N.; Fessel, W.J.; Rodell, T.; Massetto, B.; Lin, L.; Gaggar, A.; Subramanian, G.M.; et al. Randomized phase II study of GS-4774 as a therapeutic vaccine in virally suppressed patients with chronic hepatitis B. J. Hepatol. 2016, 65, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Boni, C.; Janssen, H.L.; Rossi, M.; Yoon, S.K.; Vecchi, A.; Barili, V.; Yoshida, E.M.; Trinh, H.; Rodell, T.C.; Laccabue, D.; et al. Combined GS-4774 and Tenofovir Therapy Can Improve HBV-Specific T-Cell Responses in Patients With Chronic Hepatitis. Gastroenterology 2019, 157, 227–241.e7. [Google Scholar] [CrossRef] [Green Version]
- Bazan, S.B.; Geginat, G.; Breinig, T.; Schmitt, M.J.; Breinig, F. Uptake of various yeast genera by antigen-presenting cells and influence of subcellular antigen localization on the activation of ovalbumin-specific CD8 T lymphocytes. Vaccine 2011, 29, 8165–8173. [Google Scholar] [CrossRef]
- Kim, H.; Lee, J.; Kang, H.; Lee, Y.; Park, E.-J. Oral immunization with whole yeast producing viral capsid antigen provokes a stronger humoral immune response than purified viral capsid antigen. Lett. Appl. Microbiol. 2014, 58, 285–291. [Google Scholar] [CrossRef]
- Bazan, S.B.; Walch-Rückheim, B.; Schmitt, M.J.; Breinig, F. Maturation and cytokine pattern of human dendritic cells in response to different yeasts. Med. Microbiol. Immunol. 2018, 207, 75–81. [Google Scholar] [CrossRef]
- Stewart, G.G. The Structure and Function of the Yeast Cell Wall, Plasma Membrane and Periplasm. In Brewing and Distilling Yeasts; Springer International Publishing: Cham, Switzerland, 2017; pp. 55–75. [Google Scholar] [CrossRef]
- Soutter, F.; Werling, D.; Nolan, M.; Küster, T.; Attree, E.; Marugán-Hernández, V.; Kim, S.; Tomley, F.M.; Blake, D.P. A Novel Whole Yeast-Based Subunit Oral Vaccine Against Eimeria tenella in Chickens. Front. Immunol. 2022, 13, 809711. [Google Scholar] [CrossRef]
- Jacob, D.; Ruffie, C.; Dubois, M.; Combredet, C.; Amino, R.; Formaglio, P.; Gorgette, O.; Pehau-Arnaudet, G.; Guery, C.; Puijalon, O.; et al. Whole Pichia pastoris Yeast Expressing Measles Virus Nucleoprotein as a Production and Delivery System to Multimerize Plasmodium Antigens. PLoS ONE 2014, 9, e86658. [Google Scholar] [CrossRef]
- Kiflmariam, M.G.; Yang, H.; Zhang, Z. Gene delivery to dendritic cells by orally administered recombinant Saccharomyces cerevisiae in mice. Vaccine 2013, 31, 1360–1363. [Google Scholar] [CrossRef]
- Arnold, M.; Durairaj, V.; Mundt, E.; Schulze, K.; Breunig, K.D.; Behrens, S.-E. Protective Vaccination against Infectious Bursal Disease Virus with Whole Recombinant Kluyveromyces lactis Yeast Expressing the Viral VP2 Subunit. PLoS ONE 2012, 7, e42870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Kharbikar, B.N. Lyophilized yeast powder for adjuvant free thermostable vaccine delivery. Appl. Microbiol. Biotechnol. 2021, 105, 3131–3143. [Google Scholar] [CrossRef] [PubMed]
- Capilla, J.; Clemons, K.V.; Liu, M.; Levine, H.B.; Stevens, D.A. Saccharomyces cerevisiae as a vaccine against coccidioidomycosis. Vaccine 2009, 27, 3662–3668. [Google Scholar] [CrossRef] [PubMed]
- Patterson, R.; Eley, T.; Browne, C.; Martineau, H.M.; Werling, D. Oral application of freeze-dried yeast particles expressing the PCV2b Cap protein on their surface induce protection to subsequent PCV2b challenge in vivo. Vaccine 2015, 33, 6199–6205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, A.B.; Kilgore, C.; McGlynn, M.; Jones, C.H. Improving global vaccine accessibility. Curr. Opin. Biotechnol. 2016, 42, 67–73. [Google Scholar] [CrossRef]
- Park, K.; Kwon, I.C.; Park, K. Oral protein delivery: Current status and future prospect. React. Funct. Polym. 2011, 71, 280–287. [Google Scholar] [CrossRef]
- Kumar, R.; Srivastava, V.; Baindara, P.; Ahmad, A. Thermostable vaccines: An innovative concept in vaccine development. Expert Rev. Vaccines 2022, 21, 811–824. [Google Scholar] [CrossRef]
- Kumar, R. Investigating the long-term stability of protein immunogen(s) for whole recombinant yeast-based vaccines. FEMS Yeast Res. 2018, 18, foy071. [Google Scholar] [CrossRef] [Green Version]
- Gaggar, A.; Coeshott, C.; Apelian, D.; Rodell, T.; Armstrong, B.R.; Shen, G.; Subramanian, G.M.; McHutchison, J.G. Safety, tolerability and immunogenicity of GS-4774, a hepatitis B virus-specific therapeutic vaccine, in healthy subjects: A randomized study. Vaccine 2014, 32, 4925–4931. [Google Scholar] [CrossRef]
- Gebauer, M.; Hürlimann, H.C.; Behrens, M.; Wolff, T.; Behrens, S.-E. Subunit vaccines based on recombinant yeast protect against influenza A virus in a one-shot vaccination scheme. Vaccine 2019, 37, 5578–5587. [Google Scholar] [CrossRef]
- Bolhassani, A.; Muller, M.; Roohvand, F.; Motevalli, F.; Agi, E.; Shokri, M.; Rad, M.M.; Hosseinzadeh, S. Whole recombinant Pichia pastoris expressing HPV16 L1 antigen is superior in inducing protection against tumor growth as compared to killed transgenic Leishmania. Hum. Vaccines Immunother. 2014, 10, 3499–3508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xv, Z.; Lv, J.; Jiang, J.; Wang, W.; Feng, F.; Zhang, L.; Xue, X.; Li, W. Effective Neutralizing Antibody Produced in Mice Directly Immunized with Integrated Pichia pastoris Expressing HPV16L1 Protein. Viral Immunol. 2019, 32, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Wasilenko, J.L.; Sarmento, L.; Spatz, S.; Pantin-Jackwood, M. Cell surface display of highly pathogenic avian influenza virus hemagglutinin on the surface of Pichia pastoris cells using α-agglutinin for production of oral vaccines. Biotechnol. Prog. 2009, 26, 542–547. [Google Scholar] [CrossRef]
- Bal, J.; Luong, N.N.; Park, J.; Song, K.-D.; Jang, Y.-S.; Kim, D.-H. Comparative immunogenicity of preparations of yeast-derived dengue oral vaccine candidate. Microb. Cell Factories 2018, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Chi, Z.; Li, J.; Zhang, F.; Li, M.; Yasoda, H.N.; Wu, L. The surface display of haemolysin from Vibrio harveyi on yeast cells and their potential applications as live vaccine in marine fish. Vaccine 2006, 24, 6046–6052. [Google Scholar] [CrossRef]
- Wang, J.; Stenzel, D.; Liu, A.; Liu, D.; Brown, D.; Ambrogelly, A. Quantification of a recombinant antigen in an immuno-stimulatory whole yeast cell-based therapeutic vaccine. Anal. Biochem. 2018, 545, 65–71. [Google Scholar] [CrossRef]
- Cohn, A.; Morse, M.A.; O’Neil, B.; Whiting, S.; Coeshott, C.; Ferraro, J.; Bellgrau, D.; Apelian, D.; Rodell, T.C. Whole Recombinant Saccharomyces cerevisiae Yeast Expressing Ras Mutations as Treatment for Patients With Solid Tumors Bearing Ras Mutations: Results From a Phase 1 Trial. J. Immunother. 2018, 41, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Wansley, E.K.; Chakraborty, M.; Hance, K.W.; Bernstein, M.B.; Boehm, A.L.; Guo, Z.; Quick, D.; Franzusoff, A.; Greiner, J.W.; Schlom, J.; et al. Vaccination with a Recombinant Saccharomyces cerevisiae Expressing a Tumor Antigen Breaks Immune Tolerance and Elicits Therapeutic Antitumor Responses. Clin. Cancer Res. 2008, 14, 4316–4325. [Google Scholar] [CrossRef] [Green Version]
- King, T.H.; Kemmler, C.B.; Guo, Z.; Mann, D.; Lu, Y.; Coeshott, C.; Gehring, A.J.; Bertoletti, A.; Ho, Z.Z.; Delaney, W.; et al. A Whole Recombinant Yeast-Based Therapeutic Vaccine Elicits HBV X, S and Core Specific T Cells in Mice and Activates Human T Cells Recognizing Epitopes Linked to Viral Clearance. PLoS ONE 2014, 9, e101904. [Google Scholar] [CrossRef]
- Tang, Y.Q.; Han, S.Y.; Zheng, H.; Wu, L.; Ueda, M.; Wang, X.N.; Lin, Y. Construction of cell surface-engineered yeasts displaying antigen to detect antibodies by immunofluorescence and yeast-ELISA. Appl. Microbiol. Biotechnol. 2008, 79, 1019–1026. [Google Scholar] [CrossRef]
- Liu, D.-Q.; Lu, S.; Zhang, L.; Huang, Y.-R.; Ji, M.; Sun, X.-Y.; Liu, X.-G.; Liu, R.-T. Yeast-Based Aβ1-15 Vaccine Elicits Strong Immunogenicity and Attenuates Neuropathology and Cognitive Deficits in Alzheimer’s Disease Transgenic Mice. Vaccines 2020, 8, 351. [Google Scholar] [CrossRef] [PubMed]
- Mariz, F.C.; Coimbra, E.C.; Jesus, A.L.S.; Nascimento, L.M.; Torres, F.A.G.; Freitas, A.C. Development of an IP-Free Biotechnology Platform for Constitutive Production of HPV16 L1 Capsid Protein Using the Pichia pastoris PGK1 Promoter. BioMed Res. Int. 2015, 2015, 594120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandermies, M.; Fickers, P. Bioreactor-Scale Strategies for the Production of Recombinant Protein in the Yeast Yarrowia lipolytica. Microorganisms 2019, 7, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De, S.; Mattanovich, D.; Ferrer, P.; Gasser, B. Established tools and emerging trends for the production of recombinant proteins and metabolites in Pichia pastoris. Essays Biochem. 2021, 65, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, S.d.S.; Keshavarz-Moore, E. Pichia pastoris (Komagataella phaffii) as a Cost-Effective Tool for Vaccine Production for Low- and Middle-Income Countries (LMICs). Bioengineering 2021, 8, 119. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-C.; Inwood, S.; Gong, T.; Sharma, A.; Yu, L.-Y.; Zhu, P. Fed-batch high-cell-density fermentation strategies for Pichia pastoris growth and production. Crit. Rev. Biotechnol. 2019, 39, 258–271. [Google Scholar] [CrossRef]
- Hayman, B.; Dennehy, M. Developing-country vaccine manufacturers’ technical capabilities can make a difference in global immunization. Vaccine 2021, 39, 5153–5161. [Google Scholar] [CrossRef]
- Kim, D.; Wu, Y.; Kim, Y.B.; Oh, Y.-K. Advances in vaccine delivery systems against viral infectious diseases. Drug Deliv. Transl. Res. 2021, 11, 1401–1419. [Google Scholar] [CrossRef]
- Tan, Y.; Chen, L.; Li, K.; Lou, B.; Liu, Y.; Liu, Z. Yeast as carrier for drug delivery and vaccine construction. J. Control. Release 2022, 346, 358–379. [Google Scholar] [CrossRef]
- de Moura, I.A.; Silva, A.J.D.; de Macêdo, L.S.; Invenção, M.D.C.V.; de Sousa, M.M.G.; de Freitas, A.C. Enhancing the Effect of Nucleic Acid Vaccines in the Treatment of HPV-Related Cancers: An Overview of Delivery Systems. Pathogens 2022, 11, 1444. [Google Scholar] [CrossRef]
- Hudson, L.E.; Fasken, M.B.; McDermott, C.D.; McBride, S.M.; Kuiper, E.G.; Guiliano, D.B.; Corbett, A.H.; Lamb, T.J. Functional Heterologous Protein Expression by Genetically Engineered Probiotic Yeast Saccharomyces boulardii. PLoS ONE 2014, 9, e112660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundstrom, K. Application of Viral Vectors for Vaccine Development with a Special Emphasis on COVID-19. Viruses 2020, 12, 1324. [Google Scholar] [CrossRef] [PubMed]
- Travieso, T.; Li, J.; Mahesh, S.; Mello, J.D.F.R.E.; Blasi, M. The use of viral vectors in vaccine development. NPJ Vaccines 2022, 7, 75. [Google Scholar] [CrossRef] [PubMed]
- Franzusoff, A.; Duke, R.C.; King, T.H.; Lu, Y.; Rodell, T.C. Yeasts encoding tumour antigens in cancer immunotherapy. Expert Opin. Biol. Ther. 2005, 5, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Habersetzer, F.; Baumert, T.F.; Stoll-Keller, F. GI-5005, a yeast vector vaccine expressing an NS3-core fusion protein for chronic HCV infection. Curr. Opin. Mol. Ther. 2009, 11, 456–462. [Google Scholar] [PubMed]
- Heery, C.R.; Singh, B.H.; Rauckhorst, M.; Marté, J.L.; Donahue, R.N.; Grenga, I.; Rodell, T.C.; Dahut, W.; Arlen, P.M.; Madan, R.A.; et al. Phase I Trial of a Yeast-Based Therapeutic Cancer Vaccine (GI-6301) Targeting the Transcription Factor Brachyury. Cancer Immunol. Res. 2015, 3, 1248–1256. [Google Scholar] [CrossRef] [Green Version]
- King, T.H.; Shanley, C.A.; Guo, Z.; Bellgrau, D.; Rodell, T.; Furney, S.; Henao-Tamayo, M.; Orme, I.M. GI-19007, a Novel Saccharomyces cerevisiae-Based Therapeutic Vaccine against Tuberculosis. Clin. Vaccine Immunol. 2017, 24, e00245-17. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.J.D.; Rocha, C.K.d.S.; de Freitas, A.C. Standardization and Key Aspects of the Development of Whole Yeast Cell Vaccines. Pharmaceutics 2022, 14, 2792. https://doi.org/10.3390/pharmaceutics14122792
Silva AJD, Rocha CKdS, de Freitas AC. Standardization and Key Aspects of the Development of Whole Yeast Cell Vaccines. Pharmaceutics. 2022; 14(12):2792. https://doi.org/10.3390/pharmaceutics14122792
Chicago/Turabian StyleSilva, Anna Jéssica Duarte, Crislaine Kelly da Silva Rocha, and Antonio Carlos de Freitas. 2022. "Standardization and Key Aspects of the Development of Whole Yeast Cell Vaccines" Pharmaceutics 14, no. 12: 2792. https://doi.org/10.3390/pharmaceutics14122792