
Nanoparticle-Mediated Delivery of STAT3 Inhibitors in the Treatment of Lung Cancer

Abstract
:1. Introduction
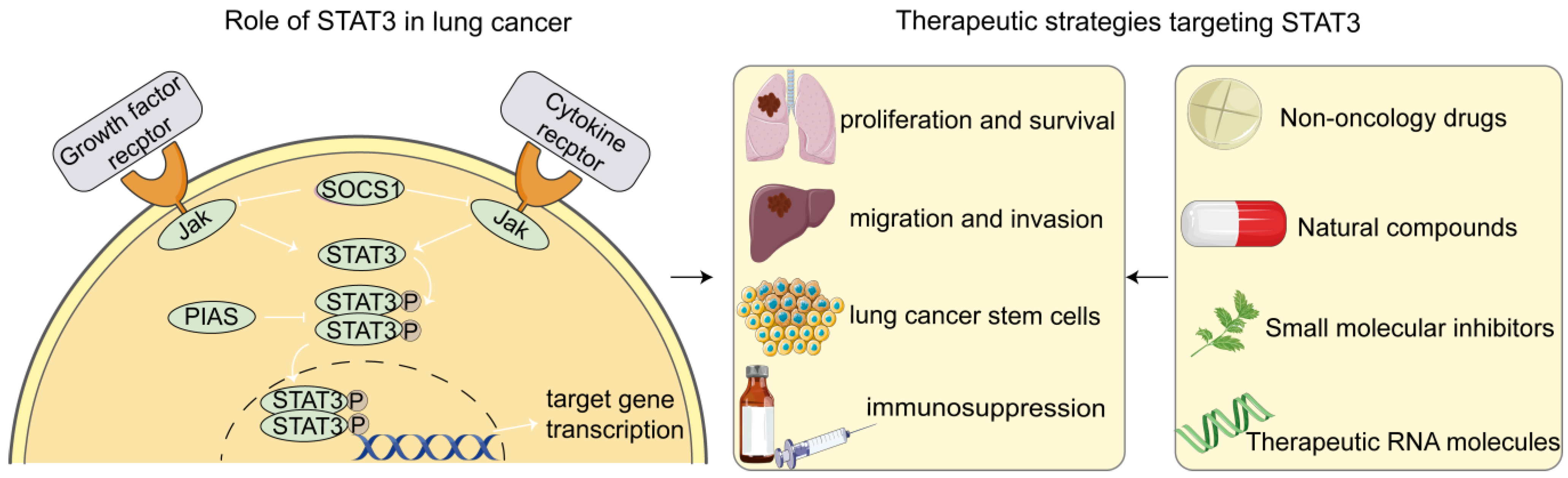
2. Role of STAT3 in Lung Cancer Cells
3. Therapeutic Strategies Targeting STAT3
3.1. Non-Oncology Drugs
3.2. Natural Compounds and Derivatives
3.3. Small Molecule Inhibitors
3.4. Therapeutic RNA Molecules
3.5. Agents Regulating the STAT3 Upstream Gene

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Drug | Mechanism | Therapeutic Effect | R&D Stage |
|---|---|---|---|---|
| Non-oncology drugs | Niclosamide [28] (FDA-approved antihelminthic drug) | Blockage of p-STAT3 binding to the promoter of PD-L1, leading to downregulation of PD-L1 | An enhancement in PD-L1 antibody by niclosamide was observed in the inhibition of NSCLC growth in vitro and in vivo | Preclinical research |
| HJC0152 [35] (Antihelminthic drug, derivative of niclosamide) | Inhibition of the phosphorylation of STAT3 at Tyr705 and alteration of metabolic pathways | HJC0152 reduced the capacity of cells to scavenge free radicals, leading to the generation and accumulation of ROS and inducing apoptosis | Preclinical research | |
| Pyrvinium [36] (FDA-approved antihelminthic drug) | Suppression of STAT3 phosphorylation at tyrosine 705 and serine 727, leading to metabolic lethality in KRAS-mutant lung cancer | Pyrvinium triggered ROS release, depolarized the mitochondrial membrane potential, and suppressed aerobic glycolysis in KRAS-mutant lung cancer cells | Preclinical research | |
| Halofuginone (HF) [37] (Anticoccidials) | Suppression of ERK and STAT3 phosphorylation, and increase in p38 phosphorylation | HF reduced cancer cell viability and induced cell cycle arrest and apoptotic cell death | Phase I clinical trial (NCT00027677) | |
| Acetaminophen (AAP) [38] (FDA-approved antipyretic and analgesic drug) | Suppression of STAT3 activity by directly binding to STAT3 with high affinity | AAP exhibited anti-CSC effects (3D spheroid formation, self-renewal, and expression of CSC markers) both in vitro and in vivo | Preclinical research | |
| Dihydroartemisinin (DHA) [39] (Antimalarial drug) | Suppression of STAT3 phosphorylation, leading to the downregulation of Mcl-1 and Survivin | DHA enhanced ABT-263-induced cytotoxicity via inhibiting STAT3 and triggered apoptosis in NSCLC cells harboring EGFR or RAS mutation | Preclinical research | |
| Natural compounds | Curcumin (CUR) and its derivative [40,41] | Inhibition of phosphorylation of STAT3, JAK, p38, and JNK, as well as their downstream genes (VEGF, Bcl-xL, Cyclin D1) | Curcumin significantly inhibited cell migration and tube formation in vitro and inhibited tumor growth and angiogenesis in vivo | Phase I clinical trial for NSCLC (NCT02321293) |
| Cucurbitacin I (CuI) [42] | Disrupting actin filaments that are physically associated with JAK2 and STAT3 in A549 cells and regulating their phosphorylation | CuI dose-dependently inhibited the phosphorylation of STAT3 and enhanced the phosphorylation of STAT1 in lung adenocarcinoma A549 cells | Preclinical research | |
| Raddeanin A [43] | Inhibition of p-STAT3 and STAT3 expression and reduction of mitochondrial membrane potential by generating ROS | Raddeanin A treatment had no obvious effect on 16HBE cells viability, but it inhibited the viability and proliferation of A549 and H1299 cells | Preclinical research | |
| Ginkgolide C (GGC) [44] | Inhibition of the phosphorylation of STAT3 and STAT3 upstream kinases | GGC exposure significantly reduced NSCLC tumour growth without significant adverse effects by reducing the level of p-STAT3 in mouse tissues | Preclinical research | |
| Eupatolide [45] | Suppression of the activation of STAT3 in NSCLC cells | Eupatolide enhanced the anti-tumor activity of the cisplatin and 5-Fluoracil (5-FU) and suppressed tumor growth in vivo | Preclinical research | |
| Proscillaridin A (PSD-A) [46] | Inhibition of both constitutive and inducible STAT3 activations and reduction of STAT3 DNA-binding activity by increasing SHP-1 expression, reducing Src phosphorylation, and binding PSD-A to STAT3 SH2 domain | PSD-A induced oxidative stress, ER stress, and mitochondrial dysfunction, resulting in apoptosis in A549 cells | Preclinical research | |
| Ethanol extract of Scutellaria baicalensis (ESB) [47] | Inhibition of STAT3 phosphorylation and downregulation of the target gene expression in EGFR TKI-resistant lung cancer cells | ESB reduced cell viability, suppressed colony formation, and induced cell cycle arrest and apoptosis in EGFR TKI-resistant lung cancer cells | Preclinical research | |
| Pterostilbene [48] | Pterostilbene plus Osimertinib reversed osimertinib-induced phosphorylation of STAT3, YAP1, and CUB domain containing protein-1 (CDCP1) | The combination of osimertinib and pterostilbene showed synergistic anti-proliferative effects in all EGFR-mutation-positive NSCLC cell lines | Preclinical research | |
| Small molecular inhibitors | W2014 [49] | Occupation of sub-pockets of the SH2 domain to prevent the dimerization and transcriptional activity of STAT3 | W2014 exhibited potent anti-tumor activities and sensitized resistant NSCLC cells to gefitinib both in vitro and in vivo | Preclinical research |
| BBI608 [51,52,54] (FDA-approved for gastro esophageal junction and pancreatic cancer) | Docking with the SH2 domain of STAT3 SH2 domain leads to disruption of STAT3 phosphorylation and dimerization | BBI608 enhanced the anti-tumor effect of gefitinib on EGFR-mutated NSCLC cells and overcame cisplatin resistance in NSCLC | Phase I, II and III clinical trials for NSCLC (NCT02347917, NCT02826161) | |
| LL1 [53] | Interference with the binding of the SH2 domain affects the phosphorylation of STAT3 | LL1 sensitized the resistance cells to gefitinib with little toxicity both in vitro and in vivo | Preclinical research | |
| ACHP [50] | Interaction with the SH2 domain to inhibit the activation and nuclear localization of STAT3 and to suppress the activation of JAK1, JAK2, and Src | ACHP blocked the proliferation activity of NSCLC, induced apoptosis, and reduced the expression of tumorigenic proteins (survivin, Bcl-2, Bcl-xl, and cyclin D1) | Preclinical research | |
| BP-1-102 [55] | Combination with RAF inhibitor AZ628 markedly abrogated the activation of the MEK/ERK signaling pathway in KRAS-mutant lung cancer cells | The combination of AZ628 and BP-1-102 showed a strong synergistic effect on KRAS-mutant cells and significantly induced cell apoptosis compared with a single inhibitor | Preclinical research | |
| YHO-1701 [56] | Preventing phosphorylated tyrosine peptides from binding to the SH2 domain of STAT3 | STAT3 inhibition by YHO-1701 effectively suppressed the adaptive survival of ALK-rearranged lung cancer cells by enhancing ALK inhibition (alectinib)-induced apoptosis | Preclinical research | |
| MMPP [57] | Binding to the hydroxyl residue of threonine 456 in the DNA-binding domain (DBD) of STAT3 inhibited its phosphorylation and DNA binding activity | MMPP showed anti-tumor activity similar to or better than docetaxel or cisplatin by inducing G1-phase cell cycle arrest and apoptosis | Preclinical research | |
| CS3D [59] | Interruption of STAT3 signaling by binding to STAT3 dimers, rendering them unable to initiate transcription at native STAT3 DNA binding sites | CS3D successfully reduced oncogenic signaling in the airway epithelium of mice exposed to the tobacco carcinogen and reduced immunosuppression | Preclinical research | |
| Therapeutic RNA molecules | miR-4500 [63] | Direct target of the 3′UTR of STAT3 to suppress STAT3 expression | miR-4500 suppressed the cell proliferation, migration, and invasion and promoted apoptosis of human NSCLC cell lines | Preclinical research |
| miR-526b-3p [21] | Direct target of the 3′UTR of STAT3 to suppress STAT3 expression | miR-526b-3p reversed cisplatin resistance, suppressed metastasis, and activated CD8+ T cells in a STAT3/PD-L1-dependent manner | Preclinical research | |
| LncRNA HAR1A [64] | Regulation of the phosphorylation of STAT3 | LncRNA HAR1A inhibited the proliferation of NSCLC cells in vitro and in vivo | Preclinical research | |
| Knockdown of LncRNA BLACAT1 [65] | Suppression of the phosphorylation of STAT3 | Knockdown of LncRNA BLACAT1 reversed the resistance of afatinib to NSCLC cells | Preclinical research | |
| Target upstream regulators of STAT3 | Acetyl-bufalin [66] | Disrupting the formation of CDK9 and STAT3 complex, and reducing the expressions of P-STAT3 and transcribed target genes | Acetyl-bufalin inhibited tumor growth in CDX and PDX models of NSCLC | Preclinical research |
| Palbociclib [67] (FDA-approved for breast cancer) | Inhibiting IL-1β and IL-6 expression, and blocking Src/STAT3 signaling in an RB-independent manner | Palbociclib-induced apoptosis in LUSC cells | Phase I, II and III clinical trials for lung cancer (NCT01291017, NCT03170206, NCT02022982, NCT04870034) | |
| DGG-100629 [68] | Inhibition of STAT3 phosphorylation via the JNK/NFATc1/DDIAS pathway | DGG-100629 inhibited the growth of lung cancer cells and patient-derived gefitinib-resistant lung cancer cells expressing NFATc1 and DDIAS | Preclinical research | |
| Miconazole [69] (FDA-approved antifungal drug) | Disruption of DDIAS/STAT3 interaction to inhibit the phosphorylation of STAT3 tyrosine Y705 and its target genes | Miconazole inhibited the growth and migration of lung cancer cells both in vitro and in vivo | Preclinical research | |
| CP466722 [70] | Suppression of JAK1/2, STAT3, and PD-L1 by inhibiting ATM | CP466722 suppressed EMT and metastatic potential of cisplatin-resistant lung cancer cells | Preclinical research | |
| NCT-80 [71] | Disruption of the interaction between Hsp90 and STAT3 and degradation of STAT3 protein | NCT-80 reduced the viability, colony formation, migration, and CSC-like phenotypes of NSCLC cells and their sublines with acquired resistance to anti-cancer drugs | Preclinical research | |
| miR218 [22] | Target the 3’-UTR of the mRNAs of the IL-6 receptor and JAK3 | miR-218 reduced cell proliferation, invasion, colony formation, and tumor sphere formation in vitro and inhibited tumor growth in vivo | Preclinical research | |
| siRNA PTPRT [72] | Silencing PTPRT with siRNA in NSCLC resulted in increased pSTAT3Tyr705 and upregulation of STAT3 target genes | / | Preclinical research |
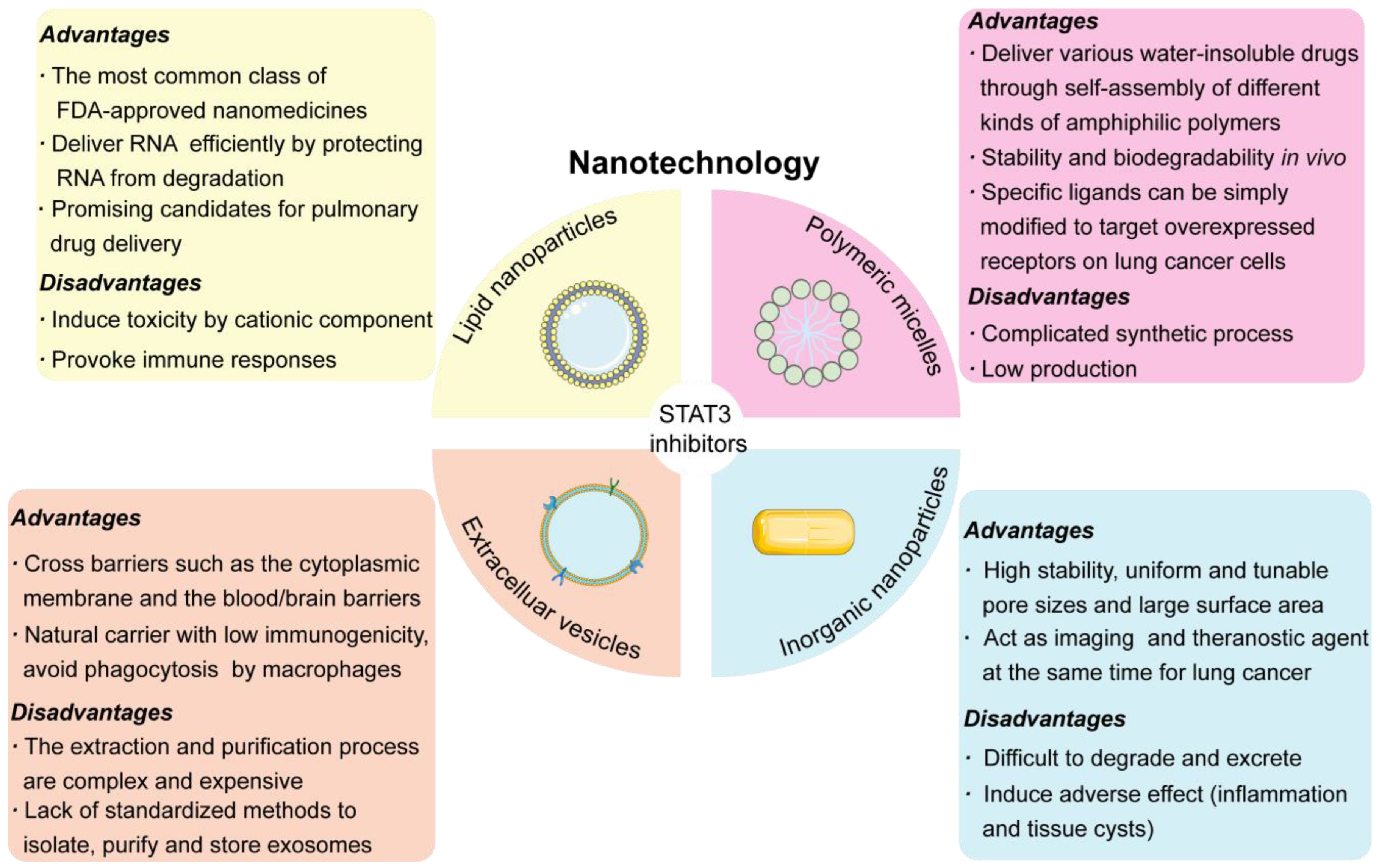
4. Nanoparticle-Based Delivery of STAT3 Inhibitors in the Treatment of Lung Cancer
4.1. LNPs/Liposomes
4.2. Inorganic Nanoparticles
4.3. Polymeric Micelles
4.4. Extracellular Vesicles
4.5. Challenges in Lung Cancer-Targeted Drug Delivery
| Name | Nanoparticles Type | Excipients | STAT3 Inhibitor | Manufacturing Methods | Size and ζ Potential | LE (%) | EE (%) | Modification | Application in Cancer Treatment |
|---|---|---|---|---|---|---|---|---|---|
| STAT3-MB [79] | Cationic lipid microbubbles | DSPC, DSEPC, DSPG and PEG-40 | STAT3 decoy oligonucleotide | DSPC, DSEPC, DSPG, and PEG-40 were dissolved at a molar ratio of 100:43:1:4.5 in chloroform and dried. The dried lipid film was hydrated using isotonic saline and sonicated. The STAT3 decoy was carried via charge–charge interaction. | 2.45 ± 0.35 μm | 10 μg of STAT3 decoy per 1 × 109 microbubbles | / | / | STAT3-MB, in conjunction with UTMC, inhibited tumor growth in HNSCC. |
| WP1066 + STAT3 siRNA-RGDK-lipopeptide [80] | Liposome | DOPC, cholesterol, DSPE-PEG(2000) Amine | STAT3 inhibitor WP1066 and STAT3 siRNA | RGDK tetrapeptides were first synthesized, and then tetrapeptides were coupled with N,N-di-n-hexadecyl-N-2-aminoethylamine. The thin-film hydration method was followed to prepare the liposome. | 138.5 ± 6.4 nm; 8.7 ± 0.4 mV | / | 96– 98% for WP106 and 91% ± 3.2 for siRNA | Modified with RGDK-lipopeptide | The liposome was internalized in glioblastoma cells via integrin α5β1 receptors and accumulated in the brain tissue of glioblastoma-bearing mice, resulting in enhanced anti-cancer efficacy. |
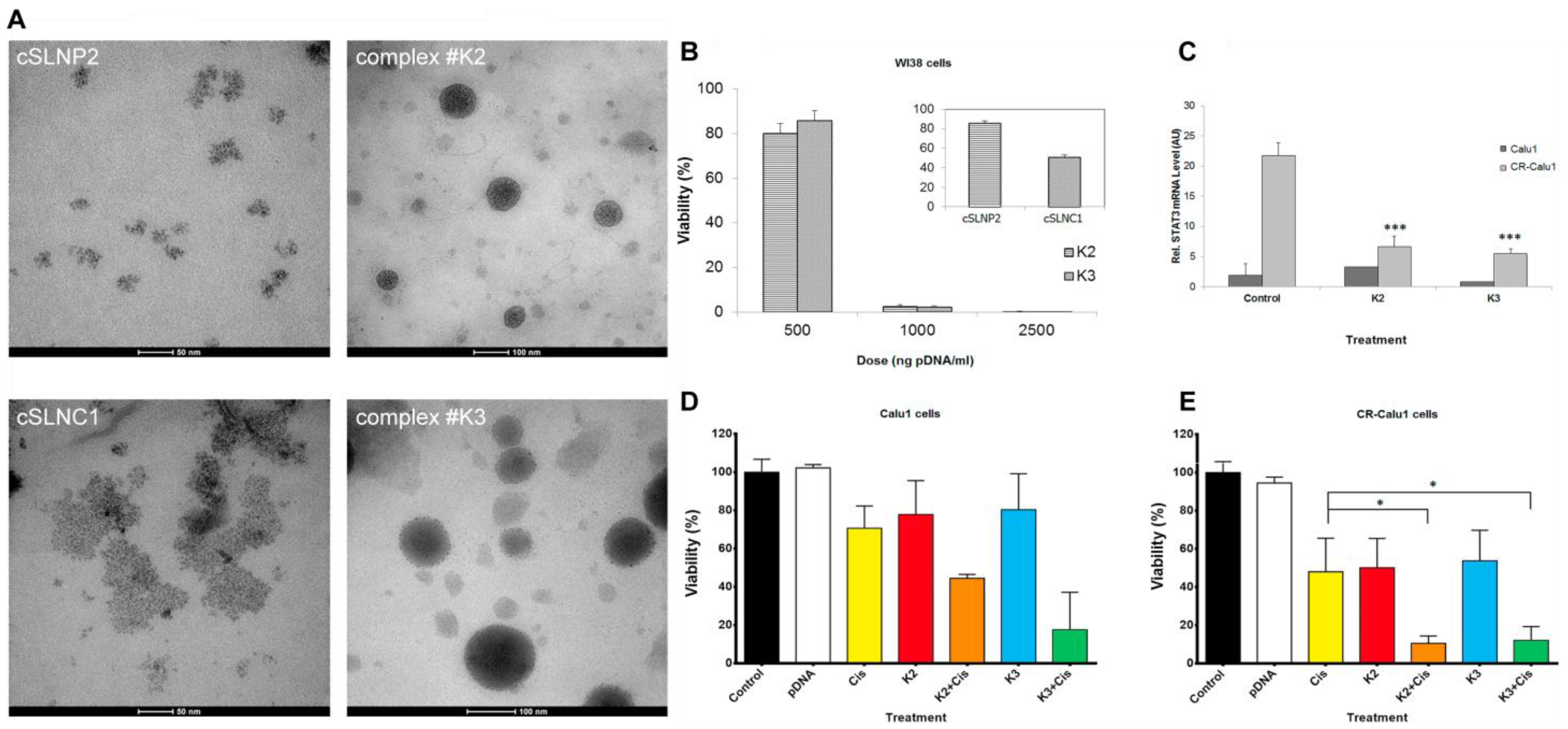
| cSLN:plasmid DNA complexes [81] | Solid lipid nanoparticles | PATO5, C888, CTAB, DDAB and EQ1 | plasmid DNA | Cationic SLNs were prepared by a modified hot microemulsion method, and CTAB, DDAB, or EQ1 were added to provide a positive charge. | 92.6 and 97.8 nm; 10.5 and 8.9 mV | / | / | / | These cSLN:plasmid DNA complexes improved the sensitivity of cisplatin-resistant cells to cisplatin by encoding anti-STAT3 short hairpin RNA. |
| CA-LCL-αCD163 [82] | Liposome | HSPC, cholesterol and mPEG2000-DSPE | STAT3 inhibitor corosolic acid | HSPC, cholesterol, and mPEG2000-DSPE were dissolved at a molar ratio of 55:40:5 in methanol and dialyzed. The lipids were then incubated with CA for 60 min at 65 °C. The lipidated anti-human CD163 antibody was inserted into the CA-LCLs. | 62.0 ± 5.4 nm | / | / | Modified with CD163 | CA-LCL-αCD163 may target macrophages with high CD163 expression and reprogram TAMs from an M2-like phenotype towards an M1-like phenotype. |
| DOX/CALP [83] | Liposome | HSPC, cholesterol and mPEG2000-DSPE | Chemotherapeutic drug DOX | HSPC, cholesterol, and mPEG2000-DSPE were mixed at a molar ratio of 56.3: 38.4: 5.3. CALP was prepared by thin film hydration methods via replacing cholesterol in cholesterol liposomes. The doxorubicin HCl was mixed with CALP for 30 min. | 97.00 ± 3.50 nm; −21.3 ± 2.46 mV | 10.76% ± 0.17 | 98.39% ± 0.51 | Cholesterol in the liposome was replaced by corosolic acid | DOX/CALP displayed strong cytotoxicity, suppressed intratumoral macrophages, and inhibited tumor growth in vivo. |
| CL4H6-LNPs [84] | Lipid nanoparticles | cationic lipid CL4H6, cholesterol, DMG-PEG2000 and DSG-PEG2000 | STAT3 siRNA and HIF-1α siRNA | The LNPs were prepared by alcohol dilution procedure at a molar ratio of 60/40/1 mol% of CL4H6, cholesterol and PEG-lipid. siRNA was mixed with LNPs via the electrostatic interaction under acidic conditions. | 94.09 ± 3.10 nm; 0.21 ± 0.63 mV | / | 92.50% ± 1.25 | / | CL4H6-LNPs increased the levels of infiltrated macrophages (CD11b+ cells) into the TME and had a tendency to increase the concentration of M1 macrophages (CD169+ cells). |
| NP-Stattic-IL20RA [85] | Liposome | DOPE, DOPC and cholesterol | STAT3 inhibitor Stattic | DOPE, DOPC and cholesterol were mixed at a molar ratio of 1:1:1 to form liposomes. IL20RA-PEG conjugates (1:100 molar ratio) and Stattic were then mixed with liposomes. | 120.4 ± 2.91 nm; −11.3 ± 0.092 mV | / | / | Modified with monoclonal anti-IL20RA antibody | NP-Stattic-IL20RA targeted IL20RA+ tumor cells to regulate the expression of PD-L1 via the JAK1-STAT3-SOX2 signaling pathway, inhibit the stemness, and modulate the immune microenvironment. |
| LP-R/C@AC NPs [86] | Liposome | Lecithin, cholesterol, DSPE-PEOz, | STAT3 inhibitor Cinobufagin and VEGFR-2 TKIs Apatinib | The drug-loaded liposome was formed via the thin-film hydration method (Lecithin, cholesterol, DSPEPEOz, AP, and CS-1 at a molar ratio of 2: 1: 0.4: 2: 0.006–0.05) and was mixed with hybrid membrane (R/C) for 3 h. | 108 nm; −7.5 mV | / | 94.2% for Apatinib and 99.9% for Cinobufagin | Coated with red blood cell membrane and cancer cell membrane | LP-R/C@AC NPs efficiently killed tumor cells and reversed tumor immunosuppression by inhibiting the VEGFR2/STAT3 signal pathway. |
| Au-CP [90] | Gold nanoparticle | HAuCl4 | STAT3 inhibitor Curcumin alone or combined with chemotherapeutic drug Paclitaxel | HAuCl4 (200 μL, 10−2 M) solution was added to a 4.8 mL water fraction of Curcumin and Paclitaxel, and Au-CP was purified by ultracentrifugation. | 128 ± 10 nm; −3.0 ± 1.1 | / | / | / | Au-CP demonstrated an excellent synergistic effect on breast cancer by downregulating the expression of STAT3 and its targeted genes both in vitro and in vivo. |
| LbL-AuNP [91] | Gold nanoparticle | HAuCl4·3H2O | STAT3 siRNA and TKI imatinib mesylate | AuNP-CS/siRNA particles were incubated with a chitosan solution containing IM and purified by centrifugation. | 197.8 ± 18.7 nm; 46.8 ± 2.7 mV | 19.2% ± 3.0 | 58.3% ± 7.5 | / | LbL-AuNP could be utilized for iontophoresis therapy to enhance the skin penetration and tumor inhibition of melanoma at an early stage. |
| SPION-TMC-ChT-TAT-H [93] | Iron oxide nanoparticles | FeCl3·6H2O, FeCl2·4H2O, chitosan, hyaluronate and TAT peptide | STAT3 siRNA and HIF-1α siRNA | FeCl3·6H2O and FeCl2·4H2O were mixed to form SPION, and then TMC and ChT were coated on SPION via electrostatic reactions. TAT peptide and hyaluronate were subsequently added. Finally, siRNA was incubated with the mixture. | 118 ± 4 nm; 20 ± 1 mV | / | / | Modified with HA and TAT peptide | The NPs targeted and penetrated CD44+ cancer cells, inhibiting STAT3/HIF-1α gene-driven proliferation, migration and metastasis of cancer. |
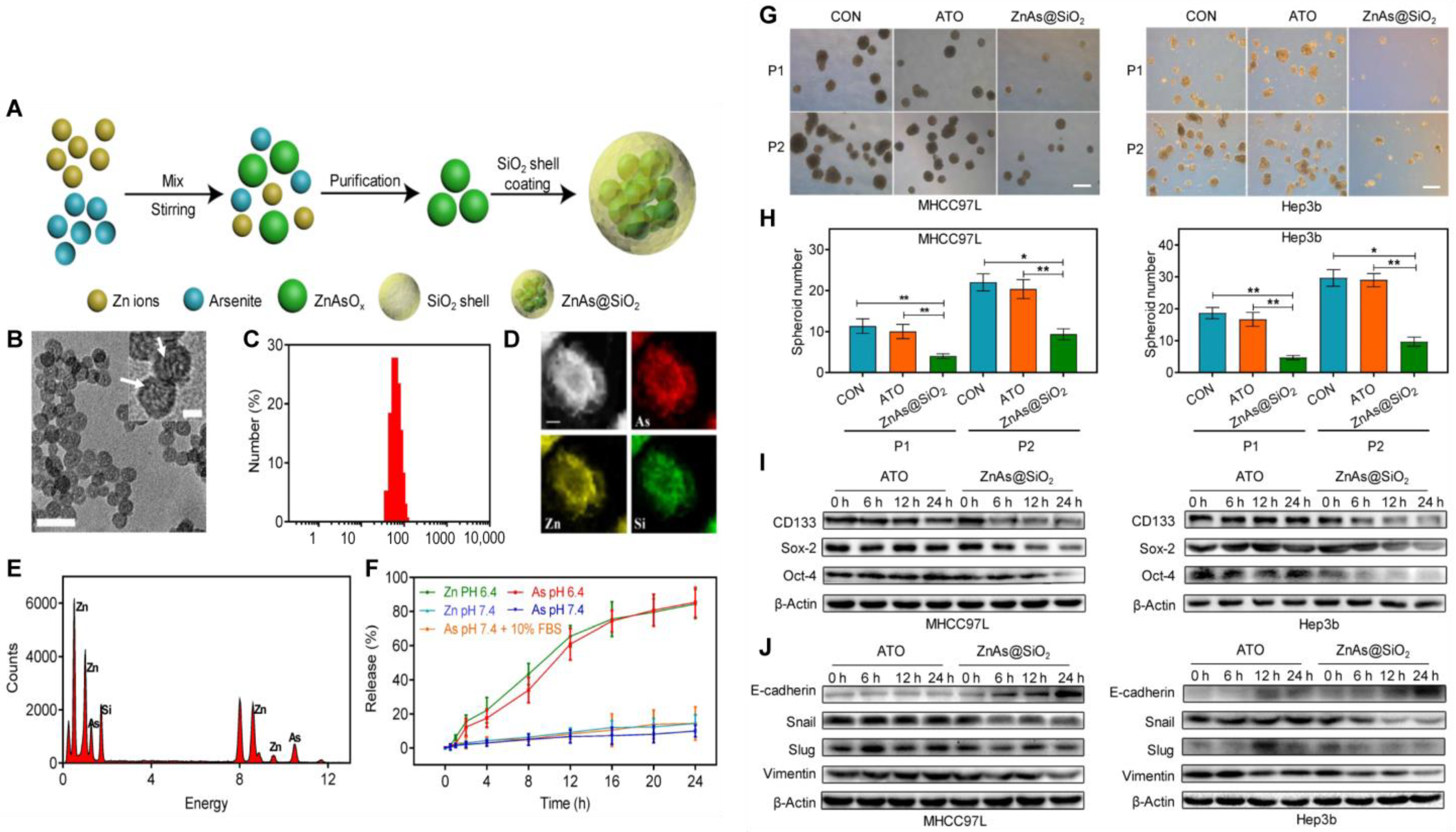
| ZnAs@SiO2 [95] | Silica nanoparticles | ZnCl2, disodium silicate, TEOS and SiO2 | STAT3 inhibitor Arsenic trioxide | Zinc arsenite (ZnAsOx) NPs were synthesized by a reverse microemulsion method, and ZnAsOx NPs were encapsulated in the SiO2 matrix. | 51 ± 3 nm | / | / | / | ZnAs@SiO2 NPs inhibited the formation of tumor spheroids and tumorigenesis and downregulated the expression of stemness and EMT markers by inhibiting the STAT3 signaling pathway. |
| CaP@HA [96] | Calcium phosphate nanoparticles | DOHAP, CaCl2 | STAT3-decoy ODNs | DOHAP were synthesized by conjugating DOAP to the carboxyl groups of HA and CaP were synthesized via an inverse microemulsion system. DOHAP and CaP were mixed, centrifuged and purified. | 51.0 ± 3.4 nm; −18.6 ± 1.9 mV | / | 78% | Modified with HA | CaP@HA effectively suppressed the expression of STAT3 and MUC4, significantly eliminated the interaction between MUC4 with TRAZ and HER2 receptors and efficiently reversed TRAZ resistance in anti-HER2 therapy. |
| CaP@LDL [97] | Calcium phosphate nanoparticles | CaCl2, low-density lipoprotein, DOPC and cholesterol | STAT3-decoy ODNs | CaP were synthesized via an inverse microemulsion system, and low-density lipoprotein was isolated from the plasma of healthy donors. CaP cores were added to the LDL-sucrose residue, together with DOPC and cholesterol, at a molar ratio of 1:1. | 40–45 nm; −4.2 ± 2.1 mV | / | 75% | Modified with low-density lipoprotein | CaP@LDL regulated TRAIL resistance by blocking STAT3 signaling and the expression of downstream anti-apoptotic genes. |
| Chol-DsiRNA Polyplexes [102] | Polymeric micelles | PEG [5K], PLL [30] | STAT3 siRNA | Add the 2X Chol-DsiRNA Complexation Solution dropwise to the 2X Polymer Complexation Solution at 1/1 (v/v), mixing the solution by pipette aspiration/dispensation (30 s) and incubating (RT, 30 min). | / | / | / | / | Chol-DsiRNA Polyplexes were therapeutically active against primary murine syngeneic breast tumors. |
| FA-OCMCS/N-2-HACC/si-STAT3 [103] | Polymeric micelles | Folic acid, OCMCS and N-2- HACC. | STAT3 siRNA | N-2-HACC/siRNA nanocomplexes (N-2-HACC: siRNA = 10:1, w/w) were prepared, and the complex solution was further mixed with FA-OCMCS (FA-OCMCS/N-2-HACC/siRNA = 15:10:1, w/w/w). | 179.8 ± 7.2 nm; −21.56 ± 2.75 mV | / | / | Modified with folic acid | This dual-targeting system shifted M2 macrophages to M1 macrophages and inhibited tumor growth by reducing STAT3 expression in cancer cells and M2 macrophages. |
| ELTN+ FDTN@PEG-PLA [104] | Polymeric micelles | PEG-OH, DGPE | FDTN (JAK2 inhibitor) and ELTN (EGFR-TKI) | PEG-PLA solution was firstly added to DGPE at a polymer/lipid molar ratio of 95:5. ELTN and FDTN solution (20 μL) were then added. The mixture was added dropwise to a citrate buffer solution and dialysed against PBS. | ∼120 nm | 12.2% ± 2.1 for ELTN and 4.2% ± 1.3 for FDTN | / | / | PEG-PLA NPs exhibited synergistic anti-cancer effects in ELTN-resistant NSCLC by inhibiting the JAK-STAT3 signal pathway. |
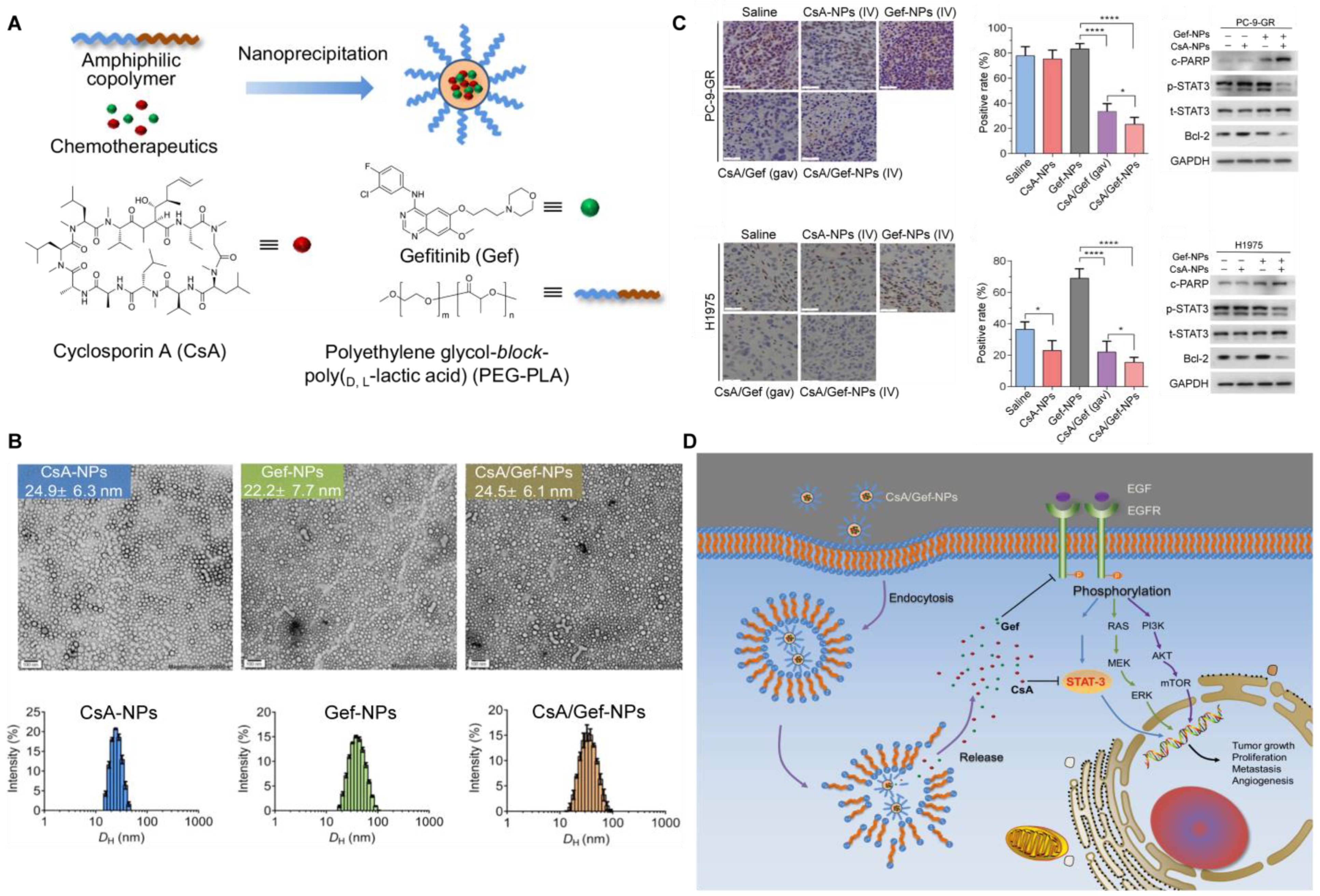
| CSA /Gef–NPs [105] | Polymeric micelles | PEG5k-b-PLA8k | Cyclosporin A (CsA) and gefitinib (Gef, EGFR-TKI) | Polymeric NPs were prepared via the nanoprecipitation method. In total, 1 mg of CsA and 40 mg of PEG-PLA were mixed and added dropwise into DI water then the solution was evaporated and concentrated. | 37.1 ± 13.1 nm; 8.160 ± 9.381 mV | / | / | / | CsA formulated in NPs sensitized Gef-resistant cells and Gef-resistant tumors to Gef treatment by inactivating the STAT3/Bcl-2 signaling pathway. |
| HA/siSTAT3 PPLPTX [106] | Polymeric micelles | PLA, LA and PEI | STAT3 siRNA and chemotherapeutic drug Paclitaxel | After activating the carboxyl group of PLA-LA by HOBt, and EDC, PEI was added dropwise, and the reaction mixture was dialyzed. PTX and siSTAT3 were added dropwise to the micelle solution, and HA was mixed subsequently. | 195 nm; −23 mV | 8.9% | 98.7% | Modified with HA | HA/siSTAT3PPLPTX exhibited anti-tumor efficacy against breast cancer and effectively reduced invasion ability and lung metastasis by silencing the expression of STAT3 and p-STAT3. |
| FM@PFC/siRNA [107] | Polymeric micelles | HMBA, AMD3100, HFBA | STAT3 siRNA and CXCR4 antagonist FM | FM was synthesized by Michael addition of an equimolar ratio of HMBA and AMD3100 and mixed with HFBA. The mixture was added with PFC and siRNA solution. | ∼190 nm; 28 mV | / | / | / | FM@PFC/siRNA induced apoptosis, anti-angiogenic effect, anti-invasive activity, overcame the immunosuppressive TME, and significantly inhibited lung metastasis. |
| P-SS-STAT3/Ca [108] | Polymeric micelles | Pluronic F108 | STAT3 siRNA | The COOP peptide having N-terminal cysteine (CACGLSGLGVA) was conjugated to the Pluronic surface by thiol-exchange reaction utilizing the pyridyl disulfide group in Pluronic F108 | 343.5 nm; −12.6 mV to −0.153 mV | / | / | / | P-SS-STAT3/Ca enhanced the efficiency of STAT3 siRNA transfection, which was successfully demonstrated in cancer cells. |
| siRNA-SS-PNIPAM [109] | Polymeric micelles | NIPAM monomer, RAFT reagent and azide group modified siRNA | STAT3 siRNA and chemotherapeutic drug TMZ | The siRNA-SS-PNIPAM polymers were synthesized by click chemistry via freeze−thaw treatment. | 40.9 nm | 11.33% | 51.12% | / | The cation-free polymeric micelles passed through the BBB and entered brain tumor cells via SRs mediated mechanism and achieved a synergistic effect against TMZ-resistant tumors. |
| DMMA-pro drugs [110] | Polymeric micelles | Nitroimidazole, stearyl chloride and DMMA | STAT3 inhibitor NI-HJC0152 and chemotherapeutic drug GEM-C18 | DMMA were obtained via modifying the side chain of mPEG5000-poly(Lys)-poly(Phe). | around 30 nm | 4.54% ± 0.33 for HJC0152 and 1.23% ± 0.99 for GEM-C18 | 82.3% ± 1.2 for HJC0152 and 90.1% ± 1.3 for GEM-C18 | / | The micelles reversed the immunosuppression of PDAC and relieved the resistance to GEM, resulting in a synergistic effect. |
| SVMAV [111] | Polymeric micelles | peptide-CSSVVR-DHA | STAT3 inhibitor static and TLR receptor 7/8 agonist R848 | Peptide-CSSVVR-DHA and R848-SS-DHA were synthesized, respectively, and they were mixed with static at a ratio of 1: 1: 1. | ~100 nm; 40.83 ± 1.52 mV | / | / | / | SVMAV efficiently stimulated DC maturation, enhanced antigen cross-presentation, and had an anti-tumor effect on primary and metastasis melanoma. |
| EVs-L-PGDS [117] | Exosome | Human Umbilical Cord MSCs | Enzyme L-PGDS | Adenovirus encoding L-PGDS (Ad-L-PGDS) and vector (Ad-Vector) were added to the MSC medium; then the conditioned medium was centrifuged and concentrated for EV isolation. | approximately 100 nm | / | / | / | EVs-L-PGDS could inhibit the phosphorylation of STAT3 and the expression of downstream stem cell markers, resulting in an efficient anti-cancer effect on gastric cancer both in vitro and in vivo. |
| NSC (STAT3-CPG ASO) [118] | Exosome | CpG-STAT3ASO conjugates and HB1.F3 NSCs | STAT3 ASO and CPG ASO | NSCs were incubated with CpG-STAT3ASO, and then culture media were collected, centrifuged at 2000× g, filtered through a 0.22 μm filter, followed by EV isolation using ultracentrifugation standard method. | range of 40–200 nm | / | / | / | NSC entered the hypoxic areas of glioma and secondary brain metastases, activated glioma-associated myeloid cells and inhibited tumor progression in mice. |
| CDNVs [119] | Nanovesicles | Cucumbers | / | CDNVs were isolated by centrifuging juice from cucumbers (CsDNVs from sarcocarp and CpDNVs from pericarp) at 100,000× g for 60 min, followed by a typical exosome isolation protocol. | 180 nm and 129 nm; −13.9 mV and −11.3 mV | / | / | / | CDNVs inhibited the proliferation of NSCLC by suppressing STAT3 activation, generating ROS, promoting cell cycle arrest, and activating the caspase pathway. |
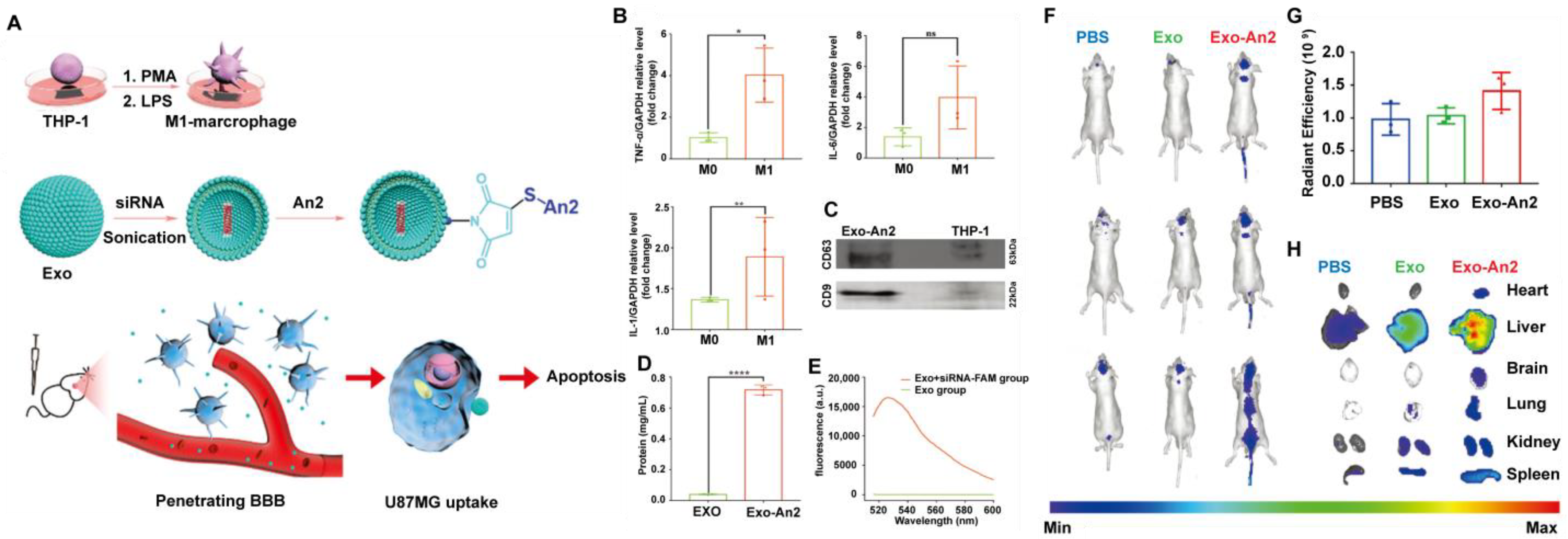
| Exo-An2-siRNA [120] | Exosome | human leukemia monocytic cell line THP-1 | STAT3 siRNA | DSPE-PEG2000-Mal was added into THP-1 cells after induction into M1 macrophages, and the exosomes were extracted using an exoEasy Maxi Kit. siRNA and An2 were added and incubated. | 150 nm; −45 mV | 8.9% | / | Modified with An2 | The exosomes could penetrate BBB and target GBM, resulting in superior anti-cancer therapeutic effects. |
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA 384. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- VanderLaan, P.A.; Roy-Chowdhuri, S. Global impact of the COVID-19 pandemic on cytopathology practice: Results from an international survey of laboratories in 23 countries. Cancer Cytopathol. 2020, 128, 885–894. [Google Scholar]
- Sankar, K.; Gadgeel, S.M.; Qin, A. Molecular therapeutic targets in non-small cell lung cancer. Expert Rev. Anticancer. Ther. 2020, 20, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Du, Y.; Wen, R.; Yang, M.; Xu, J. Drug resistance to targeted therapeutic strategies in non-small cell lung cancer. Pharmacol. Ther. 2020, 206, 107438. [Google Scholar] [CrossRef]
- Zhou, F.; Qiao, M.; Zhou, C. The cutting-edge progress of immune-checkpoint blockade in lung cancer. Cell. Mol. Immunol. 2021, 18, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Mohrherr, J.; Uras, I.Z.; Moll, H.P.; Casanova, E. STAT3: Versatile Functions in Non-Small Cell Lung Cancer. Cancers 2020, 12, 1107. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Mao, R.; Yang, J. NF-κB and STAT3 signaling pathways collaboratively link inflammation to cancer. Protein Cell 2013, 4, 176–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasetyanti, P.R.; Medema, J.P. Intra-tumor heterogeneity from a cancer stem cell perspective. Mol. Cancer 2017, 16, 41. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Xu, S.; Liang, X.; Xue, Y.; Mei, J.; Ma, Y.; Liu, Y.; Liu, Y. Nanotechnology: Breaking the Current Treatment Limits of Lung Cancer. Adv. Healthc. Mater. 2021, 10, e2100078. [Google Scholar] [CrossRef]
- Yu, H.; Lee, H.; Herrmann, A.; Buettner, R.; Jove, R. Revisiting STAT3 signalling in cancer: New and unexpected biological functions. Nat. Rev. Cancer 2014, 14, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Tong, Q.; Liu, B.; Huang, W.; Tian, Y.; Fu, X. Targeting STAT3 in Cancer Immunotherapy. Mol. Cancer 2020, 19, 145. [Google Scholar] [CrossRef] [PubMed]
- Parakh, S.; Ernst, M.; Poh, A.R. Multicellular Effects of STAT3 in Non-small Cell Lung Cancer: Mechanistic Insights and Therapeutic Opportunities. Cancers 2021, 13, 6228. [Google Scholar] [CrossRef]
- Santoni, M.; Miccini, F.; Cimadamore, A.; Piva, F.; Massari, F.; Cheng, L.; Lopez-Beltran, A.; Montironi, R.; Battelli, N. An update on investigational therapies that target STAT3 for the treatment of cancer. Expert Opin. Investig. Drugs 2021, 30, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhao, S.; Shi, Z.; Cao, L.; Liu, J.; Pan, T.; Zhou, D.; Zhang, J. Chemotherapy-elicited exosomal miR-378a-3p and miR-378d promote breast cancer stemness and chemoresistance via the activation of EZH2/STAT3 signaling. J. Exp. Clin. Cancer Res. 2021, 40, 120. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Lee, C.J.; Choi, J.H.; Kim, J.H.; Kim, J.W.; Kim, J.Y.; Nam, J.S. The JAK2/STAT3/CCND2 Axis promotes colorectal Cancer stem cell persistence and radioresistance. J. Exp. Clin. Cancer Res. 2019, 38, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, S.; Li, H.; Lou, L.; Huang, Q.; Zhang, Z.; Mo, J.; Li, M.; Lu, J.; Zhu, K.; Chu, Y.; et al. Inhibition of STAT3-ferroptosis negative regulatory axis suppresses tumor growth and alleviates chemoresistance in gastric cancer. Redox Biol. 2022, 52, 102317. [Google Scholar] [CrossRef]
- Zhang, X.; Sai, B.; Wang, F.; Wang, L.; Wang, Y.; Zheng, L.; Li, G.; Tang, J.; Xiang, J. Hypoxic BMSC-derived exosomal miRNAs promote metastasis of lung cancer cells via STAT3-induced EMT. Mol. Cancer 2019, 18, 40. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Xu, G.; Chang, Z.; Zhu, L.; Yao, J. miR-210 transferred by lung cancer cell-derived exosomes may act as proangiogenic factor in cancer-associated fibroblasts by modulating JAK2/STAT3 pathway. Clin. Sci. 2020, 134, 807–825. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, X.; Mi, Z.; Jiang, X.; Sun, L.; Zheng, B.; Wang, J.; Meng, M.; Zhang, L.; Wang, Z.; et al. STAT3/miR-135b/NF-κB axis confers aggressiveness and unfavorable prognosis in non-small-cell lung cancer. Cell Death Dis. 2021, 12, 493. [Google Scholar] [CrossRef]
- Chen, K.B.; Yang, W.; Xuan, Y.; Lin, A.J. miR-526b-3p inhibits lung cancer cisplatin-resistance and metastasis by inhibiting STAT3-promoted PD-L1. Cell Death Dis. 2021, 12, 748. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ding, L.; Hu, Q.; Xia, J.; Sun, J.; Wang, X.; Xiong, H.; Gurbani, D.; Li, L.; Liu, Y.; et al. MicroRNA-218 functions as a tumor suppressor in lung cancer by targeting IL-6/STAT3 and negatively correlates with poor prognosis. Mol. Cancer 2017, 16, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogan, D.; Grabner, A.; Yanucil, C.; Faul, C.; Ulaganathan, V.K. STAT3-enhancing germline mutations contribute to tumor-extrinsic immune evasion. J. Clin. Investig. 2018, 128, 1867–1872. [Google Scholar] [CrossRef] [PubMed]
- Jing, B.; Wang, T.; Sun, B.; Xu, J.; Xu, D.; Liao, Y.; Song, H.; Guo, W.; Li, K.; Hu, M.; et al. IL6/STAT3 Signaling Orchestrates Premetastatic Niche Formation and Immunosuppressive Traits in Lung. Cancer Res. 2020, 80, 784–797. [Google Scholar] [CrossRef] [Green Version]
- Sp, N.; Kang, D.Y.; Lee, J.M.; Jang, K.J. Mechanistic Insights of Anti-Immune Evasion by Nobiletin through Regulating miR-197/STAT3/PD-L1 Signaling in Non-Small Cell Lung Cancer (NSCLC) Cells. Int. J. Mol. Sci. 2021, 22, 9843. [Google Scholar] [CrossRef]
- Prestipino, A.; Emhardt, A.J.; Aumann, K.; O’Sullivan, D.; Gorantla, S.P.; Duquesne, S.; Melchinger, W.; Braun, L.; Vuckovic, S.; Boerries, M.; et al. Oncogenic JAK2V617F causes PD-L1 expression, mediating immune escape in myeloproliferative neoplasms. Sci. Transl. Med. 2018, 10, eaam7729. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.B.; Wang, W.J.; Xu, C.; Xie, Y.J.; Wang, X.R.; Zhang, Y.Z.; Huang, J.M.; Huang, M.; Xie, C.; Liu, P.; et al. Luteolin and its derivative apigenin suppress the inducible PD-L1 expression to improve anti-tumor immunity in KRAS-mutant lung cancer. Cancer Lett. 2021, 515, 36–48. [Google Scholar] [CrossRef]
- Luo, F.; Luo, M.; Rong, Q.X.; Zhang, H.; Chen, Z.; Wang, F.; Zhao, H.Y.; Fu, L.W. Niclosamide, an antihelmintic drug, enhances efficacy of PD-1/PD-L1 immune checkpoint blockade in non-small cell lung cancer. J. Immunother. Cancer 2019, 7, 245. [Google Scholar] [CrossRef]
- Bhummaphan, N.; Petpiroon, N.; Prakhongcheep, O.; Sritularak, B.; Chanvorachote, P. Lusianthridin targeting of lung cancer stem cells via Src-STAT3 suppression. Phytomedicine 2019, 62, 152932. [Google Scholar] [CrossRef]
- Lee, J.H.; Choi, S.I.; Kim, R.K.; Cho, E.W.; Kim, I.G. Tescalcin/c-Src/IGF1Rβ-mediated STAT3 activation enhances cancer stemness and radioresistant properties through ALDH1. Sci. Rep. 2018, 8, 10711. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Lin, J.H.; Hsu, T.W.; Su, K.; Li, A.F.; Hsu, H.S.; Hung, S.C. IL-6 enriched lung cancer stem-like cell population by inhibition of cell cycle regulators via DNMT1 upregulation. Int. J. Cancer 2015, 136, 547–559. [Google Scholar]
- Zhao, D.; Pan, C.; Sun, J.; Gilbert, C.; Drews-Elger, K.; Azzam, D.J.; Picon-Ruiz, M.; Kim, M.; Ullmer, W.; El-Ashry, D.; et al. VEGF drives cancer-initiating stem cells through VEGFR-2/Stat3 signaling to upregulate Myc and Sox2. Oncogene 2015, 34, 3107–3119. [Google Scholar] [CrossRef]
- Grabner, B.; Schramek, D.; Mueller, K.M.; Moll, H.P.; Svinka, J.; Hoffmann, T.; Bauer, E.; Blaas, L.; Hruschka, N.; Zboray, K.; et al. Disruption of STAT3 signalling promotes KRAS-induced lung tumorigenesis. Nat. Commun. 2015, 6, 6285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Qu, Z.; Yan, S.; Sun, F.; Whitsett, J.A.; Shapiro, S.D.; Xiao, G. Differential roles of STAT3 in the initiation and growth of lung cancer. Oncogene 2015, 34, 3804–3814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Li, H.; Wu, X.; Rao, J.; Zhou, J.; Fan, S.; Shen, Q. HJC0152 suppresses human non-small-cell lung cancer by inhibiting STAT3 and modulating metabolism. Cell Prolif. 2020, 53, e12777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Jiang, W.; Liu, Y.; Huang, W.; Hu, K.; Li, K.; Chen, J.; Ma, C.; Sun, Z.; Pang, X. Blocking STAT3 by pyrvinium pamoate causes metabolic lethality in KRAS-mutant lung cancer. Biochem. Pharmacol. 2020, 177, 113960. [Google Scholar] [CrossRef] [PubMed]
- Demiroglu-Zergeroglu, A.; Turhal, G.; Topal, H.; Ceylan, H.; Donbaloglu, F.; Karadeniz Cerit, K.; Odongo, R.R. Anticarcinogenic effects of halofuginone on lung-derived cancer cells. Cell Biol. Int. 2020, 44, 1934–1944. [Google Scholar] [CrossRef]
- Pingali, P.; Wu, Y.J.; Boothello, R.; Sharon, C.; Li, H.; Sistla, S.; Sankaranarayanan, N.V.; Desai, U.R.; Le, A.T.; Doebele, R.C.; et al. High dose acetaminophen inhibits STAT3 and has free radical independent anti-cancer stem cell activity. Neoplasia 2021, 23, 348–359. [Google Scholar] [CrossRef]
- Yan, X.; Li, P.; Zhan, Y.; Qi, M.; Liu, J.; An, Z.; Yang, W.; Xiao, H.; Wu, H.; Qi, Y.; et al. Dihydroartemisinin suppresses STAT3 signaling and Mcl-1 and Survivin expression to potentiate ABT-263-induced apoptosis in Non-small Cell Lung Cancer cells harboring EGFR or RAS mutation. Biochem. Pharmacol. 2018, 150, 72–85. [Google Scholar] [CrossRef]
- Zhi, T.X.; Liu, K.Q.; Cai, K.Y.; Zhao, Y.C.; Li, Z.W.; Wang, X.; He, X.H.; Sun, X.Y. Anti-Lung Cancer Activities of 1,2,3-Triazole Curcumin Derivatives via Regulation of the MAPK/NF-κB/STAT3 Signaling Pathways. ChemMedChem 2022, 17, e202100676. [Google Scholar] [CrossRef]
- Xu, X.; Zhu, Y. Curcumin inhibits human non-small cell lung cancer xenografts by targeting STAT3 pathway. Am. J. Transl. Res. 2017, 9, 3633–3641. [Google Scholar] [PubMed]
- Guo, H.; Kuang, S.; Song, Q.L.; Liu, M.; Sun, X.X.; Yu, Q. Cucurbitacin I inhibits STAT3, but enhances STAT1 signaling in human cancer cells in vitro through disrupting actin filaments. Acta Pharmacol. Sin. 2018, 39, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, M.; Li, G.; Cai, R. Raddeanin A induced apoptosis of non-small cell lung cancer cells by promoting ROS-mediated STAT3 inactivation. Tissue Cell 2021, 71, 101577. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.H.; Ha, I.J.; Lee, S.G.; Um, J.Y.; Ahn, K.S. Abrogation of STAT3 activation cascade by Ginkgolide C mitigates tumourigenesis in lung cancer preclinical model. J. Pharm. Pharmacol. 2021, 73, 1630–1642. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wu, K.; Xu, A.; Jiao, P.; Li, H.; Xing, L. The sesquiterpene lactone eupatolide induces apoptosis in non-small cell lung cancer cells by suppressing STAT3 signaling. Environ. Toxicol. Pharmacol. 2021, 81, 103513. [Google Scholar] [CrossRef]
- Maryam, A.; Mehmood, T.; Yan, Q.; Li, Y.; Khan, M.; Ma, T. Proscillaridin A Promotes Oxidative Stress and ER Stress, Inhibits STAT3 Activation, and Induces Apoptosis in A549 Lung Adenocarcinoma Cells. Oxid. Med. Cell. Longev. 2018, 2018, 3853409. [Google Scholar] [CrossRef]
- Park, H.J.; Park, S.H.; Choi, Y.H.; Chi, G.Y. The Root Extract of Scutellaria baicalensis Induces Apoptosis in EGFR TKI-Resistant Human Lung Cancer Cells by Inactivation of STAT3. Int. J. Mol. Sci. 2021, 22, 5181. [Google Scholar] [CrossRef]
- Bracht, J.W.P.; Karachaliou, N.; Berenguer, J.; Pedraz-Valdunciel, C.; Filipska, M.; Codony-Servat, C.; Codony-Servat, J.; Rosell, R. Osimertinib and pterostilbene in EGFR-mutation-positive non-small cell lung cancer (NSCLC). Int. J. Biol. Sci. 2019, 15, 2607–2614. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Dong, H.; Mo, J.; Zhang, Y.; Huang, J.; Ouyang, S.; Shi, S.; Zhu, K.; Qu, X.; Hu, W.; et al. A novel STAT3 inhibitor W2014-S regresses human non-small cell lung cancer xenografts and sensitizes EGFR-TKI acquired resistance. Theranostics 2021, 11, 824–840. [Google Scholar] [CrossRef]
- Lee, J.H.; Mohan, C.D.; Basappa, S.; Rangappa, S.; Chinnathambi, A.; Alahmadi, T.A.; Alharbi, S.A.; Kumar, A.P.; Sethi, G.; Ahn, K.S.; et al. The IκB Kinase Inhibitor ACHP Targets the STAT3 Signaling Pathway in Human Non-Small Cell Lung Carcinoma Cells. Biomolecules 2019, 9, 875. [Google Scholar] [CrossRef] [Green Version]
- MacDonagh, L.; Gray, S.G.; Breen, E.; Cuffe, S.; Finn, S.P.; O’Byrne, K.J.; Barr, M.P. BBI608 inhibits cancer stemness and reverses cisplatin resistance in NSCLC. Cancer Lett. 2018, 428, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lu, B.; Zhang, Y.; Yu, J.; Guo, J.; Zhou, Q.; Lv, H.; Sun, Y. STAT3 inhibitor BBI608 enhances the antitumor effect of gefitinib on EGFR-mutated non-small cell lung cancer cells. Hum. Cell 2021, 34, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ma, L.; Sun, Y.; Yu, W.; Wang, X. Targeting STAT3 signaling overcomes gefitinib resistance in non-small cell lung cancer. Cell Death Dis. 2021, 12, 561. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.R.; Wang, F.; Shi, X.W.; Tan, Y.X.; Zhao, J.Y.; Zhang, J.W.; Li, Q.H.; Lin, G.Q.; Gao, D.; Tian, P. Design, synthesis and biological evaluation of novel potent STAT3 inhibitors based on BBI608 for cancer therapy. Eur. J. Med. Chem. 2020, 201, 112428. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yin, M.; Chu, P.; Lou, M. STAT3 inhibitor sensitized KRAS-mutant lung cancers to RAF inhibitor by activating MEK/ERK signaling pathway. Aging 2019, 11, 7187–7196. [Google Scholar] [CrossRef]
- Yanagimura, N.; Takeuchi, S.; Fukuda, K.; Arai, S.; Tanimoto, A.; Nishiyama, A.; Ogo, N.; Takahashi, H.; Asai, A.; Watanabe, S.; et al. STAT3 inhibition suppresses adaptive survival of ALK-rearranged lung cancer cells through transcriptional modulation of apoptosis. NPJ Precis. Oncol. 2022, 6, 11. [Google Scholar] [CrossRef]
- Son, D.J.; Zheng, J.; Jung, Y.Y.; Hwang, C.J.; Lee, H.P.; Woo, J.R.; Baek, S.Y.; Ham, Y.W.; Kang, M.W.; Shong, M.; et al. MMPP Attenuates Non-Small Cell Lung Cancer Growth by Inhibiting the STAT3 DNA-Binding Activity via Direct Binding to the STAT3 DNA-Binding Domain. Theranostics 2017, 7, 4632–4642. [Google Scholar] [CrossRef]
- Dammes, N.; Peer, D. Paving the Road for RNA Therapeutics. Trends Pharmacol. Sci. 2020, 41, 755–775. [Google Scholar] [CrossRef]
- Njatcha, C.; Farooqui, M.; Almotlak, A.A.; Siegfried, J.M. Prevention of Tobacco Carcinogen-Induced Lung Tumor Development by a Novel STAT3 Decoy Inhibitor. Cancer Prev. Res. 2020, 13, 735–746. [Google Scholar] [CrossRef]
- Stoleriu, M.G.; Steger, V.; Mustafi, M.; Michaelis, M.; Cinatl, J.; Schneider, W.; Nolte, A.; Kurz, J.; Wendel, H.P.; Schlensak, C.; et al. A new strategy in the treatment of chemoresistant lung adenocarcinoma via specific siRNA transfection of SRF, E2F1, Survivin, HIF and STAT3. Eur. J. Cardiothorac. Surg. 2014, 46, 877–886. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.S.; Yeh, C.A.; Huang, I.C.; Huang, G.Y.; Chiu, C.H.; Mahalakshmi, B.; Wen, S.Y.; Huang, C.Y.; Kuo, W.W. Signal transducer and activator of transcription 3 mediates apoptosis inhibition through reducing mitochondrial ROS and activating Bcl-2 in gemcitabine-resistant lung cancer A549 cells. J. Cell Physiol. 2021, 236, 3896–3905. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Crabill, G.A.; Pritchard, T.S.; McMiller, T.L.; Wei, P.; Pardoll, D.M.; Pan, F.; Topalian, S.L. Mechanisms regulating PD-L1 expression on tumor and immune cells. J. Immunother. Cancer 2019, 7, 305. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Y.; Zhang, Z.Z.; Bi, H.; Zhang, Q.D.; Zhang, S.J.; Zhou, L.; Zhu, X.Q.; Zhou, J. MicroRNA-4500 suppresses tumor progression in non-small cell lung cancer by regulating STAT3. Mol. Med. Rep. 2019, 20, 4973–4983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Cao, K.; Ling, X.; Zhang, P.; Zhu, J. LncRNA HAR1A Suppresses the Development of Non-Small Cell Lung Cancer by Inactivating the STAT3 Pathway. Cancers 2022, 14, 2845. [Google Scholar] [CrossRef] [PubMed]
- Shu, D.; Xu, Y.; Chen, W. Knockdown of lncRNA BLACAT1 reverses the resistance of afatinib to non-small cell lung cancer via modulating STAT3 signalling. J. Drug Target. 2020, 28, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhou, F.; Zhuang, Y.; Liu, Y.; Xu, L.; Zhao, H.; Xiang, Y.; Dai, X.; Liu, Z.; Huang, X.; et al. Acetyl-bufalin shows potent efficacy against non-small-cell lung cancer by targeting the CDK9/STAT3 signalling pathway. Br. J. Cancer 2021, 124, 645–657. [Google Scholar] [CrossRef]
- Xiang, W.; Qi, W.; Li, H.; Sun, J.; Dong, C.; Ou, H.; Liu, B. Palbociclib Induces the Apoptosis of Lung Squamous Cell Carcinoma Cells via RB-Independent STAT3 Phosphorylation. Curr. Oncol. 2022, 29, 5855–5868. [Google Scholar] [CrossRef]
- Im, J.Y.; Kim, B.K.; Yoon, S.H.; Cho, B.C.; Baek, Y.M.; Kang, M.J.; Kim, N.; Gong, Y.D.; Won, M. DGG-100629 inhibits lung cancer growth by suppressing the NFATc1/DDIAS/STAT3 pathway. Exp. Mol. Med. 2021, 53, 643–653. [Google Scholar] [CrossRef]
- Yoon, S.H.; Kim, B.K.; Kang, M.J.; Im, J.Y.; Won, M. Miconazole inhibits signal transducer and activator of transcription 3 signaling by preventing its interaction with DNA damage-induced apoptosis suppressor. Cancer Sci. 2020, 111, 2499–2507. [Google Scholar] [CrossRef]
- Shen, M.; Xu, Z.; Xu, W.; Jiang, K.; Zhang, F.; Ding, Q.; Xu, Z.; Chen, Y. Inhibition of ATM reverses EMT and decreases metastatic potential of cisplatin-resistant lung cancer cells through JAK/STAT3/PD-L1 pathway. J. Exp. Clin. Cancer Res. 2019, 38, 149. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Min, H.Y.; Yong, Y.S.; Ann, J.; Nguyen, C.T.; La, M.T.; Hyun, S.Y.; Le, H.T.; Kim, H.; Kwon, H.; et al. A novel C-terminal heat shock protein 90 inhibitor that overcomes STAT3-Wnt-β-catenin signaling-mediated drug resistance and adverse effects. Theranostics 2022, 12, 105–125. [Google Scholar] [CrossRef] [PubMed]
- Sen, M.; Kindsfather, A.; Danilova, L.; Zhang, F.; Colombo, R.; LaPorte, M.G.; Kurland, B.F.; Huryn, D.M.; Wipf, P.; Herman, J.G. PTPRT epigenetic silencing defines lung cancer with STAT3 activation and can direct STAT3 targeted therapies. Epigenetics 2020, 15, 604–617. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Marioli, M.; Zhang, K. Analytical characterization of liposomes and other lipid nanoparticles for drug delivery. J. Pharm. Biomed. Anal. 2021, 192, 113642. [Google Scholar] [CrossRef] [PubMed]
- Torchilin, V.P. Recent advances with liposomes as pharmaceutical carriers. Nat. Rev. Drug Discov. 2005, 4, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Zhang, Y.; Huang, L. mRNA vaccine for cancer immunotherapy. Mol. Cancer 2021, 20, 41. [Google Scholar] [CrossRef] [PubMed]
- Crintea, A.; Dutu, A.G.; Samasca, G.; Florian, I.A.; Lupan, I.; Craciun, A.M. The Nanosystems Involved in Treating Lung Cancer. Life 2021, 11, 682. [Google Scholar] [CrossRef] [PubMed]
- Rudokas, M.; Najlah, M.; Alhnan, M.A.; Elhissi, A. Liposome Delivery Systems for Inhalation: A Critical Review Highlighting Formulation Issues and Anticancer Applications. Med. Princ. Pract. 2016, 25 (Suppl. S2), 60–72. [Google Scholar] [CrossRef]
- Clancy, J.P.; Dupont, L.; Konstan, M.W.; Billings, J.; Fustik, S.; Goss, C.H.; Lymp, J.; Minic, P.; Quittner, A.L.; Rubenstein, R.C.; et al. Phase II studies of nebulised Arikace in CF patients with Pseudomonas aeruginosa infection. Thorax 2013, 68, 818–825. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, T.; Chen, X.; Qin, B.; Johnson, D.E.; Grandis, J.R.; Villanueva, F.S. STAT3 decoy oligonucleotide-carrying microbubbles with pulsed ultrasound for enhanced therapeutic effect in head and neck tumors. PLoS ONE 2020, 15, e0241465. [Google Scholar] [CrossRef]
- Vangala, V.; Nimmu, N.V.; Khalid, S.; Kuncha, M.; Sistla, R.; Banerjee, R.; Chaudhuri, A. Combating Glioblastoma by Codelivering the Small-Molecule Inhibitor of STAT3 and STAT3siRNA with α5β1 Integrin Receptor-Selective Liposomes. Mol. Pharm. 2020, 17, 1859–1874. [Google Scholar] [CrossRef]
- Kotmakçı, M.; Çetintaş, V.B.; Kantarcı, A.G. Preparation and characterization of lipid nanoparticle/pDNA complexes for STAT3 downregulation and overcoming chemotherapy resistance in lung cancer cells. Int. J. Pharm. 2017, 525, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.N.; Etzerodt, A.; Graversen, J.H.; Holthof, L.C.; Moestrup, S.K.; Hokland, M.; Møller, H.J. STAT3 inhibition specifically in human monocytes and macrophages by CD163-targeted corosolic acid-containing liposomes. Cancer Immunol. Immunother. 2019, 68, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Widjaya, A.S.; Liu, J.; Liu, X.; Long, Z.; Jiang, Y. Cell-penetrating corosolic acid liposome as a functional carrier for delivering chemotherapeutic drugs. Acta Biomater. 2020, 106, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Shobaki, N.; Sato, Y.; Suzuki, Y.; Okabe, N.; Harashima, H. Manipulating the function of tumor-associated macrophages by siRNA-loaded lipid nanoparticles for cancer immunotherapy. J. Control. Release 2020, 325, 235–248. [Google Scholar] [CrossRef]
- Gao, W.; Wen, H.; Liang, L.; Dong, X.; Du, R.; Zhou, W.; Zhang, X.; Zhang, C.; Xiang, R.; Li, N. IL20RA signaling enhances stemness and promotes the formation of an immunosuppressive microenvironment in breast cancer. Theranostics 2021, 11, 2564–2580. [Google Scholar] [CrossRef]
- Long, Y.; Wang, Z.; Fan, J.; Yuan, L.; Tong, C.; Zhao, Y.; Liu, B. A hybrid membrane coating nanodrug system against gastric cancer via the VEGFR2/STAT3 signaling pathway. J. Mater. Chem. B 2021, 9, 3838–3855. [Google Scholar] [CrossRef]
- Wang, F.; Li, C.; Cheng, J.; Yuan, Z. Recent Advances on Inorganic Nanoparticle-Based Cancer Therapeutic Agents. Int. J. Environ. Res. Public Health 2016, 13, 1182. [Google Scholar] [CrossRef] [Green Version]
- Mottaghitalab, F.; Farokhi, M.; Fatahi, Y.; Atyabi, F.; Dinarvand, R. New insights into designing hybrid nanoparticles for lung cancer: Diagnosis and treatment. J. Control. Release 2019, 295, 250–267. [Google Scholar] [CrossRef]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, Applications and Toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Vemuri, S.K.; Halder, S.; Banala, R.R.; Rachamalla, H.K.; Devraj, V.M.; Mallarpu, C.S.; Neerudu, U.K.; Bodlapati, R.; Mukherjee, S.; Venkata, S.G.P.; et al. Modulatory Effects of Biosynthesized Gold Nanoparticles Conjugated with Curcumin and Paclitaxel on Tumorigenesis and Metastatic Pathways-In Vitro and In Vivo Studies. Int. J. Mol. Sci. 2022, 23, 2150. [Google Scholar] [CrossRef]
- Labala, S.; Jose, A.; Chawla, S.R.; Khan, M.S.; Bhatnagar, S.; Kulkarni, O.P.; Venuganti, V.V.K. Effective melanoma cancer suppression by iontophoretic co-delivery of STAT3 siRNA and imatinib using gold nanoparticles. Int. J. Pharm. 2017, 525, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Budi, H.S.; Izadi, S.; Timoshin, A.; Asl, S.H.; Beyzai, B.; Ghaderpour, A.; Alian, F.; Eshaghi, F.S.; Mousavi, S.M.; Rafiee, B.; et al. Blockade of HIF-1α and STAT3 by hyaluronate-conjugated TAT-chitosan-SPION nanoparticles loaded with siRNA molecules prevents tumor growth. Nanomedicine 2021, 34, 102373. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.A.; Choi, S.; Jeon, S.M.; Yu, J. Silica nanoparticle stability in biological media revisited. Sci. Rep. 2018, 8, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Zhou, B.; Luo, H.; Mao, J.; Huang, Y.; Zhang, K.; Mei, C.; Yan, Y.; Jin, H.; Gao, J.; et al. ZnAs@SiO2 nanoparticles as a potential anti-tumor drug for targeting stemness and epithelial-mesenchymal transition in hepatocellular carcinoma via SHP-1/JAK2/STAT3 signaling. Theranostics 2019, 9, 4391–4408. [Google Scholar] [CrossRef]
- Shi, K.; Fang, Y.; Gao, S.; Yang, D.; Bi, H.; Xue, J.; Lu, A.; Li, Y.; Ke, L.; Lin, X.; et al. Inorganic kernel—Supported asymmetric hybrid vesicles for targeting delivery of STAT3-decoy oligonucleotides to overcome anti-HER2 therapeutic resistance of BT474R. J. Control. Release 2018, 279, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Shi, K.; Xue, J.; Fang, Y.; Bi, H.; Gao, S.; Yang, D.; Lu, A.; Li, Y.; Chen, Y.; Ke, L. Inorganic Kernel-Reconstituted Lipoprotein Biomimetic Nanovehicles Enable Efficient Targeting "Trojan Horse" Delivery of STAT3-Decoy Oligonucleotide for Overcoming TRAIL Resistance. Theranostics 2017, 7, 4480–4497. [Google Scholar] [CrossRef] [Green Version]
- Wan, Z.; Zheng, R.; Moharil, P.; Liu, Y.; Chen, J.; Sun, R.; Song, X.; Ao, Q. Polymeric Micelles in Cancer Immunotherapy. Molecules 2021, 26, 1220. [Google Scholar] [CrossRef]
- Sabir, F.; Qindeel, M.; Zeeshan, M.; Ul Ain, Q.; Rahdar, A.; Barani, M.; González, E.; Aboudzadeh, M.A. Onco-Receptors Targeting in Lung Cancer via Application of Surface-Modified and Hybrid Nanoparticles: A Cross-Disciplinary Review. Processes 2021, 9, 621. [Google Scholar] [CrossRef]
- Nguyen, H.X. Targeted Delivery of Surface-Modified Nanoparticles: Modulation of Inflammation for Acute Lung Injury. In Surface Modification of Nanoparticles for Targeted Drug Delivery; Pathaked, Y.V., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 331–353. [Google Scholar]
- Qin, L.; Wu, L.; Jiang, S.; Yang, D.; He, H.; Zhang, F.; Zhang, P. Multifunctional micelle delivery system for overcoming multidrug resistance of doxorubicin. J. Drug Target. 2018, 26, 289–295. [Google Scholar] [CrossRef]
- Ye, Z.; Abdelmoaty, M.M.; Ambardekar, V.V.; Curran, S.M.; Dyavar, S.R.; Arnold, L.L.; Cohen, S.M.; Kumar, D.; Alnouti, Y.; Coulter, D.W.; et al. Preliminary preclinical study of Chol-DsiRNA polyplexes formed with PLL[30]-PEG[5K] for the RNAi-based therapy of breast cancer. Nanomedicine 2021, 33, 102363. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Dou, Y.; Tang, Y.; Zhang, X. Folate receptor-targeted RNAi nanoparticles for silencing STAT3 in tumor-associated macrophages and tumor cells. Nanomedicine 2020, 25, 102173. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhang, F.; Wang, J.; He, H.; Duan, S.; Zhu, R.; Chen, C.; Yin, L.; Chen, Y. Biodegradable Nanoparticles Mediated Co-delivery of Erlotinib (ELTN) and Fedratinib (FDTN) Toward the Treatment of ELTN-Resistant Non-small Cell Lung Cancer (NSCLC) via Suppression of the JAK2/STAT3 Signaling Pathway. Front. Pharmacol. 2018, 9, 1214. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Shi, L.; Ren, L.; Zhou, L.; Li, T.; Qiao, Y.; Wang, H. A nanomedicine approach enables co-delivery of cyclosporin A and gefitinib to potentiate the therapeutic efficacy in drug-resistant lung cancer. Signal Transduct. Target Ther. 2018, 3, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, K.; Gao, Y.; Yin, S.; Yao, Y.; Yu, H.; Wang, G.; Li, J. Co-delivery of paclitaxel and STAT3 siRNA by a multifunctional nanocomplex for targeted treatment of metastatic breast cancer. Acta Biomater. 2021, 134, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, G.; Ding, L.; Wang, Y.; Zhu, C.; Wang, K.; Li, J.; Sun, M.; Oupicky, D. Increased Survival by Pulmonary Treatment of Established Lung Metastases with Dual STAT3/CXCR4 Inhibition by siRNA Nanoemulsions. Mol. Ther. 2019, 27, 2100–2110. [Google Scholar] [CrossRef] [PubMed]
- Kadekar, S.; Nawale, G.N.; Rangasami, V.K.; Le Joncour, V.; Laakkonen, P.; Hilborn, J.; Varghese, O.P.; Oommen, O.P. Redox responsive Pluronic micelle mediated delivery of functional siRNA: A modular nano-assembly for targeted delivery. Biomater. Sci. 2021, 9, 3939–3944. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Qiao, Y.; Ruan, W.; Zhang, D.; Yang, Q.; Wang, G.; Chen, Q.; Zhu, F.; Yin, J.; Zou, Y.; et al. Cation-Free siRNA Micelles as Effective Drug Delivery Platform and Potent RNAi Nanomedicines for Glioblastoma Therapy. Adv. Mater. 2021, 33, e2104779. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, Q.; Wang, Y.; Chu, Y.; Luo, Y.; You, H.; Su, B.; Li, C.; Guo, Q.; Sun, T.; et al. Penetrating Micelle for Reversing Immunosuppression and Drug Resistance in Pancreatic Cancer Treatment. Small 2022, 18, e2107712. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, J.; Chen, X.; Pan, C.; He, Y.; Su, R.; Guo, D.; Yin, S.; Wang, S.; Zhou, L.; et al. Self-assembly nanovaccine containing TLR7/8 agonist and STAT3 inhibitor enhances tumor immunotherapy by augmenting tumor-specific immune response. J. Immunother. Cancer 2021, 9, e003132. [Google Scholar] [CrossRef]
- Bie, N.; Yong, T.; Wei, Z.; Gan, L.; Yang, X. Extracellular vesicles for improved tumor accumulation and penetration. Adv. Drug Deliv. Rev. 2022, 188, 114450. [Google Scholar] [CrossRef] [PubMed]
- Yong, T.; Wei, Z.; Gan, L.; Yang, X. Extracellular-Vesicle-Based Drug Delivery Systems for Enhanced Antitumor Therapies through Modulating the Cancer-Immunity Cycle. Adv. Mater. 2022. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Shi, J.; Xie, J.; Wang, Y.; Sun, J.; Liu, T.; Zhao, Y.; Zhao, X.; Wang, X.; Ma, Y.; et al. Large-scale generation of functional mRNA-encapsulating exosomes via cellular nanoporation. Nat. Biomed. Eng. 2020, 4, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Martin, P.; Fogarty, B.; Brown, A.; Schurman, K.; Phipps, R.; Yin, V.P.; Lockman, P.; Bai, S. Exosome delivered anticancer drugs across the blood-brain barrier for brain cancer therapy in Danio rerio. Pharm. Res. 2015, 32, 2003–2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.Y.; Liu, L.Z.; Dong, M. Progress on pivotal role and application of exosome in lung cancer carcinogenesis, diagnosis, therapy and prognosis. Mol. Cancer 2021, 20, 22. [Google Scholar] [CrossRef] [PubMed]
- You, B.; Jin, C.; Zhang, J.; Xu, M.; Xu, W.; Sun, Z.; Qian, H. MSC-Derived Extracellular Vesicle-Delivered L-PGDS Inhibit Gastric Cancer Progression by Suppressing Cancer Cell Stemness and STAT3 Phosphorylation. Stem. Cells Int. 2022, 2022, 9668239. [Google Scholar] [CrossRef]
- Adamus, T.; Hung, C.Y.; Yu, C.; Kang, E.; Hammad, M.; Flores, L.; Nechaev, S.; Zhang, Q.; Gonzaga, J.M.; Muthaiyah, K.; et al. Glioma-targeted delivery of exosome-encapsulated antisense oligonucleotides using neural stem cells. Mol. Ther. Nucleic Acids 2022, 27, 611–620. [Google Scholar] [CrossRef]
- Chen, T.; Ma, B.; Lu, S.; Zeng, L.; Wang, H.; Shi, W.; Zhou, L.; Xia, Y.; Zhang, X.; Zhang, J.; et al. Cucumber-Derived Nanovesicles Containing Cucurbitacin B for Non-Small Cell Lung Cancer Therapy. Int. J. Nanomed. 2022, 17, 3583–3599. [Google Scholar] [CrossRef]
- Liang, S.F.; Zuo, F.F.; Yin, B.C.; Ye, B.C. Delivery of siRNA based on engineered exosomes for glioblastoma therapy by targeting STAT3. Biomater. Sci. 2022, 10, 1582–1590. [Google Scholar] [CrossRef]
- Kubczak, M.; Michlewska, S.; Bryszewska, M.; Aigner, A.; Ionov, M. Nanoparticles for local delivery of siRNA in lung therapy. Adv. Drug Deliv. Rev. 2021, 179, 114038. [Google Scholar] [CrossRef]
- Lai, S.K.; Wang, Y.Y.; Hanes, J. Mucus-penetrating nanoparticles for drug and gene delivery to mucosal tissues. Adv. Drug Deliv. Rev. 2009, 61, 158–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirch, J.; Schneider, A.; Abou, B.; Hopf, A.; Schaefer, U.F.; Schneider, M.; Schall, C.; Wagner, C.; Lehr, C.M. Optical tweezers reveal relationship between microstructure and nanoparticle penetration of pulmonary mucus. Proc. Natl. Acad. Sci. USA 2012, 109, 18355–18360. [Google Scholar] [CrossRef] [PubMed]
- Ruge, C.A.; Kirch, J.; Cañadas, O.; Schneider, M.; Perez-Gil, J.; Schaefer, U.F.; Casals, C.; Lehr, C.M. Uptake of nanoparticles by alveolar macrophages is triggered by surfactant protein A. Nanomedicine 2011, 7, 690–693. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Jiao, B.; Shi, X.; Valle, R.P.; Fan, Q.; Zuo, Y.Y. Physicochemical properties of nanoparticles regulate translocation across pulmonary surfactant monolayer and formation of lipoprotein corona. ACS Nano 2013, 7, 10525–10533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patton, J.S.; Brain, J.D.; Davies, L.A.; Fiegel, J.; Gumbleton, M.; Kim, K.J.; Sakagami, M.; Vanbever, R.; Ehrhardt, C. The particle has landed--characterizing the fate of inhaled pharmaceuticals. J. Aerosol. Med. Pulm. Drug Deliv. 2010, 23 (Suppl. S2), S71–S87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, A.K.; Chellappan, D.K.; Dua, K.; Mehta, M.; Satija, S.; Singh, I. Patented therapeutic drug delivery strategies for targeting pulmonary diseases. Expert Opin. Ther. Pat. 2020, 30, 375–387. [Google Scholar] [CrossRef]
- Clark, A.G.; Vignjevic, D.M. Modes of cancer cell invasion and the role of the microenvironment. Curr. Opin. Cell Biol. 2015, 36, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, C.; Suares, D.; Yergeri, M.C. Tumor Microenvironment Targeted Nanotherapy. Front. Pharmacol. 2018, 9, 1230. [Google Scholar] [CrossRef]
- Costa, A.; de Souza Carvalho-Wodarz, C.; Seabra, V.; Sarmento, B.; Lehr, C.M. Triple co-culture of human alveolar epithelium, endothelium and macrophages for studying the interaction of nanocarriers with the air-blood barrier. Acta Biomater. 2019, 91, 235–247. [Google Scholar] [CrossRef]
- Li, H.; Yan, W.; Suo, X.; Peng, H.; Yang, X.; Li, Z.; Zhang, J.; Liu, D. Nucleus-targeted nano delivery system eradicates cancer stem cells by combined thermotherapy and hypoxia-activated chemotherapy. Biomaterials 2019, 200, 1–14. [Google Scholar] [CrossRef]
- Kim, D.M.; Kim, M.; Park, H.B.; Kim, K.S.; Kim, D.E. Anti-MUC1/CD44 Dual-Aptamer-Conjugated Liposomes for Cotargeting Breast Cancer Cells and Cancer Stem Cells. ACS Appl. Bio Mater. 2019, 2, 4622–4633. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Q.; Xiao, K. Nanoparticle-Mediated Delivery of STAT3 Inhibitors in the Treatment of Lung Cancer. Pharmaceutics 2022, 14, 2787. https://doi.org/10.3390/pharmaceutics14122787
Feng Q, Xiao K. Nanoparticle-Mediated Delivery of STAT3 Inhibitors in the Treatment of Lung Cancer. Pharmaceutics. 2022; 14(12):2787. https://doi.org/10.3390/pharmaceutics14122787
Chicago/Turabian StyleFeng, Qiyi, and Kai Xiao. 2022. "Nanoparticle-Mediated Delivery of STAT3 Inhibitors in the Treatment of Lung Cancer" Pharmaceutics 14, no. 12: 2787. https://doi.org/10.3390/pharmaceutics14122787