
Lactiplantibacillus plantarum KAU007 Extract Modulates Critical Virulence Attributes and Biofilm Formation in Sinusitis Causing Streptococcus pyogenes

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Characterization of the Lactobacillus Strains
2.2. Growth Conditions of S. pyrogens
2.3. Revival of L. plantarum KAU007
2.4. Preparation of L. plantarum KAU007 Metabolite Extract
2.5. Estimation of Antibacterial Activity of LME
2.6. Estimation of Anti-Biofilm Property of LME against S. pyogenes
2.7. Effect of LME on S. pyogenes Growth
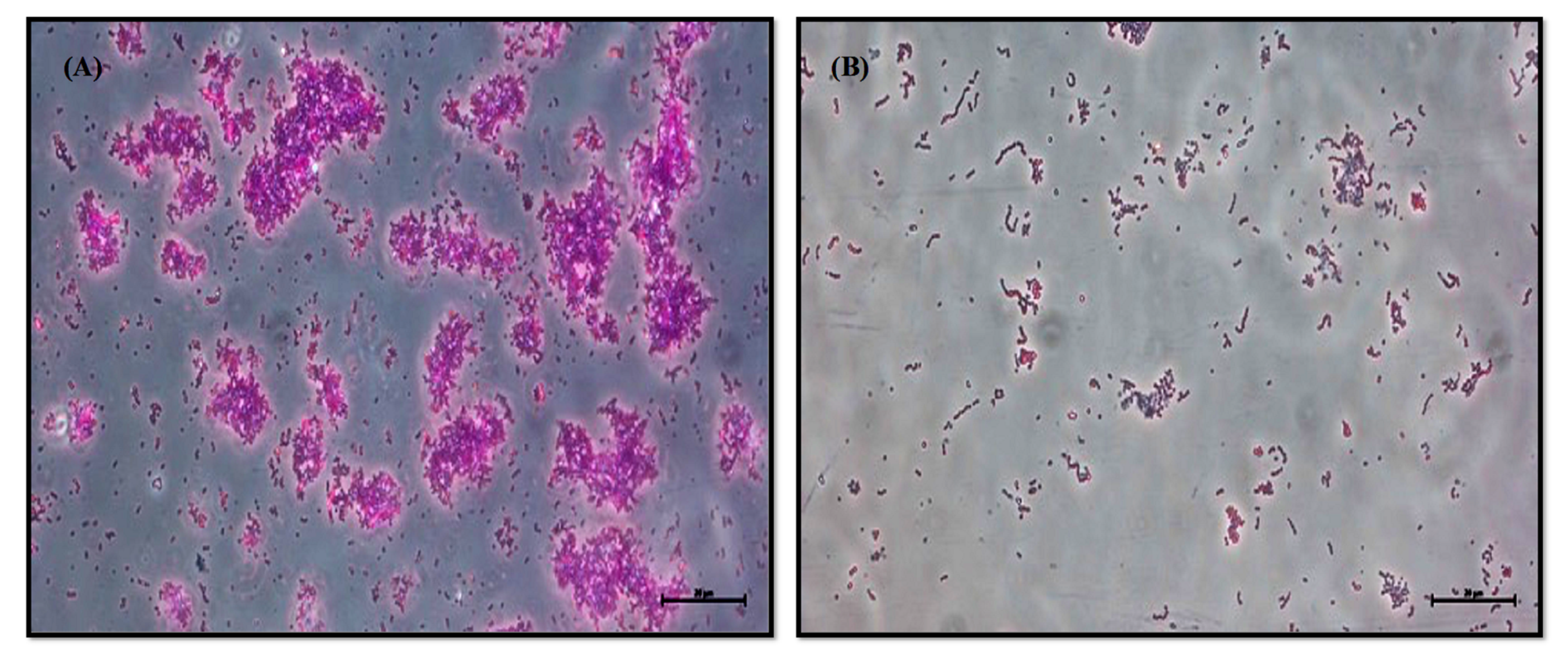
2.8. Light Microscope Micrograph
2.9. Effect of LME on Secreted Protease Activity in S. pyogenes
2.10. Effect of LME on the Hemolytic Property of S. pyogenes
2.11. Effect of LME on the Production of an Extracellular Polymeric Substance in S. pyogenes
2.12. Effect of LME on Microbial Adhesion to Hydrocarbon
2.13. Real-Time PCR Analysis
2.14. GC-MS Analysis
2.15. Statistical Analysis
3. Results and Discussion
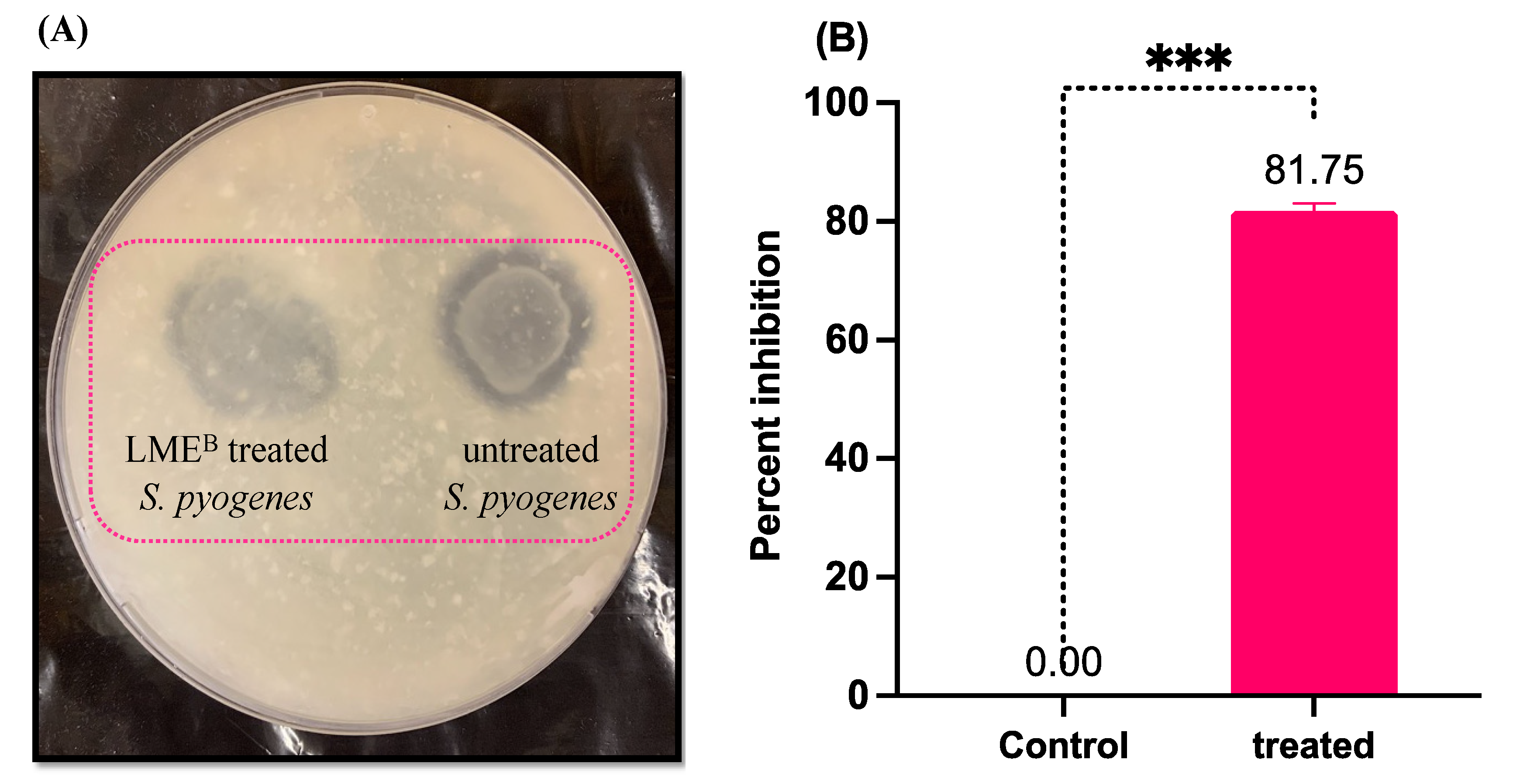
3.1. Antibacterial Activity of LME against S. pyogenes
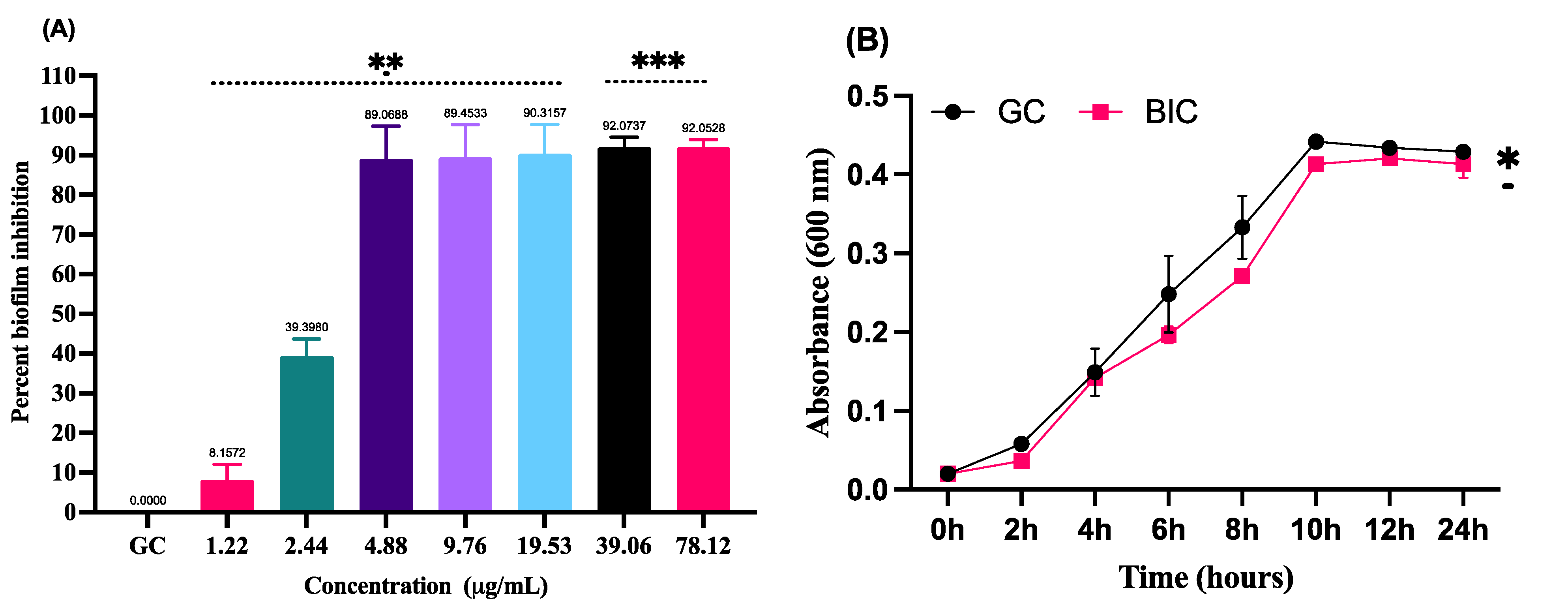
3.2. Inhibition of Biofilm Formation by LME without Hampering the Growth of S. pyogenes
3.3. LME Mitigates Production of Secreted Protease in S. pyogenes
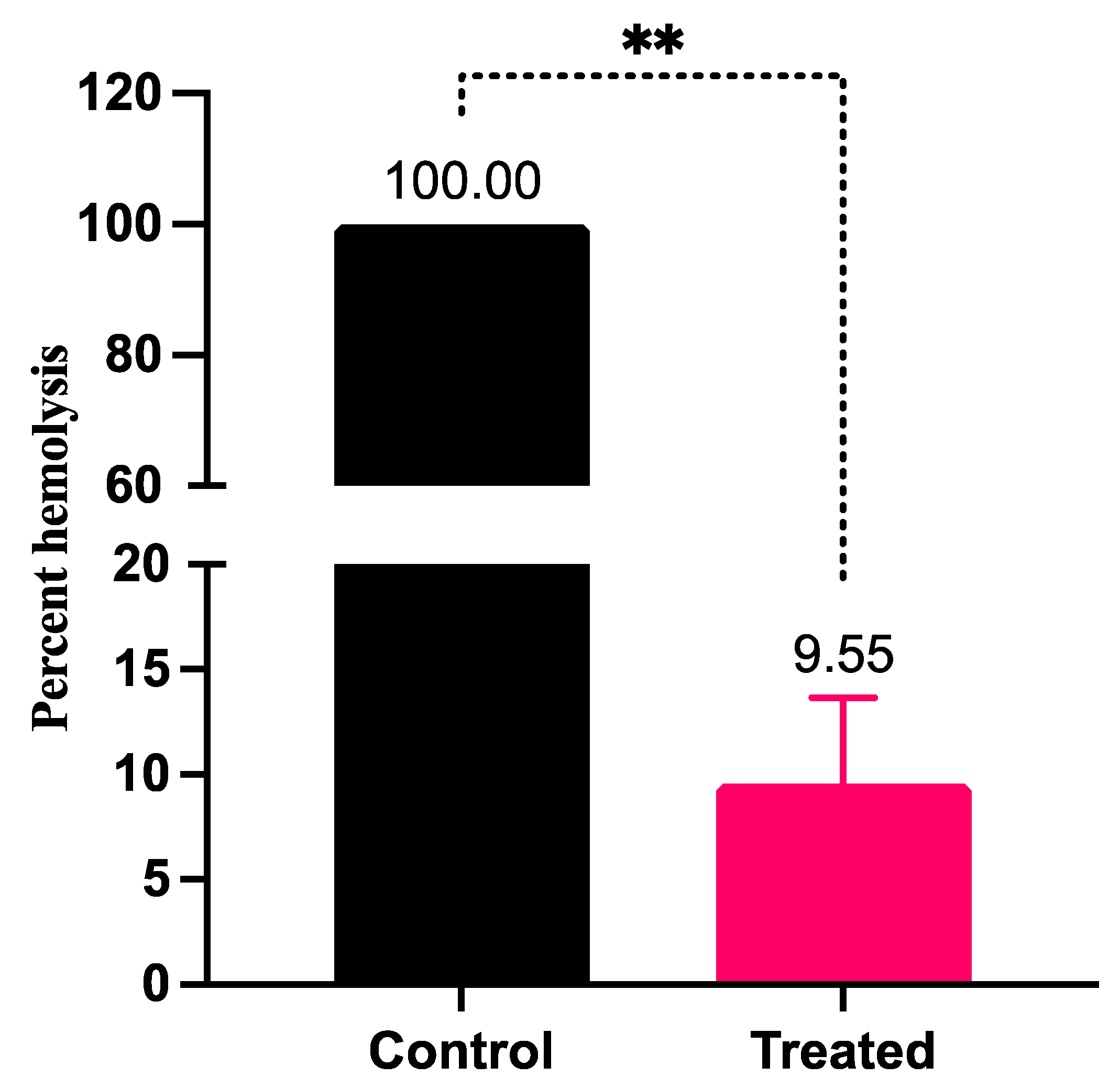
3.4. LME Mitigates Hemolysin Production in S. pyogenes
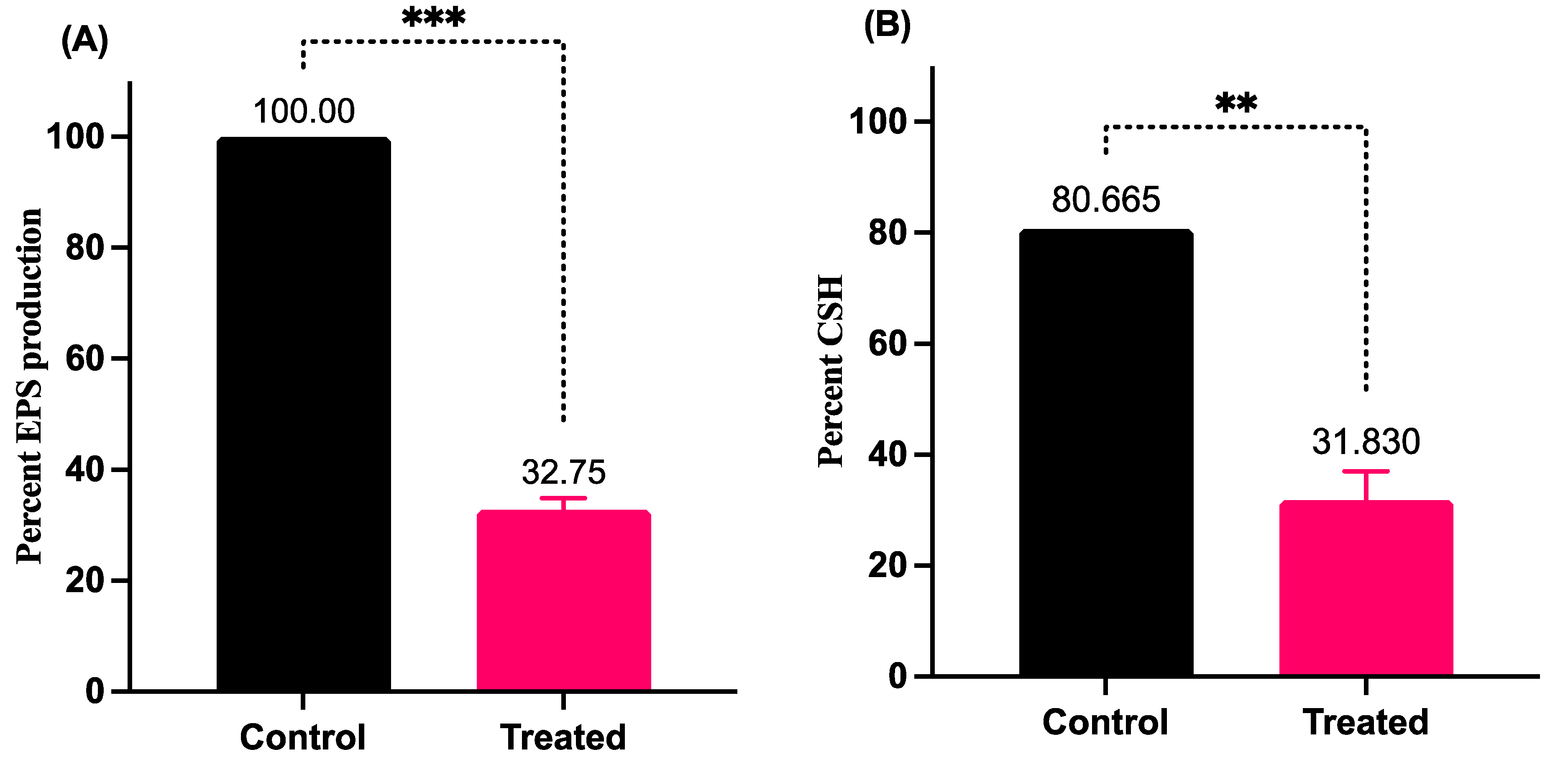
3.5. LME Abrogates the EPS Production and Cell Surface Hydropgobicity in S. pyogenes
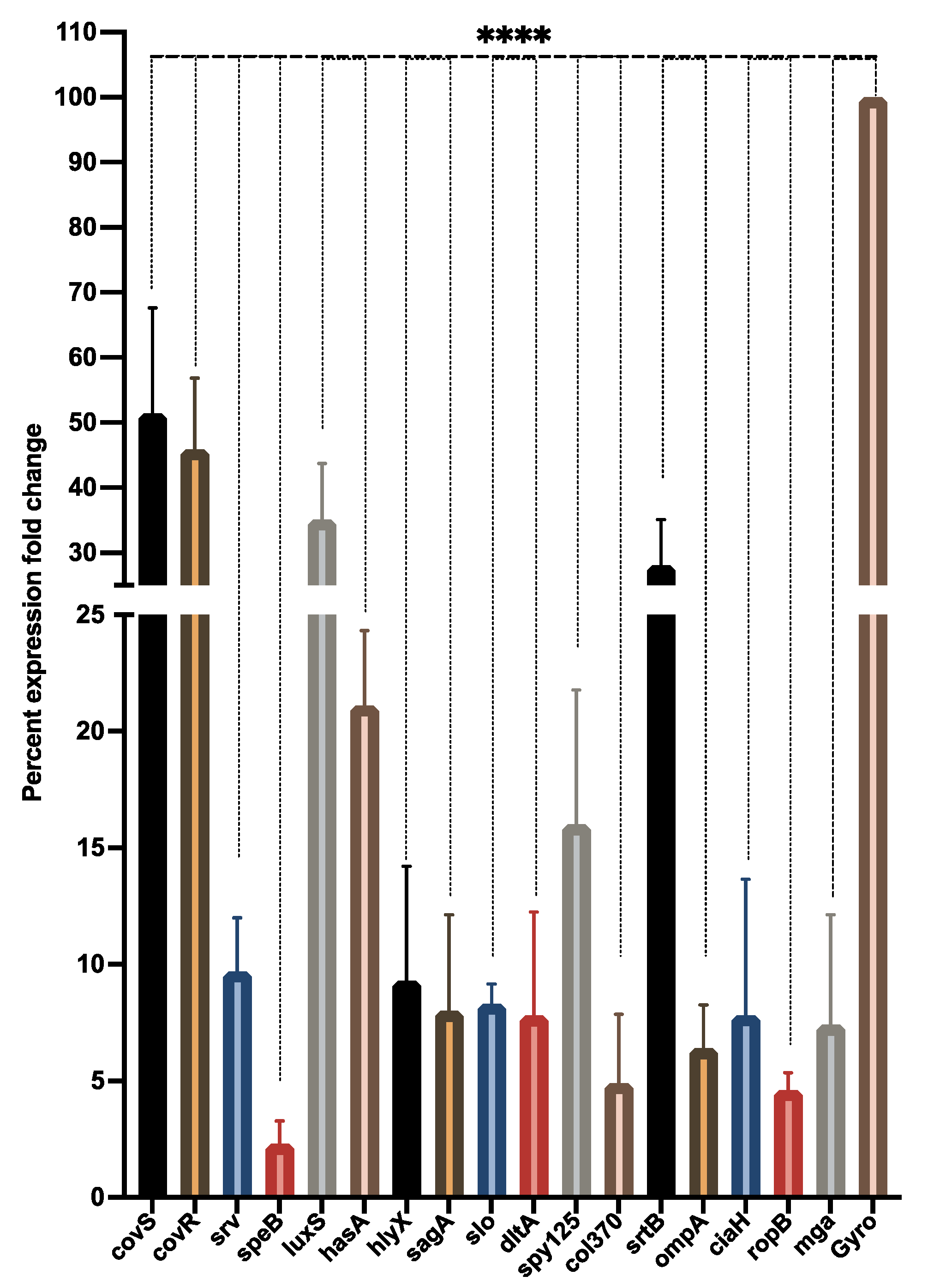
3.6. LME Modulates Expression of Biofilm and Virulence-Associated Genes in S. pyogenes
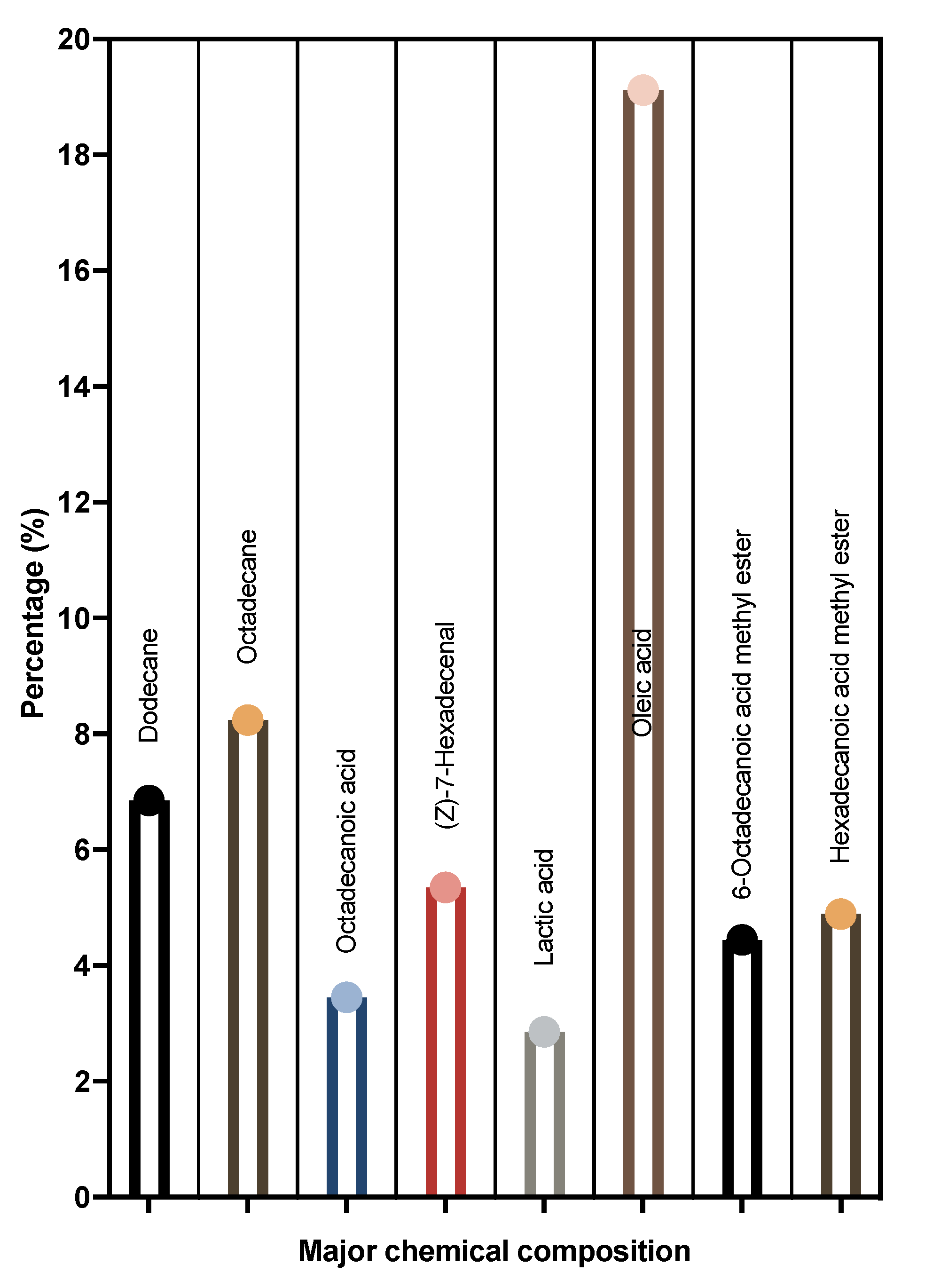
3.7. Chemical Composition of LME
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karlsson, C.A.Q.; Järnum, S.; Winstedt, L.; Kjellman, C.; Björck, L.; Linder, A.; Malmström, J.A. Streptococcus Pyogenes Infection and the Human Proteome with a Special Focus on the Immunoglobulin G-Cleaving Enzyme IdeS. Mol. Cell. Proteom. 2018, 17, 1097–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henningham, A.; Barnett, T.C.; Maamary, P.G.; Walker, M.J. Pathogenesis of Group A Streptococcal Infections. Discov. Med. 2012, 13, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, T.; Köller, T.; Kreikemeyer, B. Streptococcus Pyogenes Biofilms-Formation, Biology, and Clinical Relevance. Front. Cell Infect. Microbiol. 2015, 5, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.K.K.; Eccersley, L.R.J.; Sriskandan, S. Current Views of Haemolytic Streptococcal Pathogenesis. Curr. Opin. Infect. Dis. 2014, 27, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Krzyściak, W.; Pluskwa, K.K.; Jurczak, A.; Kościelniak, D. The Pathogenicity of the Streptococcus Genus. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1361–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golińska, E.; van der Linden, M.; Więcek, G.; Mikołajczyk, D.; Machul, A.; Samet, A.; Piórkowska, A.; Dorycka, M.; Heczko, P.B.; Strus, M. Virulence Factors of Streptococcus Pyogenes Strains from Women in Peri-Labor with Invasive Infections. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Viszwapriya, D.; Subramenium, G.A.; Prithika, U.; Balamurugan, K.; Pandian, S.K. Betulin Inhibits Virulence and Biofilm of Streptococcus Pyogenes by Suppressing RopB Core Regulon, SagA and DltA. Pathog. Dis. 2016, 74, ftw088. [Google Scholar] [CrossRef] [Green Version]
- Vega, L.A.; Malke, H.; McIver, K.S. Virulence-Related Transcriptional Regulators of Streptococcus pyogenes. 2016 Feb 10 [Updated 2016 Mar 25]. In Streptococcus pyogenes: Basic Biology to Clinical Manifestations; Ferretti, J.J., Stevens, D.L., Fischetti, V.A., Eds.; University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016. Available online: https://www.ncbi.nlm.nih.gov/books/NBK333412/ (accessed on 25 March 2016).
- Federle, M. Pathogenic streptococci speak, but what are they saying? Virulence 2012, 3, 92–94. [Google Scholar] [CrossRef] [Green Version]
- Connolly, K.L.; Roberts, A.L.; Holder, R.C.; Reid, S.D. Dispersal of Group a Streptococcal Biofilms by the Cysteine Protease SpeB Leads to Increased Disease Severity in a Murine Model. PLoS ONE 2011, 6, e18984. [Google Scholar] [CrossRef]
- Valdes, K.M.; Sundar, G.S.; Belew, A.T.; Islam, E.; El-Sayed, N.M.; Le Breton, Y.; McIver, K.S. Glucose Levels Alter the Mga Virulence Regulon in the Group A Streptococcus. Sci. Rep. 2018, 8, 4971. [Google Scholar] [CrossRef]
- Yu, D.; Zheng, Y.; Yang, Y. Is There Emergence of β-Lactam Antibiotic-Resistant Streptococcus Pyogenes in China? Infect. Drug Resist. 2020, 13, 2323. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.C.; Shen, C.F.; Lin, Y.L.; Shen, F.C.; Tsai, P.J.; Wang, S.Y.; Lin, Y.S.; Wu, J.J.; Chi, C.Y.; Liu, C.C. Emergence of Macrolide-Resistant Streptococcus Pyogenes Emm12 in Southern Taiwan from 2000 to 2019. J. Microbiol. Immunol. Infect. 2021, 54, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Kebede, D.; Admas, A.; Mekonnen, D. Prevalence and Antibiotics Susceptibility Profiles of Streptococcus Pyogenes among Pediatric Patients with Acute Pharyngitis at Felege Hiwot Comprehensive Specialized Hospital, Northwest Ethiopia. BMC Microbiol. 2021, 21, 135. [Google Scholar] [CrossRef] [PubMed]
- Alves-Barroco, C.; Paquete-Ferreira, J.; Santos-Silva, T.; Fernandes, A.R. Singularities of Pyogenic Streptococcal Biofilms—From Formation to Health Implication. Front. Microbiol. 2020, 11, 584947. [Google Scholar] [CrossRef] [PubMed]
- Subramenium, G.A.; Viszwapriya, D.; Iyer, P.M.; Balamurugan, K.; Pandian, S.K. CovR Mediated Antibiofilm Activity of 3-Furancarboxaldehyde Increases the Virulence of Group a Streptococcus. PLoS ONE 2015, 10, e0127210. [Google Scholar] [CrossRef] [Green Version]
- Fidanza, M.; Panigrahi, P.; Kollmann, T.R. Lactiplantibacillus plantarum–Nomad and Ideal Probiotic. Front. Microbiol. 2021, 12, 2911. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic Resistance: A Rundown of a Global Crisis. Infect. Drug Resist. 2018, 11, 1645. [Google Scholar] [CrossRef] [Green Version]
- Matulay, J.T.; Mlynarczyk, C.M.; Cooper, K.L. Urinary Tract Infections in Women: Pathogenesis, Diagnosis, and Management. Curr. Bladder Dysfunct. Rep. 2016, 11, 53–60. [Google Scholar] [CrossRef]
- Georgieva, R.; Yocheva, L.; Tserovska, L.; Zhelezova, G.; Stefanova, N.; Atanasova, A.; Danguleva, A.; Ivanova, G.; Karapetkov, N.; Rumyan, N.; et al. Antimicrobial Activity and Antibiotic Susceptibility of Lactobacillus and Bifidobacterium Spp. Intended for Use as Starter and Probiotic Cultures. Biotechnol. Biotechnol. Equip. 2015, 29, 84–91. [Google Scholar] [CrossRef]
- Inglin, R.C.; Stevens, M.J.A.; Meile, L.; Lacroix, C.; Meile, L. High-Throughput Screening Assays for Antibacterial and Antifungal Activities of Lactobacillus Species. J. Microbiol. Methods 2015, 114, 26–29. [Google Scholar] [CrossRef]
- Kumar, M.; Dhaka, P.; Vijay, D.; Vergis, J.; Mohan, V.; Kumar, A.; Kurkure, N.V.; Barbuddhe, S.B.; Malik, S.V.S.; Rawool, D.B. Antimicrobial Effects of Lactobacillus Plantarum and Lactobacillus Acidophilus against Multidrug-Resistant Enteroaggregative Escherichia Coli. Int. J. Antimicrob. Agents 2016, 48, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Mirnejad, R.; Vahdati, A.R.; Rashidiani, J.; Erfani, M.; Piranfar, V. The Antimicrobial Effect of Lactobacillus Casei Culture Supernatant against Multiple Drug Resistant Clinical Isolates of Shigella Sonnei and Shigella Flexneri in Vitro. Iran. Red Crescent Med. J. 2013, 15, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, K.B.; Baik, J.E.; Park, O.J.; Yun, C.H.; Han, S.H. Lactobacillus Plantarum Lipoteichoic Acid Inhibits Biofilm Formation of Streptococcus Mutans. PLoS ONE 2018, 13, e0192694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamalifar, H.; Rahimi, H.R.; Samadi, N.; Shaverdi, A.R.; Sharifian, Z.; Hosseini, F.; Eslahi, H.; Fazeli, M.R. Antimicrobial Activity of Different Lactobacillus Species against Multi-Drug Resistant Clinical Isolates of Pseudomonas Aeruginosa. Iran J. Microbiol. 2011, 3, 21–25. [Google Scholar] [PubMed]
- Kang, M.S.; Lim, H.S.; Oh, J.S.; Lim, Y.J.; Wuertz-Kozak, K.; Harro, J.M.; Shirtliff, M.E.; Achermann, Y. Antimicrobial Activity of Lactobacillus Salivarius and Lactobacillus Fermentum against Staphylococcus Aureus. Pathog. Dis. 2017, 75, ftx009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maudsdotter, L.; Jonsson, H.; Roos, S.; Jonsson, A.B. Lactobacilli Reduce Cell Cytotoxicity Caused by Streptococcus Pyogenes by Producing Lactic Acid That Degrades the Toxic Component Lipoteichoic Acid. Antimicrob. Agents Chemother. 2011, 55, 1622–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dempsey, E.; Corr, S.C. Lactobacillus spp. for Gastrointestinal Health: Current and Future Perspectives. Front. Immunol. 2022, 13, 840245. [Google Scholar] [CrossRef]
- Ostad, S.N.; Salarian, A.A.; Ghahramani, M.H.; Fazeli, M.R.; Samadi, N.; Jamalifar, H. Live and Heat-Inactivated Lactobacilli from Feces Inhibit Salmonella Typhi and Escherichia Coli Adherence to Caco-2 Cells. Folia Microbiol. 2009, 54, 157–160. [Google Scholar] [CrossRef]
- Rather, I.A.; Kamli, M.R.; Sabir, J.S.M.; Paray, B.A. Potential Antiviral Activity of Lactiplantibacillus Plantarum KAU007 against Influenza Virus H1N1. Vaccines 2022, 10, 456. [Google Scholar] [CrossRef]
- Badwaik, L.S.; Borah, P.K.; Deka, S.C. Production and Purification of Anti-Bacterial Biometabolite from Wild-Type Lactobacillus, Isolated from Fermented Bamboo Shoot: Future Suggestions and a Proposed System for Secondary Metabolite Onsite Recovery During Continuous Fermentation. Appl. Biochem. Biotechnol. 2015, 175, 1915–1925. [Google Scholar] [CrossRef]
- M100-S16CLSI 2014; Performance Standards for Antimicrobial Susceptibility Testing. 24th Informational Supplement. Clinical and Laboratory Standards (M100-S24): Wayne, PA, USA, 2014.
- Wijesundara, N.M.; Lee, S.F.; Cheng, Z.; Davidson, R.; Rupasinghe, H.P.V. Carvacrol Exhibits Rapid Bactericidal Activity against Streptococcus Pyogenes through Cell Membrane Damage. Sci. Rep. 2021, 11, 1487. [Google Scholar] [CrossRef] [PubMed]
- Viszwapriya, D.; Prithika, U.; Deebika, S.; Balamurugan, K.; Pandian, S.K. In Vitro and in Vivo Antibiofilm Potential of 2,4-Di-Tert-Butylphenol from Seaweed Surface Associated Bacterium Bacillus Subtilis against Group A Streptococcus. Microbiol. Res. 2016, 191, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Subramenium, G.A.; Vijayakumar, K.; Pandian, S.K. Limonene Inhibits Streptococcal Biofilm Formation by Targeting Surface-Associated Virulence Factors. J. Med. Microbiol. 2015, 64, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Hollands, A.; Aziz, R.K.; Kansal, R.; Kotb, M.; Nizet, V.; Walker, M.J. A Naturally Occurring Mutation in RopB Suppresses SpeB Expression and Reduces M1T1 Group A Streptococcal Systemic Virulence. PLoS ONE 2008, 3, e20592. [Google Scholar] [CrossRef] [PubMed]
- Limsuwan, S.; Voravuthikunchai, S.P. Boesenbergia Pandurata (Roxb.) Schltr., Eleutherine Americana Merr. and Rhodomyrtus Tomentosa (Aiton) Hassk. as Antibiofilm Producing and Antiquorum Sensing in Streptococcus Pyogenes. FEMS Immunol. Med. Microbiol. 2008, 53, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Doern, C.D.; Roberts, A.L.; Hong, W.; Nelson, J.; Lukomski, S.; Swords, W.E.; Reid, S.D. Biofilm Formation by Group A Streptococcus: A Role for the Streptococcal Regulator of Virulence (Srv) and Streptococcal Cysteine Protease (SpeB). Microbiology 2009, 155, 46. [Google Scholar] [CrossRef] [Green Version]
- Chaussee, M.S.; Somerville, G.A.; Reitzer, L.; Musser, J.M. Rgg Coordinates Virulence Factor Synthesis and Metabolism in Streptococcus Pyogenes. J. Bacteriol. 2003, 185, 6016–6024. [Google Scholar] [CrossRef] [Green Version]
- Kapur, V.; Topouzis, S.; Majesky, M.W.; Li, L.L.; Hamrick, M.R.; Hamill, R.J.; Patti, J.M.; Musser, J.M. A Conserved Streptococcus Pyogenes Extracellular Cysteine Protease Cleaves Human Fibronectin and Degrades Vitronectin. Microb. Pathog. 1993, 15, 327–346. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.J.; Barnett, T.C.; McArthur, J.D.; Cole, J.N.; Gillen, C.M.; Henningham, A.; Sriprakash, K.S.; Sanderson-Smith, M.L.; Nizet, V. Disease Manifestations and Pathogenic Mechanisms of Group A Streptococcus. Clin. Microbiol. Rev. 2014, 27, 264–301. [Google Scholar] [CrossRef] [Green Version]
- Patenge, N.; Fiedler, T.; Kreikemeyer, B. Common Regulators of Virulence in Streptococci. Curr. Top Microbiol. Immunol. 2013, 368, 111–153. [Google Scholar] [CrossRef]
- Datta, V.; Myskowski, S.M.; Kwinn, L.A.; Chiem, D.N.; Varki, N.; Kansal, R.G.; Kotb, M.; Nizet, V. Mutational Analysis of the Group A Streptococcal Operon Encoding Streptolysin S and Its Virulence Role in Invasive Infection. Mol. Microbiol. 2005, 56, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Cox, K.H.; Ruiz-Bustos, E.; Courtney, H.S.; Dale, J.B.; Pence, M.A.; Nizet, V.; Aziz, R.K.; Gerling, I.; Price, S.M.; Hasty, D.L. Inactivation of DltA Modulates Virulence Factor Expression in Streptococcus Pyogenes. PLoS ONE 2009, 4, e5366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.M.; Tsai, Y.S.; Wu, C.M.; Liao, S.K.; Wu, L.C.; Chang, C.S.; Liu, Y.H.; Tsai, P.J. Streptococcal Collagen-like Surface Protein 1 Promotes Adhesion to the Respiratory Epithelial Cell. BMC Microbiol. 2010, 10, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, K.R.; Nakata, M.; Sumitomo, T.; Kreikemeyer, B.; Podbielski, A.; Terao, Y.; Kawabata, S. Involvement of T6 Pili in Biofilm Formation by Serotype M6 Streptococcus Pyogenes. J. Bacteriol. 2012, 194, 804–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Wood, T.K. OmpA Influences Escherichia Coli Biofilm Formation by Repressing Cellulose Production through the CpxRA Two-Component System. Environ. Microbiol. 2009, 11, 2735–2746. [Google Scholar] [CrossRef] [PubMed]
- Gaddy, J.A.; Tomaras, A.P.; Actis, L.A. The Acinetobacter Baumannii 19606 OmpA Protein Plays a Role in Biofilm Formation on Abiotic Surfaces and in the Interaction of This Pathogen with Eukaryotic Cells. Infect. Immun. 2009, 77, 3150–3160. [Google Scholar] [CrossRef] [Green Version]
- Tatsuno, I.; Isaka, M.; Okada, R.; Zhang, Y.; Hasegawa, T. Relevance of the Two-Component Sensor Protein CiaH to Acid and Oxidative Stress Responses in Streptococcus Pyogenes. BMC Res. Notes 2014, 7, 189. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Rupasinghe, T.W.; Tull, D.L.; Augustin, M.A.; Gras, S.L. Liquid-liquid extraction and liquid chromatography-mass spectrometry detection of curcuminoids from bacterial culture medium. J. Chromatogr. B Analyt. Technol. Biomed. Life. Sci. 2015, 988, 116–120. [Google Scholar] [CrossRef]
- Chen, C.C.; Lai, C.C.; Huang, H.L.; Huang, W.Y.; Toh, H.S.; Weng, T.C.; Chuang, Y.C.; Lu, Y.C.; Tang, H.J. Antimicrobial Activity of Lactobacillus Species against Carbapenem-Resistant Enterobacteriaceae. Front. Microbiol. 2019, 10, 789. [Google Scholar] [CrossRef]
- Saroj, S.D.; Maudsdotter, L.; Tavares, R.; Jonsson, A.B. Lactobacilli Interfere with Streptococcus Pyogenes Hemolytic Activity and Adherence to Host Epithelial Cells. Front. Microbiol. 2016, 7, 1176. [Google Scholar] [CrossRef]
- Dasari, S.; Shouri, R.N.D.; Wudayagiri, R.; Valluru, L. Antimicrobial Activity of Lactobacillus against Microbial Flora of Cervicovaginal Infections. Asian Pac. J. Trop. Dis. 2014, 4, 18–24. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Rather, I.A.; Majumder, R.; Alshammari, F.H.; Nam, G.-J.; Park, Y.-H. Characterization and Antibacterial Mode of Action of Lactic Acid Bacterium Leuconostoc Mesenteroides HJ69 from Kimchi. J. Food Biochem. 2017, 41, e12290. [Google Scholar] [CrossRef]
- Humphreys, G.J.; McBain, A.J. Antagonistic Effects of Streptococcus and Lactobacillus Probiotics in Pharyngeal Biofilms. Lett. Appl. Microbiol. 2019, 68, 303–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordani, B.; Parolin, C.; Vitali, B. Lactobacilli as Anti-Biofilm Strategy in Oral Infectious Diseases: A Mini-Review. Front. Med. Technol. 2021, 3, 769172. [Google Scholar] [CrossRef] [PubMed]
- Scillato, M.; Spitale, A.; Mongelli, G.; Privitera, G.F.; Mangano, K.; Cianci, A.; Stefani, S.; Santagati, M. Antimicrobial Properties of Lactobacillus Cell-Free Supernatants against Multidrug-Resistant Urogenital Pathogens. Microbiologyopen 2021, 10, e1173. [Google Scholar] [CrossRef]
- Chaussee, M.A.; Callegari, E.A.; Chaussee, M.S. Rgg Regulates Growth Phase-Dependent Expression of Proteins Associated with Secondary Metabolism and Stress in Streptococcus Pyogenes. J. Bacteriol 2004, 186, 7091–7099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtney, H.S.; Ofek, I.; Penfound, T.; Nizet, V.; Pence, M.A.; Kreikemeyer, B.; Podbielski, A.; Hasty, D.L.; Dale, J.B. Correction: Relationship between Expression of the Family of M Proteins and Lipoteichoic Acid to Hydrophobicity and Biofilm Formation in Streptococcus Pyogenes. PLoS ONE 2009, 4, e01933385. [Google Scholar] [CrossRef]
- Fabretti, F.; Theilacker, C.; Baldassarri, L.; Kaczynski, Z.; Kropec, A.; Holst, O.; Huebner, J. Alanine Esters of Enterococcal Lipoteichoic Acid Play a Role in Biofilm Formation and Resistance to Antimicrobial Peptides. Infect. Immun. 2006, 74, 4164–4171. [Google Scholar] [CrossRef] [Green Version]
- Kitada, K.; Oho, T. Effect of Saliva Viscosity on the Co-Aggregation between Oral Streptococci and Actinomyces Naeslundii. Gerodontology 2012, 29, e981–e987. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Purpose | Primer Sequence | Source | |
|---|---|---|---|---|
| Forward | Reverse | |||
| covS | Senor kinase gene of covRS TCS. Regulates biofilm and virulence | GAGTGAGCGCGATATCACAA | GCAAGCCAGGAGATGATTCT | [8] |
| covR | Repressor gene of covRS TCS | TGCGCGTGATTCTATTATGG | GGCGGAAAATAGCACGAATA | [8] |
| srv | Biofilm formation and virulence factor regulation | CGGCATTGTGAAACAGAGTG | TCTGACTCGATGCGAACATT | [38] |
| ropB | Global transcriptional factor. Regulates stress, metabolism and virulence | TGATATGGATACGGCAAAACA | TTGACCAAGGCAAAAAGGTT | [39] |
| speB | Extracellular cysteine protease production | CTAGGATACTCTACCAGCG | CAGTAGCAACACATCCTG | [40] |
| luxS | Involved in quorum sensing | CTTTTGGCTGTCGAACAGGT | TCCAGGAACATCTTCCCAAG | [41] |
| mga | Virulence factor regulation | GATCCGTTACTACAAGGG | GTTACTTGTCTGCCTCCT | [11] |
| hasA | Hyaluronic acid capsule synthesis | AGCGTGCTGCTCAATCATTA | AACATCGATCATCCCCAATG | [41] |
| hlyX | Hemolyisn production | GCGCAATACCCAAAATCAGA | CGATTTCACCGACGATTTCT | [42] |
| sagA | Streptolysin S production | AAACAACTCAAGTTGCTCCTG | TGGCGTATAACTTCCGCTAC | [43] |
| slo | Streptolysin O production | GCCAATGTTTCAACAGCTATTG | CGGAGCTGCACTAAAGGCCGC | [42] |
| dltA | D-alanylation of Lipotheicoic acid | GCATTTGGACATCGACTCCT | GTTTTCGAGCCGTAGAAACG | [44] |
| spy125 | Minor pilin subunits on cell surface | AGAGATTAGCGACGCAACAG | ATGGCCATATGTCTCCACCA | [45] |
| col370 | Collagen-like surface protein involved in adhesion | AACCCAGATACTGCACCACA | GCGAGCTGATTACCACCTTG | [45] |
| srtB | Class C sortase production involved in aggregation | GCTGGTTTTGGTTTGTGGGA | CCCCGGGATATTTAACCAACC | [46] |
| ompA | Outer membrane protein involved in biofilm and stress response | GTGCTTCCTGGCTATGAACC | GCAGCGGGTTGGTTATTGTA | [47,48] |
| ciaH | Antioxidant stress response TCS | GGCGGTCTTACAGAATCGTC | CATGTTGCGAACCTCGTCTA | [49] |
| gyrA | Gyrase production (housekeeping gene in the present study) | CAACGCACGTAAGGAAGAAA | CGCTTGTCAAAACGACGTTA | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rather, I.A.; Wani, M.Y.; Kamli, M.R.; Sabir, J.S.M.; Hakeem, K.R.; Firoz, A.; Park, Y.-H.; Hor, Y.-Y. Lactiplantibacillus plantarum KAU007 Extract Modulates Critical Virulence Attributes and Biofilm Formation in Sinusitis Causing Streptococcus pyogenes. Pharmaceutics 2022, 14, 2702. https://doi.org/10.3390/pharmaceutics14122702
Rather IA, Wani MY, Kamli MR, Sabir JSM, Hakeem KR, Firoz A, Park Y-H, Hor Y-Y. Lactiplantibacillus plantarum KAU007 Extract Modulates Critical Virulence Attributes and Biofilm Formation in Sinusitis Causing Streptococcus pyogenes. Pharmaceutics. 2022; 14(12):2702. https://doi.org/10.3390/pharmaceutics14122702
Chicago/Turabian StyleRather, Irfan A., Mohammad Younus Wani, Majid Rasool Kamli, Jamal S. M. Sabir, Khalid Rehman Hakeem, Ahmad Firoz, Yong-Ha Park, and Yan-Yan Hor. 2022. "Lactiplantibacillus plantarum KAU007 Extract Modulates Critical Virulence Attributes and Biofilm Formation in Sinusitis Causing Streptococcus pyogenes" Pharmaceutics 14, no. 12: 2702. https://doi.org/10.3390/pharmaceutics14122702