

Involvement of Bacterial Extracellular Membrane Nanovesicles in Infectious Diseases and Their Application in Medicine
 , , , and
, , , and
Abstract
:1. Introduction
2. EMN Functions
3. Diseases Caused by Variety of Pathogen’s EMN
3.1. Diseases of the Upper and Lower Respiratory Tract
3.2. Pneumonia
3.3. Tuberculosis
3.4. Cystic Fibrosis (CF)
3.5. Whooping Cough
3.6. Sepsis
3.7. Plague
3.8. Gonorrhea
3.9. Peptic Ulcer Disease
3.10. Gastric Cancer
3.11. Gastroenteritis
3.12. Hemolytic Uremic Syndrome (HUS)
3.13. Cholera
3.14. Cystitis and Pyelonephritis
3.15. Listeriosis
3.16. Periodontitis
3.17. Meningitis
3.18. Anthrax
3.19. Tularemia
3.20. Edvarsiellosis
3.21. Endocarditis
3.22. Atherosclerosis
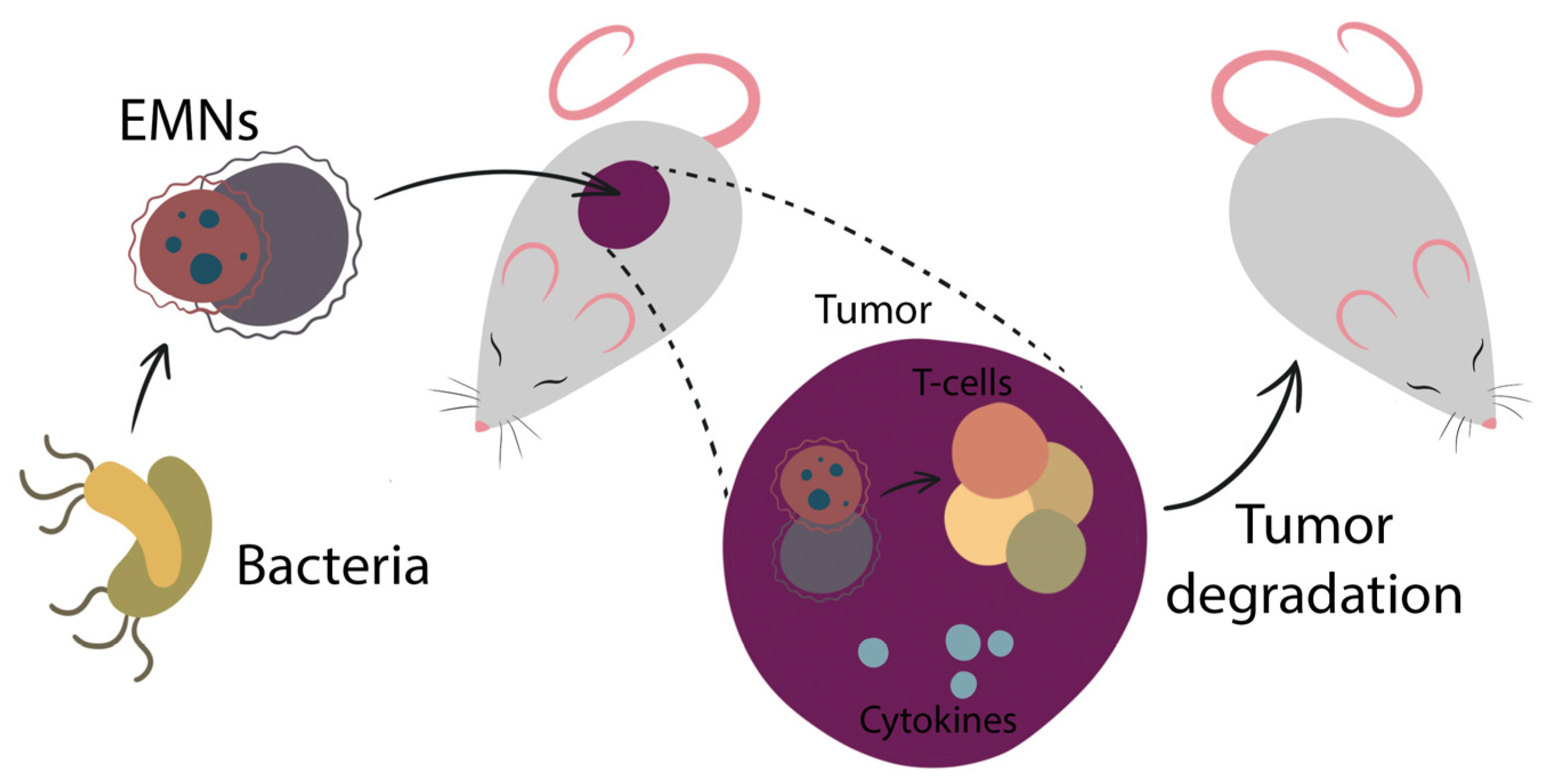
4. EMN Application in Medicine: Current State and Future Directions
5. Vaccine Development Based on EMN
5.1. EMN Vaccines against Cancer
5.2. EMN Engineering
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deatherage, B.L.; Cookson, B.T. Membrane vesicle release in bacteria, eukaryotes, and archaea: A conserved yet underappreciated aspect of microbial life. Infect. Immun. 2012, 80, 1948–1957. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocucci, E.; Racchetti, G.; Meldolesi, J. Shedding microvesicles: Artefacts no more. Trends. Cell Biol. 2009, 19, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Lusta, K.A. Bacterial outer membrane nanovesicles: Structure, biogenesis, functions, and application in biotechnology and medicine (Review). Prikl. Biokhim. Mikrobiol. 2015, 51, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Ñahui Palomino, R.A.; Vanpouille, C.; Costantini, P.E.; Margolis, L. Microbiota-host communications: Bacterial extracellular vesicles as a common language. PLoS Pathog. 2021, 17, e1009508. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sil, S.; Dagur, R.S.; Liao, K.; Peeples, E.S.; Hu, G.; Periyasamy, P.; Buch, S. Strategies for the use of extracellular vesicles for the delivery of therapeutics. J. Neuroimmune Pharmacol. 2020, 15, 422–442. [Google Scholar] [CrossRef]
- Toyofuku, M.; Nomura, N.; Eberl, L. Types and origins of bacterial membrane vesicles. Nat. Rev. Microbiol. 2019, 17, 13–24. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.; Park, J.; Gho, Y.S. Gram-negative and Gram-positive bacterial extracellular vesicles. Semin. Cell Dev. Biol. 2015, 40, 97–104. [Google Scholar] [CrossRef]
- Coelho, C.; Casadevall, A. Answers to naysayers regarding microbial extracellular vesicles. Biochem. Soc. Trans. 2019, 47, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beveridge, T.J. Structures of gram-negative cell walls and their derived membrane vesicles. J. Bacteriol. 1999, 181, 4725–4733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitto, N.J.; Kaparakis-Liaskos, M. The therapeutic benefit of bacterial membrane vesicles. Int. J. Mol. Sci. 2017, 18, 1287. [Google Scholar] [CrossRef] [Green Version]
- Tarashi, S.; Zamani, M.S.; Omrani, M.D.; Fateh, A.; Moshiri, A.; Saedisomeolia, A.; Siadat, S.D.; Kubow, S. Commensal and pathogenic bacterial-derived extracellular vesicles in host-bacterial and interbacterial dialogues: Two sides of the same coin. J. Immunol. Res. 2022, 2022, 8092170. [Google Scholar] [CrossRef]
- Bauman, S.J.; Kuehn, M.J. Purification of outer membrane vesicles from Pseudomonas aeruginosa and their activation of an IL-8 response. Microbes Infect. 2006, 8, 2400–2408. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, Y.; Uchiyama, H.; Nomura, N. Multifunctional membrane vesicles in Pseudomonas aeruginosa. Environ. Microbiol. 2012, 14, 1349–1362. [Google Scholar] [CrossRef]
- Domingues, S.; Nielsen, K.M. Membrane vesicles and horizontal gene transfer in prokaryotes. Curr. Opin. Microbiol. 2017, 38, 16–21. [Google Scholar] [CrossRef]
- Jan, A.T. Outer membrane vesicles (OMVs) of gram-negative bacteria: A perspective update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef]
- Bhar, S.; Edelmann, M.J.; Jones, M.K. Characterization and proteomic analysis of outer membrane vesicles from a commensal microbe, Enterobacter cloacae. J. Proteom. 2021, 231, 103994. [Google Scholar] [CrossRef]
- Lee, J.; Kim, O.Y.; Gho, Y.S. Proteomic profiling of Gram-negative bacterial outer membrane vesicles: Current perspectives. Proteom. Clin. Appl. 2016, 10, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.S.; Sweredoski, M.J.; Graham, R.L.; Hess, S.; Clemons, W.M., Jr. Comprehensive proteomic profiling of outer membrane vesicles from Campylobacter jejuni. J. Proteom. 2014, 98, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.-Y.; Bang, J.Y.; Park, G.W.; Choi, D.-S.; Kang, J.S.; Kim, H.-J.; Park, K.-S.; Lee, J.-O.; Kim, Y.-K.; Kwon, K.-H.; et al. Global proteomic profiling of native outer membrane vesicles derived from Escherichia coli. Proteomics 2007, 7, 3143–3153. [Google Scholar] [CrossRef] [PubMed]
- Berlanda Scorza, F.; Doro, F.; Rodríguez-Ortega, M.J.; Stella, M.; Liberatori, S.; Taddei, A.R.; Serino, L.; Moriel, D.G.; Nesta, B.; Fontana, M.R.; et al. Proteomics characterization of outer membrane vesicles from the extraintestinal pathogenic Escherichia coli DeltatolR IHE3034 mutant. Mol. Cell Proteom. 2008, 7, 473–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galka, F.; Wai, S.N.; Kusch, H.; Engelmann, S.; Hecker, M.; Schmeck, B.; Serino, L.; Moriel, D.G.; Nesta, B.; Fontana, M.R.; et al. Proteomic characterization of the whole secretome of Legionella pneumophila and functional analysis of outer membrane vesicles. Infect. Immun. 2008, 76, 1825–1836. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.O.; Gho, Y.S.; Lee, J.C.; Kim, S.I. Proteome analysis of outer membrane vesicles from a clinical Acinetobacter baumannii isolate. FEMS Microbiol. Lett. 2009, 297, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.Y.; Choi, D.S.; Kim, K.P.; Gho, Y.S. Proteomics in gram-negative bacterial outer membrane vesicles. Mass Spectrom. Rev. 2008, 27, 535–555. [Google Scholar] [CrossRef]
- Sidhu, V.K.; Vorhölter, F.J.; Niehaus, K.; Watt, S.A. Analysis of outer membrane vesicle associated proteins isolated from the plant pathogenic bacterium Xanthomonas campestris pv. campestris. BMC Microbiol. 2008, 8, 87. [Google Scholar] [CrossRef] [Green Version]
- Villageliu, D.N.; Samuelson, D.R. The role of bacterial membrane vesicles in human health and disease. Front. Microbiol. 2022, 13, 828704. [Google Scholar] [CrossRef]
- Turner, L.; Bitto, N.J.; Steer, D.L.; Lo, C.; D’Costa, K.; Ramm, G.; Shambrook, M.; Hill, A.F.; Ferrero, R.; Kaparakis-Liaskos, M. Helicobacter pylori outer membrane vesicle size determines their mechanisms of host cell entry and protein content. Front. Immunol. 2018, 9, 1466. [Google Scholar] [CrossRef]
- Ayalew, S.; Confer, A.W.; Shrestha, B.; Wilson, A.E.; Montelongo, M. Proteomic analysis and immunogenicity of Mannheimia haemolytica vesicles. Clin. Vaccine Immunol. 2013, 20, 191–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naradasu, D.; Miran, W.; Sharma, S.; Takenawa, S.; Soma, T.; Nomura, N.; Toyofuku, M.; Okamoto, A. Biogenesis of outer membrane vesicles concentrates the unsaturated fatty acid of phosphatidylinositol in Capnocytophaga ochracea. Front. Microbiol. 2021, 12, 682685. [Google Scholar] [CrossRef] [PubMed]
- Zavan, L.; Bitto, N.J.; Johnston, E.L.; Greening, D.W.; Kaparakis-Liaskos, M. Helicobacter pylori growth stage determines the size, protein composition, and preferential cargo packaging of outer membrane vesicles. Proteomics 2019, 19, e1800209. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, Y.; Takaki, K.; Futamata, H. Targeted delivery using membrane vesicles in prokaryotes. Biophys. Physicobiol. 2019, 16, 114–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Vázquez, C.; Villarroya-Beltri, C.; Mittelbrunn, M.; Sánchez-Madrid, F. Transfer of extracellular vesicles during immune cell-cell interactions. Immunol. Rev. 2013, 251, 125–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Yin, S.; Wang, M. Extracellular vesicles of bacteria as potential targets for immune interventions. Hum. Vaccines Immunother. 2021, 17, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H. Bacterial outer membrane vesicles as a delivery system for virulence regulation. J. Microbiol. Biotechnol. 2016, 26, 1343–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, N.; Yeruva, L. Role of bacterial infections in extracellular vesicles release and impact on immune response. Biomed. J. 2021, 44, 157–164. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial outer membrane vesicles as a platform for biomedical applications: An update. J. Control Release 2020, 323, 253–268. [Google Scholar] [CrossRef]
- Schetters, S.T.T.; Jong, W.S.P.; Horrevorts, S.K.; Kruijssen, L.J.W.; Engels, S.; Stolk, D.; Daleke-Schermerhorn, M.H.; Garcia-Vallejo, J.; Houben, D.; Unger, W.W.; et al. Outer membrane vesicles engineered to express membrane-bound antigen program dendritic cells for cross-presentation to CD8+ T cells. Acta Biomater. 2019, 91, 248–257. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Q.; Wang, S.; Weng, W.; Jing, Y.; Su, J. Bacterial extracellular vesicles as bioactive nanocarriers for drug delivery: Advances and perspectives. Bioact. Mater. 2021, 14, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Kulp, A.; Kuehn, M.J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schertzer, J.W.; Whiteley, M. Bacterial outer membrane vesicles in trafficking, communication and the host-pathogen interaction. J. Mol. Microbiol. Biotechnol. 2013, 23, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Berleman, J.; Auer, M. The role of bacterial outer membrane vesicles for intra- and interspecies delivery. Environ. Microbiol. 2013, 15, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Dell’Annunziata, F.; Folliero, V.; Giugliano, R.; De Filippis, A.; Santarcangelo, C.; Izzo, V.; Daglia, M.; Galdiero, M.; Arciola, C.; Franci, G. Gene transfer potential of outer membrane vesicles of gram-negative bacteria. Int. J. Mol. Sci. 2021, 22, 5985. [Google Scholar] [CrossRef]
- Jang, S.C.; Kim, S.R.; Yoon, Y.J.; Park, K.S.; Kim, J.H.; Lee, J.; Kim, O.Y.; Choi, E.-J.; Kim, D.-K.; Choi, D.-S.; et al. In vivo kinetic biodistribution of nano-sized outer membrane vesicles derived from bacteria. Small 2015, 11, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, H.M.; Jagannadham, M.V. Biogenesis and multifaceted roles of outer membrane vesicles from Gram-negative bacteria. Microbiology 2014, 160 Pt 10, 2109–2121. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, I.A.; Kuehn, M.J. Offense and defense: Microbial membrane vesicles play both ways. Res. Microbiol. 2012, 163, 607–618. [Google Scholar] [CrossRef] [Green Version]
- McBroom, A.J.; Kuehn, M.J. Outer membrane vesicles. EcoSal Plus 2005, 1. [Google Scholar] [CrossRef] [PubMed]
- McBroom, A.J.; Kuehn, M.J. Release of outer membrane vesicles by Gram-negative bacteria is a novel envelope stress response. Mol. Microbiol. 2007, 63, 545–558. [Google Scholar] [CrossRef] [PubMed]
- McBroom, A.J.; Johnson, A.P.; Vemulapalli, S.; Kuehn, M.J. Outer membrane vesicle production by Escherichia coli is independent of membrane instability. J. Bacteriol. 2006, 188, 5385–5392. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, S.W.; Kuehn, M.J.; Mason, K.M. Elicitation of epithelial cell-derived immune effectors by outer membrane vesicles of nontypeable Haemophilus influenzae. Infect. Immun. 2011, 79, 4361–4369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueter, C.; Bielaszewska, M. Secretion and delivery of intestinal pathogenic Escherichia coli virulence factors via outer membrane vesicles. Front. Cell Infect. Microbiol. 2020, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Macia, L.; Nanan, R.; Hosseini-Beheshti, E.; Grau, G.E. Host- and microbiota-derived extracellular vesicles, immune function, and disease development. Int. J. Mol. Sci. 2019, 21, 107. [Google Scholar] [CrossRef] [Green Version]
- Bielaszewska, M.; Greune, L.; Bauwens, A.; Dersch, P.; Mellmann, A.; Rüter, C. Virulence factor cargo and host cell interactions of shiga toxin-producing Escherichia coli outer membrane vesicles. Methods Mol. Biol. 2021, 2291, 177–205. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Rüter, C.; Bauwens, A.; Greune, L.; Jarosch, K.A.; Steil, D.; Zhang, W.; He, X.; Lloubes, R.; Fruth, A.; et al. Host cell interactions of outer membrane vesicle-associated virulence factors of enterohemorrhagic Escherichia coli O157: Intracellular delivery, trafficking and mechanisms of cell injury. PLoS Pathog. 2017, 13, e1006159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, T.N.; Kuehn, M.J. Virulence and immunomodulatory roles of bacterial outer membrane vesicles. Microbiol. Mol. Biol. Rev. 2010, 74, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Ayala, G.; Torres, L.; Espinosa, M.; Fierros-Zarate, G.; Maldonado, V.; Meléndez-Zajgla, J. External membrane vesicles from Helicobacter pylori induce apoptosis in gastric epithelial cells. FEMS Microbiol. Lett. 2006, 260, 178–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bomberger, J.M.; Maceachran, D.P.; Coutermarsh, B.A.; Ye, S.; O’Toole, G.A.; Stanton, B.A. Long-distance delivery of bacterial virulence factors by Pseudomonas aeruginosa outer membrane vesicles. PLoS Pathog. 2009, 5, e1000382. [Google Scholar] [CrossRef]
- Thay, B.; Damm, A.; Kufer, T.A.; Wai, S.N.; Oscarsson, J. Aggregatibacter actinomycetemcomitans outer membrane vesicles are internalized in human host cells and trigger NOD1- and NOD2-dependent NF-κB activation. Infect. Immun. 2014, 82, 4034–4046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, N.; Tsuda, K.; Omori, H.; Yoshimori, T.; Yoshimura, F.; Amano, A. Porphyromonas gingivalis outer membrane vesicles enter human epithelial cells via an endocytic pathway and are sorted to lysosomal compartments. Infect. Immun. 2009, 77, 4187–4196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wai, S.N.; Lindmark, B.; Söderblom, T.; Takade, A.; Westermark, M.; Oscarsson, J.; Jass, J.; Richter-Dahlfors, A.; Mizunoe, Y.; Uhlin, B.E. Vesicle-mediated export and assembly of pore-forming oligomers of the enterobacterial ClyA cytotoxin. Cell 2003, 115, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchatchouang, S.; Nzouankeu, A.; Kenmoe, S.; Ngando, L.; Penlap, V.; Fonkoua, M.-C.; Pefura-Yone, E.-W.; Njouom, R. Bacterial aetiologies of lower respiratory tract infections among adults in Yaoundé, Cameroon. Biomed. Res. Int. 2019, 2019, 4834396. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.; Ng, D.; Schryvers, A.B. The role of the Moraxella catarrhalis CopB protein in facilitating iron acquisition from human transferrin and lactoferrin. Front. Microbiol. 2021, 12, 714815. [Google Scholar] [CrossRef]
- De Vries, S.P.; Bootsma, H.J.; Hays, J.P.; Hermans, P.W. Molecular aspects of Moraxella catarrhalis pathogenesis. Microbiol. Mol. Biol. Rev. 2009, 73, 389–406. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.T.; Morgelin, M.; Forsgren, A.; Riesbeck, K. Haemophilus influenzae survival during complement-mediated attacks is promoted by Moraxella catarrhalis outer membrane vesicles. J. Infect. Dis. 2007, 195, 1661–1670. [Google Scholar] [CrossRef] [Green Version]
- O’Donoghue, E.J.; Krachler, A.M. Mechanisms of outer membrane vesicle entry into host cells. Cell Microbiol. 2016, 18, 1508–1517. [Google Scholar] [CrossRef]
- Clementi, C.F.; Murphy, T.F. Non-typeable Haemophilus influenzae invasion and persistence in the human respiratory tract. Front. Cell Infect. Microbiol. 2011, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Vidakovics, M.L.; Jendholm, J.; Mörgelin, M.; Månsson, A.; Larsson, C.; Cardell, L.O.; Riesbeck, K. B cell activation by outer membrane vesicles—A novel virulence mechanism. PLoS Pathog. 2010, 6, e1000724. [Google Scholar] [CrossRef]
- Deknuydt, F.; Nordström, T.; Riesbeck, K. Diversion of the host humoral response: A novel virulence mechanism of Haemophilus influenzae mediated via outer membrane vesicles. J. Leukoc. Biol. 2014, 95, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Grief, S.N.; Loza, J.K. Guidelines for the evaluation and treatment of pneumonia. Prim. Care 2018, 45, 485–503. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.S.; Novak, R.; Gao, G.; Murray, P.J.; Shenep, J.L. Pneumolysin, a protein toxin of Streptococcus pneumoniae, induces nitric oxide production from macrophages. Infect. Immun. 1999, 67, 3750–3756. [Google Scholar] [CrossRef] [Green Version]
- Kolios, G.; Valatas, V.; Ward, S.G. Nitric oxide in inflammatory bowel disease: A universal messenger in an unsolved puzzle. Immunology 2004, 113, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, T.J.; Dalziel, C.E. The biology of pneumolysin. Subcell. Biochem. 2014, 80, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Orman, K.L.; Shenep, J.L.; English, B.K. Pneumococci stimulate the production of the inducible nitric oxide synthase and nitric oxide by murine macrophages. J. Infect. Dis. 1998, 178, 1649–1657. [Google Scholar] [CrossRef]
- Salim, T.; Sershen, C.L.; May, E.E. Investigating the role of TNF-α and IFN-γ activation on the dynamics of iNOS gene expression in LPS stimulated macrophages. PLoS ONE 2016, 11, e0153289. [Google Scholar] [CrossRef] [Green Version]
- Hirst, R.A.; Kadioglu, A.; O’callaghan, C.; Andrew, P.W. The role of pneumolysin in pneumococcal pneumonia and meningitis. Clin. Exp. Immunol. 2004, 138, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Olaya-Abril, A.; Prados-Rosales, R.; McConnell, M.J.; Martín-Peña, R.; González-Reyes, J.A.; Jiménez-Munguía, I.; Gómez-Gascón, L.; Fernández, J.; Luque-García, J.L.; García-Lidón, C.; et al. Characterization of protective extracellular membrane-derived vesicles produced by Streptococcus pneumoniae. J. Proteom. 2014, 106, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Parveen, S.; Subramanian, K. Emerging roles of extracellular vesicles in pneumococcal infections: Immunomodulators to potential novel vaccine candidates. Front. Cell Infect. Microbiol. 2022, 12, 836070. [Google Scholar] [CrossRef]
- Jin, J.S.; Kwon, S.O.; Moon, D.C.; Gurung, M.; Lee, J.H.; Kim, S.I.; Lee, J.C. Acinetobacter baumannii secretes cytotoxic outer membrane protein A via outer membrane vesicles. PLoS ONE 2011, 6, e17027. [Google Scholar] [CrossRef] [Green Version]
- Park, K.S.; Lee, J.; Jang, S.C.; Kim, S.R.; Jang, M.H.; Lötvall, J.; Kim, Y.-K.; Gho, Y.S. Pulmonary inflammation induced by bacteria-free outer membrane vesicles from Pseudomonas aeruginosa. Am. J. Respir. Cell Mol. Biol. 2013, 49, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Behrens, F.; Funk-Hilsdorf, T.C.; Kuebler, W.M.; Simmons, S. Bacterial membrane vesicles in pneumonia: From mediators of virulence to innovative vaccine candidates. Int. J. Mol. Sci. 2021, 22, 3858. [Google Scholar] [CrossRef] [PubMed]
- Bussi, C.; Gutierrez, M.G. Mycobacterium tuberculosis infection of host cells in space and time. FEMS Microbiol. Rev. 2019, 43, 341–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Kim, S.-H.; Choi, D.-S.; Lee, J.S.; Kim, D.-K.; Go, G.; Park, S.-M.; Shin, J.H.; Chang, C.L.; Gho, Y.S. Proteomic analysis of extracellular vesicles derived from Mycobacterium tuberculosis. Proteomics 2015, 15, 3331–3337. [Google Scholar] [CrossRef]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: Extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Microbiol. 2015, 13, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Jiménez, V.D.; Leyva-Paredes, K.; García-Martínez, M.; Vázquez-Flores, L.; García-Paredes, V.G.; Campillo-Navarro, M.; Romo-Cruz, I.; Rosales-García, V.H.; Castañeda-Casimiro, J.; González-Pozos, S.; et al. Extracellular vesicles released from mycobacterium tuberculosis-infected neutrophils promote macrophage autophagy and decrease intracellular mycobacterial survival. Front. Immunol. 2018, 9, 272. [Google Scholar] [CrossRef] [Green Version]
- Dryahina, K.; Sovová, K.; Nemec, A.; Španěl, P. Differentiation of pulmonary bacterial pathogens in cystic fibrosis by volatile metabolites emitted by their in vitro cultures: Pseudomonas aeruginosa, Staphylococcus aureus, Stenotrophomonas maltophilia and the Burkholderia cepacia complex. J. Breath Res. 2016, 10, 037102. [Google Scholar] [CrossRef]
- Haurat, M.F.; Elhenawy, W.; Feldman, M.F. Prokaryotic membrane vesicles: New insights on biogenesis and biological roles. Biol. Chem. 2015, 396, 95–109. [Google Scholar] [CrossRef]
- Olsen, I.; Amano, A. Outer membrane vesicles—Offensive weapons or good Samaritans? J. Oral. Microbiol. 2015, 7, 27468. [Google Scholar] [CrossRef] [PubMed]
- Sebo, P.; Osicka, R.; Masin, J. Adenylate cyclase toxin-hemolysin relevance for pertussis vaccines. Expert Rev. Vaccines 2014, 13, 1215–1227. [Google Scholar] [CrossRef] [PubMed]
- Skopova, K.; Tomalova, B.; Kanchev, I.; Rossmann, P.; Svedova, M.; Adkins, I.; Bibova, I.; Tomala, J.; Masin, J.; Guiso, N.; et al. Cyclic, AMP-elevating capacity of adenylate cyclase toxin-hemolysin is sufficient for lung infection but not for full virulence of Bordetella pertussis. Infect. Immun. 2017, 85, e00937-16. [Google Scholar] [CrossRef] [Green Version]
- Eby, J.C.; Gray, M.C.; Mangan, A.R.; Donato, G.M.; Hewlett, E.L. Role of CD11b/CD18 in the process of intoxication by the adenylate cyclase toxin of Bordetella pertussis. Infect. Immun. 2012, 80, 850–859. [Google Scholar] [CrossRef] [Green Version]
- Donato, G.M.; Goldsmith, C.S.; Paddock, C.D.; Eby, J.C.; Gray, M.C.; Hewlett, E.L. Delivery of Bordetella pertussis adenylate cyclase toxin to target cells via outer membrane vesicles. FEBS Lett. 2012, 586, 459–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, J.C. Sepsis definitions: A work in progress. Crit. Care Clin. 2018, 34, 1–14. [Google Scholar] [CrossRef]
- Soult, M.C.; Lonergan, N.E.; Shah, B.; Kim, W.K.; Britt, L.D.; Sullivan, C.J. Outer membrane vesicles from pathogenic bacteria initiate an inflammatory response in human endothelial cells. J. Surg. Res. 2013, 184, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Van der Poll, T.; Meijers, J.C. Systemic inflammatory response syndrome and compensatory anti-inflammatory response syndrome in sepsis. J. Innate Immun. 2010, 2, 379–380. [Google Scholar] [CrossRef]
- Jean-Baptiste, E. Cellular mechanisms in sepsis. J. Intensive Care Med. 2007, 22, 63–72. [Google Scholar] [CrossRef]
- Du, Z.; Wang, X. Pathology and pathogenesis of Yersinia pestis. Adv. Exp. Med. Biol. 2016, 918, 193–222. [Google Scholar] [CrossRef] [PubMed]
- Pechous, R.D.; Sivaraman, V.; Stasulli, N.M.; Goldman, W.E. Pneumonic plague: The darker side of Yersinia pestis. Trends. Microbiol. 2016, 24, 190–197. [Google Scholar] [CrossRef]
- Eddy, J.L.; Gielda, L.M.; Caulfield, A.J.; Rangel, S.M.; Lathem, W.W. Production of outer membrane vesicles by the plague pathogen Yersinia pestis. PLoS ONE 2014, 9, e107002. [Google Scholar] [CrossRef] [PubMed]
- Eren, E.; Murphy, M.; Goguen, J.; van den Berg, B. An active site water network in the plasminogen activator pla from Yersinia pestis. Structure 2010, 18, 809–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielke, R.A.; Wierzbicki, I.H.; Weber, J.V.; Gafken, P.R.; Sikora, A.E. Quantitative proteomics of the Neisseria gonorrhoeae cell envelope and membrane vesicles for the discovery of potential therapeutic targets. Mol. Cell Proteom. 2014, 13, 1299–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyanova, L. Role of Helicobacter pylori virulence factors for iron acquisition from gastric epithelial cells of the host and impact on bacterial colonization. Future Microbiol. 2011, 6, 843–846. [Google Scholar] [CrossRef]
- Lanas, A.; Chan, F.K.L. Peptic ulcer disease. Lancet 2017, 390, 613–624. [Google Scholar] [CrossRef]
- Nejati, S.; Karkhah, A.; Darvish, H.; Validi, M.; Ebrahimpour, S.; Nouri, H.R. Influence of Helicobacter pylori virulence factors CagA and VacA on pathogenesis of gastrointestinal disorders. Microb. Pathog. 2018, 117, 43–48. [Google Scholar] [CrossRef]
- Chmiela, M.; Walczak, N.; Rudnicka, K. Helicobacter pylori outer membrane vesicles involvement in the infection development and Helicobacter pylori-related diseases. J. Biomed. Sci. 2018, 25, 78. [Google Scholar] [CrossRef] [Green Version]
- Parker, H.; Keenan, J.I. Composition and function of Helicobacter pylori outer membrane vesicles. Microbes Infect. 2012, 14, 9–16. [Google Scholar] [CrossRef]
- Olofsson, A.; Nygård Skalman, L.; Obi, I.; Lundmark, R.; Arnqvist, A. Uptake of Helicobacter pylori vesicles is facilitated by clathrin-dependent and clathrin-independent endocytic pathways. mBio 2014, 5, e00979-14. [Google Scholar] [CrossRef] [Green Version]
- Parker, H.; Chitcholtan, K.; Hampton, M.B.; Keenan, J.I. Uptake of Helicobacter pylori outer membrane vesicles by gastric epithelial cells. Infect. Immun. 2010, 78, 5054–5061. [Google Scholar] [CrossRef]
- Ajani, J.A.; Lee, J.; Sano, T.; Janjigian, Y.Y.; Fan, D.; Song, S. Gastric adenocarcinoma. Nat. Rev. Dis. Prim. 2017, 3, 17036. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Graham, D.Y. Helicobacter pylori virulence and cancer pathogenesis. Future Oncol. 2014, 10, 1487–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olofsson, A.; Vallström, A.; Petzold, K.; Tegtmeyer, N.; Schleucher, J.; Carlsson, S.; Haas, R.; Backert, S.; Wai, S.N.; Gröbner, G.; et al. Biochemical and functional characterization of Helicobacter pylori vesicles. Mol. Microbiol. 2010, 77, 1539–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chitcholtan, K.; Hampton, M.B.; Keenan, J.I. Outer membrane vesicles enhance the carcinogenic potential of Helicobacter pylori. Carcinogenesis 2008, 29, 2400–2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, N.P.; Schiellerup, P.; Boisen, N.; Klein, B.M.; Locht, H.; Abuoun, M.; Newell, D.; Krogfelt, K.A. The role of Campylobacter jejuni cytolethal distending toxin in gastroenteritis: Toxin detection, antibody production, and clinical outcome. APMIS 2011, 119, 626–634. [Google Scholar] [CrossRef]
- Elmi, A.; Watson, E.; Sandu, P.; Gundogdu, O.; Mills, D.C.; Inglis, N.F.; Manson, E.; Imrie, L.; Bajaj-Elliott, M.; Wren, B.W.; et al. Campylobacter jejuni outer membrane vesicles play an important role in bacterial interactions with human intestinal epithelial cells. Infect. Immun. 2012, 80, 4089–4098. [Google Scholar] [CrossRef] [Green Version]
- Schwidder, M.; Heinisch, L.; Schmidt, H. Genetics, Toxicity, and distribution of enterohemorrhagic Escherichia coli hemolysin. Toxins 2019, 11, 502. [Google Scholar] [CrossRef] [Green Version]
- Bielaszewska, M.; Rüter, C.; Kunsmann, L.; Greune, L.; Bauwens, A.; Zhang, W.; Kuczius, T.; Kim, K.S.; Mellmann, A.; Schmidt, M.A.; et al. Enterohemorrhagic Escherichia coli hemolysin employs outer membrane vesicles to target mitochondria and cause endothelial and epithelial apoptosis. PLoS Pathog. 2013, 9, e1003797. [Google Scholar] [CrossRef] [Green Version]
- Pantano, S.; Montecucco, C. A molecular model of the Vibrio cholerae cytolysin transmembrane pore. Toxicon 2006, 47, 35–40. [Google Scholar] [CrossRef]
- Rai, A.K.; Chattopadhyay, K. Revisiting the oligomerization mechanism of Vibrio cholerae cytolysin, a beta-barrel pore-forming toxin. Biochem. Biophys. Res. Commun. 2016, 474, 421–427. [Google Scholar] [CrossRef]
- Elluri, S.; Enow, C.; Vdovikova, S.; Rompikuntal, P.K.; Dongre, M.; Carlsson, S.; Pal, A.; Uhlin, B.E.; Wai, S.N. Outer membrane vesicles mediate transport of biologically active Vibrio cholerae cytolysin (VCC) from V. cholerae strains. PLoS ONE 2014, 9, e106731. [Google Scholar] [CrossRef] [PubMed]
- Bielig, H.; Rompikuntal, P.K.; Dongre, M.; Zurek, B.; Lindmark, B.; Ramstedt, M.; Wai, S.N.; Kufer, T.A. NOD-like receptor activation by outer membrane vesicles from Vibrio cholerae non-O1 non-O139 strains is modulated by the quorum-sensing regulator HapR. Infect. Immun. 2011, 79, 1418–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolman, K.B. Cystitis and pyelonephritis: Diagnosis, treatment, and prevention. Prim. Care 2019, 46, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Garcia, T.A.; Ventura, C.L.; Smith, M.A.; Merrell, D.S.; O’Brien, A.D. Cytotoxic necrotizing factor 1 and hemolysin from uropathogenic Escherichia coli elicit different host responses in the murine bladder. Infect. Immun. 2013, 81, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouokam, J.C.; Wai, S.N.; Fällman, M.; Dobrindt, U.; Hacker, J.; Uhlin, B.E. Active cytotoxic necrotizing factor 1 associated with outer membrane vesicles from uropathogenic Escherichia coli. Infect. Immun. 2006, 74, 2022–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLauchlin, J.; Crofts, N.; Campbell, D.M. A possible outbreak of listeriosis caused by an unusual strain of Listeria monocytogenes. J. Infect. 1989, 18, 179–187. [Google Scholar] [CrossRef]
- Šrajer Gajdošik, M.; Andjelković, U.; Gašo-Sokač, D.; Pavlović, H.; Shevchuk, O.; Martinović, T.; Clifton, J.; Begić, M.; Josić, D. Proteomic analysis of pyridoxal oxime derivatives treated Listeria monocytogenes reveals down-regulation of the main virulence factor, Listeriolysin O. Food Res. Int. 2020, 131, 108951. [Google Scholar] [CrossRef]
- Lee, J.H.; Choi, C.W.; Lee, T.; Kim, S.I.; Lee, J.C.; Shin, J.H. Transcription factor σB plays an important role in the production of extracellular membrane-derived vesicles in Listeria monocytogenes. PLoS ONE 2013, 8, e73196. [Google Scholar] [CrossRef] [Green Version]
- Vdovikova, S.; Luhr, M.; Szalai, P.; Nygård Skalman, L.; Francis, M.K.; Lundmark, R.; Engedal, N.; Johansson, J.; Wai, S.N. A novel role of Listeria monocytogenes membrane vesicles in inhibition of autophagy and cell death. Front. Cell Infect. Microbiol. 2017, 7, 154. [Google Scholar] [CrossRef] [Green Version]
- Mysak, J.; Podzimek, S.; Sommerova, P.; Lyuya-Mi, Y.; Bartova, J.; Janatova, T.; Prochazkova, J.; Duskova, J. Porphyromonas gingivalis: Major periodontopathic pathogen overview. J. Immunol. Res. 2014, 2014, 476068. [Google Scholar] [CrossRef]
- Ho, M.H.; Chen, C.H.; Goodwin, J.S.; Wang, B.Y.; Xie, H. Functional advantages of Porphyromonas gingivalis vesicles. PLoS ONE 2015, 10, e0123448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groeger, S.; Jarzina, F.; Mamat, U.; Meyle, J. Induction of B7-H1 receptor by bacterial cells fractions of Porphyromonas gingivalis on human oral epithelial cells: B7-H1 induction by Porphyromonas gingivalis fractions. Immunobiology 2017, 222, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Nakao, R.; Takashiba, S.; Kosono, S.; Yoshida, M.; Watanabe, H.; Ohnishi, M.; Senpuku, H. Effect of Porphyromonas gingivalis outer membrane vesicles on gingipain-mediated detachment of cultured oral epithelial cells and immune responses. Microbes Infect. 2014, 16, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Furuta, N.; Takeuchi, H.; Amano, A. Entry of Porphyromonas gingivalis outer membrane vesicles into epithelial cells causes cellular functional impairment. Infect. Immun. 2009, 77, 4761–4770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, L.; Han, N.; Du, J.; Guo, L.; Luo, Z.; Liu, Y. Pathogenesis of important virulence factors of Porphyromonas gingivalis via toll-like receptors. Front. Cell Infect. Microbiol. 2019, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, D.; Liu, S.; Zhang, S.; Pan, Y. The Role of Porphyromonas gingivalis outer membrane vesicles in periodontal disease and related systemic diseases. Front. Cell Infect. Microbiol. 2021, 10, 585917. [Google Scholar] [CrossRef]
- Kato, S.; Kowashi, Y.; Demuth, D.R. Outer membrane-like vesicles secreted by Actinobacillus actinomycetemcomitans are enriched in leukotoxin. Microb. Pathog. 2002, 32, 1–13. [Google Scholar] [CrossRef]
- Loh, L.N.; Ward, T.H. Escherichia coli K1 invasion of human brain microvascular endothelial cells. Methods Enzymol. 2012, 506, 93–113. [Google Scholar] [CrossRef]
- Yu, H.; Kim, K.S. YgfZ contributes to secretion of cytotoxic necrotizing factor 1 into outer-membrane vesicles in Escherichia coli. Microbiology 2012, 158 Pt 3, 612–621. [Google Scholar] [CrossRef] [Green Version]
- Balsalobre, C.; Silván, J.M.; Berglund, S.; Mizunoe, Y.; Uhlin, B.E.; Wai, S.N. Release of the type I secreted alpha-haemolysin via outer membrane vesicles from Escherichia coli. Mol. Microbiol. 2006, 59, 99–112. [Google Scholar] [CrossRef]
- Bhatnagar, R.; Batra, S. Anthrax toxin. Crit. Rev. Microbiol. 2001, 27, 167–200. [Google Scholar] [CrossRef] [PubMed]
- Hammamieh, R.; Ribot, W.J.; Abshire, T.G.; Jett, M.; Ezzell, J. Activity of the Bacillus anthracis 20 kDa protective antigen component. BMC Infect. Dis. 2008, 8, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.J.; Guo, Q. The adenylyl cyclase activity of anthrax edema factor. Mol. Asp. Med. 2009, 30, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Rivera, J.; Cordero, R.J.; Nakouzi, A.S.; Frases, S.; Nicola, A.; Casadevall, A. Bacillus anthracis produces membrane-derived vesicles containing biologically active toxins. Proc. Natl. Acad. Sci. USA 2010, 107, 19002–19007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimentova, J.; Pavkova, I.; Horcickova, L.; Bavlovic, J.; Kofronova, O.; Benada, O.; Stulik, J. Francisella tularensis subsp. holarctica releases differentially loaded outer membrane vesicles under various stress conditions. Front. Microbiol. 2019, 10, 2304. [Google Scholar] [CrossRef] [Green Version]
- Mainous, M.E.; Smith, S.A.; Kuhn, D.D. Effect of common aquaculture chemicals against Edwardsiella ictaluri and E. tarda. J. Aquat. Anim. Health 2010, 22, 224–228. [Google Scholar] [CrossRef]
- Park, S.B.; Jang, H.B.; Nho, S.W.; Cha, I.S.; Hikima, J.; Ohtani, M.; Aoki, T.; Jung, T.S. Outer membrane vesicles as a candidate vaccine against edwardsiellosis. PLoS ONE 2011, 6, e17629. [Google Scholar] [CrossRef]
- Sharara, S.L.; Tayyar, R.; Kanafani, Z.A.; Kanj, S.S. HACEK endocarditis: A review. Expert Rev. Anti Infect. Ther. 2016, 14, 539–545. [Google Scholar] [CrossRef]
- Haraszthy, V.I.; Zambon, J.J.; Trevisan, M.; Zeid, M.; Genco, R.J. Identification of periodontal pathogens in atheromatous plaques. J. Periodontol. 2000, 71, 1554–1560. [Google Scholar] [CrossRef]
- Piñon-Esteban, P.; Núñez, L.; Moure, R.P.; Marrón-Liñares, G.M.; Flores-Rios, X.; Aldama-Lopez, G.; Salgado-Fernandez, J.; Calviño-Santos, R.; Rebollal-Leal, F.; Pan-Lizcano, R.; et al. Presence of bacterial DNA in thrombotic material of patients with myocardial infarction. Sci. Rep. 2020, 10, 16299. [Google Scholar] [CrossRef]
- Xie, M.; Tang, Q.; Nie, J.; Zhang, C.; Zhou, X.; Yu, S.; Sun, J.; Cheng, X.; Dong, N.; Hu, Y.; et al. BMAL1-downregulation aggravates Porphyromonas Gingivalis-induced atherosclerosis by encouraging oxidative stress. Circ. Res. 2020, 126, e15–e29. [Google Scholar] [CrossRef]
- Gibson, F.C., 3rd; Genco, C.A. Porphyromonas gingivalis mediated periodontal disease and atherosclerosis: Disparate diseases with commonalities in pathogenesis through TLRs. Curr. Pharm. Des. 2007, 13, 3665–3675. [Google Scholar] [CrossRef] [PubMed]
- Pothineni, N.V.K.; Subramany, S.; Kuriakose, K.; Shirazi, L.F.; Romeo, F.; Shah, P.K.; Mehta, J.L. Infections, atherosclerosis, and coronary heart disease. Eur. Heart J. 2017, 38, 3195–3201. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Hu, J.; Tian, M.; Li, Y.; Ren, C.; Deng, Y.; Jiang, Y. Extracellular vesicles from Helicobacter pylori-infected cells and Helicobacter pylori outer membrane vesicles in atherosclerosis. Helicobacter 2022, 27, e12877. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xia, Y.-P.; Luo, X.-Y.; Chen, S.-L.; Li, B.-W.; Ye, Z.-M.; Chen, S.-C.; Mao, L.; Jin, H.-J.; Li, Y.-N.; et al. Exosomal CagA derived from Helicobacter pylori-infected gastric epithelial cells induces macrophage foam cell formation and promotes atherosclerosis. J. Mol. Cell Cardiol. 2019, 135, 40–51. [Google Scholar] [CrossRef]
- Pap, E.; Pállinger, E.; Pásztói, M.; Falus, A. Highlights of a new type of intercellular communication: Microvesicle-based information transfer. Inflamm. Res. 2009, 58, 1–8. [Google Scholar] [CrossRef]
- Unal, C.M.; Schaar, V.; Riesbeck, K. Bacterial outer membrane vesicles in disease and preventive medicine. Semin. Immunopathol. 2011, 33, 395–408. [Google Scholar] [CrossRef]
- Shin, H.S.; Gedi, V.; Kim, J.K.; Lee, D.K. Detection of Gram-negative bacterial outer membrane vesicles using DNA aptamers. Sci. Rep. 2019, 9, 13167. [Google Scholar] [CrossRef] [Green Version]
- Richter, R.; Lehr, C.M. Extracellular vesicles as novel assay tools to study cellular interactions of anti-infective compounds—A perspective. Adv. Drug Deliv. Rev. 2021, 173, 492–503. [Google Scholar] [CrossRef]
- Sutaria, D.S.; Badawi, M.; Phelps, M.A.; Schmittgen, T.D. Achieving the promise of therapeutic extracellular vesicles: The devil is in details of therapeutic loading. Pharm. Res. 2017, 34, 1053–1066. [Google Scholar] [CrossRef]
- Schorey, J.S.; Cheng, Y.; Singh, P.P.; Smith, V.L. Exosomes and other extracellular vesicles in host-pathogen interactions. EMBO Rep. 2015, 16, 24–43. [Google Scholar] [CrossRef] [Green Version]
- Barile, L.; Vassalli, G. Exosomes: Therapy delivery tools and biomarkers of diseases. Pharmacol. Ther. 2017, 174, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.M.; Brown, A.C. Bacterial outer membrane vesicles as antibiotic delivery vehicles. Front. Immunol. 2021, 12, 733064. [Google Scholar] [CrossRef] [PubMed]
- Anderson, H.C.; Mulhall, D.; Garimella, R. Role of extracellular membrane vesicles in the pathogenesis of various diseases, including cancer, renal diseases, atherosclerosis, and arthritis. Lab. Investig. 2010, 90, 1549–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, N.J.; Turner, K.B.; Medintz, I.L.; Walper, S.A. Emerging therapeutic delivery capabilities and challenges utilizing enzyme/protein packaged bacterial vesicles. Ther. Deliv. 2015, 6, 873–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Meng, L.; Chen, Y.; Dong, Z.; Peng, Q. Bacterial outer membrane vesicles as potential biological nanomaterials for antibacterial therapy. Acta Biomater. 2022, 140, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Li, C.; Peng, X.; Wu, S.; Li, Y.; Tan, J.P.K.; Yang, Y.Y.; Yuan, P.; Ding, X. Fight bacteria with bacteria: Bacterial membrane vesicles as vaccines and delivery nanocarriers against bacterial infections. Nanomedicine 2021, 35, 102398. [Google Scholar] [CrossRef]
- Jiang, L.; Schinkel, M.; van Essen, M.; Schiffelers, R.M. Bacterial membrane vesicles as promising vaccine candidates. Eur. J. Pharm. Biopharm. 2019, 145, 1–6. [Google Scholar] [CrossRef]
- Van der Pol, L.; Stork, M.; van der Ley, P. Outer membrane vesicles as platform vaccine technology. Biotechnol. J. 2015, 10, 1689–1706. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, R.; Fernández, S.; Zayas, C.; Acosta, A.; Sarmiento, M.E.; Ferro, V.A.; Rosenqvist, E.; Campa, C.; Cardoso, D.; Garcia, L.; et al. Bacterial outer membrane vesicles and vaccine applications. Front. Immunol. 2014, 5, 121. [Google Scholar] [CrossRef]
- Holst, J.; Oster, P.; Arnold, R.; Tatley, M.V.; Næss, L.M.; Aaberge, I.S.; Galloway, Y.; McNicholas, A.; O’Hallahan, J.; Rosenqvist, E.; et al. Vaccines against meningococcal serogroup B disease containing outer membrane vesicles (OMV): Lessons from past programs and implications for the future. Hum. Vaccines Immunother. 2013, 9, 1241–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakao, R.; Hasegawa, H.; Dongying, B.; Ohnishi, M.; Senpuku, H. Assessment of outer membrane vesicles of periodontopathic bacterium Porphyromonas gingivalis as possible mucosal immunogen. Vaccine 2016, 34, 4626–4634. [Google Scholar] [CrossRef]
- Daleke-Schermerhorn, M.H.; Felix, T.; Soprova, Z.; Ten Hagen-Jongman, C.M.; Vikström, D.; Majlessi, L.; Beskers, J.; Follmann, F.; de Punder, K.; van der Wel, N.N.; et al. Decoration of outer membrane vesicles with multiple antigens by using an autotransporter approach. Appl. Environ. Microbiol. 2014, 80, 5854–5865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Yang, F.; Zou, J.; Wu, W.; Jing, H.; Gou, Q.; Li, H.; Gu, J.; Zou, Q.; Zhang, J. Immunization with Pseudomonas aeruginosa outer membrane vesicles stimulates protective immunity in mice. Vaccine 2018, 36, 1047–1054. [Google Scholar] [CrossRef]
- Bottero, D.; Gaillard, M.E.; Errea, A.; Moreno, G.; Zurita, E.; Pianciola, L.; Rumbo, M.; Hozbor, D. Outer membrane vesicles derived from Bordetella parapertussis as an acellular vaccine against Bordetella parapertussis and Bordetella pertussis infection. Vaccine 2013, 31, 5262–5268. [Google Scholar] [CrossRef] [PubMed]
- Asensio, C.J.; Gaillard, M.E.; Moreno, G.; Bottero, D.; Zurita, E.; Rumbo, M.; van der Ley, P.; van der Ark, A.; Hozbor, D. Outer membrane vesicles obtained from Bordetella pertussis Tohama expressing the lipid A deacylase PagL as a novel acellular vaccine candidate. Vaccine 2011, 29, 1649–1656. [Google Scholar] [CrossRef]
- Nieves, W.; Petersen, H.; Judy, B.M.; Blumentritt, C.A.; Russell-Lodrigue, K.; Roy, C.J.; Torres, A.; Morici, L.A. A Burkholderia pseudomallei outer membrane vesicle vaccine provides protection against lethal sepsis. Clin. Vaccine Immunol. 2014, 21, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Byvalov, A.A.; Konyshev, I.V.; Uversky, V.N.; Dentovskaya, S.V.; Anisimov, A.P. Yersinia outer membrane vesicles as potential vaccine candidates in protecting against plague. Biomolecules 2020, 10, 1694. [Google Scholar] [CrossRef] [PubMed]
- Maurakis, S.A.; Cornelissen, C.N. Recent progress towards a gonococcal vaccine. Front. Cell Infect. Microbiol. 2022, 12, 881392. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, X.; Zhang, Y.; Song, Z.; Li, R.; Ruan, H.; Huang, X. Orally-administered outer-membrane vesicles from Helicobacter pylori reduce H. pylori infection via Th2-biased immune responses in mice. Pathog. Dis. 2019, 77, ftz050. [Google Scholar] [CrossRef] [PubMed]
- Fingermann, M.; Avila, L.; De Marco, M.B.; Vázquez, L.; Di Biase, D.N.; Müller, A.V.; Lescano, M.; Dokmetjian, J.C.; Castillo, S.F.; Quiñoy, J.L.P. OMV-based vaccine formulations against Shiga toxin producing Escherichia coli strains are both protective in mice and immunogenic in calves. Hum. Vaccines Immunother. 2018, 14, 2208–2213. [Google Scholar] [CrossRef] [Green Version]
- Godlewska, R.; Kuczkowski, M.; Wyszyńska, A.; Klim, J.; Derlatka, K.; Woźniak-Biel, A.; Jagusztyn-Krynicka, E.K. Evaluation of a protective effect of in ovo delivered Campylobacter jejuni OMVs. Appl. Microbiol. Biotechnol. 2016, 100, 8855–8864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schild, S.; Nelson, E.J.; Bishop, A.L.; Camilli, A. Characterization of Vibrio cholerae outer membrane vesicles as a candidate vaccine for cholera. Infect. Immun. 2009, 77, 472–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierson, T.; Matrakas, D.; Taylor, Y.U.; Manyam, G.; Morozov, V.N.; Zhou, W.; van Hoek, M.L. Proteomic characterization and functional analysis of outer membrane vesicles of Francisella novicida suggests possible role in virulence and use as a vaccine. J. Proteome Res. 2011, 10, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Hagan, E.C.; Mobley, H.L. Uropathogenic Escherichia coli outer membrane antigens expressed during urinary tract infection. Infect. Immun. 2007, 75, 3941–3949. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.; Moreno, G.; Bottero, D.; Gaillard, M.E.; Fingermann, M.; Graieb, A.; Rumbo, M.; Hozbor, D. Outer membrane vesicles as acellular vaccine against pertussis. Vaccine 2008, 26, 4639–4646. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Rao, Y.; Zhu, K.; Rao, X.; Zhou, R. Engineered remolding and application of bacterial membrane vesicles. Front. Microbiol. 2021, 12, 729369. [Google Scholar] [CrossRef]
- Jain-Gupta, N.; Contreras-Rodriguez, A.; Vemulapalli, R.; Witonsky, S.G.; Boyle, S.M.; Sriranganathan, N. Pluronic P85 enhances the efficacy of outer membrane vesicles as a subunit vaccine against Brucella melitensis challenge in mice. FEMS Immunol. Med. Microbiol. 2012, 66, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Fang, Z.; Li, R.; Huang, X.; Liu, Q. Design of outer membrane vesicles as cancer vaccines: A new toolkit for cancer therapy. Cancers 2019, 11, 1314. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.Y.; Park, H.T.; Nth, D.; Choi, S.J.; Lee, J.; Kim, J.H.; Lee, S.-W.; Gho, Y.S. Bacterial outer membrane vesicles suppress tumor by interferon-γ-mediated antitumor response. Nat. Commun. 2017, 8, 626. [Google Scholar] [CrossRef]
- Gujrati, V.; Kim, S.; Kim, S.H.; Min, J.J.; Choy, H.E.; Kim, S.C.; Jon, S. Bioengineered bacterial outer membrane vesicles as cell-specific drug-delivery vehicles for cancer therapy. ACS Nano 2014, 8, 1525–1537. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, R.; Cheng, K.; Zhang, K.; Wang, Y.; Zhang, Y.; Li, Y.; Liu, G.; Xu, J.; Xu, J.; et al. Bacterial outer membrane vesicles presenting programmed death 1 for improved cancer immunotherapy via immune activation and checkpoint inhibition. ACS Nano 2020, 14, 16698–16711. [Google Scholar] [CrossRef] [PubMed]
- Aly, R.G.; El-Enbaawy, M.I.; Abd El-Rahman, S.S.; Ata, N.S. Antineoplastic activity of Salmonella Typhimurium outer membrane nanovesicles. Exp. Cell Res. 2021, 399, 112423. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, C.; You, S.; Zhang, K.; Li, M.; Cao, Y.; Wang, C.; Dong, H.; Zhang, X. Bacterial vesicle-cancer cell hybrid membrane-coated nanoparticles for tumor specific immune activation and photothermal therapy. ACS Appl. Mater. Interfaces 2020, 12, 41138–41147. [Google Scholar] [CrossRef]
- Van den Berg van Saparoea, H.B.; Houben, D.; de Jonge, M.I.; Jong, W.S.P.; Luirink, J. Display of recombinant proteins on bacterial outer membrane vesicles by using protein ligation. Appl. Environ. Microbiol. 2018, 84, e02567-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-H.; Kim, K.-S.; Lee, S.-R.; Kim, E.; Kim, M.-S.; Lee, E.-Y.; Gho, Y.S.; Kim, J.-W.; Bishop, R.E.; Chang, K.-T. Structural modifications of outer membrane vesicles to refine them as vaccine delivery vehicles. Biochim. Biophys. Acta 2009, 1788, 2150–2159. [Google Scholar] [CrossRef] [Green Version]
- Vader, P.; Fuhrmann, G. Approaches to surface engineering of extracellular vesicles. Adv. 2021, 173, 416–426. [Google Scholar] [CrossRef]
- Li, R.; Liu, Q. Engineered bacterial outer membrane vesicles as multifunctional delivery platforms. Front. Mater. 2020, 7, 202. [Google Scholar] [CrossRef]
- Gerritzen, M.J.H.; Salverda, M.L.M.; Martens, D.E.; Wijffels, R.H.; Stork, M. Spontaneously released Neisseria meningitidis outer membrane vesicles as vaccine platform: Production and purification. Vaccine 2019, 37, 6978–6986. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Driedonks, T.A.P.; Jong, W.S.P.; Dhakal, S.; Bart van den Berg van Saparoea, H.; Sitaras, I.; Zhou, R.; Caputo, C.; Littlefield, K.; Lowman, M.; et al. A bacterial extracellular vesicle-based intranasal vaccine against SARS-CoV-2 protects against disease and elicits neutralizing antibodies to wild-type and Delta variants. J. Extracell. Vesicles 2022, 11, e12192. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Disease | Infectious Agent | Approach | References |
|---|---|---|---|
| Pneumonia | Acinetobacter baumannii, Bordetella pertussis, Streptococcus pneumoniae, Klebsiella pneumonia | intramuscular, intraperitoneal or the intranasal administration of A. baumannii EMNs, including multidrug-resistant strains; intranasal administration of B. pertussis EMNs; intranasal administration of native S. pneumoniae MVs; intraperitoneal administration of native K. pneumoniae-secreted EMNs | [83] |
| Meningitis | Neisseria meningitides | the complex formed from the adjuvant lipopolysaccharide (LPS) and bacterial surface antigen allows EMNs to produce an adaptive immune response | [170,171] |
| Periodontitis | Porphyromonas gingivalis | intranasal immunization with a combination of P. gingivalis EMNs and a TLR3 agonist, Poly(I:C) | [172] |
| Tuberculosis | Mycobacterium tuberculosis | Immunization with an Ag85B-ESAT6-Rv2660c fusion protein stimulated stronger immune responses and better protection against M. tuberculosis | [173] |
| Cystic fibrosis | Pseudomonas aeruginosa | Immunization with Pseudomonas aeruginosa EMNs was efficient to launch a complex humoral and cellular immune response in mice | [174] |
| Whooping cough | Bordetella parapertussis Bordetella pertussis | intranasal administration of B. pertussis EMNs | [175,176] |
| Sepsis (Melioidosis) | Burkholderia pseudomallei | multivalent EMN vaccine derived from B. pseudomallei strain 1026b | [177] |
| Plague | Yersinia pestis | Immunization with EV of engineered strain of Y. pestis producing a detoxified 1-dephosphorylated hexa-acylated version instead of highly inflammatory bisphosphoryl hexa-acylated lipid A (monophosphoryl lipid A, MPLA) | [178] |
| Gonorrhea | Neisseria gonorrhoeae | Intravaginal administration of EMN preparation with microencapsulated IL-12 as an adjuvant appeared to accelerate gonococcal clearance and to induce N.gonorrhoeae-specific antibodies to BALB/C mice | [179] |
| Peptic ulcer disease | Helicobacter pylori | Intragastrical immunization with the H. pylori EMNs plus CT as an adjuvant, or a WCV supplemented with CT | [180] |
| Hemolitic uremic syndrome | E. coli (STEC) | chemical-inactivated EMN (EMNi) obtained from a virulent STEC strain (O157:H7 serotype) | [181] |
| Gastroenteritis | Campylobacter jejuni | In ovo vaccination with wtEMNs, 639-EMNs, and 1405-EMNs of chicks | [182] |
| Cholera | Vibrio cholera | intranasal, intragastric, or intraperitoneal administration of EMNs derived from V. cholerae | [183] |
| Tularemia | Francisellanovicida | EMNs were shown to induce both cell mediated and humoral immunity | [184] |
| Cystitis and pyelonephritis | Escherichia coli (UPEC) | OMPs purified from E. coli CFT073 | [185] |
| Pertussis | Bordetella pertussis | EMNs derived from B. pertussis | [186] |
| Shigellosis | Shigella flexneri | EMNs derived from S. flexneri | [187] |
| Brucellosis | Brucella melitensis | Pluronic P85 enhances the efficacy of B. melitensis EMNs | [188] |
| Edwardsiellosis | Edwardsiella tarda | EMNs naturally released from E. tarda injected into olive flounders | [147] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lusta, K.A.; Poznyak, A.V.; Litvinova, L.; Poggio, P.; Orekhov, A.N.; Melnichenko, A.A. Involvement of Bacterial Extracellular Membrane Nanovesicles in Infectious Diseases and Their Application in Medicine. Pharmaceutics 2022, 14, 2597. https://doi.org/10.3390/pharmaceutics14122597
Lusta KA, Poznyak AV, Litvinova L, Poggio P, Orekhov AN, Melnichenko AA. Involvement of Bacterial Extracellular Membrane Nanovesicles in Infectious Diseases and Their Application in Medicine. Pharmaceutics. 2022; 14(12):2597. https://doi.org/10.3390/pharmaceutics14122597
Chicago/Turabian StyleLusta, Konstantin A., Anastasia V. Poznyak, Larisa Litvinova, Paolo Poggio, Alexander N. Orekhov, and Alexandra A. Melnichenko. 2022. "Involvement of Bacterial Extracellular Membrane Nanovesicles in Infectious Diseases and Their Application in Medicine" Pharmaceutics 14, no. 12: 2597. https://doi.org/10.3390/pharmaceutics14122597