
A Comparative Analysis of the Wound Healing-Related Heterogeneity of Adipose-Derived Stem Cells Donors


Abstract
:1. Introduction
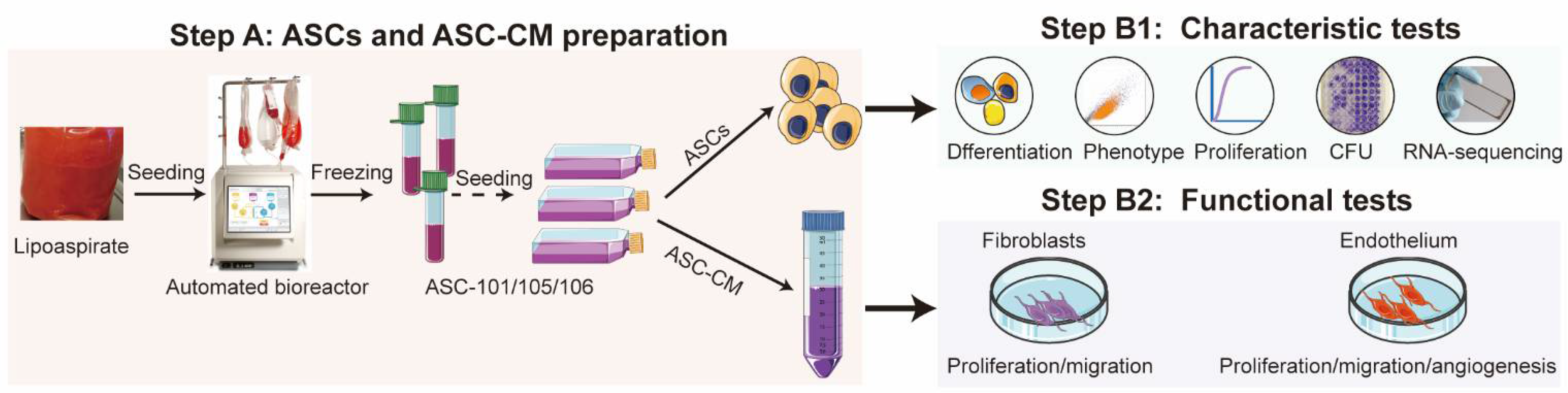
2. Materials and Methods
2.1. Cell Isolation and Culture
2.2. Stem Cell Characterization
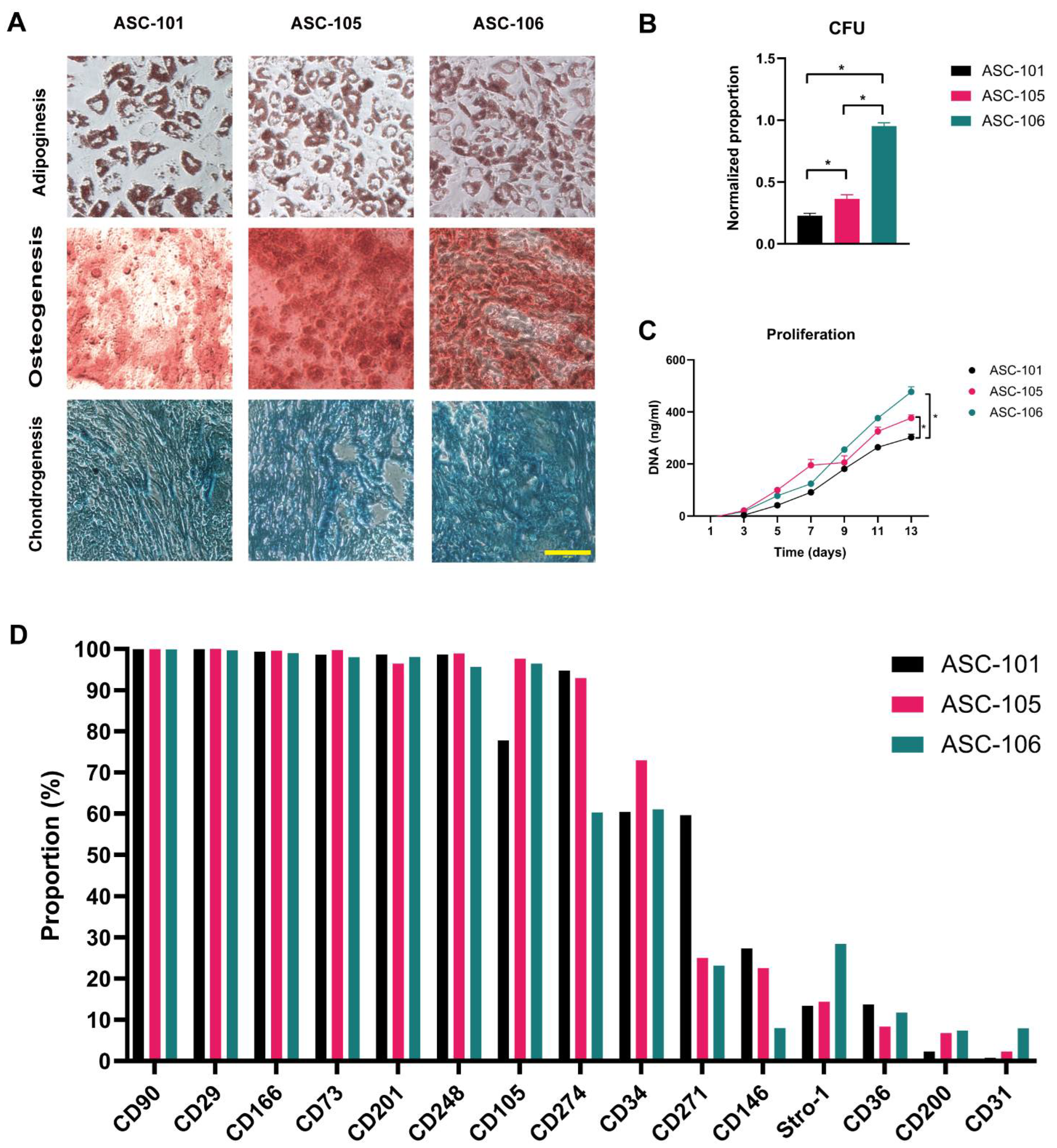
2.3. Trilineage Differentiation
2.3.1. Adipogenic Differentiation
2.3.2. Osteogenic Differentiation
2.3.3. Chondrogenic Differentiation
2.4. Colony-Forming Unit
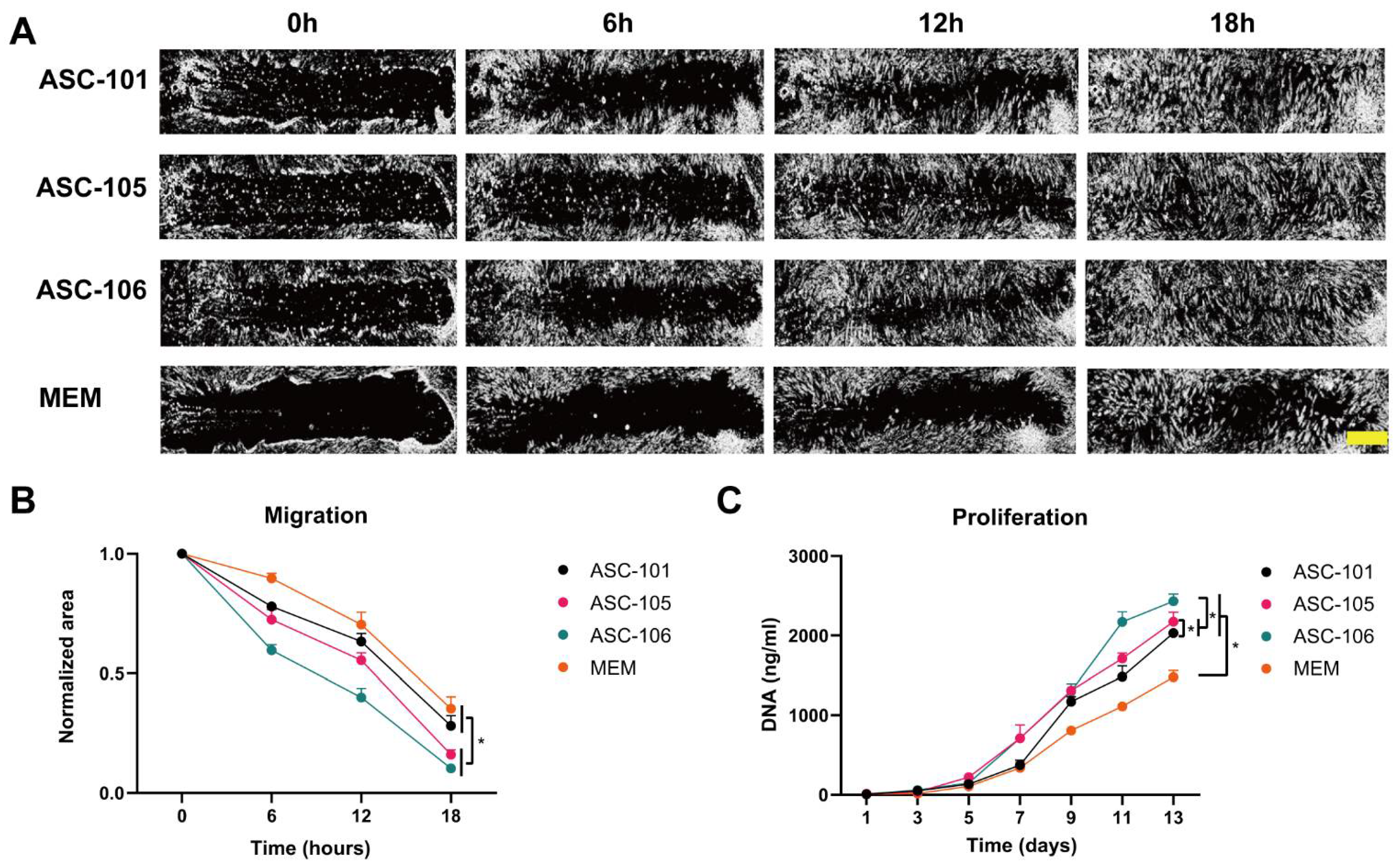
2.5. Proliferation
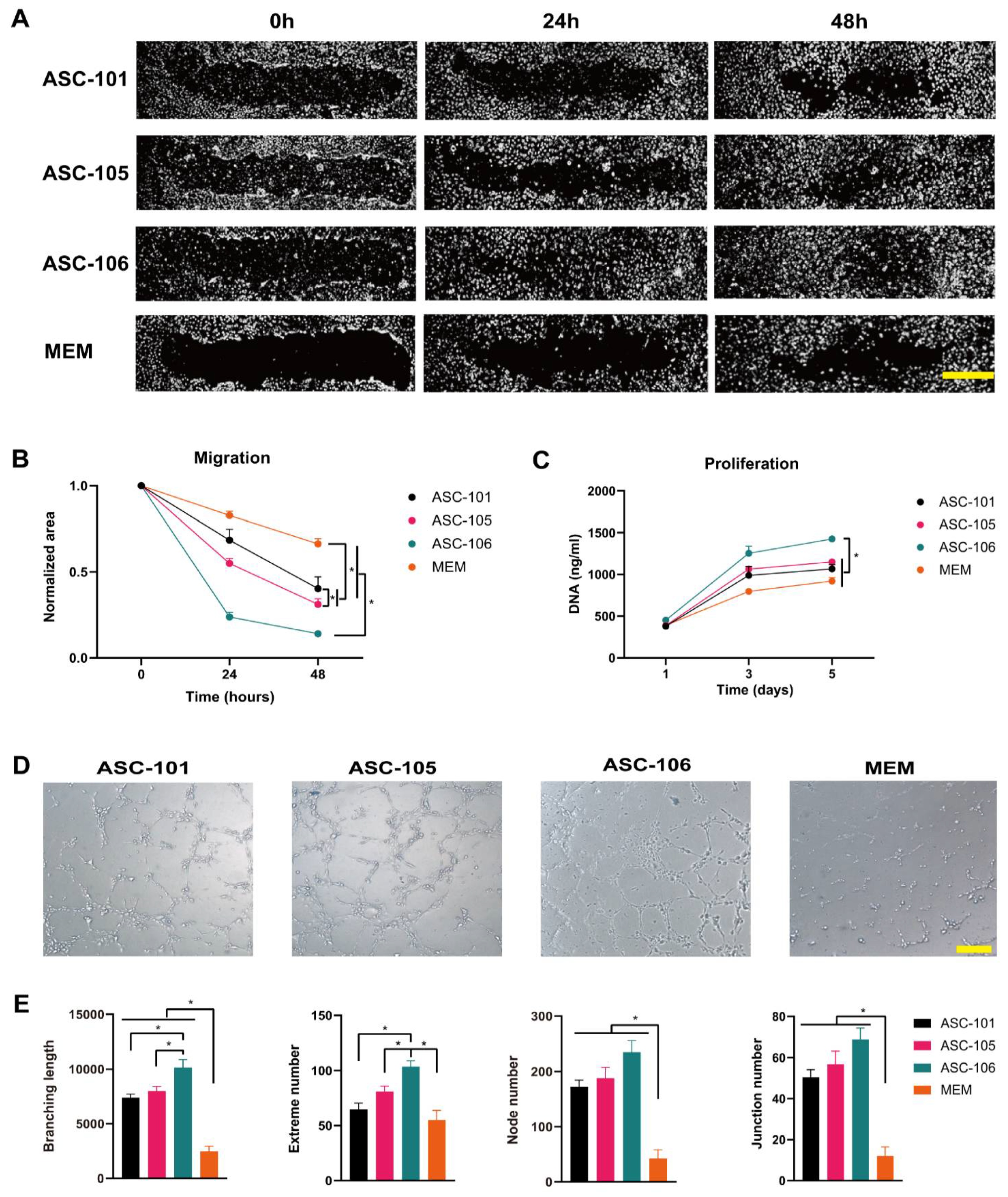
2.6. Migration
2.7. Tube Formation
2.8. RNA Sequencing
2.9. Statistical Analysis
3. Results
3.1. Identification of ASCs and Characteristic Comparison
3.2. Donor-Dependent Difference in ASC Function during Wound Healing
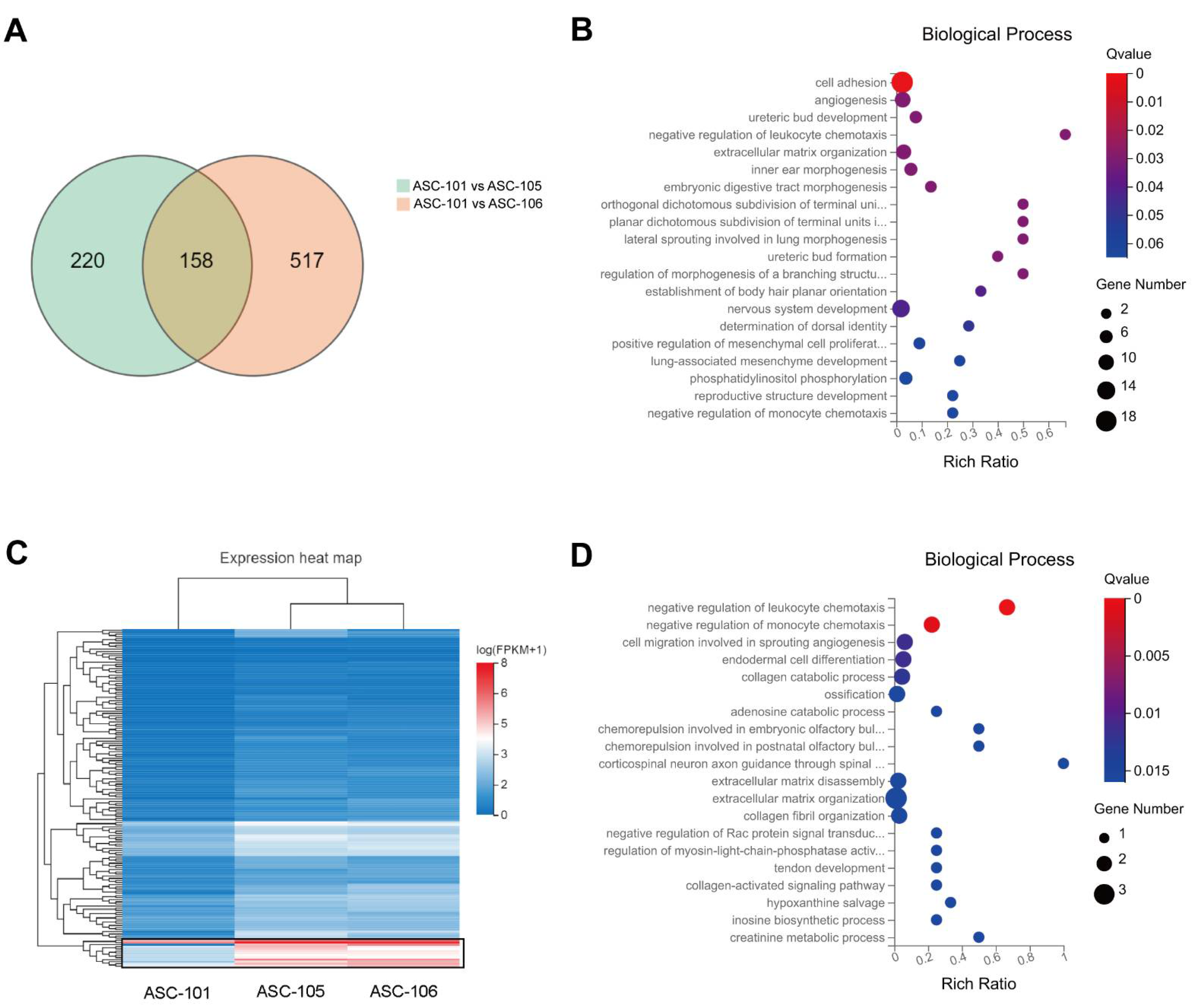
3.3. Differences in Gene Expression between Donors and GO Analysis
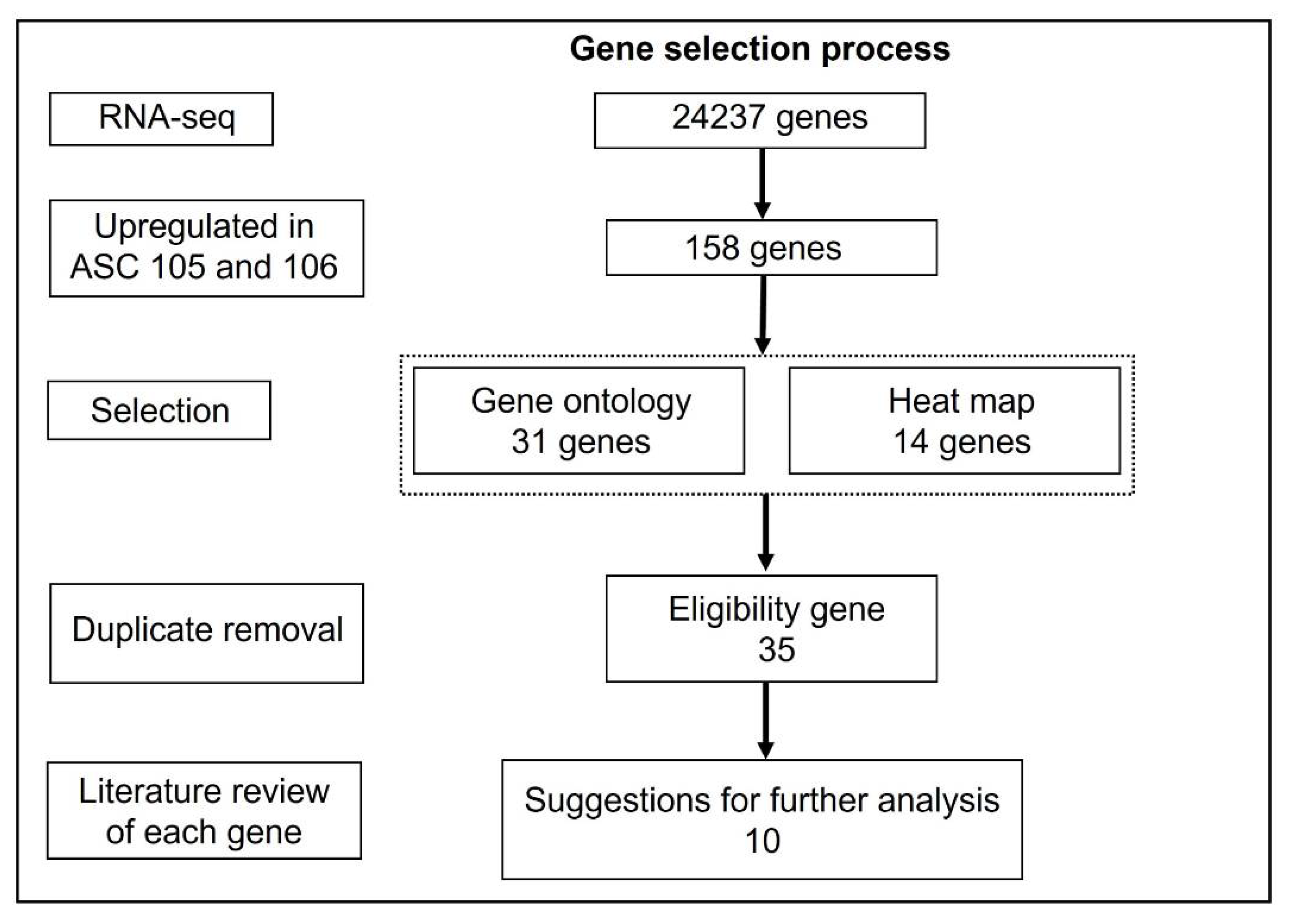
3.4. Selection of Putative Markers Predicting Wound Healing Capacity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Menke, N.B.; Ward, K.R.; Witten, T.M.; Bonchev, D.G.; Diegelmann, R.F. Impaired wound healing. Clin. Dermatol. 2007, 25, 19–25. [Google Scholar] [CrossRef]
- Gosain, A.; DiPietro, L.A. Aging and Wound Healing. World J. Surg. 2004, 28, 321–326. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhao, J.; Liu, J.; Gong, N.; Chen, L.; Margolis, D.J.; Bartus, C.; Hoffstad, O.; Malay, S.; Berlin, J.A.; et al. Adipose tissue-derived stem cells in clinical applications. Expert Opin. Biol. Ther. 2013, 13, 1357–1370. [Google Scholar]
- Cherubino, M.; Rubin, J.P.; Miljkovic, N.; Kelmendi-Doko, A.; Marra, K.G. Adipose-derived stem cells for wound healing applications. Ann. Plast. Surg. 2011, 66, 210–215. [Google Scholar] [CrossRef]
- Zuk, P. Adipose-Derived Stem Cells in Tissue Regeneration: A Review. ISRN Stem Cells 2013, 2013, 713959. [Google Scholar] [CrossRef] [Green Version]
- Isakson, M.; de Blacam, C.; Whelan, D.; McArdle, A.; Clover, A.J.P. Mesenchymal Stem Cells and Cutaneous Wound Healing: Current Evidence and Future Potential. Stem Cells Int. 2015, 2015, 831095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamora, D.O.; Natesan, S.; Becerra, S.; Wrice, N.; Chung, E.; Suggs, L.J.; Christy, R.J. Enhanced wound vascularization using a dsASCs seeded FPEG scaffold. Angiogenesis 2013, 16, 745–757. [Google Scholar] [CrossRef]
- Zografou, A.; Papadopoulos, O.; Tsigris, C.; Kavantzas, N.; Michalopoulos, E.; Chatzistamatiou, T.; Papassavas, A.; Stavropoulou-Gioka, C.; Dontas, I.; Perrea, D. Autologous Transplantation of Adipose-Derived Stem Cells Enhances Skin Graft Survival and Wound Healing in Diabetic Rats. Ann. Plast. Surg. 2013, 71, 225–232. [Google Scholar] [CrossRef]
- Khairoun, M.; Korevaar, S.S.; Roemeling-van Rhijn, M.; Khairoun, M.; Korevaar, S.; Lievers, E.; Leuning, D.; Ijzermans, J.; Betjes, M.; Genever, P.; et al. Human Bone Marrow- and Adipose Tissue-derived Mesenchymal Stromal Cells are Immunosuppressive In vitro and in a Humanized Allograft Rejection Model. J. Stem Cell Res. 2013, 1 (Suppl. S6), 20780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, G.; Chen, B.; Zhang, X.; Chen, H. Human Adipose-Derived Mesenchymal Stem Cells-Derived Exosomal microRNA-19b Promotes the Healing of Skin Wounds Through Modulation of the CCL1/TGF-β Signaling Axis. Clin. Cosmet. Investig. Dermatol. 2020, 13, 957–971. [Google Scholar] [CrossRef]
- Lu, Y.; Wen, H.; Huang, J.; Liao, P.; Liao, H.; Tu, J.; Zeng, Y. Extracellular vesicle-enclosed miR-486-5p mediates wound healing with adipose-derived stem cells by promoting angiogenesis. J. Cell. Mol. Med. 2020, 24, 9590–9604. [Google Scholar] [CrossRef]
- Moon, M.H.; Kim, S.Y.; Kim, Y.J.; Kim, S.J.; Lee, J.B.; Bae, Y.C.; Sung, S.M.; Jung, J.S. Human Adipose Tissue-Derived Mesenchymal Stem Cells Improve Postnatal Neovascularization in a Mouse Model of Hindlimb Ischemia. Cell. Physiol. Biochem. 2006, 17, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Lim, J.S.; Jun, J.B.; Choi, W.; Hong, I.S.; Jun, H.S. Exendin-4 in combination with adipose-derived stem cells promotes angiogenesis and improves diabetic wound healing. J. Transl. Med. 2017, 15, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brett, E.; Zielins, E.R.; Chin, M.; Januszyk, M.; Blackshear, C.P.; Findlay, M.; Momeni, A.; Gurtner, G.C.; Longaker, M.T.; Wan, D.C. Isolation of CD248-expressing stromal vascular fraction for targeted improvement of wound healing. Wound Repair Regen. 2017, 25, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Wang, J.; Zhou, X.; Xiong, Z.; Zhao, J.; Yu, R.; Huang, F.; Zhang, H.; Chen, L. Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts. Sci. Rep. 2016, 6, 32993. [Google Scholar] [CrossRef] [Green Version]
- Rehman, J.; Traktuev, D.; Li, J.; Merfeld-Clauss, S.; Temm-Grove, C.J.; Bovenkerk, J.E.; Pell, C.L.; Johnstone, B.H.; Considine, R.V.; March, K.L. Secretion of angiogenic and antiapoptotic factors by human adipose stromal cells. Circulation 2004, 109, 1292–1298. [Google Scholar] [CrossRef]
- Riis, S.; Zachar, V.; Boucher, S.; Vemuri, M.C.; Pennisi, C.P.; Fink, T. Critical steps in the isolation and expansion of adipose-derived stem cells for translational therapy. Expert Rev. Mol. Med. 2015, 17, e11. [Google Scholar] [CrossRef] [PubMed]
- Roemeling-Van Rhijn, M.; Mensah, F.K.F.; Korevaar, S.S.; Leijs, M.J.; Van Osch, G.J.V.M.; IJzermans, J.N.M.; Betjes, M.G.H.; Baan, C.C.; Weimar, W.; Hoogduijn, M.J. Effects of Hypoxia on the Immunomodulatory Properties of Adipose Tissue-Derived Mesenchymal Stem cells. Front. Immunol. 2013, 4, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, J.G.; Riis, S.E.; Frøbert, O.; Yang, S.; Kastrup, J.; Zachar, V.; Simonsen, U.; Fink, T. Activation of Protease-Activated Receptor 2 Induces VEGF Independently of HIF-1. PLoS ONE 2012, 7, e46087. [Google Scholar] [CrossRef] [Green Version]
- Thangarajah, H.; Vial, I.N.; Chang, E.; El-Ftesi, S.; Januszyk, M.; Chang, E.I.; Paterno, J.; Neofytou, E.; Longaker, M.T.; Gurtner, G.C. IFATS Collection: Adipose Stromal Cells Adopt a Proangiogenic Phenotype Under the Influence of Hypoxia. Stem Cells 2009, 27, 266–274. [Google Scholar] [CrossRef]
- Rubina, K.; Kalinina, N.; Efimenko, A.; Lopatina, T.; Melikhova, V.; Tsokolaeva, Z.; Sysoeva, V.; Tkachuk, V.; Parfyonova, Y. Adipose stromal cells stimulate angiogenesis via promoting progenitor cell differentiation, secretion of angiogenic factors, and enhancing vessel maturation. Tissue Eng. Part A 2009, 15, 2039–2050. [Google Scholar] [CrossRef] [Green Version]
- Efimenko, A.; Starostina, E.; Kalinina, N.; Stolzing, A. Angiogenic properties of aged adipose derived mesenchymal stem cells after hypoxic conditioning. J. Transl. Med. 2011, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Gao, J.; Yuan, Y.; Chang, Q.; Liao, Y.; Lu, F. Hypoxia preconditioned human adipose derived mesenchymal stem cells enhance angiogenic potential via secretion of increased VEGF and bFGF. Cell Biol. Int. 2013, 37, 551–560. [Google Scholar] [CrossRef]
- Lee, H.C.; An, S.G.; Lee, H.W.; Park, J.-S.; Cha, K.S.; Hong, T.J.; Park, J.H.; Lee, S.Y.; Kim, S.-P.; Kim, Y.D.; et al. Safety and Effect of Adipose Tissue-Derived Stem Cell Implantation in Patients With Critical Limb Ischemia. Circ. J. 2012, 76, 1750–1760. [Google Scholar] [CrossRef] [Green Version]
- Riis, S.; Newman, R.; Ipek, H.; Andersen, J.I.; Kuninger, D.; Boucher, S.; Vemuri, M.C.; Pennisi, C.P.; Zachar, V.; Fink, T. Hypoxia enhances the wound-healing potential of adipose-derived stem cells in a novel human primary keratinocyte-based scratch assay. Int. J. Mol. Med. 2017, 39, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Riis, S.; Stensballe, A.; Emmersen, J.; Pennisi, C.P.; Birkelund, S.; Zachar, V.; Fink, T. Mass spectrometry analysis of adipose-derived stem cells reveals a significant effect of hypoxia on pathways regulating extracellular matrix. Stem Cell Res. Ther. 2016, 7, 52. [Google Scholar] [CrossRef] [Green Version]
- Alt, E.U.; Senst, C.; Murthy, S.N.; Slakey, D.P.; Dupin, C.L.; Chaffin, A.E.; Kadowitz, P.J.; Izadpanah, R. Aging alters tissue resident mesenchymal stem cell properties. Stem Cell Res. 2012, 8, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor age negatively impacts adipose tissue-derived mesenchymal stem cell expansion and differentiation. J. Transl. Med. 2014, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- De Girolamo, L.; Lopa, S.; Arrigoni, E.; Sartori, M.F.; Baruffaldi Preis, F.W.; Brini, A.T. Human adipose-derived stem cells isolated from young and elderly women: Their differentiation potential and scaffold interaction during in vitro osteoblastic differentiation. Cytotherapy 2009, 11, 793–803. [Google Scholar] [CrossRef]
- Van Harmelen, V.; Röhrig, K.; Hauner, H. Comparison of proliferation and differentiation capacity of human adipocyte precursor cells from the omental and subcutaneous adipose tissue depot of obese subjects. Metabolism 2004, 53, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Xiong, H.; Chen, J.; Yang, X.; Liu, Y.; Guo, J.; Jiang, T.; Xu, Z.; Yuan, M.; Liu, Y.; et al. The whole profiling and competing endogenous RNA network analyses of noncoding RNAs in adipose-derived stem cells from diabetic, old, and young patients. Stem Cell Res. Ther. 2021, 12, 313. [Google Scholar] [CrossRef]
- De Barros, S.; Dehez, S.; Arnaud, E.; Barreau, C.; Cazavet, A.; Perez, G.; Galinier, A.; Casteilla, L.; Planat-Bénard, V. Aging-related decrease of human ASC angiogenic potential is reversed by hypoxia preconditioning through ROS production. Mol. Ther. 2013, 21, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Aksu, A.E.; Rubin, J.P.; Dudas, J.R.; Marra, K.G. Role of gender and anatomical region on induction of osteogenic differentiation of human adipose-derived stem cells. Ann. Plast. Surg. 2008, 60, 306–322. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, R.; Mizuno, H.; Watanabe, A.; Migita, M.; Hyakusoku, H.; Shimada, T. Adipogenic differentiation by adipose-derived stem cells harvested from GFP transgenic mice-Including relationship of sex differences. Biochem. Biophys. Res. Commun. 2004, 319, 511–517. [Google Scholar] [CrossRef]
- Juntunen, M.; Heinonen, S.; Huhtala, H.; Rissanen, A.; Kaprio, J.; Kuismanen, K.; Pietiläinen, K.H.; Miettinen, S.; Patrikoski, M. Evaluation of the effect of donor weight on adipose stromal/stem cell characteristics by using weight-discordant monozygotic twin pairs. Stem Cell Res. Ther. 2021, 12, 516. [Google Scholar] [CrossRef]
- Mohiuddin, O.A.; Motherwell, J.M.; Rogers, E.; Bratton, M.R.; Zhang, Q.; Wang, G.; Bunnell, B.; Hayes, D.J.; Gimble, J.M. Characterization and Proteomic Analysis of Decellularized Adipose Tissue Hydrogels Derived from Lean and Overweight/Obese Human Donors. Adv. Biosyst. 2020, 4, e2000124. [Google Scholar] [CrossRef]
- Stroncek, D.F.; Jin, P.; Wang, E.; Jett, B.; Zielins, E.R.; Atashroo, D.A.; Maan, Z.N.; Duscher, D.; Walmsley, G.G.; Hu, M.; et al. Potency analysis of cellular therapies: The emerging role of molecular assays. J. Transl. Med. 2016, 5, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nations, U.; Programme, E.; Criteria, E.H.; Management, S.; Programme, T.I.; Safety, C.; Co-operation, E.; Organizations, P.; Conference, U.N.; Library, W.H.O.; et al. Biomarkers in Risk Assessment: Validity and Validation; WHO: Geneva, Switzerland, 2014; pp. 1–21. [Google Scholar]
- Haack-Sørensen, M.; Juhl, M.; Follin, B.; Harary Søndergaard, R.; Kirchhoff, M.; Kastrup, J.; Ekblond, A. Development of large-scale manufacturing of adipose-derived stromal cells for clinical applications using bioreactors and human platelet lysate. Scand. J. Clin. Lab. Investig. 2018, 78, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Mohamed-Ahmed, S.; Fristad, I.; Lie, S.A.; Suliman, S.; Mustafa, K.; Vindenes, H.; Idris, S.B. Adipose-derived and bone marrow mesenchymal stem cells: A donor-matched comparison. Stem Cell Res. Ther. 2018, 9, 168. [Google Scholar] [CrossRef] [PubMed]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics (IFATS) and Science and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641. [Google Scholar]
- Wu, X.Y.; Xu, H.; Wu, Z.F.; Chen, C.; Liu, J.Y.; Wu, G.N.; Yao, X.Q.; Liu, F.K.; Li, G.; Shen, L. Formononetin, a novel FGFR2 inhibitor, potently inhibits angiogenesis and tumor growth in preclinical models. Oncotarget 2015, 6, 44563. [Google Scholar] [CrossRef]
- Xu, N.; Brodin, P.; Wei, T.; Meisgen, F.; Eidsmo, L.; Nagy, N.; Kemeny, L.; Ståhle, M.; Sonkoly, E.; Pivarcsi, A. MiR-125b, a MicroRNA Downregulated in Psoriasis, Modulates Keratinocyte Proliferation by Targeting FGFR2. J. Investig. Dermatol. 2011, 131, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Kaabeche, K.; Lemonnier, J.; Le Mée, S.; Caverzasio, J.; Marie, P.J. Cbl-mediated degradation of Lyn and Fyn induced by constitutive fibroblast growth factor receptor-2 activation supports osteoblast differentiation. J. Biol. Chem. 2004, 279, 36259–36267. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Kakudo, N.; Morimoto, N.; Lai, F.; Taketani, S.; Kusumoto, K. Fibroblast growth factor-2 stimulates proliferation of human adipose-derived stem cells via Src activation. Stem Cell Res. Ther. 2019, 10, 350. [Google Scholar] [CrossRef] [PubMed]
- Moyle, M.; Napier, M.A.; McLean, J.W. Cloning and expression of a divergent integrin subunit beta 8. J. Biol. Chem. 1991, 266, 19650–19658. [Google Scholar] [CrossRef]
- Mu, D.; Cambier, S.; Fjellbirkeland, L.; Baron, J.L.; Munger, J.S.; Kawakatsu, H.; Sheppard, D.; Courtney Broaddus, V.; Nishimura, S.L. The integrin alpha(v)beta8 mediates epithelial homeostasis through MT1-MMP-dependent activation of TGF-beta1. J. Cell Biol. 2002, 157, 493–507. [Google Scholar] [CrossRef] [Green Version]
- Gaudet, P.; Livstone, M.S.; Lewis, S.E.; Thomas, P.D. Phylogenetic-based propagation of functional annotations within the Gene Ontology consortium. Brief. Bioinform. 2011, 12, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Pankov, R.; Yamada, K.M. Fibronectin at a glance. J. Cell Sci. 2002, 115, 3861–3863. [Google Scholar] [CrossRef] [Green Version]
- LaPointe, V.L.S.; Verpoorte, A.; Stevens, M.M. The Changing Integrin Expression and a Role for Integrin β8 in the Chondrogenic Differentiation of Mesenchymal Stem Cells. PLoS ONE 2013, 8, e82035. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Kim, W.K.; Oh, K.J.; Lee, E.W.; Han, B.S.; Lee, S.C.; Bae, K.H. Protein Tyrosine Phosphatase, Receptor Type B (PTPRB) Inhibits Brown Adipocyte Differentiation through Regulation of VEGFR2 Phosphorylation. J. Microbiol. Biotechnol. 2019, 29, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Bollag, R.J.; Siegfried, Z.; Cebra-Thomas, J.A.; Garvey, N.; Davison, E.M.; Silver, L.M. An ancient family of embryonically expressed mouse genes sharing a conserved protein motif with the T locus. Nat. Genet. 1994, 7, 383–389. [Google Scholar] [CrossRef]
- Markan, K.R.; Boland, L.K.; King-McAlpin, A.Q.; Claflin, K.E.; Leaman, M.P.; Kemerling, M.K.; Stonewall, M.M.; Amendt, B.A.; Ankrum, J.A.; Potthoff, M.J. Adipose TBX1 regulates β-adrenergic sensitivity in subcutaneous adipose tissue and thermogenic capacity in vivo. Mol. Metab. 2020, 36, 100965. [Google Scholar] [CrossRef]
- Liu, M.; Lei, H.; Dong, P.; Fu, X.; Yang, Z.; Yang, Y.; Ma, J.; Liu, X.; Cao, Y.; Xiao, R. Adipose-Derived Mesenchymal Stem Cells from the Elderly Exhibit Decreased Migration and Differentiation Abilities with Senescent Properties. Cell Transplant. 2017, 26, 1505–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Yu, J.; Pan, Q.; Yang, J.; Hao, G.; Wang, Y.; Li, L.; Cao, H. Hypoxia-inducible factor-2 alpha promotes the proliferation of human placenta-derived mesenchymal stem cells through the MAPK/ERK signaling pathway. Sci. Rep. 2016, 6, 35489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joannes, A.; Brayer, S.; Besnard, V.; Marchal-Sommé, J.; Jaillet, M.; Mordant, P.; Mal, H.; Borie, R.; Crestani, B.; Mailleux, A.A. FGF9 and FGF18 in idiopathic pulmonary fibrosis promote survival and migration and inhibit myofibroblast differentiation of human lung fibroblasts in vitro. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L615–L629. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zeng, Q.S.; Zou, M.; Zeng, J.; Nie, J.; Chen, D.F.; Gan, H.T. Targeting Gremlin 1 Prevents Intestinal Fibrosis Progression by Inhibiting the Fatty Acid Oxidation of Fibroblast Cells. Front. Pharmacol. 2021, 12, 663774. [Google Scholar] [CrossRef]
- O’Reilly, S.; Ciechomska, M.; Cant, R.; Van Laar, J.M. Interleukin-6 (IL-6) trans signaling drives a STAT3-dependent pathway that leads to hyperactive transforming growth factor-β (TGF-β) signaling promoting SMAD3 activation and fibrosis via Gremlin protein. J. Biol. Chem. 2014, 289, 9952–9960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffy, L.; Henderson, J.; Brown, M.; Pryzborski, S.; Fullard, N.; Summa, L.; Distler, J.H.W.; Stratton, R.; O’Reilly, S. Bone Morphogenetic Protein Antagonist Gremlin-1 Increases Myofibroblast Transition in Dermal Fibroblasts: Implications for Systemic Sclerosis. Front. Cell Dev. Biol. 2021, 9, 1451. [Google Scholar] [CrossRef]
- O’Reilly, S. Gremlin: A complex molecule regulating wound healing and fibrosis. Cell. Mol. Life Sci. 2021, 78, 7917–7923. [Google Scholar] [CrossRef]
- Puxeddu, I.; Bader, R.; Piliponsky, A.M.; Reich, R.; Levi-Schaffer, F.; Berkman, N. The CC chemokine eotaxin/CCL11 has a selective profibrogenic effect on human lung fibroblasts. J. Allergy Clin. Immunol. 2006, 117, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Olczyk, P.; Mencner, Ł.; Komosinska-Vassev, K. The role of the extracellular matrix components in cutaneous wound healing. Biomed. Res. Int. 2014, 2014, 747584. [Google Scholar] [CrossRef]
- Clark, R.A.F. Biology of Dermal Wound Repair. Dermatol. Clin. 1993, 11, 647–666. [Google Scholar] [CrossRef]
- Nishikai-Yan Shen, T.; Kado, M.; Hagiwara, H.; Fujimura, S.; Mizuno, H.; Tanaka, R. MMP9 secreted from mononuclear cell quality and quantity culture mediates STAT3 phosphorylation and fibroblast migration in wounds. Regen. Ther. 2021, 18, 464–471. [Google Scholar] [CrossRef]
- Xue, M.; McKelvey, K.; Shen, K.; Minhas, N.; March, L.; Park, S.Y.; Jackson, C.J. Endogenous MMP-9 and not MMP-2 promotes rheumatoid synovial fibroblast survival, inflammation and cartilage degradation. Rheumatology 2014, 53, 2270–2279. [Google Scholar] [CrossRef] [Green Version]
- Tracy, L.E.; Minasian, R.A.; Caterson, E.J. Extracellular Matrix and Dermal Fibroblast Function in the Healing Wound. Adv. Wound Care 2016, 5, 119. [Google Scholar] [CrossRef]
- Chen, P.; Parks, W.C. Role of Matrix Metalloproteinases in Epithelial Migration. J. Cell. Biochem. 2009, 108, 1233. [Google Scholar] [CrossRef] [PubMed]
- Herrera, I.; Cisneros, J.; Maldonado, M.; Ramírez, R.; Ortiz-Quintero, B.; Anso, E.; Chandel, N.S.; Selman, M.; Pardo, A. Matrix Metalloproteinase (MMP)-1 Induces Lung Alveolar Epithelial Cell Migration and Proliferation, Protects from Apoptosis, and Represses Mitochondrial Oxygen Consumption. J. Biol. Chem. 2013, 288, 25964–25975. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Stamenkovic, I. Cell surface-localized matrix metalloproteinase-9 proteolytically activates TGF-beta and promotes tumor invasion and angiogenesis. Genes Dev. 1999, 13, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Ji, J.; Hou, J.; Tan, Y.; Han, X. Identification of Key Candidate Genes Involved in the Progression of Idiopathic Pulmonary Fibrosis. Molecules 2021, 26, 1123. [Google Scholar] [CrossRef]
- Kendall, R.T.; Feghali-Bostwick, C.A. Fibroblasts in fibrosis: Novel roles and mediators. Front. Pharmacol. 2014, 5, 123. [Google Scholar] [CrossRef] [Green Version]
- Behr, B.; Leucht, P.; Longaker, M.T.; Quarto, N. Fgf-9 is required for angiogenesis and osteogenesis in long bone repair. Proc. Natl. Acad. Sci. USA 2010, 107, 11853–11858. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, Y. Therapeutic uses of FGFs. Semin. Cell Dev. Biol. 2016, 53, 144–154. [Google Scholar] [CrossRef]
- Frontini, M.J.; Nong, Z.; Gros, R.; Drangova, M.; O’Neil, C.; Rahman, M.N.; Akawi, O.; Yin, H.; Ellis, C.G.; Pickering, J.G. Fibroblast growth factor 9 delivery during angiogenesis produces durable, vasoresponsive microvessels wrapped by smooth muscle cells. Nat. Biotechnol. 2011, 29, 421–427. [Google Scholar] [CrossRef]
- Wallner, C.; Schira, J.; Wagner, J.M.; Schulte, M.; Fischer, S.; Hirsch, T.; Richter, W.; Abraham, S.; Kneser, U.; Lehnhardt, M.; et al. Application of VEGFA and FGF-9 Enhances Angiogenesis, Osteogenesis and Bone Remodeling in Type 2 Diabetic Long Bone Regeneration. PLoS ONE 2015, 10, e0118823. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Khan, A.W.; Kim, M.S.; Choi, S. The Role of Fibroblast Growth Factor (FGF) Signaling in Tissue Repair and Regeneration. Cells 2021, 10, 3242. [Google Scholar] [CrossRef]
- Said, S.S.; O’Neil, C.; Yin, H.; Nong, Z.; Pickering, J.G.; Mequanint, K. Concurrent and Sustained Delivery of FGF2 and FGF9 from Electrospun Poly(ester amide) Fibrous Mats for Therapeutic Angiogenesis. Tissue Eng.-Part A 2016, 22, 584–596. [Google Scholar] [CrossRef]
- Mitola, S.; Ravelli, C.; Moroni, E.; Salvi, V.; Leali, D.; Ballmer-Hofer, K.; Zammataro, L.; Presta, M. Gremlin is a novel agonist of the major proangiogenic receptor VEGFR2. Blood 2010, 116, 3677–3680. [Google Scholar] [CrossRef] [Green Version]
- Corsini, M.; Moroni, E.; Ravelli, C.; Andrés, G.; Grillo, E.; Ali, I.H.; Brazil, D.P.; Presta, M.; Mitola, S. Cyclic adenosine monophosphate-response element-binding protein mediates the proangiogenic or proinflammatory activity of gremlin. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Salcedo, R.; Young, H.A.; Ponce, M.L.; Ward, J.M.; Kleinman, H.K.; Murphy, W.J.; Oppenheim, J.J. Eotaxin (CCL11) Induces In Vivo Angiogenic Responses by Human CCR3+ Endothelial Cells. J. Immunol. 2001, 166, 7571–7578. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Kang, Y.W.; Choi, B.Y.; Yang, Y.C.; Cho, B.P.; Cho, W.G. CCL11 promotes angiogenic activity by activating the PI3K/Akt pathway in HUVECs. J. Recept. Signal Transduct. Res. 2017, 37, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Ammann, K.R.; DeCook, K.J.; Li, M.; Slepian, M.J. Migration versus proliferation as contributor to in vitro wound healing of vascular endothelial and smooth muscle cells. Exp. Cell Res. 2019, 376, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Sottile, J. Regulation of angiogenesis by extracellular matrix. Biochim. Biophys. Acta 2004, 1654, 13–22. [Google Scholar] [CrossRef]
- Song, Y.H.; Shon, S.H.; Shan, M.; Stroock, A.D.; Fischbach, C. Adipose-derived Stem Cells Increase Angiogenesis through Matrix Metalloproteinase-dependent Collagen Remodeling. Integr. Biol. 2016, 8, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazor, R.; Alsaigh, T.; Shaked, H.; Altshuler, A.E.; Pocock, E.S.; Kistler, E.B.; Karin, M.; Schmid-Schönbein, G.W. Matrix metalloproteinase-1-mediated up-regulation of vascular endothelial growth factor-2 in endothelial cells. J. Biol. Chem. 2013, 288, 598–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergers, G.; Brekken, R.; McMahon, G.; Vu, T.H.; Itoh, T.; Tamaki, K.; Tanzawa, K.; Thorpe, P.; Itohara, S.; Werb, Z.; et al. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nat. Cell Biol. 2000, 2, 737–744. [Google Scholar] [CrossRef]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez-Sánchez, M.E. Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Front. Oncol. 2019, 9, 1370. [Google Scholar] [CrossRef] [Green Version]
- Lamalice, L.; Le Boeuf, F.; Huot, J. Endothelial cell migration during angiogenesis. Circ. Res. 2007, 100, 782–794. [Google Scholar] [CrossRef] [PubMed]
- Bonanno, E.; Iurlaro, M.; Madri, J.A.; Nicosia, R.F. Type IV collagen modulates angiogenesis and neovessel survival in the rat aorta model. In Vitro Cell. Dev. Biol. Anim. 2000, 36, 336–340. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Marker | ASC-101 | ASC-105 | ASC-106 |
|---|---|---|---|---|
| MSC markers | CD105 | 77.73 | 97.65 | 96.43 |
| CD73 | 98.60 | 99.71 | 97.99 | |
| CD90 | 99.95 | 99.96 | 99.92 | |
| ASC markers | CD146 | 27.33 | 22.52 | 7.99 |
| CD34 | 60.41 | 72.94 | 61.01 | |
| CD31 | 0.77 | 2.33 | 7.94 | |
| Differentiation capacity | Stro-1 | 13.38 | 14.38 | 28.36 |
| CD201 | 98.65 | 96.44 | 98.08 | |
| CD36 | 13.67 | 8.33 | 11.74 | |
| Immune regulation | CD29 | 99.99 | 100 | 99.69 |
| CD200 | 2.29 | 6.77 | 7.37 | |
| CD274 | 94.79 | 92.93 | 60.28 | |
| Wound healing | CD248 | 98.68 | 98.87 | 95.66 |
| CD166 | 99.39 | 99.62 | 99.03 | |
| CD271 | 59.61 | 24.98 | 23.12 |
| Description | Gene | Go Term |
|---|---|---|
| ECM and tissue development | ||
| Cell adhesion | TENM2, HAPLN1, SVEP1, PCDHB3, PGM5, CADM3, PCDHB2, SPON1, EFS, CCL11, LZTS1, ITGB8, CELSR1, NUAK1, PDZD2, ITGA11, PCDH19 | GO:0007155 |
| Cell-matrix adhesion | ITGB8, ITGA11 | GO:0007160 |
| ECM | MMP1, LOC102724770-DGCR6, COL4A4, HAPLN1, RARRES2, | GO:0031012 |
| ECM disassembly | MMP1 | GO:0022617 |
| ECM organization | MMP9, TNFRSF11B, COL10A1, COL11A1, ITGB8, ITGA11 | GO:0030198 |
| Extracellular region | ADA2 | GO:0005576 |
| Collagen-containing extracellular matrix | MMP9, COL10A1 | GO:0062023 |
| Collagen fibril organization | COL11A1 | GO:0030199 |
| Collagen catabolic process | MMP1 | GO:0030574 |
| Angiogenesis development | ||
| Angiogenesis | FGF9, TBX1, FGFR4, GREM1, FGFR2, EPAS1, PTPRB | GO:0001525 |
| Positive regulation of angiogenesis | PIK3R6, GREM1, CCL11, ITGB8 | GO:0045766 |
| Endothelial cell morphogenesis | COL4A4 | GO:0001886 |
| Blood vessel morphogenesis/development | TBX1 | GO:0048514 GO:0001568 |
| Blood vessel remodeling | EPAS1 | GO:0001974 |
| Differentiation | ||
| Cell differentiation | FGF9, RARRES2, FGFR4, FGFR2, EPAS1 | GO:0030154 |
| Epithelial cell differentiation | TBX1 | GO:0030855 |
| Positive regulation of fat cell differentiation | RARRES2 | GO:0045600 |
| Osteoblast differentiation | FGF9, FGFR2, ITGA11 | GO:0001649 GO:0045667 |
| Mesenchymal cell differentiation | FGFR2 | GO:0048762 |
| Proliferation | ||
| Positive regulation of cell proliferation | TBX1, FGF9, TNFRSF11B, FGFR4, GREM1, NUAK1 | GO:0008284 |
| Positive regulation of mesenchymal cell proliferation | TBX1, FGFR2 | GO:0002053 |
| Positive regulation of epithelial cell proliferation | TBX1, FGF9, FGFR2 | GO:0050679 |
| Positive regulation of endothelial cell proliferation | CCL11 | GO:0001938 |
| Migration | ||
| Positive regulation of cell migration | MMP9, FGFR4, EFS, CCL11 | GO:0030335 GO:0016477 |
| Positive regulation of keratinocyte migration | MMP9 | GO:0051549 |
| Endothelial cell migration | GREM1 | GO:0043542 |
| Cell migration involved in sprouting angiogenesis | GREM1, SLIT2 | GO:0002042 |
| Others | ||
| Wound healing | FGFR2, CELSR1 | GO:0042060 |
| Chondrocyte development | COL11A1 | GO:0002063 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, G.; Peng, Q.; Emmersen, J.; Zachar, V.; Fink, T.; Porsborg, S.R. A Comparative Analysis of the Wound Healing-Related Heterogeneity of Adipose-Derived Stem Cells Donors. Pharmaceutics 2022, 14, 2126. https://doi.org/10.3390/pharmaceutics14102126
Ren G, Peng Q, Emmersen J, Zachar V, Fink T, Porsborg SR. A Comparative Analysis of the Wound Healing-Related Heterogeneity of Adipose-Derived Stem Cells Donors. Pharmaceutics. 2022; 14(10):2126. https://doi.org/10.3390/pharmaceutics14102126
Chicago/Turabian StyleRen, Guoqiang, Qiuyue Peng, Jeppe Emmersen, Vladimir Zachar, Trine Fink, and Simone R. Porsborg. 2022. "A Comparative Analysis of the Wound Healing-Related Heterogeneity of Adipose-Derived Stem Cells Donors" Pharmaceutics 14, no. 10: 2126. https://doi.org/10.3390/pharmaceutics14102126