
Characterization and Differential Cytotoxicity of Gramicidin Nanoparticles Combined with Cationic Polymer or Lipid Bilayer

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Gramicidin and Gramicidin/Poly (Diallyldimethylammonium Chloride) Dispersions in Water
2.3. Preparation of Supported Dioctadecyldimethylammonium Bromide (DODAB) Bilayers on Silica Particles with or without Gramicidin Inserted in the DODAB Bilayer
2.4. Determination of Physical Properties of Gr, Gr/PDDA, or SiO2/DODAB/Gr Dispersions
2.5. Determination of Colloidal Stability
2.6. Determination of Circular Dichroism Spectra for Gramicidin in Ethanol, Trifluoroethanol, Water, and Poly (Diallyldimethylammonium) Chloride Solutions
2.7. Scanning Electron Microscopy for Gr or Gr/PDDA Dispersions
2.8. Determination of Cell Viability for Staphylococcus aureus, Candida albicans, and Pseudomonas aeruginosa over a Range of Gramicidin, Gramicidin/Poly (Diallyldimethylammonium) Chloride, or Dioctadecyldimethylammonium Bromide Concentrations
3. Results
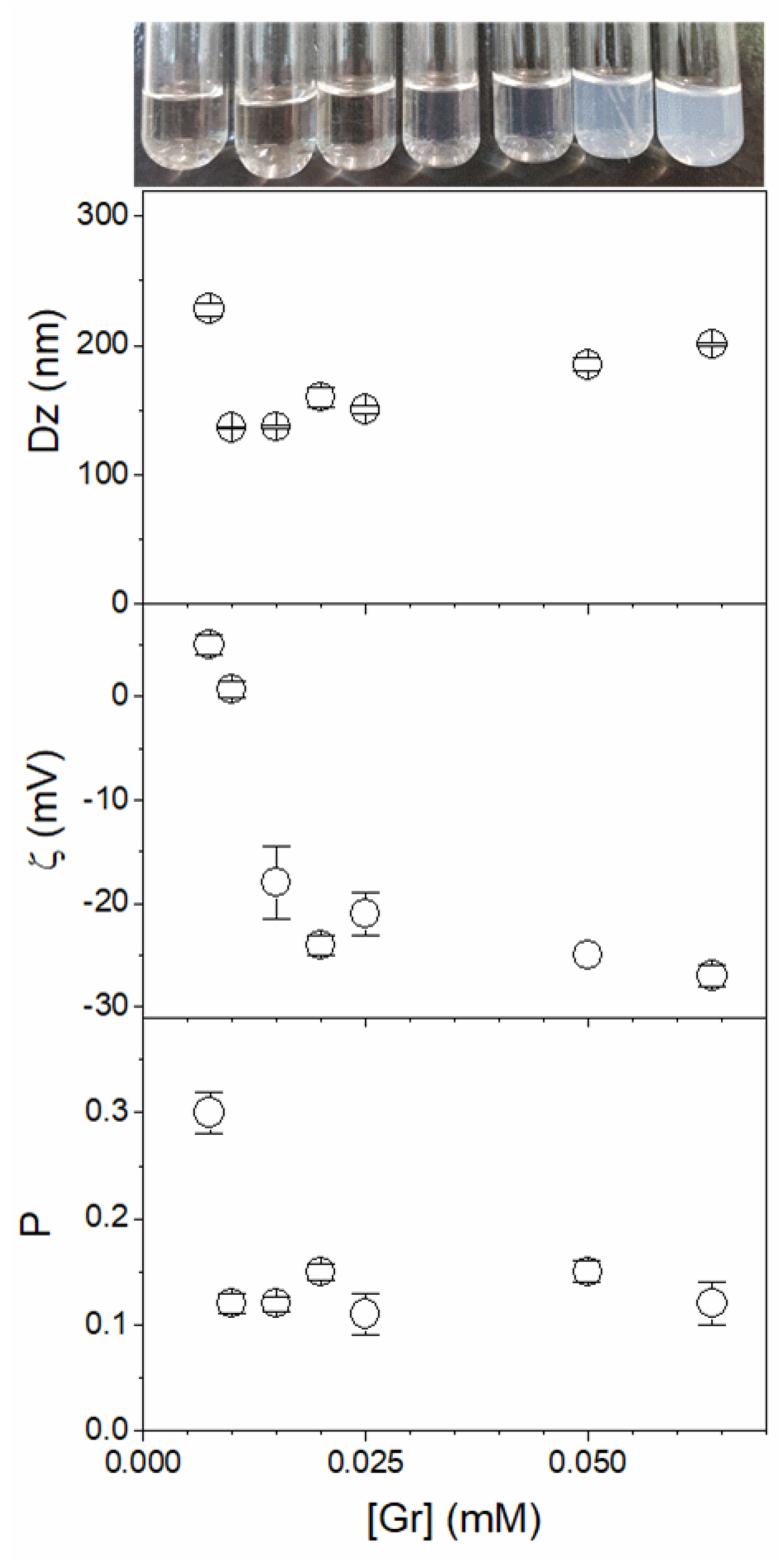
3.1. Assembly of Gramicidin and Gramicidin/Poly (Diallyldimethylammonium) Chloride in Water as Nanoparticles
3.2. Incorporation of Gramicidin in Dioctadecyldimethylammonium Bromide Bilayers Reduces Gramicidin Antimicrobial Activity against Staphylococcus aureus
3.3. Combinations of Gramicidin Nanoparticles (Gr NPs) and Poly (Diallyldimethyl Ammonium Chloride) (PDDA) in Water Dispersions Exhibit High and Broad Microbicidal Activity in Absence of Cytotoxicity against Mammalian Cells at Reduced Doses of Both Antimicrobials
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tian, X.; Sun, F.; Zhou, X.-R.; Luo, S.-Z.; Chen, L. Role of Peptide Self-Assembly in Antimicrobial Peptides. J. Pept. Sci. 2015, 21, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Tirrell, M. Polymorphism in Peptide Self-Assembly Visualized. Proc. Natl. Acad. Sci. USA 2022, 119, e2123197119. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus Epidermidis––The “accidental” Pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef]
- Dubos, R.J. Studies on a Bactericidal Agent Extracted from a Soil Bacillus. J. Exp. Med. 1939, 70, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Stein, W.D.; Litman, T. Chapter 3—Ion Channels Across Cell Membranes. In Channels, Carriers, and Pumps, 2nd ed.; Stein, W.D., Litman, T., Eds.; Elsevier: London, UK, 2015; pp. 81–130. ISBN 978-0-12-416579-3. [Google Scholar]
- Steinem, C.; Janshoff, A.; Ulrich, W.P.; Sieber, M.; Galla, H.J. Impedance Analysis of Supported Lipid Bilayer Membranes: A Scrutiny of Different Preparation Techniques. Biochim. Biophys. Acta 1996, 1279, 169–180. [Google Scholar] [CrossRef]
- Burkhart, B.M.; Gassman, R.M.; Langs, D.A.; Pangborn, W.A.; Duax, W.L.; Pletnev, V. Gramicidin D Conformation, Dynamics and Membrane Ion Transport. Biopolymers 1999, 51, 129–144. [Google Scholar] [CrossRef]
- Ketchem, R.R.; Lee, K.C.; Huo, S.; Cross, T.A. Macromolecular Structural Elucidation with Solid-State NMR-Derived Orientational Constraints. J. Biomol. NMR 1996, 8, 1–14. [Google Scholar] [CrossRef]
- Kelkar, D.A.; Chattopadhyay, A. The Gramicidin Ion Channel: A Model Membrane Protein. Biochim. Biophys. Acta (BBA)-Biomembr. 2007, 1768, 2011–2025. [Google Scholar] [CrossRef]
- Wang, F.; Qin, L.; Pace, C.J.; Wong, P.; Malonis, R.; Gao, J. Solubilized Gramicidin A as Potential Systemic Antibiotics. ChemBioChem 2012, 13, 51–55. [Google Scholar] [CrossRef]
- Sun, Z.; Barboiu, M. Artificial Gramicidins. Front. Chem. 2019, 7, 611. [Google Scholar] [CrossRef] [Green Version]
- Bamberg, E.; Noda, K.; Läuger, P. Single-Channel Parameters of Gramicidin A, B, and C. Biochim. Biophys. Acta (BBA)-Biomembr. 1976, 419, 223–228. [Google Scholar] [CrossRef]
- Svensson, F.R.; Lincoln, P.; Nordén, B.; Esbjörner, E.K. Tryptophan Orientations in Membrane-Bound Gramicidin and Melittin—A Comparative Linear Dichroism Study on Transmembrane and Surface-Bound Peptides. Biochim. Biophys. Acta (BBA)-Biomembr. 2011, 1808, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Lee, K.; Cross, T. Tryptophans in Membrane Proteins: Indole Ring Orientations and Functional Implications in the Gramicidin Channel. Biochemistry 1993, 32, 7035–7047. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wei, G.-W. A Review of Mathematical Modeling, Simulation and Analysis of Membrane Channel Charge Transport. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2017; ISBN 978-0-12-809633-8. [Google Scholar]
- Killian, J.A.; Burger, K.N.J.; de Kruijff, B. Phase Separation and Hexagonal HII Phase Formation by Gramicidins A, B and C in Dioleoylphosphatidylcholine Model Membranes. A Study on the Role of the Tryptophan Residues. Biochim. Biophys. Acta (BBA)–Biomembr. 1987, 897, 269–284. [Google Scholar] [CrossRef]
- Kelkar, D.A.; Chattopadhyay, A. Modulation of Gramicidin Channel Conformation and Organization by Hydrophobic Mismatch in Saturated Phosphatidylcholine Bilayers. Biochim. Biophys. Acta (BBA)-Biomembr. 2007, 1768, 1103–1113. [Google Scholar] [CrossRef]
- Schmitt, E.K.; Weichbrodt, C.; Steinem, C. Impedance Analysis of Gramicidin D in Pore-Suspending Membranes. Soft Matter 2009, 5, 3347–3353. [Google Scholar] [CrossRef]
- Ragioto, D.A.M.T.; Carrasco, L.D.M.; Carmona-Ribeiro, A.M. Novel Gramicidin Formulations in Cationic Lipid as Broad-Spectrum Microbicidal Agents. Int. J. Nanomed. 2014, 9, 3183–3192. [Google Scholar] [CrossRef]
- Pavithrra, G.; Rajasekaran, R. Gramicidin Peptide to Combat Antibiotic Resistance: A Review. Int. J. Pept. Res. Ther. 2020, 26, 191–199. [Google Scholar] [CrossRef]
- Ribeiro, R.T.; Braga, V.H.A.; Carmona-Ribeiro, A.M. Biomimetic Cationic Nanoparticles Based on Silica: Optimizing Bilayer Deposition from Lipid Films. Biomimetics 2017, 2, 20. [Google Scholar] [CrossRef]
- Sanches, L.M.; Petri, D.F.S.; de Melo Carrasco, L.D.; Carmona-Ribeiro, A.M. The Antimicrobial Activity of Free and Immobilized Poly (Diallyldimethylammonium) Chloride in Nanoparticles of Poly (Methylmethacrylate). J. Nanobiotechnol. 2015, 13, 58. [Google Scholar] [CrossRef] [Green Version]
- Kayitmazer, A.B.; Strand, S.P.; Tribet, C.; Jaeger, W.; Dubin, P.L. Effect of Polyelectrolyte Structure on Protein-Polyelectrolyte Coacervates: Coacervates of Bovine Serum Albumin with Poly(Diallyldimethylammonium Chloride) versus Chitosan. Biomacromolecules 2007, 8, 3568–3577. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zheng, K.; Si, Y.; Guo, X.; Xu, Y. Protein-Polyelectrolyte Interaction: Thermodynamic Analysis Based on the Titration Method (†). Polymers 2019, 11, 82. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Betancourt, Y.; Távora, B.C.L.F.; Colombini, M.; Faquim-Mauro, E.L.; Carmona-Ribeiro, A.M. Simple Nanoparticles from the Assembly of Cationic Polymer and Antigen as Immunoadjuvants. Vaccines 2020, 8, 105. [Google Scholar] [CrossRef]
- Borro, B.C.; Malmsten, M. Complexation between Antimicrobial Peptides and Polyelectrolytes. Adv. Colloid Interface Sci. 2019, 270, 251–260. [Google Scholar] [CrossRef]
- López, A.D.L.R.; Lee, M.-R.; Ortiz, B.J.; Gastfriend, B.D.; Whitehead, R.; Lynn, D.M.; Palecek, S.P. Preventing S. Aureus Biofilm Formation on Titanium Surfaces by the Release of Antimicrobial β-Peptides from Polyelectrolyte Multilayers. Acta Biomater. 2019, 93, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, M.; Beito, B.; Oliveira, M.; Tudela Martins, M.; Gallas, B.; Salmain, M.; Boujday, S.; Humblot, V. Strategies for Antimicrobial Peptides Immobilization on Surfaces to Prevent Biofilm Growth on Biomedical Devices. Antibiotics 2022, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Ng, V.W.L.; Ke, X.; Lee, A.L.Z.; Hedrick, J.L.; Yang, Y.Y. Synergistic Co-Delivery of Membrane-Disrupting Polymers with Commercial Antibiotics against Highly Opportunistic Bacteria. Adv. Mater. 2013, 25, 6730–6736. [Google Scholar] [CrossRef]
- Leong, J.; Yang, C.; Tan, J.; Tan, B.Q.; Hor, S.; Hedrick, J.L.; Yang, Y.Y. Combination of Guanidinium and Quaternary Ammonium Polymers with Distinctive Antimicrobial Mechanisms Achieving a Synergistic Antimicrobial Effect. Biomater. Sci. 2020, 8, 6920–6929. [Google Scholar] [CrossRef]
- Si, Z.; Hou, Z.; Vikhe, Y.S.; Thappeta, K.R.V.; Marimuthu, K.; De, P.P.; Ng, O.T.; Li, P.; Zhu, Y.; Pethe, K.; et al. Antimicrobial Effect of a Novel Chitosan Derivative and Its Synergistic Effect with Antibiotics. ACS Appl. Mater. Interfaces 2021, 13, 3237–3245. [Google Scholar] [CrossRef]
- Moir, R.D.; Lathe, R.; Tanzi, R.E. The Antimicrobial Protection Hypothesis of Alzheimer’s Disease. Alzheimer’s Dement. 2018, 14, 1602–1614. [Google Scholar] [CrossRef]
- de Carrasco, L.D.M.; Sampaio, J.L.M.; Carmona-Ribeiro, A.M. Supramolecular Cationic Assemblies against Multidrug-Resistant Microorganisms: Activity and Mechanism of Action. Int. J. Mol. Sci. 2015, 16, 6337–6352. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Betancourt, Y.; de Távora, B.C.L.F.; Faquim-Mauro, E.L.; Carmona-Ribeiro, A.M. Biocompatible Lipid Polymer Cationic Nanoparticles for Antigen Presentation. Polymers 2021, 13, 185. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, E.; Morrison, I. Particle Size Distribution from Analysis of Quasi-Elastic Light Scattering Data. In Measurement of Suspended Particles by Quasi-Elastic Light Scattering; Dahneke, B., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 1983; pp. 199–236. [Google Scholar]
- Rahme, L.G.; Stevens, E.J.; Wolfort, S.F.; Shao, J.; Tompkins, R.G.; Ausubel, F.M. Common Virulence Factors for Bacterial Pathogenicity in Plants and Animals. Science 1995, 268, 1899–1902. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Baldini, R.L.; Déziel, E.; Saucier, M.; Zhang, Q.; Liberati, N.T.; Lee, D.; Urbach, J.; Goodman, H.M.; Rahme, L.G. The Broad Host Range Pathogen Pseudomonas Aeruginosa Strain PA14 Carries Two Pathogenicity Islands Harboring Plant and Animal Virulence Genes. Proc. Natl. Acad. Sci. USA 2004, 101, 2530–2535. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, S.-H.; Russell, D.H. An Experimental Study of the Solvent-Dependent Self-Assembly/Disassembly and Conformer Preferences of Gramicidin A. Anal. Chem. 2013, 85, 7826–7833. [Google Scholar] [CrossRef]
- Zhou, Y.; Franks, G.V. Flocculation Mechanism Induced by Cationic Polymers Investigated by Light Scattering. Langmuir ACS J. Surf. Colloids 2006, 22, 6775–6786. [Google Scholar] [CrossRef]
- Xavier, G.R.S.; Carmona-Ribeiro, A.M. Cationic Biomimetic Particles of Polystyrene/Cationic Bilayer/Gramicidin for Optimal Bactericidal Activity. Nanomaterials 2017, 7, 422. [Google Scholar] [CrossRef]
- Mathiazzi, B.I.; Carmona-Ribeiro, A.M. Hybrid Nanoparticles of Poly (Methyl Methacrylate) and Antimicrobial Quaternary Ammonium Surfactants. Pharmaceutics 2020, 12, 340. [Google Scholar] [CrossRef]
- Yala, J.-F.; Thebault, P.; Héquet, A.; Humblot, V.; Pradier, C.-M.; Berjeaud, J.-M. Elaboration of Antibiofilm Materials by Chemical Grafting of an Antimicrobial Peptide. Appl. Microbiol. Biotechnol. 2011, 89, 623–634. [Google Scholar] [CrossRef]
- Gregoriadis, G. Engineering Liposomes for Drug Delivery: Progress and Problems. Trends Biotechnol. 1995, 13, 527–537. [Google Scholar] [CrossRef]
- Jeswani, G.; Alexander, A.; Saraf, S.; Saraf, S.; Qureshi, A. Ajazuddin Recent Approaches for Reducing Hemolytic Activity of Chemotherapeutic Agents. J. Control. Release 2015, 211, 10–21. [Google Scholar] [CrossRef]
- Fischer, D.; Li, Y.; Ahlemeyer, B.; Krieglstein, J.; Kissel, T. In Vitro Cytotoxicity Testing of Polycations: Influence of Polymer Structure on Cell Viability and Hemolysis. Biomaterials 2003, 24, 1121–1131. [Google Scholar] [CrossRef]
- Milović, N.M.; Wang, J.; Lewis, K.; Klibanov, A.M. Immobilized N-Alkylated Polyethylenimine Avidly Kills Bacteria by Rupturing Cell Membranes with No Resistance Developed. Biotechnol. Bioeng. 2005, 90, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Hsu, B.B.; Ouyang, J.; Wong, S.Y.; Hammond, P.T.; Klibanov, A.M. On Structural Damage Incurred by Bacteria upon Exposure to Hydrophobic Polycationic Coatings. Biotechnol. Lett. 2011, 33, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Poon, Y.F.; Li, W.; Zhu, H.-Y.; Yeap, S.H.; Cao, Y.; Qi, X.; Zhou, C.; Lamrani, M.; Beuerman, R.W.; et al. A Polycationic Antimicrobial and Biocompatible Hydrogel with Microbe Membrane Suctioning Ability. Nat. Mater. 2011, 10, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Nizet, V. Antimicrobial Peptide Resistance Mechanisms of Human Bacterial Pathogens. Curr. Issues Mol. Biol. 2006, 8, 11–26. [Google Scholar] [CrossRef]
- Silva, G.A.; Czeisler, C.; Niece, K.L.; Beniash, E.; Harrington, D.A.; Kessler, J.A.; Stupp, S.I. Selective Differentiation of Neural Progenitor Cells by High-Epitope Density Nanofibers. Science 2004, 303, 1352–1355. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Photo | Dz/nm | P | ζ/mV | G/µS | Turbidity/400 nm |
|---|---|---|---|---|---|---|
| 0.25 mg/mL PDDA in water |  | - | - | - | 245 ± 10 | 0 |
| 0.1 mM Gr/TFE/water |  | 159 ± 1 | 0.14 ± 0.01 | -26 ± 3 | 4 ± 2 | 0.43 ± 0.01 |
| 0.25 mg/mL PDDA/TFE/water |  | - | - | - | 246 ± 11 | 0 |
| 0.1 mM Gr/0.25 mg/mL PDDA/TFE/water |  | 426 ± 31 | 0.24 ± 0.04 | 49 ± 1 | 236 ± 6 | 0.64 ± 0.01 |
| Cells | [Gr]/µM | [PDDA]/µg·mL−1 | [Gr](µM)/[PDDA](µg·mL−1) |
|---|---|---|---|
| Macrophages/104 cells | * 10 (0.05) | ||
| Fibroblasts/104 cells | * 10 (0.05) | ||
| E. coli/108 CFU | ** 5 (0.3) | *** 5 (8) | |
| P. aeruginosa/107 CFU | 3 (7) | 0.5/0.5 (7) | |
| P. aeruginosa MDR/108 CFU | **** 2 (8) | ||
| C. albicans/107 CFU | 2 (7) | *** 5 (5) | 0.5/0.5 (7) |
| C. albicans fluconazol R/105 CFU | **** 1 (5) | ||
| S. aureus | 20 (7) (from 107 to 1 CFU) | *** 10 (6) (from 108 to 102 CFU) | 10/10 (6) (from 106 to 1 CFU) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Betancourt, Y.; Zaia, R.; Evangelista, M.F.; Ribeiro, R.T.; Roncoleta, B.M.; Mathiazzi, B.I.; Carmona-Ribeiro, A.M. Characterization and Differential Cytotoxicity of Gramicidin Nanoparticles Combined with Cationic Polymer or Lipid Bilayer. Pharmaceutics 2022, 14, 2053. https://doi.org/10.3390/pharmaceutics14102053
Pérez-Betancourt Y, Zaia R, Evangelista MF, Ribeiro RT, Roncoleta BM, Mathiazzi BI, Carmona-Ribeiro AM. Characterization and Differential Cytotoxicity of Gramicidin Nanoparticles Combined with Cationic Polymer or Lipid Bilayer. Pharmaceutics. 2022; 14(10):2053. https://doi.org/10.3390/pharmaceutics14102053
Chicago/Turabian StylePérez-Betancourt, Yunys, Rachel Zaia, Marina Franchi Evangelista, Rodrigo Tadeu Ribeiro, Bruno Murillo Roncoleta, Beatriz Ideriha Mathiazzi, and Ana Maria Carmona-Ribeiro. 2022. "Characterization and Differential Cytotoxicity of Gramicidin Nanoparticles Combined with Cationic Polymer or Lipid Bilayer" Pharmaceutics 14, no. 10: 2053. https://doi.org/10.3390/pharmaceutics14102053