
The Impact of Fullerenes as Doxorubicin Nano-Transporters on Metallothionein and Superoxide Dismutase Status in MCF-10A Cells

 and
and
Abstract
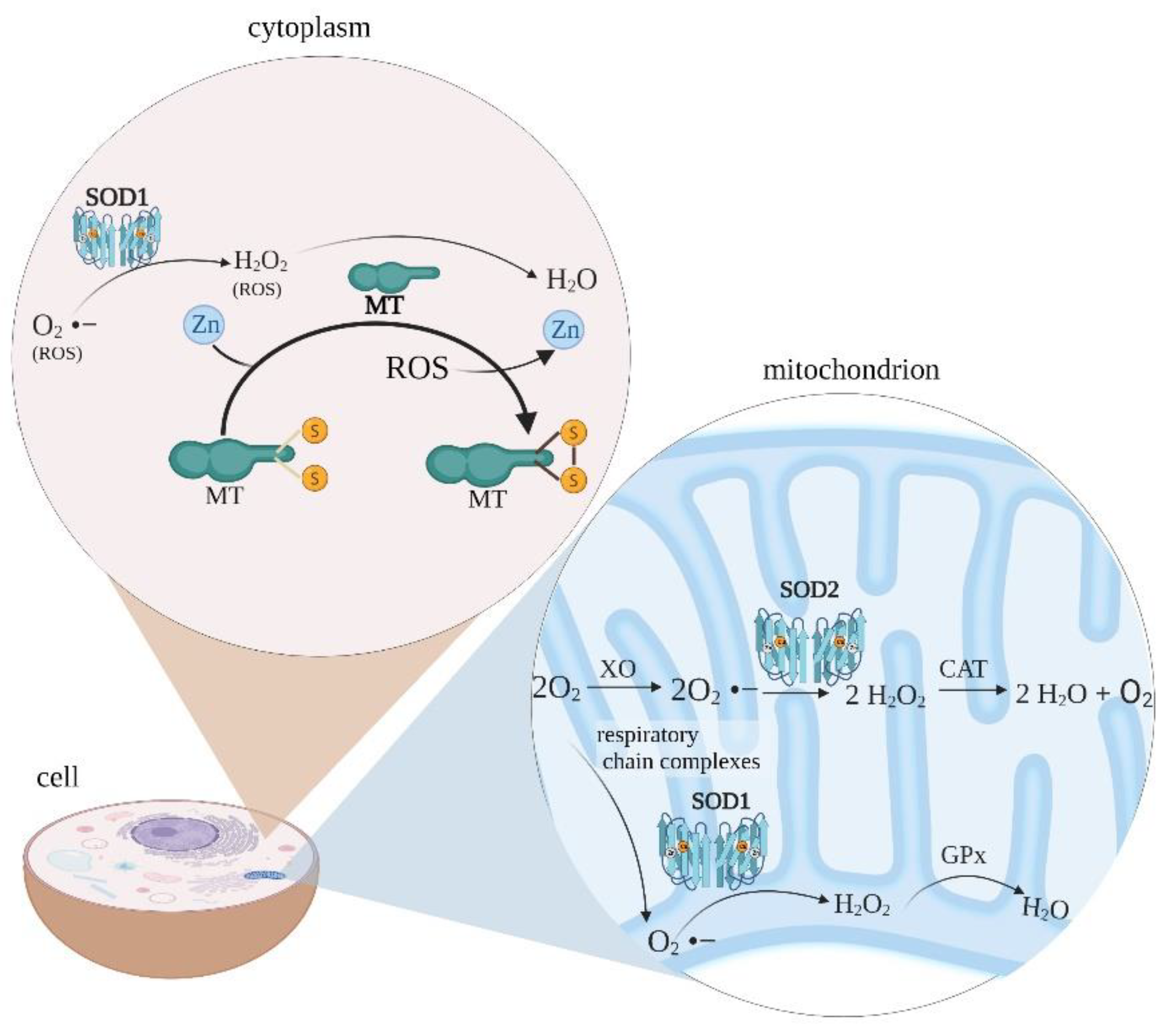
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Preparation of Fullerene–Doxorubicin Complexes
2.2.2. Size and Zeta Potential Measurements
2.2.3. Fluorescence Measurements of DOX
2.2.4. Culture and Preparation of MCF-10A Cells
2.2.5. Cell Lysis
2.2.6. Total Protein Concentration Measurement by the Bradford Method
2.2.7. Metallothionein Concentration
2.2.8. Concentration of SOD1 and Total SOD Activity
2.2.9. Statistical Analysis
3. Results
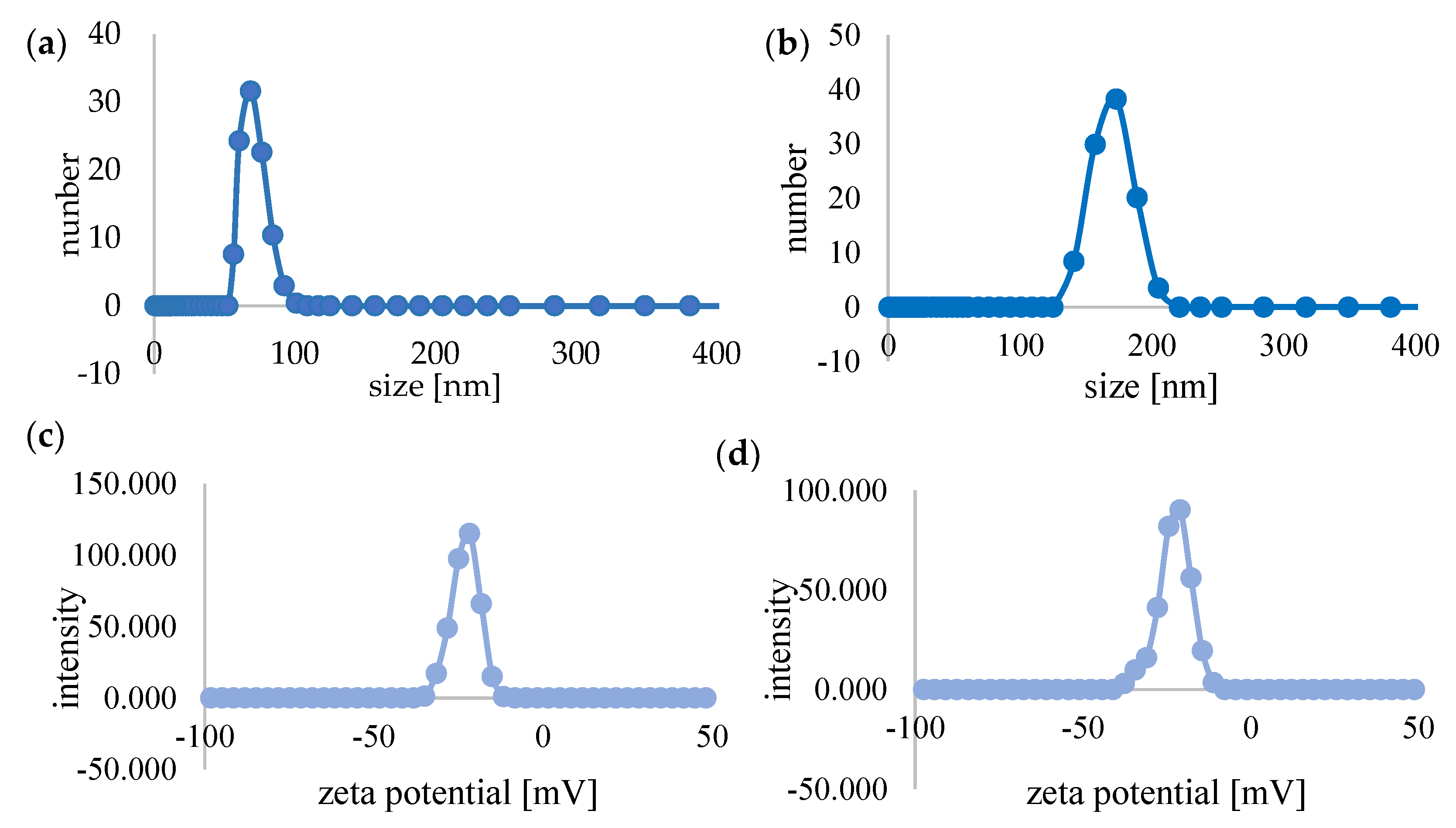
3.1. Biophysical Characteristics of C60–DOX Complexes
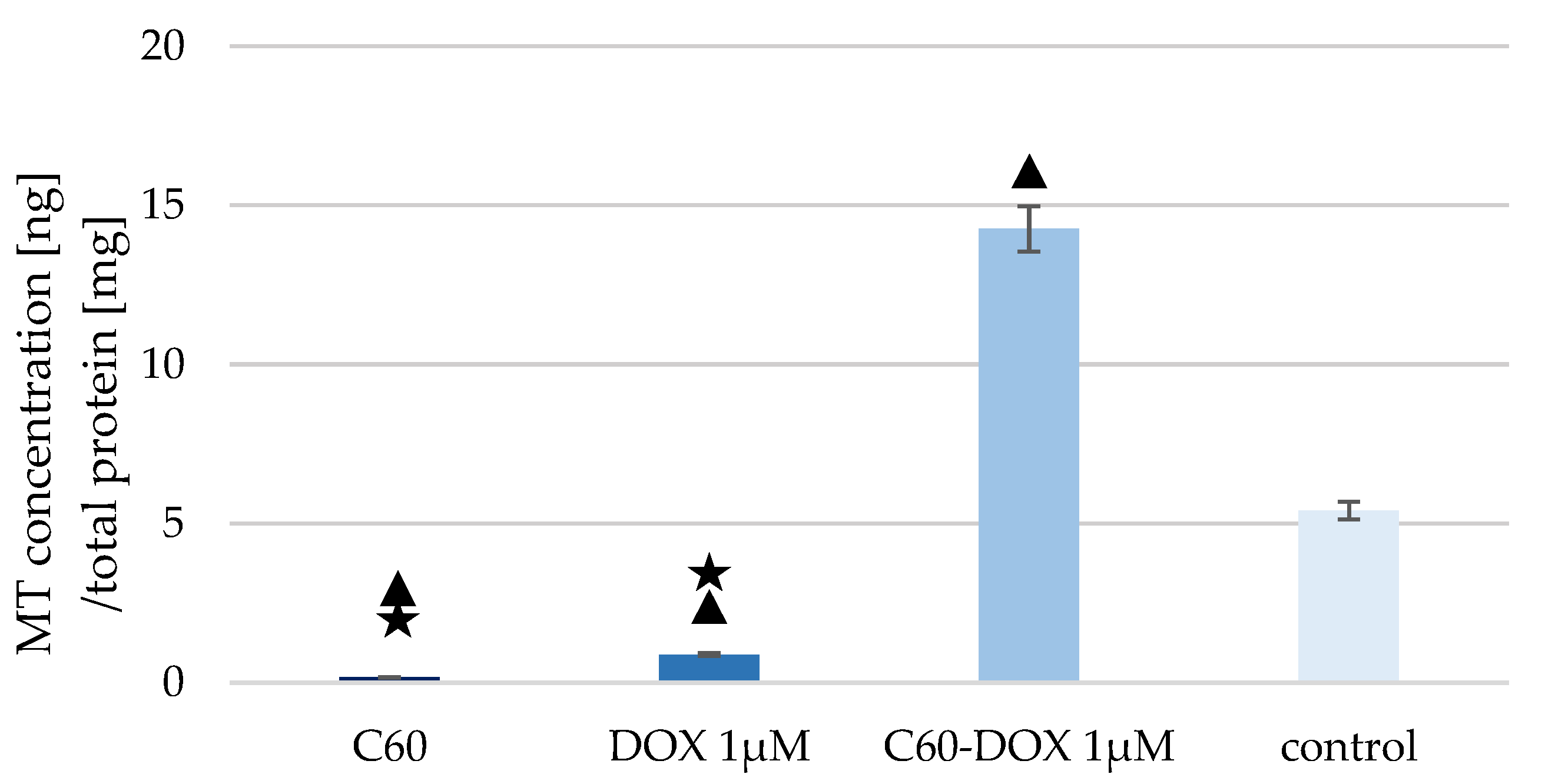
3.2. The Effect of C60–DOX Complexes on Metallothionein Concentration in MCF-10A Cells
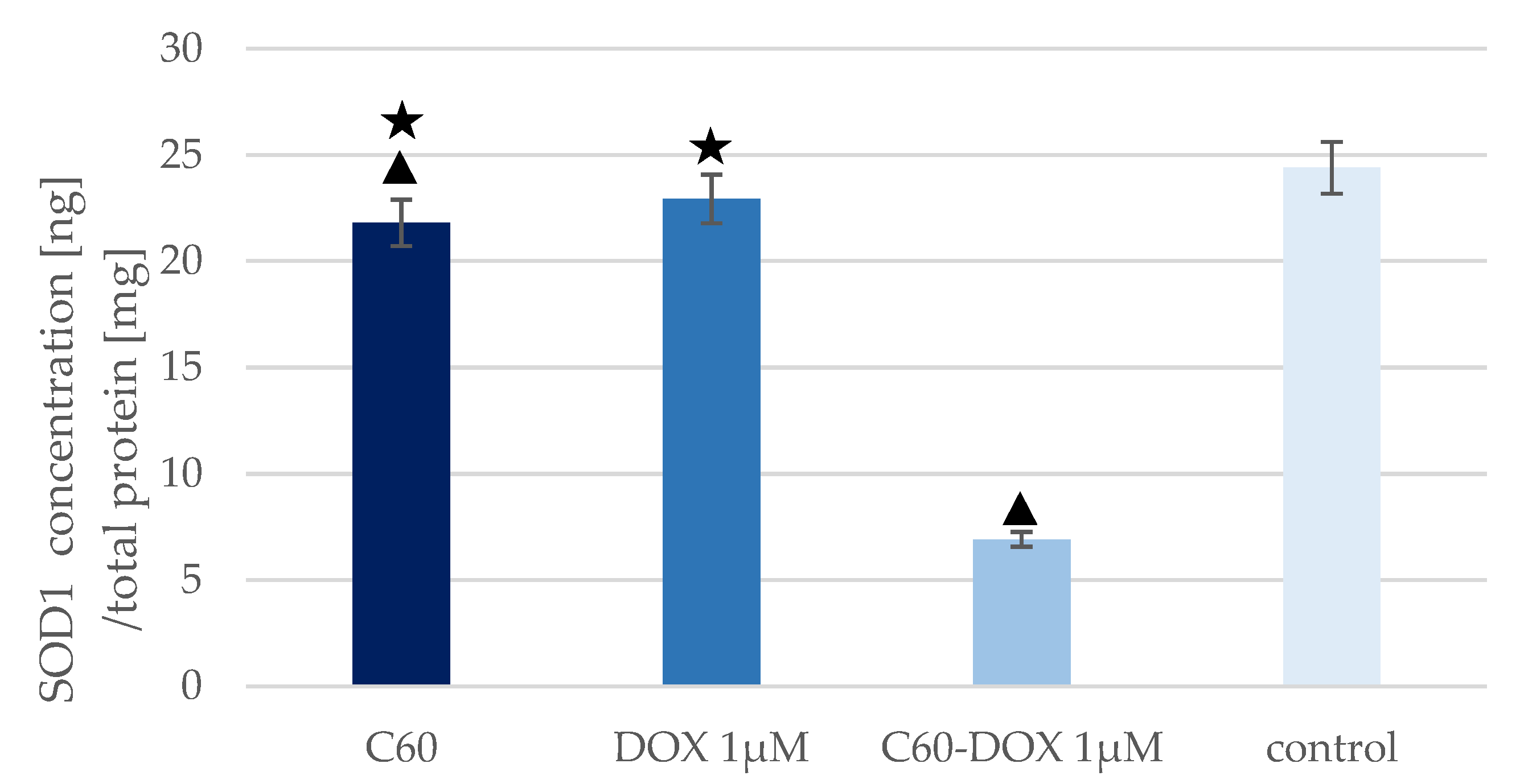
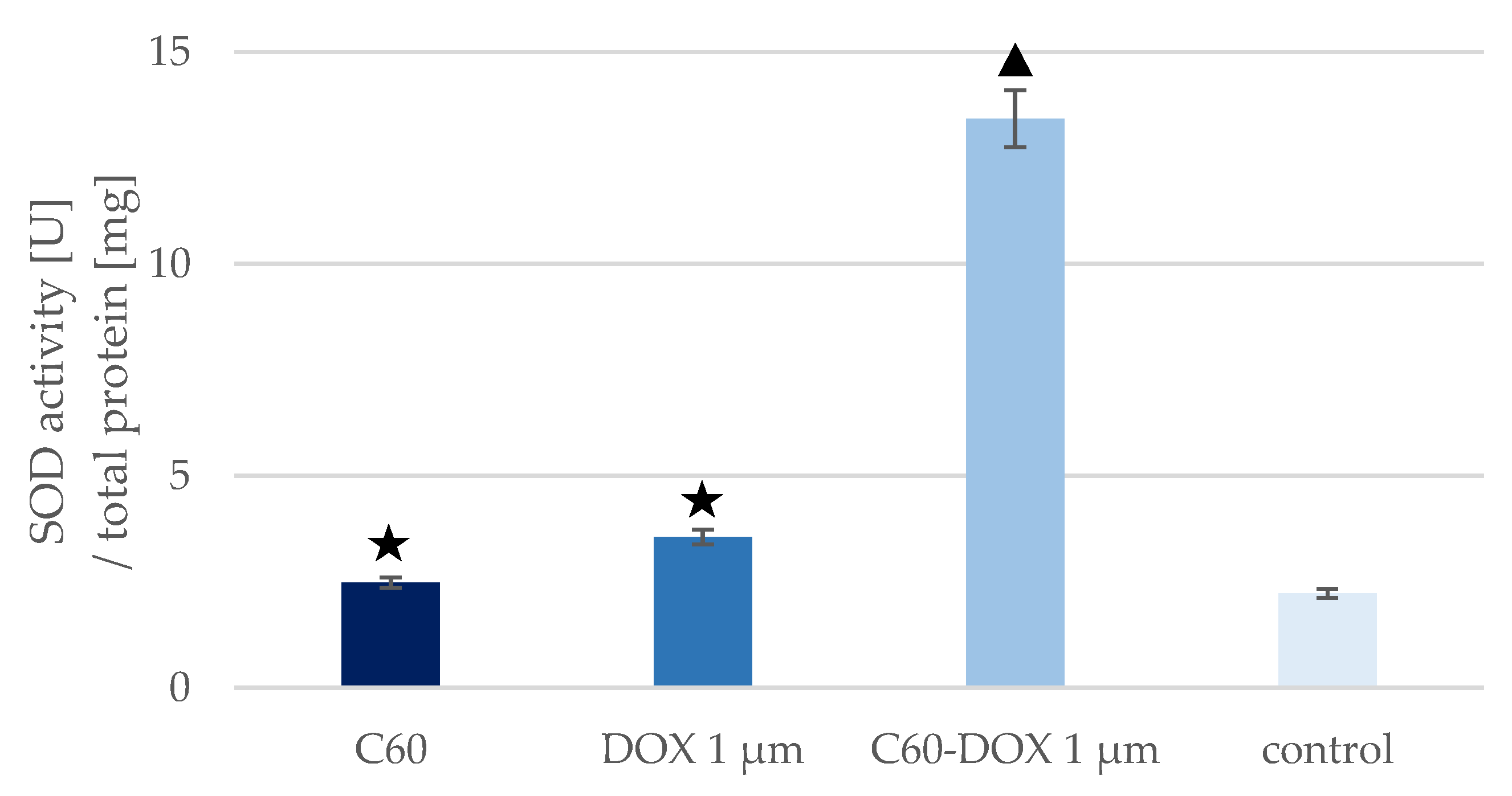
3.3. The Effect of C60–DOX Complexes on SOD Concentration and Activity in MCF-10A Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGowan, J.V.; Chung, R.; Maulik, A.; Piotrowska, I.; Walker, J.M.; Yellon, D.M. Anthracycline Chemotherapy and Cardiotoxicity. Cardiovasc. Drugs Ther. 2017, 31, 63–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korbut, R. Farmakologia; PZWL Wydawnictwo Lekarskie: Warszawa, Poland, 2017; pp. 240–248. [Google Scholar]
- Pilco-Ferreto, N.; Calaf, G.M. Influence of doxorubicin on apoptosis and oxidative stress in breast cancer cell lines. Int. J. Oncol. 2016, 49, 753–762. [Google Scholar] [CrossRef] [Green Version]
- International Agency for Research on Cancer. Available online: https://www.iarc.who.int/ (accessed on 23 September 2021).
- Van der Zanden, S.Y.; Qiao, X.; Neefjes, J. New insights into the activities and toxicities of the old anticancer drug doxorubicin. FEBS J. 2020, 288, 6095–6111. [Google Scholar] [CrossRef] [PubMed]
- Tacar, O.; Sriamornsak, P.; Dass, C.R. Doxorubicin: An update on anticancer molecular action, toxicity and novel drug delivery systems. J. Pharm. Pharmacol. 2013, 65, 157–170. [Google Scholar] [CrossRef]
- Kudryavtseva, A.V.; Krasnov, G.S.; Dmitriev, A.; Alekseev, B.Y.; Kardymon, O.; Sadritdinova, A.F.; Fedorova, M.S.; Pokrovsky, A.V.; Melnikova, N.V.; Kaprin, A.D.; et al. Mitochondrial dysfunction and oxidative stress in aging and cancer. Oncotarget 2016, 7, 44879–44905. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Villani, R.M.; Wang, H.; Simpson, M.J.; Roberts, MS.; Tang, M.; Liang, X. The role of cellular reactive oxygen species in cancer chemotherapy. J. Exp. Clin. Cancer Res. 2018, 37, 266. [Google Scholar] [CrossRef] [PubMed]
- Borgstahl, G.E.O.; Oberley-Deegan, R.E. Superoxide dismutases (SODs) and SOD mimetics. Antioxidants. 2018, 7, 156. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, F.; Ramnath, N.; Nagrath, D. Reactive oxygen species in the tumor. Cancers 2019, 11, 1191. [Google Scholar] [CrossRef] [Green Version]
- Case, A.J. On the origin of superoxide dismutase: An evolutionary perspective of superoxide-mediated redox signaling. Antioxidants 2017, 6, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gałecka, E.; Jacewicz, R.; Mrowicka, M.; Gałecki, P. Antioxidative enzymes-structure, properties, functions. Pol. Merkur. Lek. 2008, 25, 266–268. [Google Scholar]
- Zaręba, N.; Kepinska, M. The Function of Transthyretin Complexes with Metallothionein in Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 9003. [Google Scholar] [CrossRef]
- Ostrakhovitch, E.A.; Olsson, P.E.; Jiang, S.; Cherian, M.G. Interaction of metallothionein with tumor suppressor p53 protein. FEBS Lett. 2006, 580, 1235–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoda, R.; Achanzar, W.E.; Qu, W.; Nagamine, T.; Takagi, H.; Mori, M.; Waalkes, M.P. Metallothionein is a potential negative regulator of apoptosis. Toxicol Sci. 2003, 73, 294–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, M.; Lang, J. The roles of metallothioneins in carcinogenesis. J. Hematol. Oncol. 2018, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. Alterations in Concentration/Activity of Superoxide Dismutases in Context of Obesity and Selected Single Nucleotide Polymorphisms in Genes: SOD1, SOD2, SOD3. Int. J. Mol. Sci. 2020, 21, 5069. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. Inhibition of copper-zinc superoxide dismutase activity by selected environmental xenobiotics. Environ. Toxicol. Pharmacol. 2018, 58, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Papa, L.; Hahn, M.; Marsh, E.L.; Evans, B.S.; Germain, D. SOD2 to SOD1 switch in breast cancer. J. Biol. Chem. 2014, 289, 5412–5416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papa, L.; Manfredi, G.; Germain, D. SOD1, an unexpected novel target for cancer therapy. Genes Cancer 2018, 5, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Songbo, M.; Lang, H.; Xinyong, C.; Bin, X.; Ping, Z.; Liang, S. Oxidative stress injury in doxorubicin-induced cardiotoxicity. Toxicol. Lett. 2019, 307, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Gomari, H.; Moghadam, M.F.; Soleimani, M.; Ghavami, M.; Khodashenas, S. Targeted delivery of doxorubicin to HER2 positive tumor models. Int. J. Nanomed. 2019, 14, 5679–5690. [Google Scholar] [CrossRef] [Green Version]
- Rašović, I. Water-soluble fullerenes for medical applications. Mater. Sci. Technol. 2016, 33, 777–794. [Google Scholar] [CrossRef]
- Goodarzi, S.; da Ros, T.; Conde, J.; Sefat, F.; Mozafari, M. Fullerene: Biomedical engineers get to revisit an old friend. Mater. Today 2017, 20, 460–480. [Google Scholar] [CrossRef] [Green Version]
- Castro, E.; Hernandez Garcia, A.; Zavala, G.; Echegoyen, L. Fullerenes in Biology and Medicine. J. Mater. Chem. B 2017, 5, 6523–6535. [Google Scholar] [CrossRef] [PubMed]
- Kerna, A.; Pruitt, K.; Flores, J.; Nwokorie, U. The application of fullerene derivatives in medicine and specific endocrinological conditions. EC Endocrinol. Metab. Res. 2020, 5, 56–66. [Google Scholar]
- Mirkamali, E.S.; Ahmadi, R.; Kalateh, K.; Zarei, G. Adsorption of melphalan anticancer drug on the surface of fullerene (C24): A comprehensive DFT study. Nanomed. J. 2019, 6, 112–119. [Google Scholar]
- Kazemzadeh, H.; Mozafari, M. Fullerene-based delivery systems. Drug Discov. Today 2019, 24, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Misra, C.; Kumar, M.; Sharma, G.; Kumar, R.; Singh, B.; Katare, O.P.; Raza, K. Glycinated fullerenes for tamoxifen intracellular delivery with improved anticancer activity and pharmacokinetics. Nanomedicine 2017, 12, 1011–1023. [Google Scholar] [CrossRef]
- Lichota, A.; Krokosz, A. Fullerenols in therapy and diagnosis of cancer. Med. Pr. 2016, 67, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Kepinska, M.; Kizek, R.; Milnerowicz, H. Fullerene as a doxorubicin nanotransporter for targeted breast cancer therapy: Capillary electrophoresis analysis. Electrophoresis 2018, 39, 2370–2379. [Google Scholar] [CrossRef]
- Milnerowicz, H.; Bizoń, A. Determination of metallothionein in biological fluids using enzyme-linked immunoassay with commercial antibody. Acta Bioch. Pol. 2010, 57, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Buzdar, A.U. Preoperative chemotherapy treatment of breast cancer-A review. Cancer 2007, 110, 2394–2407. [Google Scholar] [CrossRef] [PubMed]
- Mahalingaiah, P.K.S.; Singh, K.P. Chronic oxidative stress increases growth and tumorigenic potential of MCF-7 breast cancer cells. PLoS ONE 2014, 9, e87371. [Google Scholar]
- Prylutska, S.; Grynyuk, I.; Matyshevska, O.; Prylutskyy, Y.; Evstigneev, M.; Scharff, P.; Ritter, U. C60 Fullerene as Synergistic Agent in Tumor-Inhibitory Doxorubicin Treatment. Drugs R D 2014, 14, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Prylutskyy, Y.I.; Evstigneev, M.P.; Pashkova, I.S.; Wyrzykowski, D. Characterization of C 60 fullerene complexation with antibiotic doxorubicin. Phys. Chem. Chem. Phys. 2014, 16, 23164–23172. [Google Scholar] [CrossRef]
- Brant, J. Aggregation and deposition characteristics of fullerene nanoparticles in aqueous systems. J. Nanoparticle Res. 2005, 7, 545–553. [Google Scholar] [CrossRef]
- Alargova, R.G.; Deguchi, S.; Tsujii, K. Stable colloidal dispersions of fullerenes in polar organic solvents. J. Am. Chem. Soc. 2001, 123, 10460–10467. [Google Scholar] [CrossRef]
- Kumar, A.; Dixit, C.K. Methods for Characterization of Nanoparticles. In Advances in Nanomedicine for the Delivery of Therapeutic Nucleic Acids; Woodhead Publishing: Cambridge, UK, 2017. [Google Scholar]
- Joseph, E.; Singhvi, G. Multifunctional Nanocrystals for Cancer Therapy: A Potential Nanocarrier. In Nanomaterials for Drug Delivery and Therapy; William Andrew Publishing: Norwich, NY, USA, 2019; pp. 91–116. [Google Scholar]
- Mchedlov-Petrossyan, N.O.; Klochkov, V.K.; Andrievsky, G.V. Colloidal dispersions of fullerene C60 in water: Some properties and regularities of coagulation by electrolytes. J. Chem. Soc. Faraday Trans. 1997, 93, 4343–4346. [Google Scholar] [CrossRef]
- Kepinska, M.; Kizek, R.; Milnerowicz, H. Metallothionein and superoxide dismutase—Antioxidative protein status in fullerene-doxorubicin delivery to MCF-7 human breast cancer cells. Int. J. Mol. Sci. 2018, 19, 3253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilghami, R.; Barzegari, A.; Mashayekhi, M.R.; Letourneur, D.; Crepin, M.; Pavon-Djavid, G. The conundrum of dietary antioxidants in cancer chemotherapy. Nutr. Rev. 2020, 78, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Gajewski, E.; Gaur, S.; Akman, S.A.; Matsumoto, L.; Balgooy, J.N.A.; Doroshow, J.H. Oxidative DNA Base Damage in MCF-10A Breast Epithelial Cells at Clinically Achievable Concentrations of Doxorubicin. Biochem. Pharmacol. 2007, 73, 1947. [Google Scholar] [CrossRef] [Green Version]
- Aminipour, Z.; Khorshid, M.; Keshvari, H.; Bonakdar, S.; Wagner, P.; Van der Bruggen, B. Passive permeability assay of doxorubicin through model cell membranes under cancerous and normal membrane potential conditions. Eur. J. Pharm. Biopharm. 2020, 146, 133–142. [Google Scholar] [CrossRef]
- Grebinyk, A.; Prylutska, S.; Chepurna, O.; Grebinyk, S.; Prylutskyy, Y.; Ritter, U.; Ohulchanskyy, T.Y.; Matyshevska, O.; Dandekar, T.; Frohme, M. Synergy of Chemo- and Photodynamic Therapies with C60 Fullerene-Doxorubicin Nanocomplex. Nanomaterials 2019, 9, 1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, L.; Li, L.; Zhao, J.; Sun, Z.; Peng, S. Zinc-induced metallothionein overexpression prevents doxorubicin toxicity in cardiomyocytes by regulating the peroxiredoxins. Xenobiotica 2016, 46, 715–725. [Google Scholar] [CrossRef]
- Merlos Rodrigo, M.A.; Jimenez Jimemez, A.M.; Haddad, Y.; Bodoor, K.; Adam, P.; Krizkova, S.; Heger, Z.; Adam, V. Metallothionein isoforms as double agents-Their roles in carcinogenesis, cancer progression and chemoresistance. Drug Resist Updat. 2020, 52, 100691. [Google Scholar] [CrossRef]
- Srdjenovic, B.; Milic-Torres, V.; Grujic, N.; Stankov, K.; Djordjevic, A.; Vasovic, V. Antioxidant properties of fullerenol C60(OH)24 in rat kidneys, testes, and lungs treated with doxorubicin. Toxicol. Mech. Methods 2010, 20, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.C.; Lai, H.C.; Ting, C.T.; Lee, W.L.; Wang, L.C.; Wang, K.Y.; Lai, H.C.; Liu, T.J. Protection by doxycycline against doxorubicin-induced oxidative stress and apoptosis in mouse testes. Biochem. Pharmacol. 2007, 74, 969–980. [Google Scholar] [CrossRef]
- Ali, S.S.; Hardt, J.I.; Quick, K.L.; Sook Kim-Han, J.; Erlanger, B.F.; Huang, T.T.; Epstein, C.J.; Dugan, L.L. A biologically effective fullerene (C 60) derivative with superoxide dismutase mimetic properties. Free Radic. Biol. Med. 2004, 37, 1191–1202. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | SOD1 Concentration [ng/mg Total Protein] | SOD Activity [U/mg Total Protein] | SOD Activity/SOD1 Concentration [U/ng] |
|---|---|---|---|
| Control | 24.402 | 2.226 | 0.091 |
| C60 | 21.807 | 2.483 | 0.114 |
| DOX | 22.931 | 3.557 | 0.155 |
| C60–DOX | 6.913 | 13.424 | 1.942 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaręba, N.; Więcławik, K.; Kizek, R.; Hosnedlova, B.; Kepinska, M. The Impact of Fullerenes as Doxorubicin Nano-Transporters on Metallothionein and Superoxide Dismutase Status in MCF-10A Cells. Pharmaceutics 2022, 14, 102. https://doi.org/10.3390/pharmaceutics14010102
Zaręba N, Więcławik K, Kizek R, Hosnedlova B, Kepinska M. The Impact of Fullerenes as Doxorubicin Nano-Transporters on Metallothionein and Superoxide Dismutase Status in MCF-10A Cells. Pharmaceutics. 2022; 14(1):102. https://doi.org/10.3390/pharmaceutics14010102
Chicago/Turabian StyleZaręba, Natalia, Klaudia Więcławik, Rene Kizek, Bozena Hosnedlova, and Marta Kepinska. 2022. "The Impact of Fullerenes as Doxorubicin Nano-Transporters on Metallothionein and Superoxide Dismutase Status in MCF-10A Cells" Pharmaceutics 14, no. 1: 102. https://doi.org/10.3390/pharmaceutics14010102