
Purification and Identification of Novel Antioxidant Peptides Isolated from Geoffroea decorticans Seeds with Anticoagulant Activity


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Chemicals
2.1.2. Sample Collection
2.2. Crude Extract Preparation and Determination of the Protein Content
2.3. Purification of Antioxidant Peptides
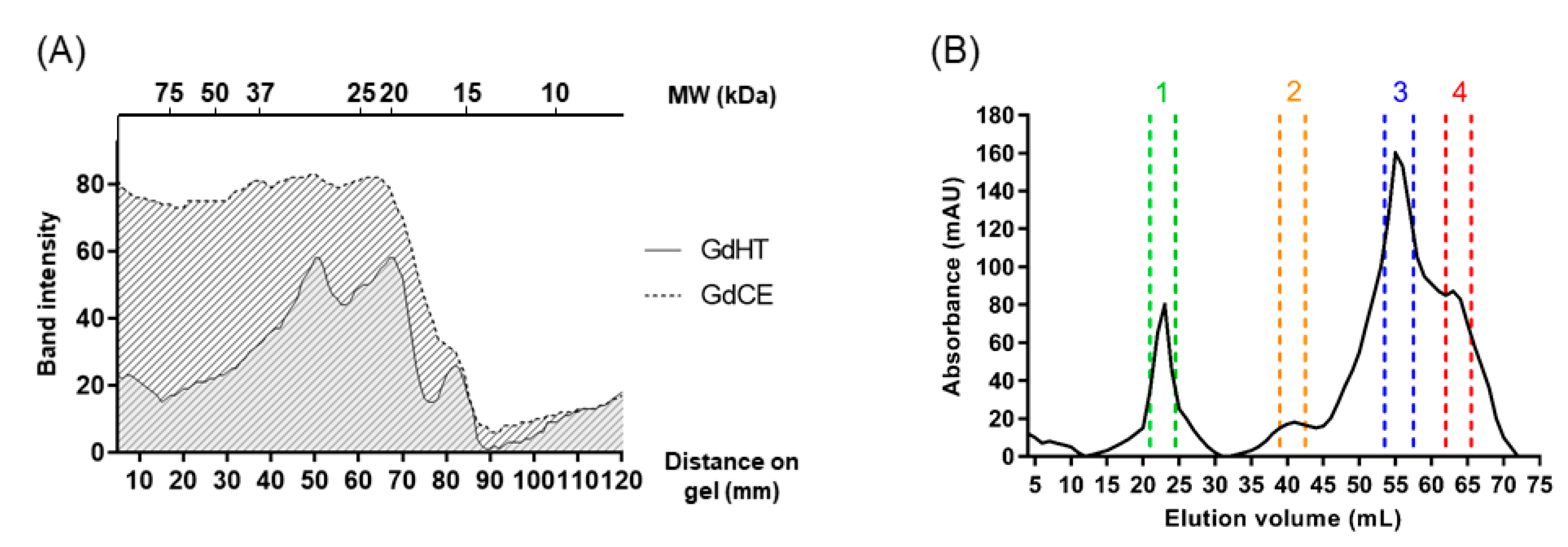
2.3.1. Initial Purification by Heat Treatment
2.3.2. Size-Exclusion Chromatography
2.4. MALDI-TOF Mass Spectrometric Analysis
2.5. Antioxidant Activity Evaluation
2.5.1. DPPH Radical-Scavenging Activity
2.5.2. ABTS Radical-Scavenging Activity
2.6. Anticoagulant Activity
2.7. Statistical Analysis
3. Results and Discussion
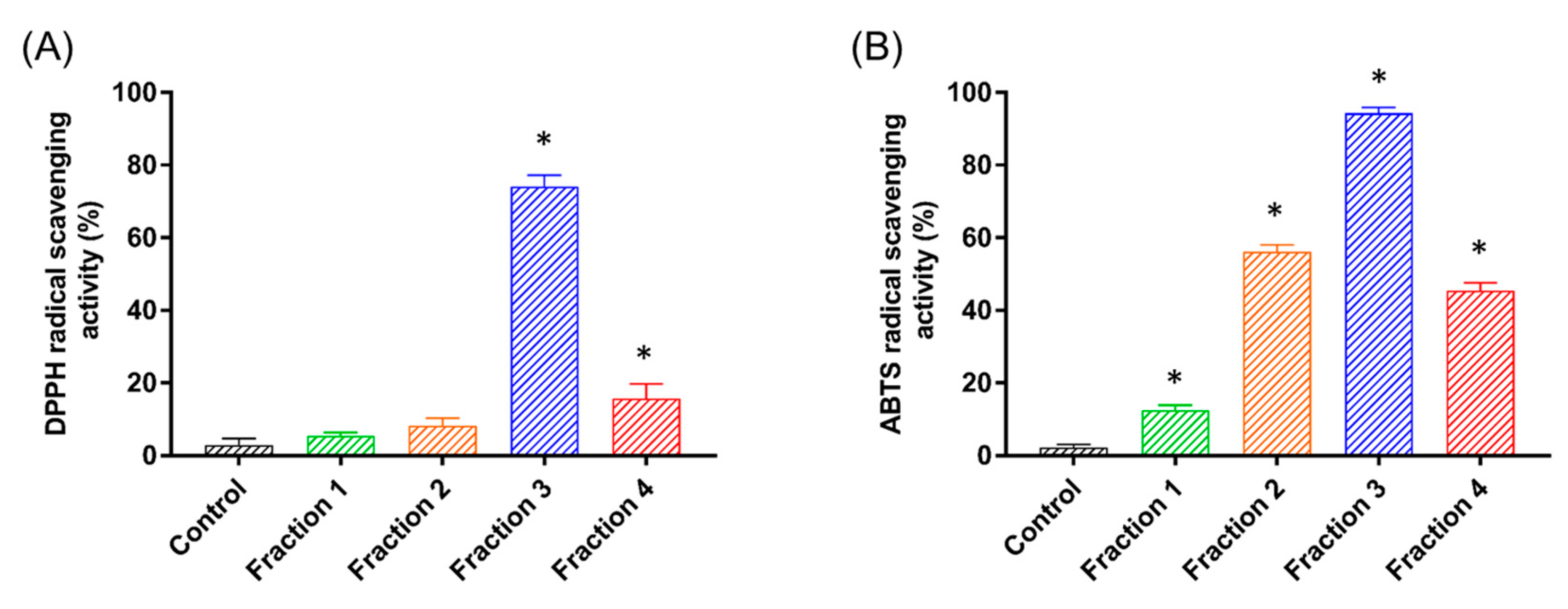
3.1. Isolation and Purification of the Antioxidant Peptides
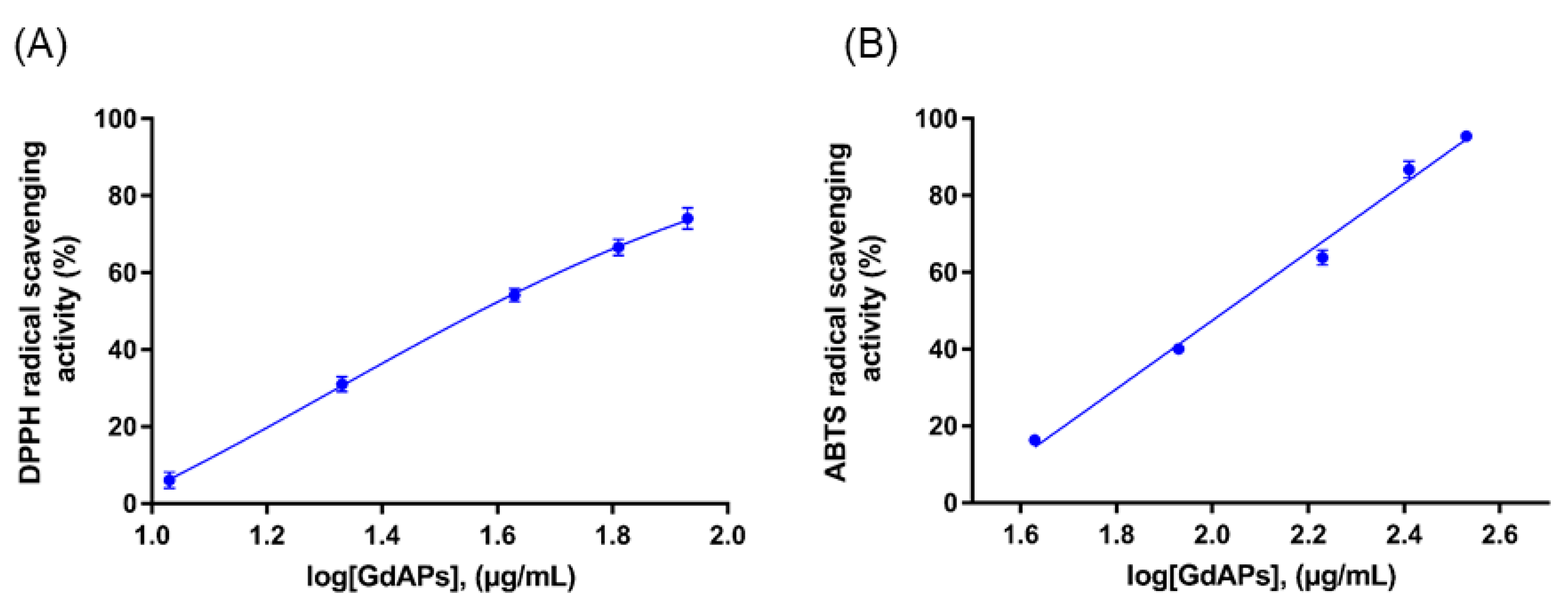
3.2. Identification and Antioxidant Properties of GdAPs
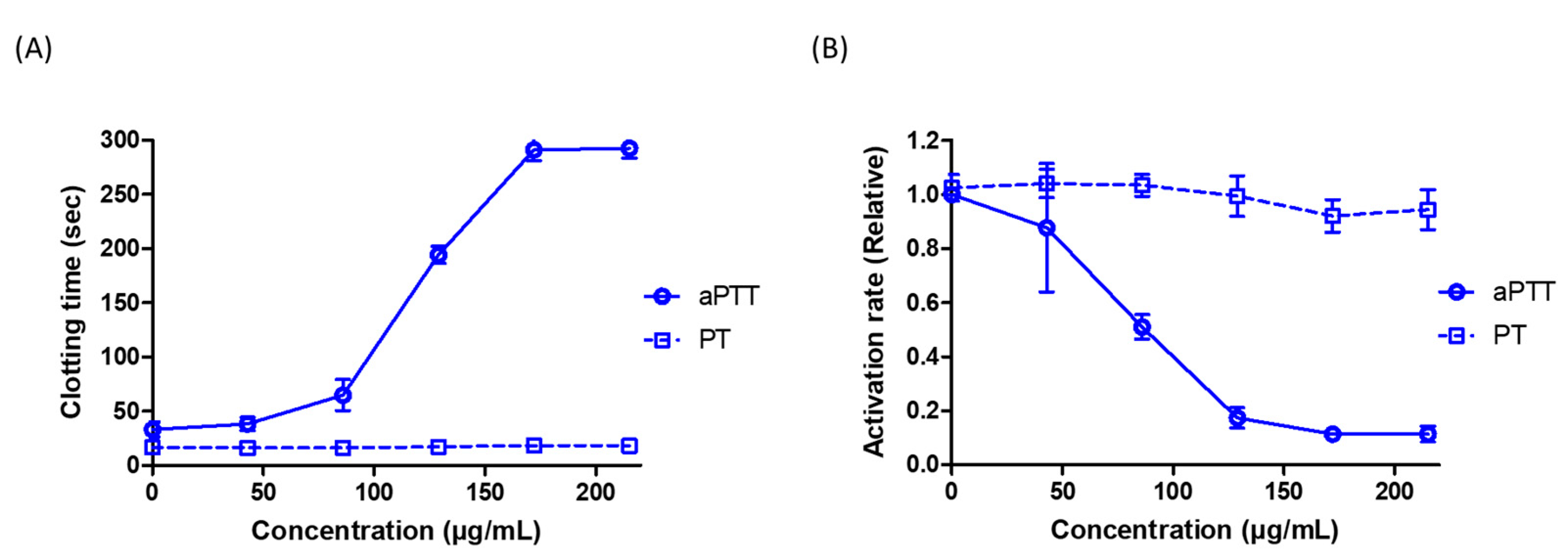
3.3. GdAPs Display Strong Anticoagulant Activiy against the Intrinsic Pathway
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dias, D.A.; Urban, S.; Roessner, U. A Historical Overview of Natural Products in Drug Discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [Green Version]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural Products in Drug Discovery: Advances and Opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Lautié, E.; Russo, O.; Ducrot, P.; Boutin, J.A. Unraveling Plant Natural Chemical Diversity for Drug Discovery Purposes. Front. Pharmacol. 2020, 11, 1–37. [Google Scholar] [CrossRef]
- Cotabarren, J.; Lufrano, D.; Parisi, M.G.; Obregón, W.D. Biotechnological, Biomedical, and Agronomical Applications of Plant Protease Inhibitors with High Stability: A Systematic Review. Plant Sci. 2020, 292, 110398. [Google Scholar] [CrossRef]
- Cotabarren, J.; Claver, S.; Payrol, J.A.; Garcia-pardo, J.; Obregón, W.D. Purification and Characterization of a Novel Thermostable Pa-Pain Inhibitor from Moringa Oleifera with Antimicrobial and an-Ticoagulant Properties. Pharmaceutics 2021, 1–12. [Google Scholar]
- Palta, S.; Saroa, R.; Palta, A. Overview of the Coagulation System. Indian J. Anaesth. 2014, 58, 515–523. [Google Scholar] [CrossRef]
- Chapin, J.C.; Hajjar, K.A. Fibrinolysis and the Control of Blood Coagulation John NIH Public Access. Blood Rev. 2015, 29, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furie, B.; Furie, B.C. Mechanisms of Thrombus Formation. Mechanisms of Disease. N. Engl. J. Med. 2008, 359, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.P.; Darbousset, R.; Schoenwaelder, S.M. Thromboinflammation: Challenges of Therapeutically Targeting Coagulation and Other Host Defense Mechanisms. Blood 2019, 133, 906–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Executive Summary: Report of the 22nd WHO Expert Committee on Selection and Use of Essential Medicines. WHO Med. Web 2019, No. April, 1–15. Available online: https://apps.who.int/iris/handle/10665/325773 (accessed on 10 June 2021).
- Franchini, M.; Marano, G.; Cruciani, M.; Mengoli, C.; Pati, I.; Masiello, F.; Veropalumbo, E.; Pupella, S.; Vaglio, S.; Liumbruno, G.M. COVID-19-Associated Coagulopathy. Diagnosis 2020, 7, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Bang, O.Y.; Chung, J.-W.; Lee, M.J.; Seo, W.-K.; Kim, G.-M.; Ahn, M.-J.; Investigators, O.-C.S. Cancer-Related Stroke: An Emerging Subtype of Ischemic Stroke with Unique Pathomechanisms. J. Stroke 2020, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Adewole, K.E.; Adebayo, J.O. Antioxidant Defense System Induced by Cysteine-Stabilized Peptide Fraction of Aqueous Extract of Morinda Lucida Leaf in Selected Tissues of Plasmodium Berghei-Infected Mice. J. Integr. Med. 2017, 15, 388–397. [Google Scholar] [CrossRef]
- Herrera Chalé, F.G.; Ruiz Ruiz, J.C.; Acevedo Fernández, J.J.; Betancur Ancona, D.A.; Segura Campos, M.R. ACE Inhibitory, Hypotensive and Antioxidant Peptide Fractions from Mucuna Pruriens Proteins. Process Biochem. 2014, 49, 1691–1698. [Google Scholar] [CrossRef]
- Indiano-Romacho, P.; Fernández-Tomé, S.; Amigo, L.; Hernández-Ledesma, B. Multifunctionality of Lunasin and Peptides Released during Its Simulated Gastrointestinal Digestion. Food Res. Int. 2019, 125, 108513. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Tang, X.; Ren, Y.; Wang, E.; Shi, L.; Wu, X.; Wu, H. Novel Antioxidant Peptides Purified from Mulberry (Morus Atropurpurea Roxb.) Leaf Protein Hydrolysates with Hemolysis Inhibition Ability and Cellular Antioxidant Activity. J. Agric. Food Chem. 2019, 67, 7650–7659. [Google Scholar] [CrossRef]
- Bilska, A.; Wlodek, L. Lipoic Acid-the Drug of the Future? Pharmacol. Rep. 2005, 57, 570–577. [Google Scholar]
- Butterfield, D.A.; Abdul, H.M.; Opii, W.; Newman, S.F.; Joshi, G.; Ansari, M.A.; Sultana, R. Pin1 in Alzheimer’s Disease. J. Neurochem. 2006, 98, 1697–1706. [Google Scholar] [CrossRef]
- Ngo, D.-H.; Wijesekara, I.; Vo, T.-S.; Van Ta, Q.; Kim, S.-K. Marine Food-Derived Functional Ingredients as Potential Antioxidants in the Food Industry: An Overview. Food Res. Int. 2011, 44, 523–529. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Victor, V.M.; Rocha, M. Targeting Antioxidants to Mitochondria: A Potential New Therapeutic Strategy for Cardiovascular Diseases. Curr. Pharm. Des. 2007, 13, 845–863. [Google Scholar] [CrossRef] [PubMed]
- Sivonova, M.; Tatarkova, Z.; Durackova, Z.; Dobrota, D.; Lehotsky, J.; Matakova, T.; Kaplan, P. Relationship between Antioxidant Potential and Oxidative Damage to Lipids, Proteins and DNA in Aged Rats. Physiol. Res. 2007, 56, 757–764. [Google Scholar] [PubMed]
- Diaz, M.; Dunn, C.M.; McClements, D.J.; Decker, E.A. Use of Caseinophosphopeptides as Natural Antioxidants in Oil-in-Water Emulsions. J. Agric. Food Chem. 2003, 51, 2365–2370. [Google Scholar] [CrossRef]
- Ndiaye, F.; Vuong, T.; Duarte, J.; Aluko, R.E.; Matar, C. Anti-Oxidant, Anti-Inflammatory and Immunomodulating Properties of an Enzymatic Protein Hydrolysate from Yellow Field Pea Seeds. Eur. J. Nutr. 2012, 51, 29–37. [Google Scholar] [CrossRef]
- Torres-Fuentes, C.; Contreras, M.D.M.; Recio, I.; Alaiz, M.; Vioque, J. Identification and Characterization of Antioxidant Peptides from Chickpea Protein Hydrolysates. Food Chem. 2015, 180, 194–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.-Y.; Shyu, Y.-S.; Wang, Y.-T.; Hsu, C.-K. Antioxidative Properties of Protein Hydrolysate from Defatted Peanut Kernels Treated with Esperase. LWT-Food Sci. Technol. 2010, 43, 285–290. [Google Scholar] [CrossRef]
- Girjal, V.U.; Neelagund, S.; Krishnappa, M. Antioxidant Properties of the Peptides Isolated From Ganoderma Lucidum Fruiting Body. Int. J. Pept. Res. Ther. 2012, 18, 319–325. [Google Scholar] [CrossRef]
- Hong, J.; Chen, T.T.; Hu, P.; Yang, J.; Wang, S.Y. Purification and Characterization of an Antioxidant Peptide (GSQ) from Chinese Leek (Allium Tuberosum Rottler) Seeds. J. Funct. Foods 2014, 10, 144–153. [Google Scholar] [CrossRef]
- Ningappa, M.B.; Srinivas, L. Purification and Characterization of ∼35 KDa Antioxidant Protein from Curry Leaves (Murraya koenigii L.). Toxicol. In Vitro 2008, 22, 699–709. [Google Scholar] [CrossRef]
- Siswoyo, T.A.; Mardiana, E.; Lee, K.O.; Hoshokawa, K. Isolation and Characterization of Antioxidant Protein Fractions from Melinjo (Gnetum Gnemon) Seeds. J. Agric. Food Chem. 2011, 59, 5648–5656. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.; Raut, H.N.; Wagh, R.S.; Puntambekar, H.M.; Kulkarni, M.J. Purification and Characterization of an Antioxidant Protein (∼16 KDa) from Terminalia Chebula Fruit. Food Chem. 2012, 131, 141–148. [Google Scholar] [CrossRef]
- Costamagna, M.S.; Ordoñez, R.M.; Zampini, I.C.; Sayago, J.E.; Isla, M.I. Nutritional and Antioxidant Properties of Geoffroea Decorticans, an Argentinean Fruit, and Derived Products (Flour, Arrope, Decoction and Hydroalcoholic Beverage). Food Res. Int. 2013, 54, 160–168. [Google Scholar] [CrossRef]
- Maestri, D.M.; Fortunato, R.H.; Greppi, J.A.; Lamarque, A.L. Compositional Studies of Seeds and Fruits from Two Varieties of Geoffroea Decorticans. J. Food Compos. Anal. 2001, 14, 585–590. [Google Scholar] [CrossRef]
- Lamarque, A.L.; Maestri, D.M.; Zygadlo, J.A.; Guzmán, C.A. Chemical Evaluation of Geoffroea Decorticans Seeds as Source of Oil and Protein. Grasas y Aceites. 2000, 241–243. [Google Scholar] [CrossRef] [Green Version]
- Costamagna, M.S.; Zampini, I.C.; Alberto, M.R.; Cuello, S.; Torres, S.; Pérez, J.; Quispe, C.; Schmeda-Hirschmann, G.; Isla, M.I. Polyphenols Rich Fraction from Geoffroea Decorticans Fruits Flour Affects Key Enzymes Involved in Metabolic Syndrome, Oxidative Stress and Inflammatory Process. Food Chem. 2016, 190, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Orrabalis, C.J.; Mufari, R.Y.; Gorostegui, H.A.; Calandri, E.L.; Guzmán, C.A. Chemical and Nutritional Characterization of Fruits from Geoffroea Decorticans Tree (Chañar) and Their Parts, from Argentine Subtropical Forest. Int. J. Adv. Res. Bot. 2015, 1, 23–33. [Google Scholar]
- Pereira, R.D.A.; Valencia-Jiménez, A.; Magalhães, C.P.; Prates, M.V.; Melo, J.A.T.; De Lima, L.M.; De Sales, M.P.; Nakasu, E.Y.T.; Da Silva, M.C.M.; Grossi-De-Sá, M.F. Effect of a Bowman-Birk Proteinase Inhibitor from Phaseolus Coccineus on Hypothenemus Hampei Gut Proteinases in Vitro. J. Agric. Food Chem. 2007, 55, 10714–10719. [Google Scholar] [CrossRef]
- Quiroga, E.N.; Sampietro, D.A.; Sgariglia, M.A.; Soberón, J.R.; Vattuone, M.A. Antimycotic Activity of 5′-Prenylisoflavanones of the Plant Geoffroea Decorticans, against Aspergillus Species. Int. J. Food Microbiol. 2009, 132, 42–46. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chevallet, M.; Luche, S.; Rabilloud, T. Silver Staining of Proteins in Polyacrylamide Gels. Nat. Protoc. 2006, 1, 1852–1858. [Google Scholar] [CrossRef] [Green Version]
- Obregón, W.D.; Liggieri, C.S.; Morcelle, S.R.; Trejo, S.A.; Avilés, F.X.; Priolo, N.S. Biochemical and PMF MALDI-TOF Analyses of Two Novel Papain-like Plant Proteinases. Protein Pept. Lett. 2009, 16, 1323–1333. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of Free Radical Method to Evaluate Antioxidant Activity. Leb. Und-Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Pukalskas, A.; van Beek, T.A.; Venskutonis, R.P.; Linssen, J.P.H.; van Veldhuizen, A.; de Groot, A. Identification of Radical Scavengers in Sweet Grass (Hierochloe Odorata). J. Agric. Food Chem. 2002, 50, 2914–2919. [Google Scholar] [CrossRef] [PubMed]
- Cotabarren, J.; Broitman, D.J.; Quiroga, E.; Obregón, W.D. GdTI, the First Thermostable Trypsin Inhibitor from Geoffroea Decorticans Seeds. A Novel Natural Drug with Potential Application in Biomedicine. Int. J. Biol. Macromol. 2020, 148, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T.J. Antioxidative Peptides Derived from Plants for Human Nutrition: Their Production, Mechanisms and Applications. Eur. Food Res. Technol. 2020. [Google Scholar] [CrossRef]
- Daliri, E.B.M.; Oh, D.H.; Lee, B.H. Bioactive Peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef]
- Maleki, M.H.; Daneshniya, M.; Keshavarzbahadori, N. A Review of Biological Properties of Bioactive Peptides: Antioxidant Activity A Review of Biological Properties of Bioactive Peptides: Antioxidant Activity. In Proceedings of the 3rd International Congress of Science, Hamburg, Germany, 19–22 March 2020. [Google Scholar]
- Wong, F.C.; Xiao, J.; Wang, S.; Ee, K.Y.; Chai, T.T. Advances on the Antioxidant Peptides from Edible Plant Sources. Trends Food Sci. Technol. 2020, 99, 44–57. [Google Scholar] [CrossRef]
- Arnao, M.B. Some Methodological Problems in the Determination of Antioxidant Activity Using Chromogen Radicals: A Practical Case. Trends Food Sci. Technol. 2000, 11, 419–421. [Google Scholar] [CrossRef]
- Zou, T.B.; He, T.P.; Li, H.B.; Tang, H.W.; Xia, E.Q. The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins. Molecules 2016, 21, 72. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Food-Derived Peptidic Antioxidants: A Review of Their Production, Assessment, and Potential Applications. J. Funct. Foods 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Malomo, S.; Offengenden, M.; Wu, J.; Aluko, R.E. Structural and Functional Characterization of Hemp Seed (Cannabis Sativa L.) Protein-Derived Antioxidant and Antihypertensive Peptides. J. Funct. Foods 2014, 6, 384–394. [Google Scholar] [CrossRef]
- Huang, W.; Deng, Q.; Xie, B.; Shi, J.; Huang, F.H.; Tian, B.; Huang, Q.; Xue, S. Purification and Characterization of an Antioxidant Protein from Ginkgo Biloba Seeds. Food Res. Int. 2010, 43, 86–94. [Google Scholar] [CrossRef]
- Ashaolu, T.J.; Yupanqui, C.T. Hypoallergenic and Immunomodulatory Prospects of Pepsin-Educed Soy Protein Hydrolysates. Croat. J. Food Sci. Technol. 2018, 10, 270–278. [Google Scholar] [CrossRef]
- Reddi, S.; Kapila, R.; Dang, A.K.; Kapila, S. Evaluation of Allergenic Response of Milk Bioactive Peptides Using Mouse Mast Cell. Milchwissenschaft 2012, 67, 189–191. [Google Scholar]
- Yili, A.; Ma, Q.L.; Gao, Y.H.; Zhao, B.; Jun, D.; Aisa, H.A. Isolation of Two Antioxidant Peptides from Seeds of Apium Graveolens Indigenous to China. Chem. Nat. Compd. 2012, 48, 719–720. [Google Scholar] [CrossRef]
- Boonmee, A.; Srisomsap, C.; Karnchanatat, A.; Sangvanich, P. An Antioxidant Protein in Curcuma Comosa Roxb. Rhizomes. Food Chem. 2011, 124, 476–480. [Google Scholar] [CrossRef]
- Sarkar, M.K.; Kinter, M.; Mazumder, B.; Sil, P.C. Purification and Characterisation of a Novel Antioxidant Protein Molecule from Phyllanthus Niruri. Food Chem. 2009, 114, 1405–1412. [Google Scholar] [CrossRef]
- World Health Organization. Cardiovascular Diseases (CVDs). Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 10 June 2021).
- Hylek, E.M.; Evans-Molina, C.; Shea, C.; Henault, L.E.; Regan, S. Major Hemorrhage and Tolerability of Warfarin in the First Year of Therapy among Elderly Patients with Atrial Fibrillation. Circulation 2007, 115, 2689–2696. [Google Scholar] [CrossRef] [Green Version]
- Indumathi, P.; Mehta, A. A Novel Anticoagulant Peptide from the Nori Hydrolysate. J. Funct. Foods 2016, 20, 606–617. [Google Scholar] [CrossRef]
- Oliva, M.L.; Souza-Pinto, J.C.; Batista, I.F.; Araujo, M.S.; Silveira, V.F.; Auerswald, E.A.; Mentele, R.; Eckerskorn, C.; Sampaio, M.U.; Sampaio, C.A. Leucaena Leucocephala Serine Proteinase Inhibitor: Primary Structure and Action on Blood Coagulation, Kinin Release and Rat Paw Edema. Biochim. Biophys. Acta 2000, 1477, 64–74. [Google Scholar] [CrossRef]
- Batista, I.F.; Oliva, M.L.; Araujo, M.S.; Sampaio, M.U.; Richardson, M.; Fritz, H.; Sampaio, C.A. Primary Structure of a Kunitz-Type Trypsin Inhibitor from Enterolobium Contortisiliquum Seeds. Phytochemistry 1996, 41, 1017–1022. [Google Scholar] [CrossRef]
- Lazza, C.; Trejo, S.; Obregon, W.; Pistaccio, L.; Caffini, N.; Lopez, L. A Novel Trypsin and α-Chymotrypsin Inhibitor from Maclura Pomifera Seeds. Lett. Drug Des. Discov. 2010, 7, 244–249. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Soluble Protein (mg) | Yield | Inhibition of DPPH IC50 (µg/mL) | Purification Fold (a) | Inhibition of ABTS IC50 (µg/mL) | Purification Fold (b) |
|---|---|---|---|---|---|---|
| Crude extract | 21.4 ± 0.4 | 100 | 82.5 ± 1.2 | 1 | 175.7 ± 3.1 | 1 |
| 90 °C heat treatment | 17.7 ± 0.5 | 82.6 ± 0.7 | 63.2 ± 0.8 | 1.3 ± 0.1 | 141.3 ± 2.7 | 1.2 ± 0.1 |
| Size-exclusion chromatography | 3.1 ± 0.2 | 14.1 ± 0.3 | 35.5 ± 0.5 | 2.3 ± 0.1 | 95.1 ± 0.9 | 1.9 ± 0.1 |
| AntioxidantPeptide/Protein | Plant Source | MW (Da) | IC50 by DPPH (µg/mL) | IC50 by ABTS (µg/mL) | References |
|---|---|---|---|---|---|
| GSQ | Allium tuberosum Rottler | 290.10 | 610 | 40 | [28] |
| G4b | Ginkgo biloba seeds | 29,247 | 100.7 | n/d | [53] |
| QCZDE | Apium graveolens seeds | 6500 | n/d | 158.7 | [56] |
| QCZCM | Apium graveolens seeds | 11,390 | n/d | 193.2 | [56] |
| Gg-AOPI | Gnetum gnemon seeds | 30,000 | 21 | 10 | [30] |
| Gg-AOPII | Gnetum gnemon seeds | 12,000 | 27 | 12 | [30] |
| Antioxidant protein from Curcuma comosa | Curcuma comosa Roxb. Rhizomes | 18,000 | n/d | n/d | [57] |
| PNP | Phyllanthus nuri leaves | 35,000 | n/d | n/d | [58] |
| TCP-III | Terminalia chebula fruit | 16,267 | 291 | n/d | [31] |
| APC | Murraya koenigii leaves | 35,000 | 70 | n/d | [29] |
| GdAPs | Geoffrea decorticans | 2000–8000 | 35.5 | 95.1 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cotabarren, J.; Ozón, B.; Claver, S.; Garcia-Pardo, J.; Obregón, W.D. Purification and Identification of Novel Antioxidant Peptides Isolated from Geoffroea decorticans Seeds with Anticoagulant Activity. Pharmaceutics 2021, 13, 1153. https://doi.org/10.3390/pharmaceutics13081153
Cotabarren J, Ozón B, Claver S, Garcia-Pardo J, Obregón WD. Purification and Identification of Novel Antioxidant Peptides Isolated from Geoffroea decorticans Seeds with Anticoagulant Activity. Pharmaceutics. 2021; 13(8):1153. https://doi.org/10.3390/pharmaceutics13081153
Chicago/Turabian StyleCotabarren, Juliana, Brenda Ozón, Santiago Claver, Javier Garcia-Pardo, and Walter David Obregón. 2021. "Purification and Identification of Novel Antioxidant Peptides Isolated from Geoffroea decorticans Seeds with Anticoagulant Activity" Pharmaceutics 13, no. 8: 1153. https://doi.org/10.3390/pharmaceutics13081153