
Cymbopogon winterianus Essential Oil Attenuates Bleomycin-Induced Pulmonary Fibrosis in a Murine Model

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site and Plant Material
2.2. Extraction and Analysis of the Essential Oil
2.3. Experimental Procedures of Lung Fibrosis: Induction and Treatment
2.4. Assessment of the Body Weight of the Animals
2.5. Hematological Analysis of the Peripheral Blood
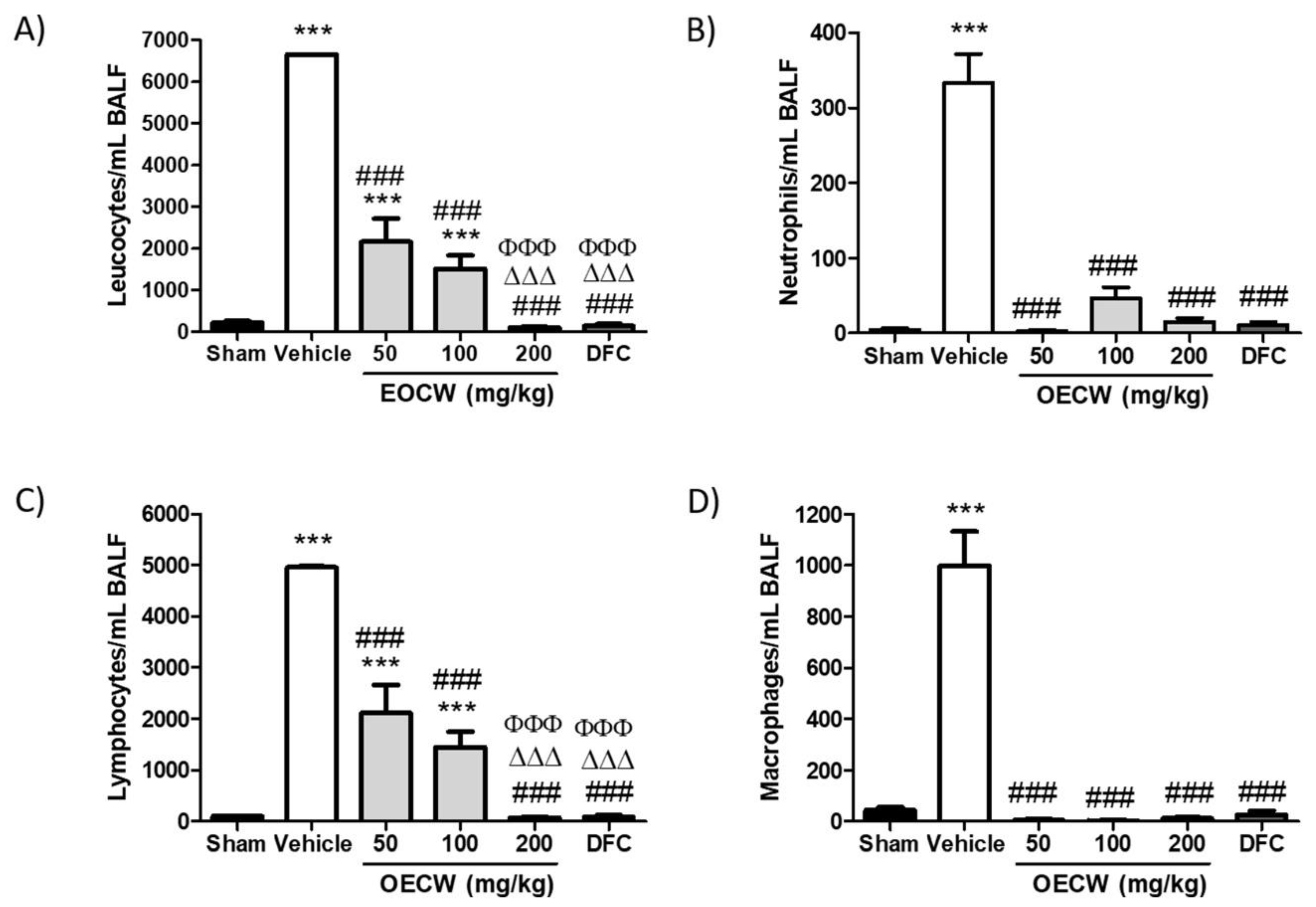
2.6. Analysis of the Bronchoalveolar Lavage Fluid (BALF)
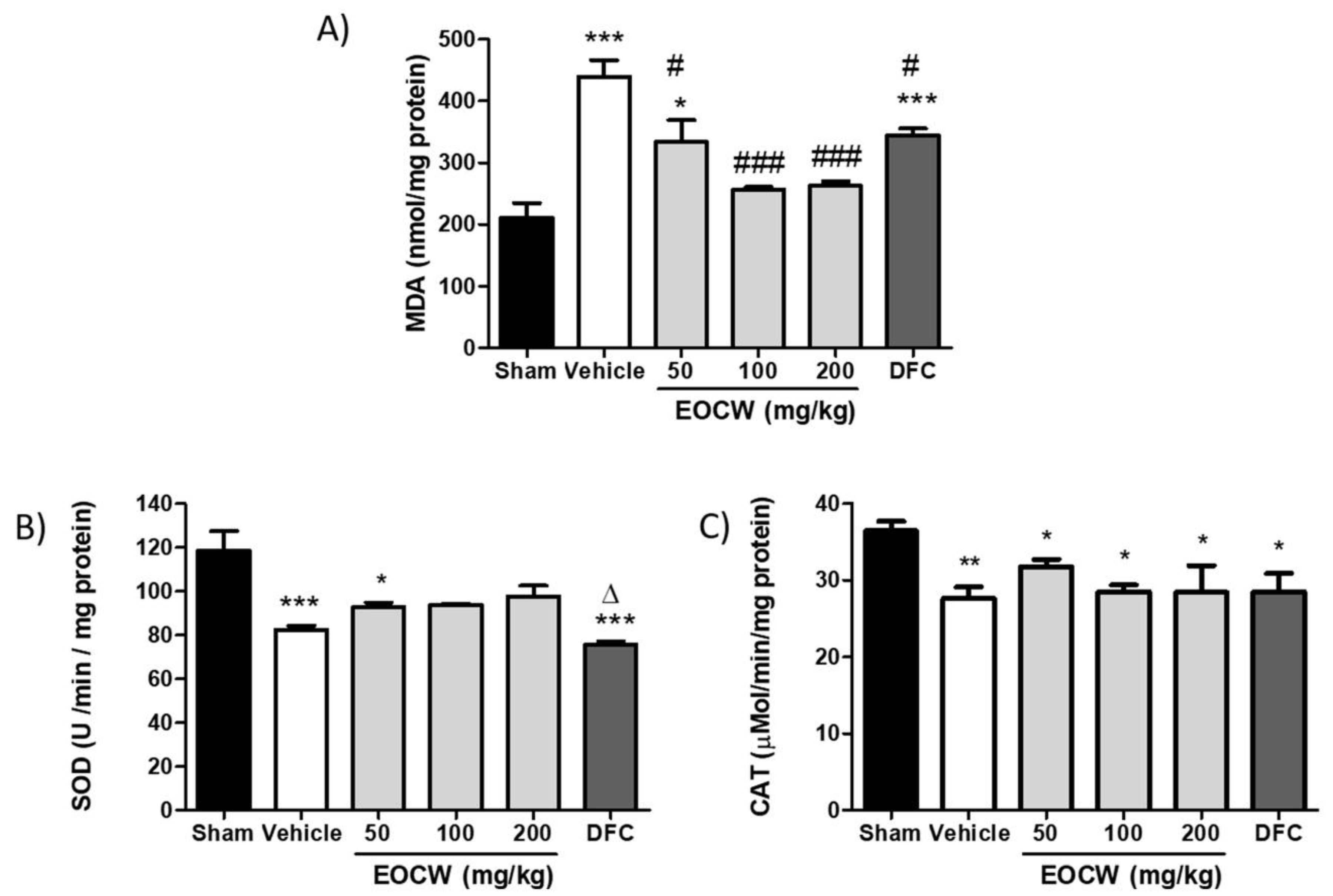
2.7. Assessment of Lipid Peroxidation and Mda Tissue Levels
2.8. Assessment of Superoxide Dismutase (SOD) Tissue Activity
2.9. Assessment of Catalase (CAT) Tissue Activity
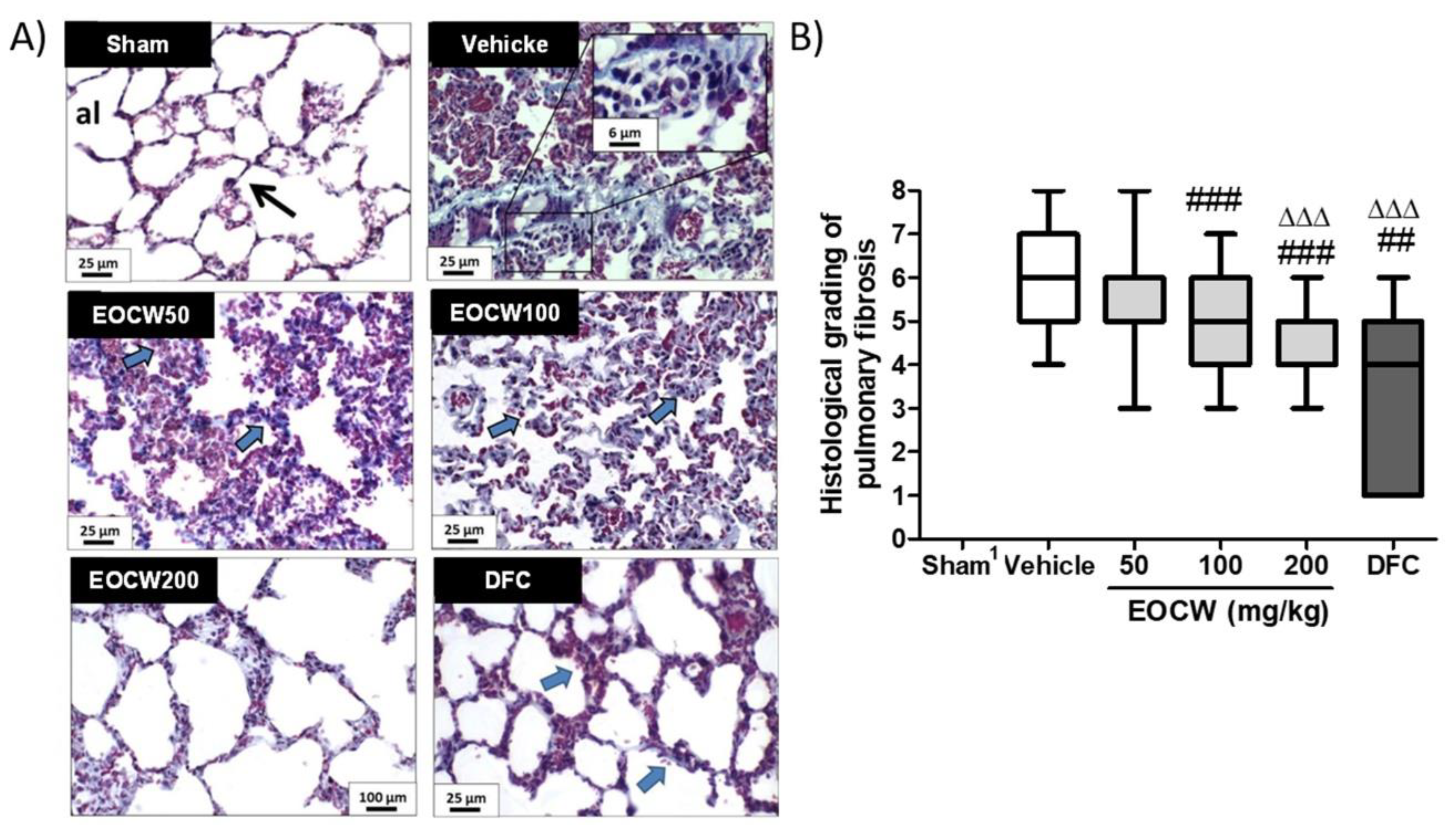
2.10. Histological Procedures and Assessment of the Histological Grading of Bleomycin-Induced Lung Fibrosis
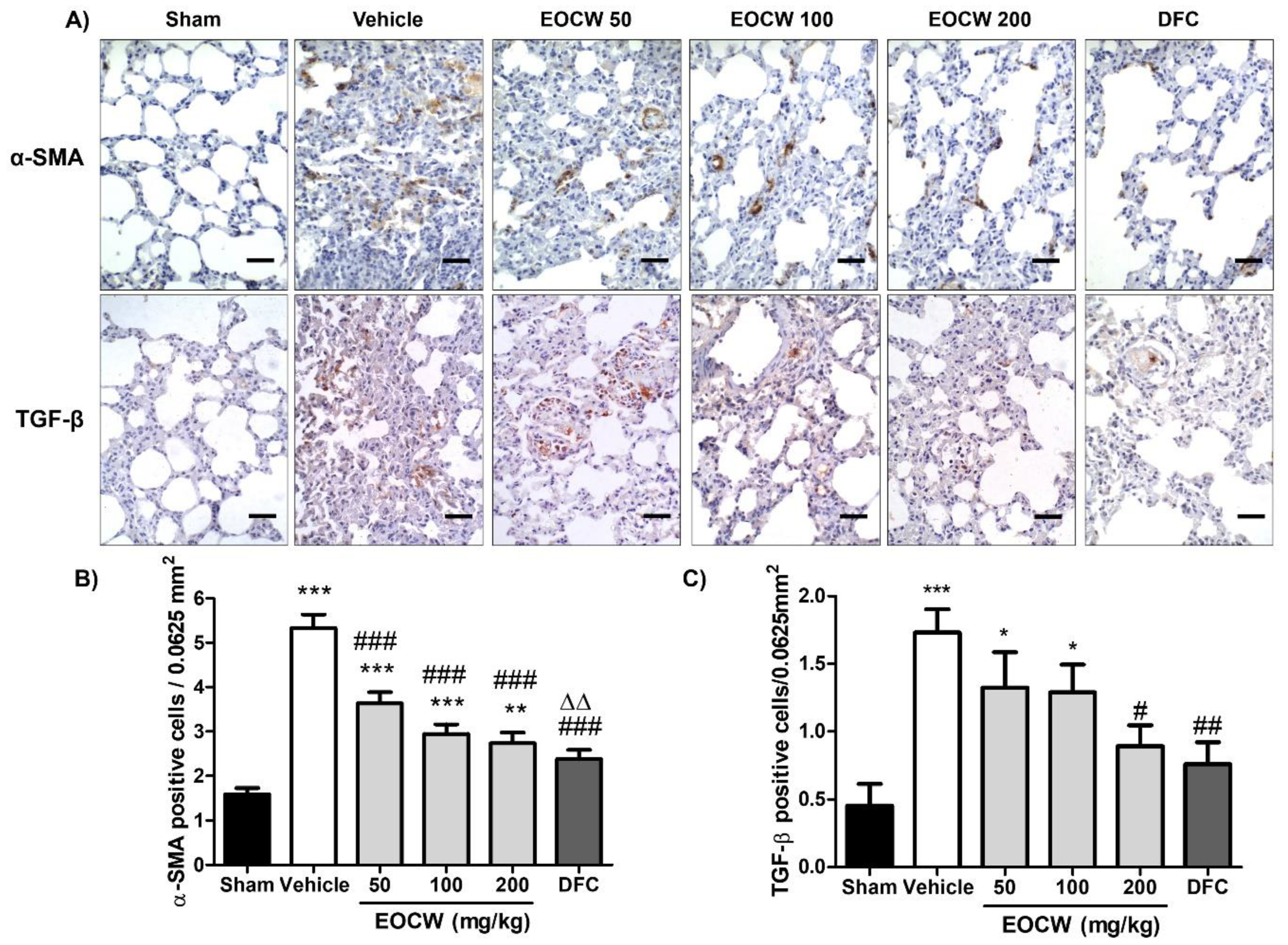
2.11. Analysis of the Immunohistochemical Expression of α-SMA and TGF-β in the Lung Tissue
2.12. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martinez, F.J.; Collard, H.R.; Pardo, A.; Raghu, G.; Richeldi, L.; Selman, M.; Swigris, J.J.; Taniguchi, H.; Wells, A.U. Idiopathic pulmonary fibrosis. Nat. Rev. Dis. Primers 2017, 3, 17074. [Google Scholar] [CrossRef] [PubMed]
- Sgalla, G.; Iovene, B.; Calvello, M.; Ori, M.; Varone, F.; Richeldi, L. Idiopathic pulmonary fibrosis: Pathogenesis and management. Respir. Res. 2018, 19, 32. [Google Scholar] [CrossRef] [PubMed]
- Jafri, S.; Ahmed, N.; Saifullah, N.; Musheer, M. Epidemiology and Clinico-radiological features of Interstitial Lung Diseases. Pak. J. Med. Sci. 2020, 36, 365–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, K.C. Pulmonary fibrosis, part I: Epidemiology, pathogenesis, and diagnosis. Expert Rev. Respir. Med. 2017, 11, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Bahri, S.; Ben Ali, R.; Abidi, A.; Jameleddine, S. The efficacy of plant extract and bioactive compounds approaches in the treatment of pulmonary fibrosis: A systematic review. Biomed. Pharmacother. 2017, 93, 666–673. [Google Scholar] [CrossRef]
- Evans, C.M.; Fingerlin, T.E.; Schwarz, M.I.; Lynch, D.; Kurche, J.; Warg, L.; Yang, I.V.; Schwartz, D.A. Idiopathic Pulmonary Fibrosis: A Genetic Disease That Involves Mucociliary Dysfunction of the Peripheral Airways. Physiol. Rev. 2016, 96, 1567–1591. [Google Scholar] [CrossRef]
- Raghu, G.; Remy-Jardin, M.; Myers, J.L.; Richeldi, L.; Ryerson, C.J.; Lederer, D.J.; Behr, J.; Cottin, V.; Danoff, S.K.; Morell, F.; et al. Diagnosis of Idiopathic Pulmonary Fibrosis. An Official ATS/ERS/JRS/ALAT Clinical Practice Guideline. Am. J. Respir. Crit. Care Med. 2018, 198, e44–e68. [Google Scholar] [CrossRef]
- Salton, F.; Volpe, M.C.; Confalonieri, M. Epithelial⁻Mesenchymal Transition in the Pathogenesis of Idiopathic Pulmonary Fibrosis. Medicina 2019, 55, 83. [Google Scholar] [CrossRef] [Green Version]
- Zaafan, M.A.; Haridy, A.R.; Abdelhamid, A.M. Amitriptyline attenuates bleomycin-induced pulmonary fibrosis: Modulation of the expression of NF-κβ, iNOS, and Nrf2. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 279–286. [Google Scholar] [CrossRef]
- Li, L.; Li, Q.; Wei, L.; Wang, Z.; Ma, W.; Liu, F.; Shen, Y.; Zhang, S.; Zhang, X.; Li, H.; et al. Dexamethasone combined with berberine is an effective therapy for bleomycin-induced pulmonary fibrosis in rats. Exp. Ther. Med. 2019, 18, 2385–2392. [Google Scholar] [CrossRef]
- Yue, X.; Shan, B.; Lasky, J.A. TGF-β: Titan of Lung Fibrogenesis. Curr. Enzym. Inhib. 2010, 6. [Google Scholar] [CrossRef]
- Warshamana, G.S.; Pociask, D.A.; Fisher, K.J.; Liu, J.-Y.; Sime, P.J.; Brody, A.R. Titration of non-replicating adenovirus as a vector for transducing active TGF-beta1 gene expression causing inflammation and fibrogenesis in the lungs of C57BL/6 mice. Int. J. Exp. Pathol. 2002, 83, 183–201. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, A.; Heldin, C.-H. Mechanisms of TGFβ-Induced Epithelial-Mesenchymal Transition. J. Clin. Med. 2016, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Ma, Q. Myofibroblasts and lung fibrosis induced by carbon nanotube exposure. Part. Fibre Toxicol. 2016, 13, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sontake, V.; Gajjala, P.R.; Kasam, R.K.; Madala, S.K. New therapeutics based on emerging concepts in pulmonary fibrosis. Expert Opin. Ther. Targets 2019, 23, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Gharaee-Kermani, M.; Gyetko, M.R.; Hu, B.; Phan, S.H. New insights into the pathogenesis and treatment of idiopathic pulmonary fibrosis: A potential role for stem cells in the lung parenchyma and implications for therapy. Pharm. Res. 2007, 24, 819–841. [Google Scholar] [CrossRef]
- Bahri, S.; Ben Ali, R.; Nahdi, A.; Mlika, M.; Abdennabi, R.; Jameleddine, S. Salvia officinalis attenuates bleomycin-induced oxidative stress and lung fibrosis in rats. Nutr. Cancer 2020, 72, 1135–1145. [Google Scholar] [CrossRef]
- Sun, L.; Mao, M.; Yan, Z.; Zuo, C.; Zhang, X. A Chinese Traditional Therapy for Bleomycin-Induced Pulmonary Fibrosis in Mice. Can. Respir. J. 2018, 2018, 8491487. [Google Scholar] [CrossRef] [PubMed]
- Leite, B.L.S.; Souza, T.T.; Antoniolli, Â.R.; Guimarães, A.G.; Siqueira, R.S.; Quintans, J.S.S.; Bonjardim, L.R.; Alves, P.B.; Blank, A.F.; Botelho, M.A.; et al. Volatile constituents and behavioral change induced by Cymbopogon winterianus leaf essential oil in rodents. Afr. J. Biotechnol. 2011, 10, 8312–8319. [Google Scholar]
- Sharma, R.; Rao, R.; Kumar, S.; Mahant, S.; Khatkar, S. Therapeutic Potential of Citronella Essential Oil: A Review. Curr. Drug Discov. Technol. 2019, 16, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Quintans-Júnior, L.J.; Souza, T.T.; Leite, B.S.; Lessa, N.M.; Bonjardim, L.R.; Santos, M.R.; Alves, P.B.; Blank, A.F.; Antoniolli, A.R. Phythochemical screening and anticonvulsant activity of Cymbopogon winterianus Jowitt (Poaceae) leaf essential oil in rodents. Phytomedicine Int. J. Phytother. Phytopharm. 2008, 15, 619–624. [Google Scholar] [CrossRef]
- Tiwari, M.; Kakkar, P. Plant derived antioxidants—Geraniol and camphene protect rat alveolar macrophages against t-BHP induced oxidative stress. Toxicol. In Vitro 2009, 23, 295–301. [Google Scholar] [CrossRef]
- Tsai, M.-L.; Lin, C.-D.; Khoo, K.A.; Wang, M.-Y.; Kuan, T.-K.; Lin, W.-C.; Zhang, Y.-N.; Wang, Y.-Y. Composition and Bioactivity of Essential Oil from Citrus grandis (L.) Osbeck ‘Mato Peiyu’ Leaf. Molecules 2017, 22, 2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, P.L.; Matos, J.; Picot, L.; Almeida, J.; Quintans, J.S.S.; Quintans-Júnior, L.J. Citronellol, a monoterpene alcohol with promising pharmacological activities—A systematic review. Food Chem. Toxicol. 2019, 123, 459–469. [Google Scholar] [CrossRef]
- Jayachandran, M.; Chandrasekaran, B.; Namasivayam, N. Geraniol attenuates fibrosis and exerts anti-inflammatory effects on diet induced atherogenesis by NF-κB signaling pathway. Eur. J. Pharmacol. 2015, 762, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.-M.; Wang, X.-B.; Zou, N.; Han, C.; Xu, J. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis of the Volatile Oil of Cichorium Glandulosum Boiss et Huet and its Effects on Carbon Tetrachloride-Induced Liver Fibrosis in Rats. Med. Sci. Monit. 2019, 25, 3591–3604. [Google Scholar] [CrossRef]
- Silveira, G.d.O.; Loddi, S.; de Oliveira, C.D.R.; Zucoloto, A.D.; Fruchtengarten, L.V.G.; Yonamine, M. Headspace solid-phase microextraction and gas chromatography—Mass spectrometry for determination of cannabinoids in human breast milk. Forensic Toxicol. 2017, 35, 125–132. [Google Scholar] [CrossRef]
- Sparkman, O.D. Identification of essential oil components by gas chromatography/quadrupole mass spectroscopy Robert P. Adams. J. Am. Soc. Mass Spectrom. 2005, 16, 1902–1903. [Google Scholar] [CrossRef] [Green Version]
- van Den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas–liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Ashcroft, T.; Simpson, J.M.; Timbrell, V. Simple method of estimating severity of pulmonary fibrosis on a numerical scale. J. Clin. Pathol. 1988, 41, 467–470. [Google Scholar] [CrossRef] [Green Version]
- Hübner, R.H.; Gitter, W.; El Mokhtari, N.E.; Mathiak, M.; Both, M.; Bolte, H.; Freitag-Wolf, S.; Bewig, B. Standardized quantification of pulmonary fibrosis in histological samples. BioTechniques 2008, 44, 507–511, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.M.; Lima, A.K.; Melo, M.G.D.; Dória, G.A.A.; Serafini, M.R.; Albuquerque-Júnor, R.L.C.; Araújo, A.A.S. Valores de referência hematológicos e bioquímicos de ratos (Rattus novergicus linhagem Wistar) provenientes do biotério da Universidade Tiradentes. Scientia Plena 2014, 10. [Google Scholar]
- Gonzalez-Perez, O.; Luquin, S.; Garcia-Estrada, J.; Ramos-Remus, C. Deflazacort: A glucocorticoid with few metabolic adverse effects but important immunosuppressive activity. Adv. Ther. 2007, 24, 1052–1060. [Google Scholar] [CrossRef]
- de Castro Pereira, C.A.; Baddini-Martinez, J.A.; Baldi, B.G.; de Oliveira Jezler, S.F.; Rubin, A.S.; Rufino Alves, R.L.; Alves Zonzin, G.; Quaresma, M.; Trampisch, M.; Rabahi, M.F. Segurança e tolerabilidade de Nintedanibe em pacientes com fibrose pulmonar idiopática no Brasil. J. Bras. Pneumol. 2019, 45, e20180414. [Google Scholar] [CrossRef] [PubMed]
- National Clinical Guideline Centre. National Institute for Health and Care Excellence: Clinical Guidelines. In Diagnosis and Management of Suspected Idiopathic Pulmonary Fibrosis: Idiopathic Pulmonary Fibrosis; Royal College of Physicians: London, UK, 2013. [Google Scholar]
- Ding, W.; Ding, L.; Li, W.B.; Pan, H.; Lin, H.D. Pharmacokinetics of deflazacort tablets in healthy Chinese volunteers. Yao Xue Xue Bao Acta Pharm. Sin. 2014, 49, 921–926. [Google Scholar]
- Leite, B.L.; Bonfim, R.R.; Antoniolli, A.R.; Thomazzi, S.M.; Araújo, A.A.; Blank, A.F.; Estevam, C.S.; Cambui, E.V.; Bonjardim, L.R.; Albuquerque, R.L., Jr.; et al. Assessment of antinociceptive, anti-inflammatory and antioxidant properties of Cymbopogon winterianus leaf essential oil. Pharm. Biol. 2010, 48, 1164–1169. [Google Scholar] [CrossRef]
- Silva, C.T.S.; Wanderley-Teixeira, V.; Cunha, F.M.; Oliveira, J.V.; Dutra, K.A.; Navarro, D.M.A.F.; Teixeira, A.A.C. Effects of citronella oil (Cymbopogon winterianus Jowitt ex Bor) on Spodoptera frugiperda (J. E. Smith) midgut and fat body. Biotech. Histochem. 2018, 93, 36–48. [Google Scholar] [CrossRef]
- Okoh, S.O.; Okoh, O.O.; Okoh, A.I. Seasonal variation of volatile oil composition and antioxidant property of aerial parts of Syzygium paniculatum Gaertn. grown in the Eastern Cape, South Africa. Nat. Prod. Res. 2019, 33, 2276–2280. [Google Scholar] [CrossRef] [PubMed]
- Zribi, I.; Bleton, J.; Moussa, F.; Abderrabba, M. GC-MS analysis of the volatile profile and the essential oil compositions of Tunisian Borago Officinalis L.: Regional locality and organ dependency. Ind. Crop. Prod. 2019, 129, 290–298. [Google Scholar] [CrossRef]
- Niyibituronsa, M.; Onyango, A.N.; Gaidashova, S.; Imathiu, S.; Ming, Z.; Ruinan, Y.; Weiqi, Z.; XiuPin, W.; Qi, Z.; Zhaowei, Z.; et al. Evaluation of Five Essential Oils by Gas Chromatography-Mass Spectrometry and their Effect on Fungal Growth Inhibition and Sensory Acceptability of Soymilk. J. Food Res. 2020, 9, 36–47. [Google Scholar] [CrossRef]
- Solanki, K.P.; Desai, M.A.; Parikh, J.K. Sono hydrodistillation for isolation of citronella oil: A symbiotic effect of sonication and hydrodistillation towards energy efficiency and environment friendliness. Ultrason. Sonochem. 2018, 49, 145–153. [Google Scholar] [CrossRef]
- Barbayianni, I.; Ninou, I.; Tzouvelekis, A.; Aidinis, V. Bleomycin Revisited: A Direct Comparison of the Intratracheal Micro-Spraying and the Oropharyngeal Aspiration Routes of Bleomycin Administration in Mice. Front. Med. 2018, 5, 269. [Google Scholar] [CrossRef]
- Jost, R.T.; Dias, H.B.; Krause, G.C.; de Souza, R.G.; de Souza, T.R.; Nuñez, N.K.; Dos Santos, F.G.; Haute, G.V.; da Silva Melo, D.A.; Pitrez, P.M.; et al. Fructose-1,6-Bisphosphate Prevents Bleomycin-Induced Pulmonary Fibrosis in Mice and Inhibits the Proliferation of Lung Fibroblasts. Inflammation 2018, 41, 1987–2001. [Google Scholar] [CrossRef]
- Dias, H.B.; de Oliveira, J.R.; Donadio, M.V.F.; Kimura, S. Fructose-1,6-bisphosphate prevents pulmonary fibrosis by regulating extracellular matrix deposition and inducing phenotype reversal of lung myofibroblasts. PLoS ONE 2019, 14, e0222202. [Google Scholar] [CrossRef]
- Hajjaji, N.; Couet, C.; Besson, P.; Bougnoux, P. DHA effect on chemotherapy-induced body weight loss: An exploratory study in a rodent model of mammary tumors. Nutr. Cancer 2012, 64, 1000–1007. [Google Scholar] [CrossRef]
- Rizzello, F.; Ricci, C.; Scandella, M.; Cavazza, E.; Giovanardi, E.; Valerii, M.C.; Campieri, M.; Comparone, A.; De Fazio, L.; Candela, M.; et al. Dietary geraniol ameliorates intestinal dysbiosis and relieves symptoms in irritable bowel syndrome patients: A pilot study. BMC Complement. Altern. Med. 2018, 18, 338. [Google Scholar] [CrossRef]
- Zeng, H.; Lazarova, D.L.; Bordonaro, M. Mechanisms linking dietary fiber, gut microbiota and colon cancer prevention. World J. Gastrointest. Oncol. 2014, 6, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Cowley, P.M.; Roberts, C.R.; Baker, A.J. Monitoring the Health Status of Mice with Bleomycin-induced Lung Injury by Using Body Condition Scoring. Comp. Med. 2019, 69, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Kaplan, A.M.; Goud, S.N. Immunologic alterations in bleomycin-treated mice: Role of pulmonary fibrosis in the modulation of immune responses. Am. J. Respir. Crit. Care Med. 1996, 153, 1924–1930. [Google Scholar] [CrossRef] [PubMed]
- Arun, K.; Vankatachalam, V.V.; Deepak, U.; Tiwari, V.K.; Aakash, S. Combined Effect of N-Acetyl Cysteine and Clarithromycin on Bleomycin Induced Pulmonary Fibrosis. Int. Biol. Biomed. J. 2018, 4, 14–23. [Google Scholar]
- Ding, Y.; Xiao, L.; Chen, R.; Qing, Y.; Gao, Y.; Jiang, Q.; Chen, X. Efficacy and Safety of Inhaled Anti-Fibrotic Peptide HTPEP-001 in Rat Models. In A64. Therapeutics, Drug Delivery, and Tissue Engineering. American Journal of Respiratory and Critical Care Medicine 2020; American Thoraic Society: New York, NY, USA, 2020; Volume 201, p. A2286. [Google Scholar] [CrossRef]
- Qiu, M.; An, M.; Bian, M.; Yu, S.; Liu, C.; Liu, Q. Terrestrosin D from Tribulus terrestris attenuates bleomycin-induced inflammation and suppresses fibrotic changes in the lungs of mice. Pharm. Biol. 2019, 57, 694–700. [Google Scholar] [CrossRef] [Green Version]
- Lucattelli, M.; Fineschi, S.; Selvi, E.; Garcia Gonzalez, E.; Bartalesi, B.; De Cunto, G.; Lorenzini, S.; Galeazzi, M.; Lungarella, G. Ajulemic acid exerts potent anti-fibrotic effect during the fibrogenic phase of bleomycin lung. Respir. Res. 2016, 17, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, W.; Zhang, X.-Y.; Pan, M.; Zhao, B.; Chen, C.; Niu, Z.-H.; Huang, C.-L.; Li, Y.-Y.; Fan, X.-M.; Ma, Y.-M.; et al. Interleukin-17A promotes the formation of inflammation in the lung tissues of rats with pulmonary fibrosis. Exp Ther. Med 2015, 10, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-S.; Li, Q.; Youn, H.-Y.; Kim, D.Y. Oral Administration of Chitosan Attenuates Bleomycin-induced Pulmonary Fibrosis in Rats. In Vivo 2019, 33, 1455–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaghloul, M.S.; Said, E.; Suddek, G.M.; Salem, H.A. Crocin attenuates lung inflammation and pulmonary vascular dysfunction in a rat model of bleomycin-induced pulmonary fibrosis. Life Sci. 2019, 235, 116794. [Google Scholar] [CrossRef] [PubMed]
- Kabel, A.M.; Estfanous, R.S.; Alrobaian, M.M. Targeting oxidative stress, proinflammatory cytokines, apoptosis and toll like receptor 4 by empagliflozin to ameliorate bleomycin-induced lung fibrosis. Respir. Physiol. Neurobiol. 2020, 273, 103316. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cai, L.; Zheng, L.; Hu, Y.; Yuan, W.; Guo, Z.; Li, W. Gefitinib Inhibits Bleomycin-Induced Pulmonary Fibrosis via Alleviating the Oxidative Damage in Mice. Oxid. Med. Cell. Longev. 2018, 2018, 8249693. [Google Scholar] [CrossRef]
- Mansouri, N.; Willis, G.R.; Fernandez-Gonzalez, A.; Reis, M.; Nassiri, S.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal stromal cell exosomes prevent and revert experimental pulmonary fibrosis through modulation of monocyte phenotypes. JCI Insight 2019, 4, e128060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.X.; Qiu, J.; Zhou, H.H.; Yu, Y.; Zhou, D.L.; Xu, Y.; Zhu, M.Z.; Ge, X.P.; Li, J.M.; Lv, C.J.; et al. Dihydroartemisinin alleviates oxidative stress in bleomycin-induced pulmonary fibrosis. Life Sci. 2018, 205, 176–183. [Google Scholar] [CrossRef]
- Qian, Y.; Han, Q.; Zhao, X.; Song, J.; Cheng, Y.; Fang, Z.; Ouyang, Y.; Yuan, W.E.; Fan, C. 3D melatonin nerve scaffold reduces oxidative stress and inflammation and increases autophagy in peripheral nerve regeneration. J. Pineal Res. 2018, 65, e12516. [Google Scholar] [CrossRef]
- Prasad, S.N.; Muralidhara, M. Analysis of the antioxidant activity of geraniol employing various in vitro models: Relevance to neurodegeneration in diabetic neuropathy. Asian J. Pharm. Clin. Res. 2017, 10, 101–105. [Google Scholar] [CrossRef]
- El-Emam, S.Z.; Soubh, A.A.; Al-Mokaddem, A.K.; Abo El-Ella, D.M. Geraniol activates Nrf-2/HO-1 signaling pathway mediating protection against oxidative stress-induced apoptosis in hepatic ischemia-reperfusion injury. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 1849–1858. [Google Scholar] [CrossRef]
- Sahakyan, N.; Petrosyan, M.; Trchounian, A. Some peculiarities of essential oil of Damask Rose growing in high altitude Armenian Landscapes. Proc. YSU B Chem. Biol. Sci. 2020, 54, 68–74. [Google Scholar] [CrossRef]
- Liu, T.; De Los Santos, F.G.; Phan, S.H. The Bleomycin Model of Pulmonary Fibrosis. In Fibrosis; Rittié, L., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2017; Volume 1627, pp. 27–42. [Google Scholar] [CrossRef]
- Ruscitti, F.; Ravanetti, F.; Essers, J.; Ridwan, Y.; Belenkov, S.; Vos, W.; Ferreira, F.; KleinJan, A.; van Heijningen, P.; Van Holsbeke, C.; et al. Longitudinal assessment of bleomycin-induced lung fibrosis by Micro-CT correlates with histological evaluation in mice. Multidiscip. Respir. Med. 2017, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, M.C.; Mano, R.A.; Oliveira, D.H.; Maia, D.S.V.; Silva, W.P.; Savegnago, L.; Lenardão, E.J.; Jacob, R.G. Synthesis, Antimicrobial, and Antioxidant Activities of Chalcogen-Containing Nitrone Derivatives from (R)-citronellal. Medicines 2017, 4, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintans-Júnior, L.; da Rocha, R.F.; Caregnato, F.F.; Moreira, J.C.; da Silva, F.A.; Araújo, A.A.; dos Santos, J.P.; Melo, M.S.; de Sousa, D.P.; Bonjardim, L.R.; et al. Antinociceptive action and redox properties of citronellal, an essential oil present in lemongrass. J. Med. Food 2011, 14, 630–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saibabu, V.; Singh, S.; Ansari, M.A.; Fatima, Z.; Hameed, S. Insights into the intracellular mechanisms of citronellal in Candida albicans: Implications for reactive oxygen species-mediated necrosis, mitochondrial dysfunction, and DNA damage. Rev. Soc. Bras. Med. Trop. 2017, 50, 524–529. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, S.M.; El- Denshary, E.S.; Abdallah, D.M. Geraniol, Alone and in Combination with Pioglitazone, Ameliorates Fructose-Induced Metabolic Syndrome in Rats via the Modulation of Both Inflammatory and Oxidative Stress Status. PLoS ONE 2015, 10, e0117516. [Google Scholar] [CrossRef]
- Gerarduzzi, C.; Di Battista, J.A. Myofibroblast repair mechanisms post-inflammatory response: A fibrotic perspective. Inflamm. Res. 2017, 66, 451–465. [Google Scholar] [CrossRef]
- Sousa, A.M.; Liu, T.; Guevara, O.; Stevens, J.; Fanburg, B.L.; Gaestel, M.; Toksoz, D.; Kayyali, U.S. Smooth muscle alpha-actin expression and myofibroblast differentiation by TGFβ are dependent upon MK2. J. Cell. Biochem. 2007, 100, 1581–1592. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Kim, J.-D.; Ugai, K.; Matsuda, S.; Mikami, H.; Yoshioka, K.; Ikari, J.; Hatano, M.; Fukamizu, A.; Tatsumi, K.; et al. Transcriptomic changes involved in the dedifferentiation of myofibroblasts derived from the lung of a patient with idiopathic pulmonary fibrosis. Mol. Med. Rep. 2020, 22, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- Koga, M.; Kuramochi, M.; Karim, M.R.; Izawa, T.; Kuwamura, M.; Yamate, J. Immunohistochemical characterization of myofibroblasts appearing in isoproterenol-induced rat myocardial fibrosis. J. Vet. Med. Sci. 2019, 81, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.Y.; Duan, J.X.; Yang, H.H.; Sun, C.C.; Zhong, W.J.; Tao, J.H.; Guan, X.X.; Jiang, H.L.; Hammock, B.D.; Hwang, S.H.; et al. COX-2/sEH dual inhibitor PTUPB alleviates bleomycin-induced pulmonary fibrosis in mice via inhibiting senescence. FEBS J. 2020, 287, 1666–1680. [Google Scholar] [CrossRef]
- Phan, S.H. Genesis of the myofibroblast in lung injury and fibrosis. Proc. Am. Thorac. Soc. 2012, 9, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.W.; Herzog, E.L. Regulation and Relevance of Myofibroblast Responses in Idiopathic Pulmonary Fibrosis. Curr. Pathobiol. Rep. 2013, 1, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Tong, M.; Ou, B.; Liu, C.; Hu, C.; Yang, Y. Isorhamnetin protects against bleomycin-induced pulmonary fibrosis by inhibiting endoplasmic reticulum stress and epithelial-mesenchymal transition. Int. J. Mol. Med. 2019, 43, 117–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallée, A.; Lecarpentier, Y. TGF-β in fibrosis by acting as a conductor for contractile properties of myofibroblasts. Cell Biosci. 2019, 9, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-P.; Yu, H.-H.M.; Chiang, E.-R.; Wang, J.-Y.; Chou, P.-H.; Hung, S.-C. Corticosteroid inhibits differentiation of palmar fibromatosis-derived stem cells (FSCs) through downregulation of transforming growth factor-β1 (TGF-β1). PLoS ONE 2018, 13, e0198326. [Google Scholar] [CrossRef]
- Ueshima, E.; Fujimori, M.; Kodama, H.; Felsen, D.; Chen, J.; Durack, J.C.; Solomon, S.B.; Coleman, J.A.; Srimathveeravalli, G. Macrophage-secreted TGF-β1 contributes to fibroblast activation and ureteral stricture after ablation injury. Am. J. Physiol. Ren. Physiol. 2019, 317, F52–F64. [Google Scholar] [CrossRef]
- Rago, F.; Melo, E.M.; Kraemer, L.; Galvão, I.; Cassali, G.D.; Santos, R.A.S.; Russo, R.C.; Teixeira, M.M. Effect of preventive or therapeutic treatment with angiotensin 1–7 in a model of bleomycin-induced lung fibrosis in mice. J. Leukoc. Biol. 2019, 106, 677–686. [Google Scholar] [CrossRef]
- Zhou, Y.; He, Z.; Gao, Y.; Zheng, R.; Zhang, X.; Zhao, L.; Tan, M. Induced Pluripotent Stem Cells Inhibit Bleomycin-Induced Pulmonary Fibrosis in Mice through Suppressing TGF-β1/Smad-Mediated Epithelial to Mesenchymal Transition. Front. Pharm. 2016, 7, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups (n = 6) | Pulmonary Damage Procedure | Oral Administration (Treatment) a |
|---|---|---|
| Sham b | Saline solution | Soybean oil |
| Vehicle c | Bleomycin sulfate (5 mg/kg) | Soybean oil |
| EOCW 50 c | Bleomycin sulfate (5 mg/kg) | 50 mg/kg essential oil of Cymbopogon winterianus |
| EOCW 100 c | Bleomycin sulfate (5 mg/kg) | 100 mg/kg essential oil of Cymbopogon winterianus |
| EOCW 200 c | Bleomycin sulfate (5 mg/kg) | 200 mg/kg essential oil of Cymbopogon winterianus |
| DFC c | Bleomycin sulfate (5 mg/kg) | 1.0 mg/kg Deflazacort |
| Grade of Fibrosis | Modified Scale |
|---|---|
| 0 | Alveolar Septa: No fibrotic burden at the flimsiest small fibers in some alveolar walls Lung structure: Normal lung |
| 1 | Alveolar Septa: Isolated gentle fibrotic changes (septum ≤3× thicker than normal) Lung structure: Alveoli partly enlarged and rarefied, but no fibrotic masses present. |
| 2 | Alveolar Septa: Clearly fibrotic changes (septum >3× thicker than normal) with not-like formation but not connected to each other Lung structure: Alveoli partly enlarged and rarefied, but no fibrotic masses. |
| 3 | Alveolar Septa: Contiguous fibrotic walls (septum >3× thicker than normal) predominantly in whole microscopic field Lung structure: Alveoli partly enlarged and rarefied, but no fibrotic masses. |
| 4 | Alveolar Septa: Variable Lung structure: Single fibrotic masses (≤10% microscopic field) |
| 5 | Alveolar Septa: Variable Lung structure: Confluent fibrotic masses (>10% and ≤50% of microscopic field). Lung structure severely damaged but still preserved. |
| 6 | Alveolar Septa: Variable, most not existent Lung structure: Large contiguous fibrotic masses (>50% of microscopic field). Lung architecture mostly not preserved. |
| 7 | Alveolar Septa: Non-existent Lung structure: Alveoli nearly obliterated with fibrous masses but still up to five air bubbles. |
| 8 | Alveolar Septa: Non-existent Lung structure: Microscopic field with complete obliteration with fibrotic masses. |
| RT (min) a | Compounds b | (%) c | RI d |
|---|---|---|---|
| 6.4 | 3-Hexanone, 2-methyl- | 0.01 | 871 |
| 11.0 | 5-Hepten-2-one, 6-methyl- | 0.03 | 987 |
| 11.1 | β-Myrcene | 0.10 | 990 |
| 12.7 | Limonene | 3.21 | 1027 |
| 13.9 | Melonal | 0.16 | 1053 |
| 15.4 | Terpinolene | 0.05 | 1087 |
| 16.0 | Linalool | 1.17 | 1101 |
| 16.5 | cis-Rose oxide | 0.05 | 1110 |
| 17.2 | trans-Rose oxide | 0.03 | 1127 |
| 18.1 | Isopulegol | 1.55 | 1145 |
| 18.7 | Citronellal | 32.61 | 1159 |
| 19.2 | Isopulegol | 0.10 | 1170 |
| 19.6 | 4-Terpineol | 0.07 | 1178 |
| 19.8 | Carane, 4,5-epoxy-, trans | 0.06 | 1183 |
| 20.2 | α-Terpineol | 0.09 | 1191 |
| 20.3 | cis-4-Decenal | 0.05 | 1194 |
| 20.8 | Decanal | 0.20 | 1206 |
| 22.1 | Citronellol | 14.37 | 1233 |
| 22.5 | Neral | 1.32 | 1242 |
| 23.4 | Geraniol | 22.83 | 1262 |
| 23.8 | Geranial | 1.56 | 1272 |
| 25.9 | Citronellic acid | 0.26 | 1320 |
| 27.4 | Citronellol acetate | 0.93 | 1354 |
| 27.6 | Phenol, 4-allyl-2-methoxy- | 0.94 | 1359 |
| 28.7 | Geranyl acetate | 1.14 | 1384 |
| 29.0 | β-Element | 0.56 | 1392 |
| 30.1 | Caryophyllene | 0.04 | 1419 |
| 31.5 | Humulene | 0.07 | 1454 |
| 32.5 | Naphthalene, 1,2,4a,5,6,8a-hexahydro-4,7-dimethyl-1-(1-methylethyl)- | 0.07 | 1477 |
| 32.7 | Germacrene D | 0.97 | 1482 |
| 33.4 | α-Muurolene | 0.30 | 1501 |
| 33.6 | δ-Guaiene | 0.56 | 1506 |
| 34.0 | α-Amorphene | 0.24 | 1514 |
| 34.4 | β-Cadinene | 1.16 | 1524 |
| 35.4 | o-Menth-8-ene-4-methanol, α,α-dimethyl-1-vinyl-, (1S,2S,4R)-(−)- | 3.64 | 1552 |
| 36.4 | trans-Sesquisabinene hydrate | 1.02 | 1577 |
| 36.7 | Caryophyllene oxide | 0.14 | 1584 |
| 38.1 | Eudesmol | 0.49 | 1621 |
| 38.4 | α-Acorenol | 0.46 | 1629 |
| 38.5 | α-Eudesmol | 1.26 | 1633 |
| 38.9 | α-Muurolol | 1.79 | 1644 |
| 39.0 | Cadinol | 0.45 | 1648 |
| 39.2 | β-Eudesmol | 0.67 | 1652 |
| 39.4 | α-Cadinol | 2.95 | 1657 |
| 41.5 | (Z,E)-Farnesol | 0.04 | 1716 |
| 41.7 | (E,E)-Farnesol | 0.21 | 1723 |
| Body Weight | Sham | Vehicle | EOCW 50 | EOCW 100 | EOCW 200 | DFC |
|---|---|---|---|---|---|---|
| Initial (g) | 174 ± 11.33 | 236.8 ± 20.7 | 213.33 ± 20.81 | 200.25 ± 12.01 | 179.83 ± 9.04 | 174.66 ± 6.31 |
| Final (g) | 183.4 ± 9.55 | 228.6 ± 15.14 | 206.66 ± 20.64 | 195 ± 7.02 | 182.5 ± 8.31 | 172.5 ± 6.22 |
| Body weight gain (%) | 5.17 ± 1.60 a | −3.46 ± 2.32 b | –3.24 ± 1.21 b | –2.63 ± 2.85 b | 1.42 ± 0.55 c | –1.27± 0.48 b,c |
| Hematological Parameters | Sham | Vehicle | EOCW (mg/kg) | DFC | Reference Range * | ||
|---|---|---|---|---|---|---|---|
| 50 | 100 | 200 | |||||
| Total leukocytes (×103/µL) | 4.7 ± 0.6 a | 3.1 ± 1.2 b | 5.0 ± 0.3 a | 4.5 ± 0.6 a | 6.1 ± 1.4 a | 3.6 ± 0.4 b | 2.3−9.9 |
| Red blood cells (×103/µL) | 8.0 ± 1.0 a | 5.8 ± 0.7 b | 6.1 ± 1.7 b | 6.1 ± 0.9 b | 7.6 ± 1.2 a | 9.0 ± 0.4 a | 5.2−8.8 |
| Hematocrit (%) | 51.8 ±1.9 a | 38.4 ± 4.0 b | 40.5 ± 0.70 b | 39.2 ± 1.70 b | 46.0 ± 2.4 a | 52.8 ± 2.9 a | 27.2–48.5 |
| Hemoglobin (g/dL) | 15.7 ± 0.9 | 13.7 ± 1.7 | 13.6 ± 0.9 | 13. 9 ± 0.8 | 14.6 ± 1.0 | 16.0 ± 0.8 | 11.1–17.1 |
| Platelets (×103/µL) | 1.0 ± 0.2 | 1.3 ± 0.7 | 1.0 ± 0.4 | 1.2 ± 0.2 | 0.9 ± 0.1 | 0.8 ± 0.1 | 0.76–1.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tavares, L.A.; Rezende, A.A.; Santos, J.L.; Estevam, C.S.; Silva, A.M.O.; Schneider, J.K.; Cunha, J.L.S.; Droppa-Almeida, D.; Correia-Neto, I.J.; Cardoso, J.C.; et al. Cymbopogon winterianus Essential Oil Attenuates Bleomycin-Induced Pulmonary Fibrosis in a Murine Model. Pharmaceutics 2021, 13, 679. https://doi.org/10.3390/pharmaceutics13050679
Tavares LA, Rezende AA, Santos JL, Estevam CS, Silva AMO, Schneider JK, Cunha JLS, Droppa-Almeida D, Correia-Neto IJ, Cardoso JC, et al. Cymbopogon winterianus Essential Oil Attenuates Bleomycin-Induced Pulmonary Fibrosis in a Murine Model. Pharmaceutics. 2021; 13(5):679. https://doi.org/10.3390/pharmaceutics13050679
Chicago/Turabian StyleTavares, Lívia A., Allan A. Rezende, Jymmys L. Santos, Charles S. Estevam, Ana M. O. Silva, Jaderson K. Schneider, John L. S. Cunha, Daniela Droppa-Almeida, Ivan J. Correia-Neto, Juliana C. Cardoso, and et al. 2021. "Cymbopogon winterianus Essential Oil Attenuates Bleomycin-Induced Pulmonary Fibrosis in a Murine Model" Pharmaceutics 13, no. 5: 679. https://doi.org/10.3390/pharmaceutics13050679