
The Effect of a TLR4 Agonist/Cationic Liposome Adjuvant on Varicella-Zoster Virus Glycoprotein E Vaccine Efficacy: Antigen Presentation, Uptake, and Delivery to Lymph Nodes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Materials
2.3. Preparation of dLOS and Recombinant VZV gE Antigen
2.4. Preparation and Characterization of Cationic Liposomes
2.4.1. Liposome Preparation
2.4.2. Measurement of Particle Size and Zeta Potential of Liposomes
2.4.3. Morphology
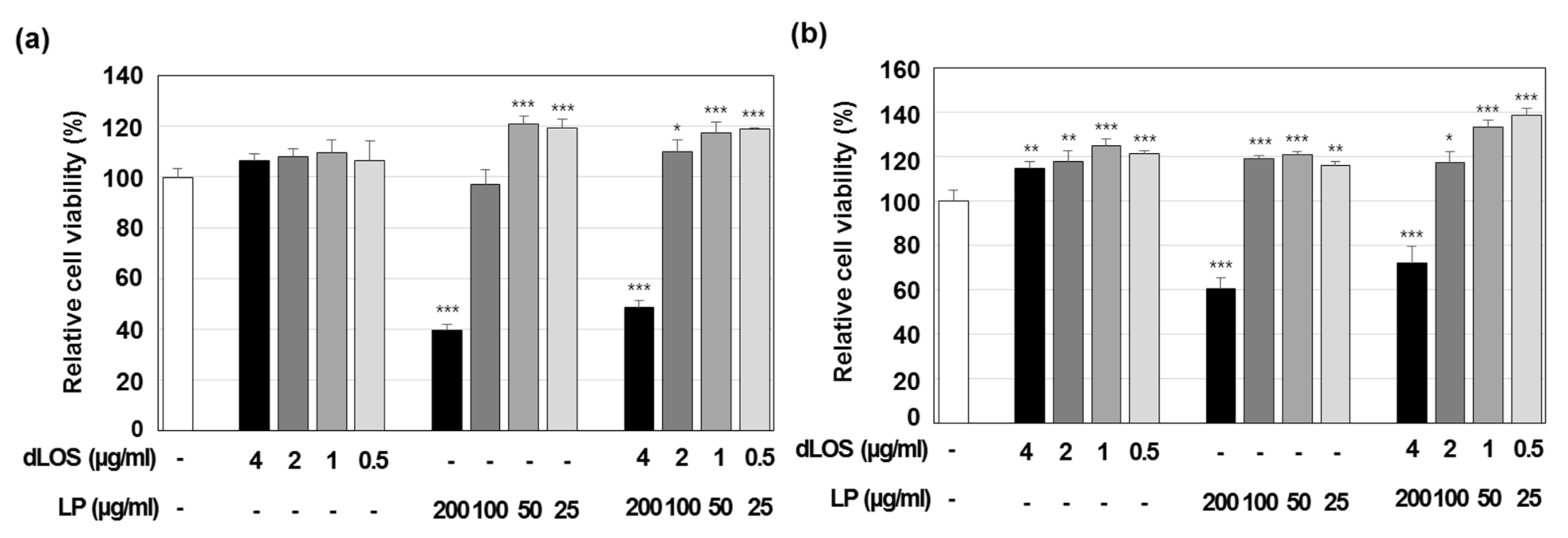
2.4.4. Cytotoxicity Assay
2.5. Measurement of gE Antigen Adsorption on Liposomes
2.6. Determination of Cellular Uptake of gE Antigen
2.6.1. Imaging of Cellular Antigen-Uptake by Confocal Microscopy
2.6.2. Measurement of Cellular Antigen Uptake by Flow Cytometry
2.7. Measurement of Cytokines and Chemokines Secreted from Mouse Immune Cells
2.8. Measurement of Cytokine and Chemokine Secretion at the SOI
2.9. Immune Cell Phenotyping of Mouse Muscle Samples
2.10. In Vivo Imaging Analysis
2.10.1. Fluorescence Imaging Analysis
2.10.2. SPECT/CT Imaging Analysis
2.11. Measurement of In Vivo Cellular Antigen Uptake and Delivery to Lymph Nodes
2.12. Antigen-Presenting Cell Assay
2.13. Statistical Analysis
3. Results
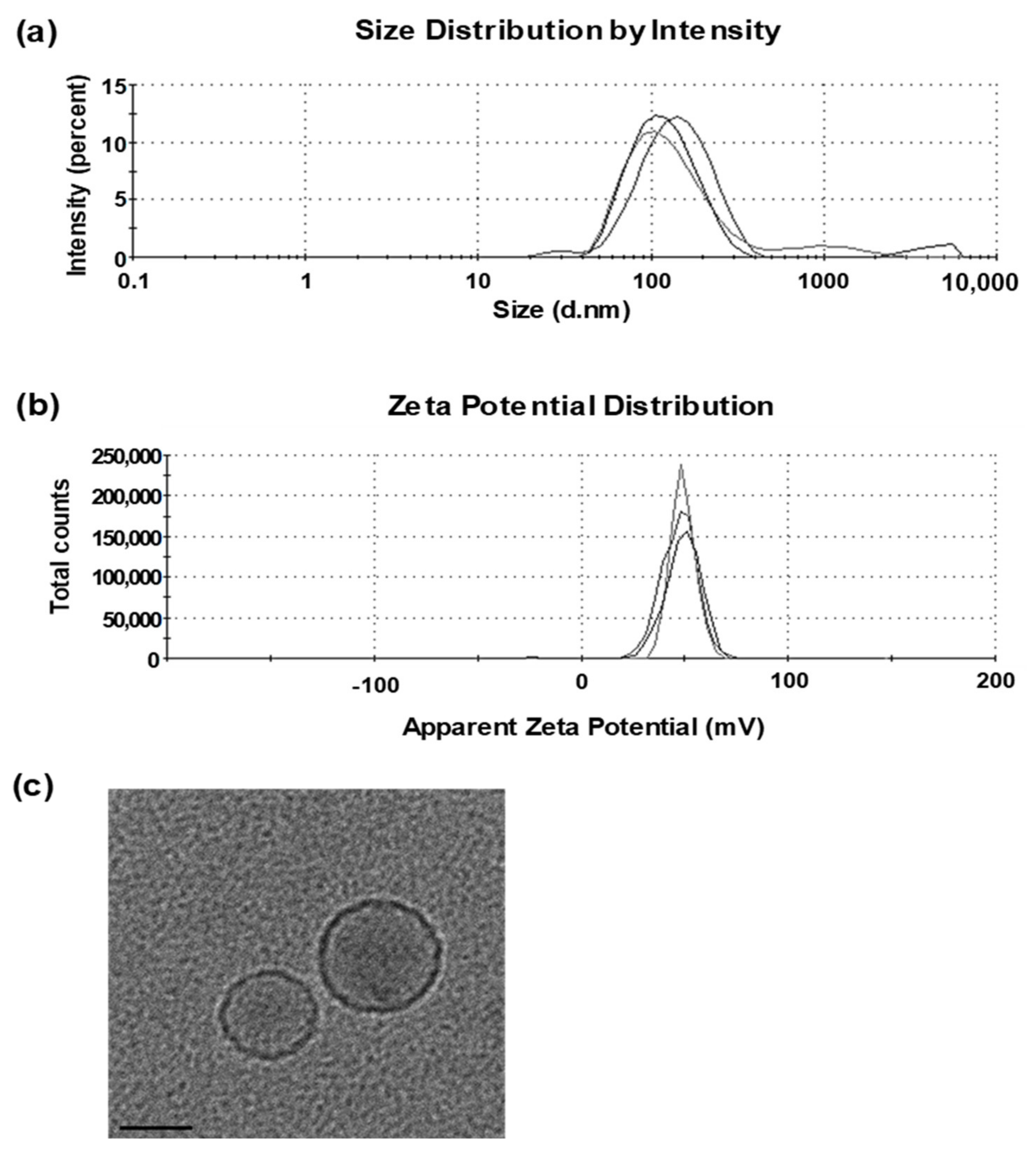
3.1. Characterization of Liposomes
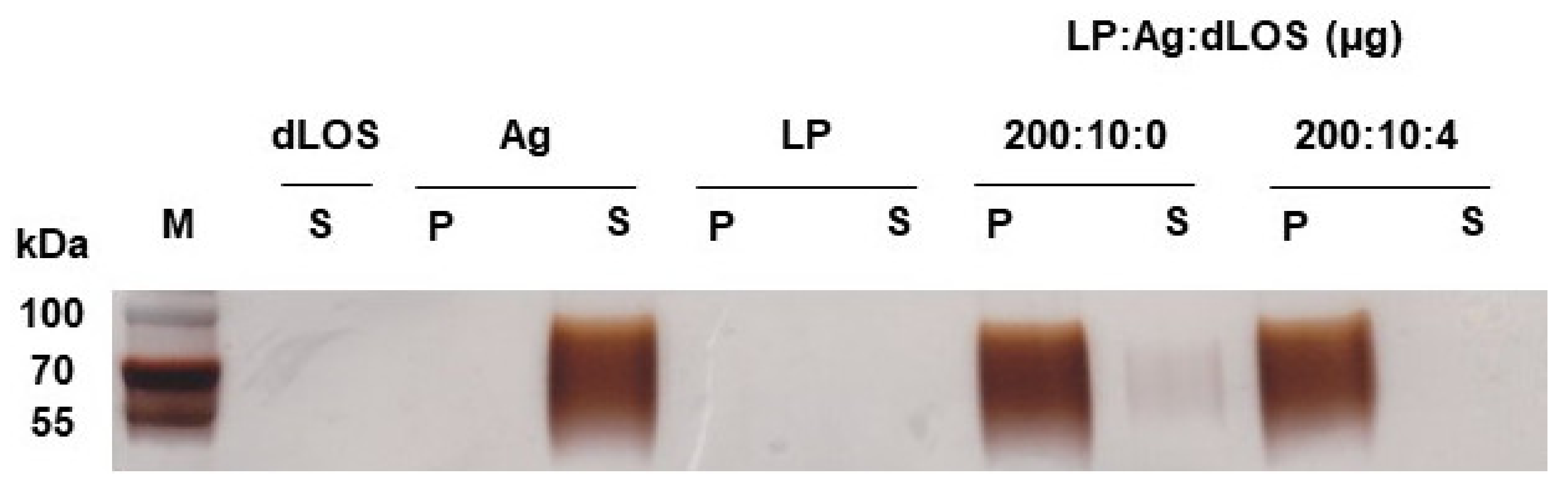
3.2. Adsorption of VZV gE Antigen to Liposomes
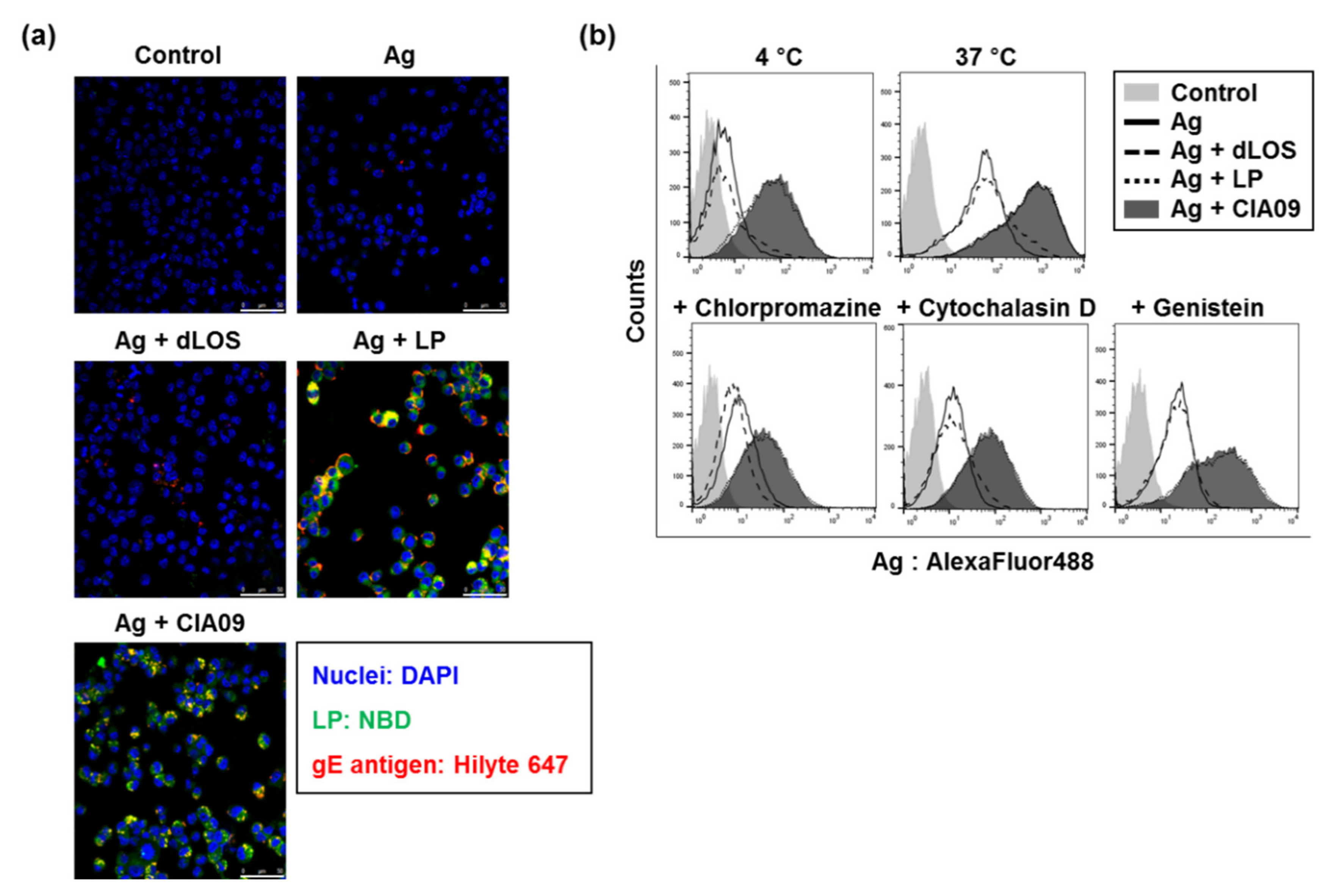
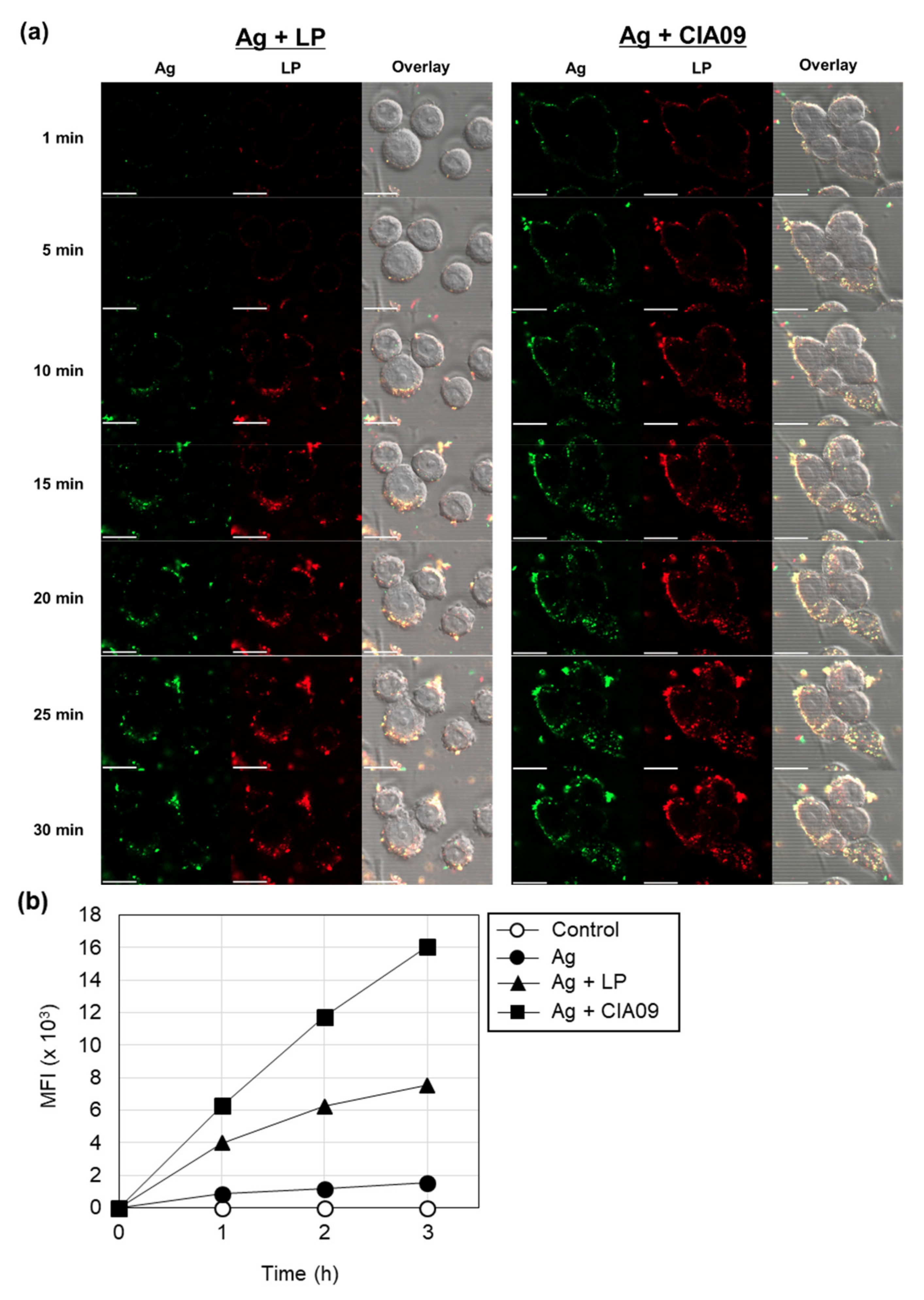
3.3. Enhancement of In Vitro Cellular Uptake of VZV gE Antigen by Liposomes
3.3.1. Liposome-Mediated Cellular Uptake of gE Antigen
3.3.2. Cooperative Effects of Liposomes and dLOS on the Cellular Uptake of gE antigen
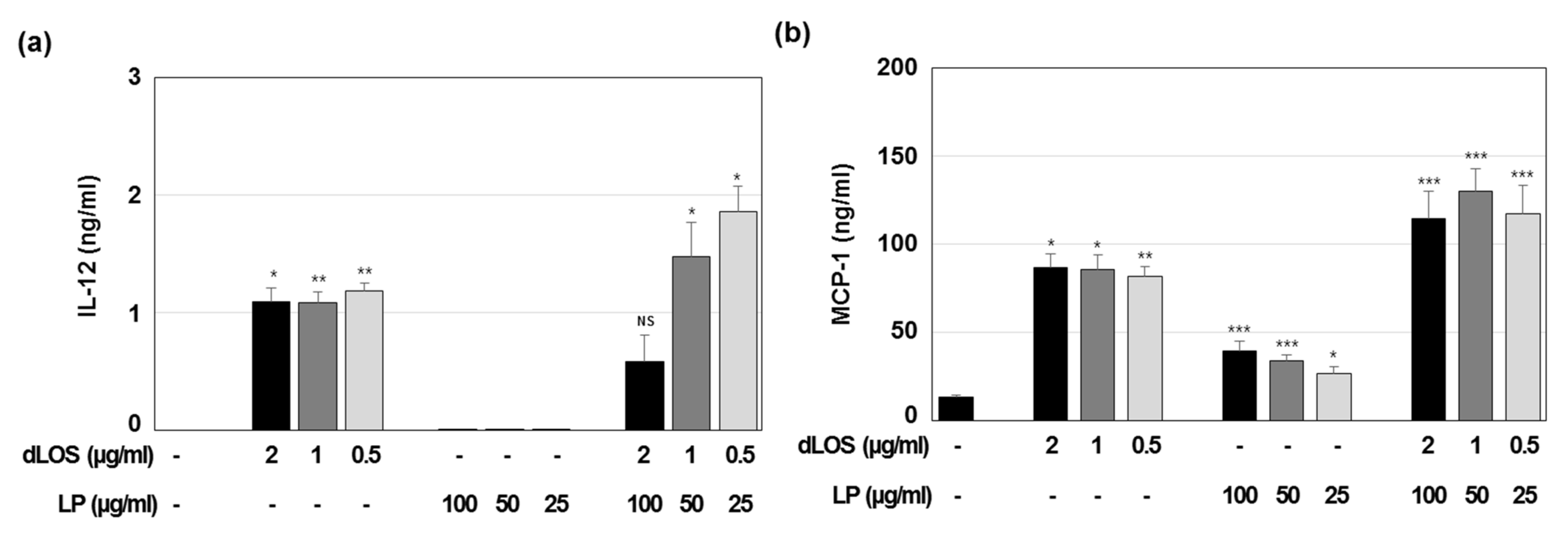
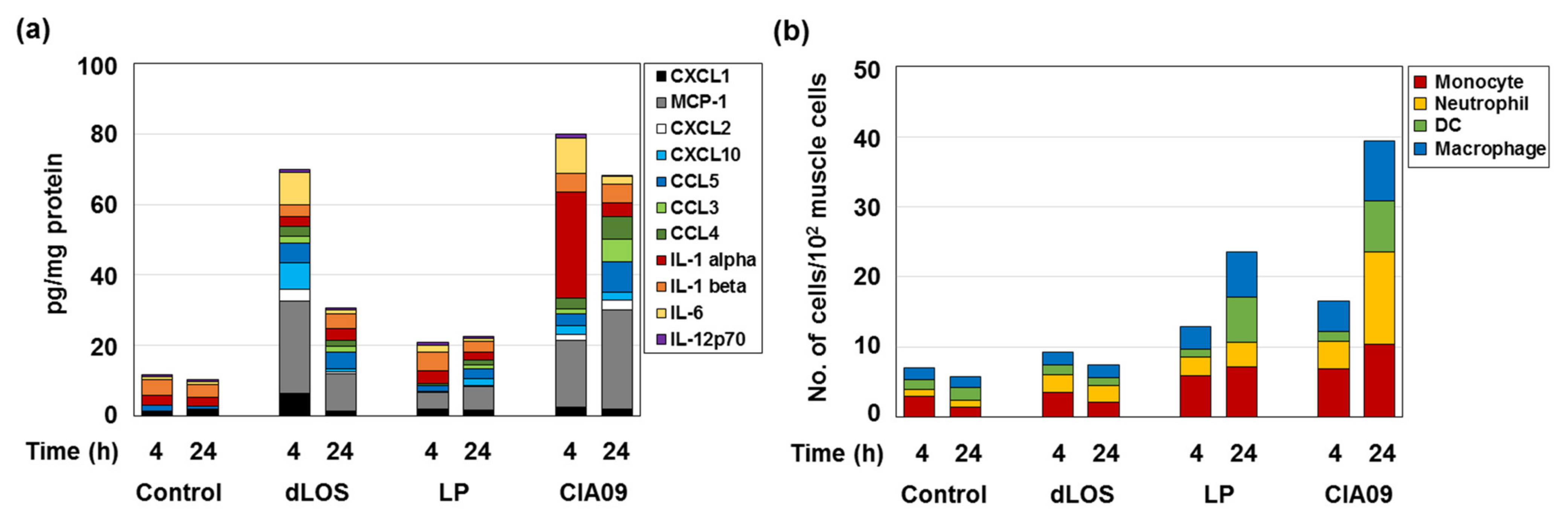
3.4. Immune Cell Activation and Recruitment by CIA09
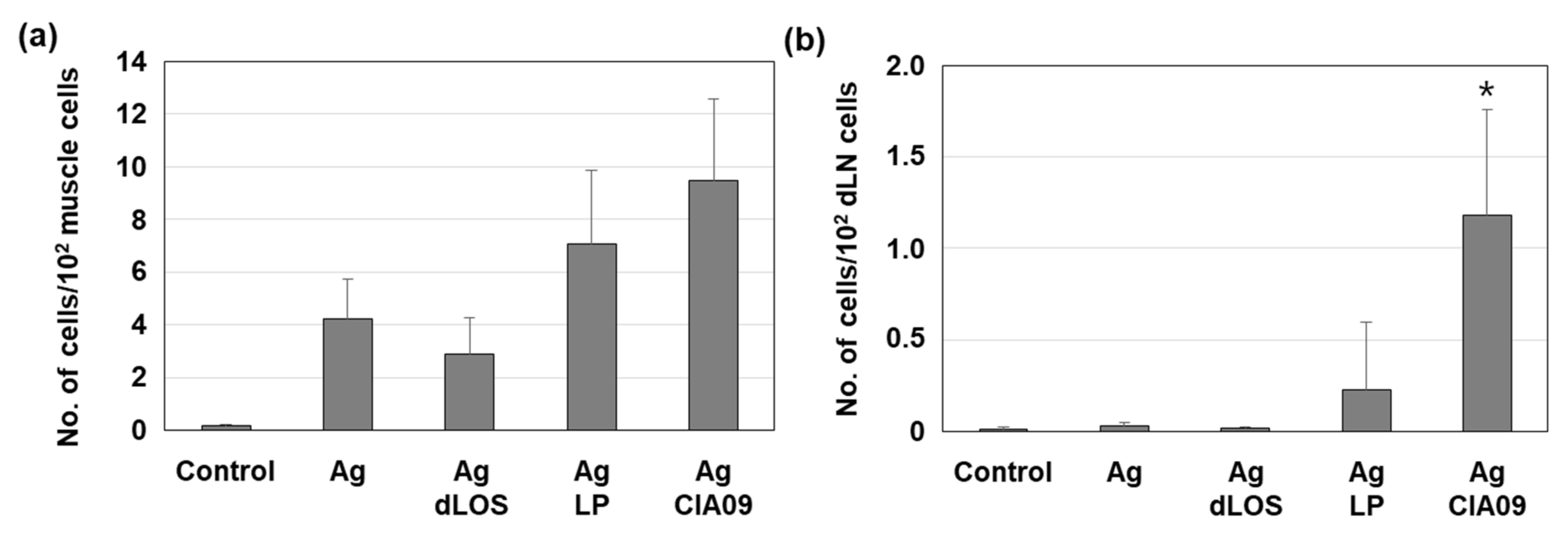
3.5. Increase in Antigen Uptake at the Site of Injection by CIA09
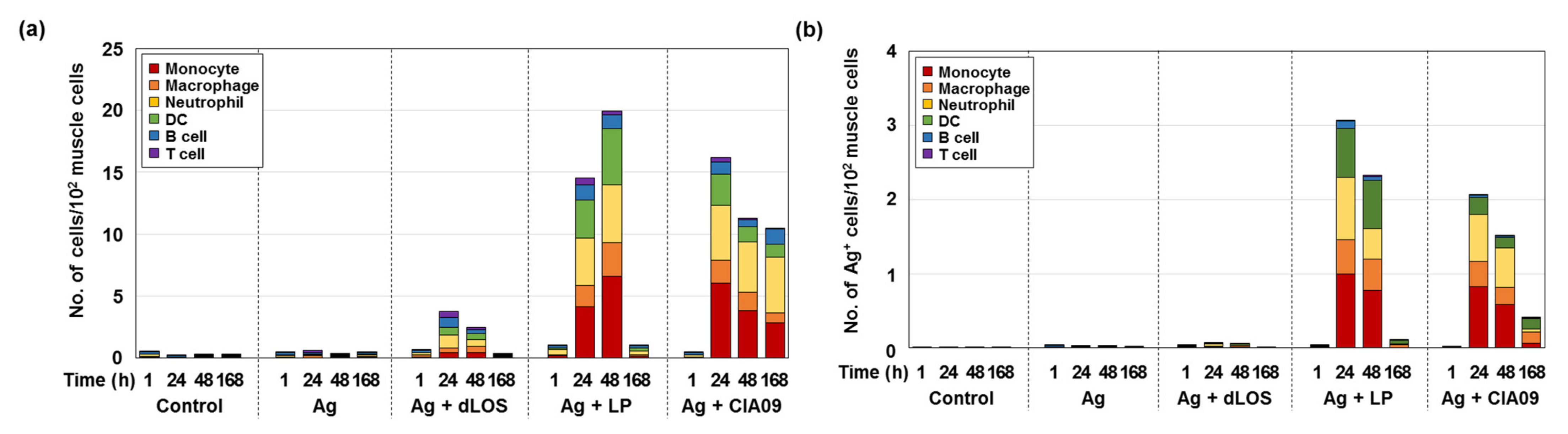
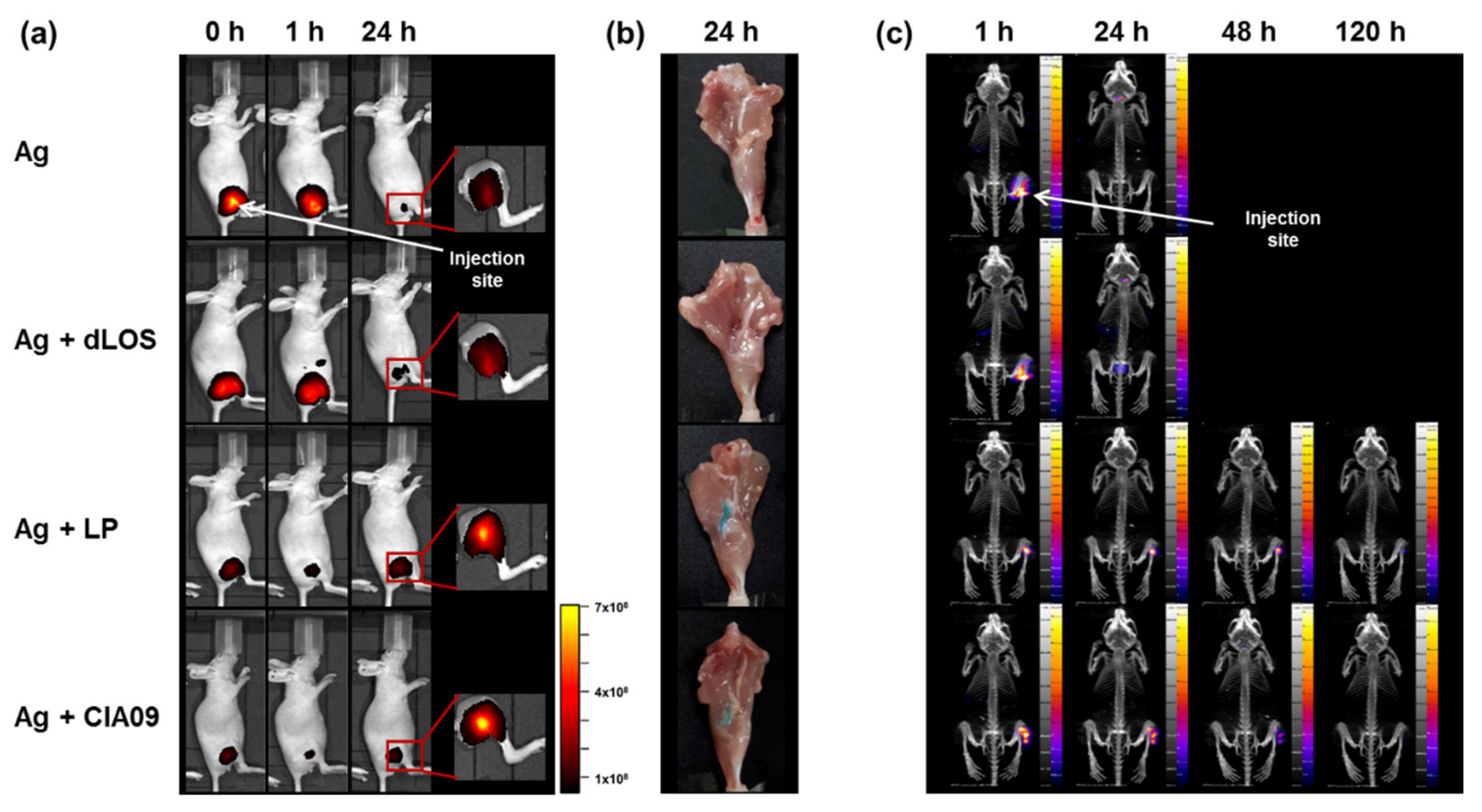
3.6. Increased In Vivo Stability of Liposome-Associated gE Antigen
3.7. Efficient Delivery to Lymph Nodes of CIA09-Adjuvanted gE Antigen
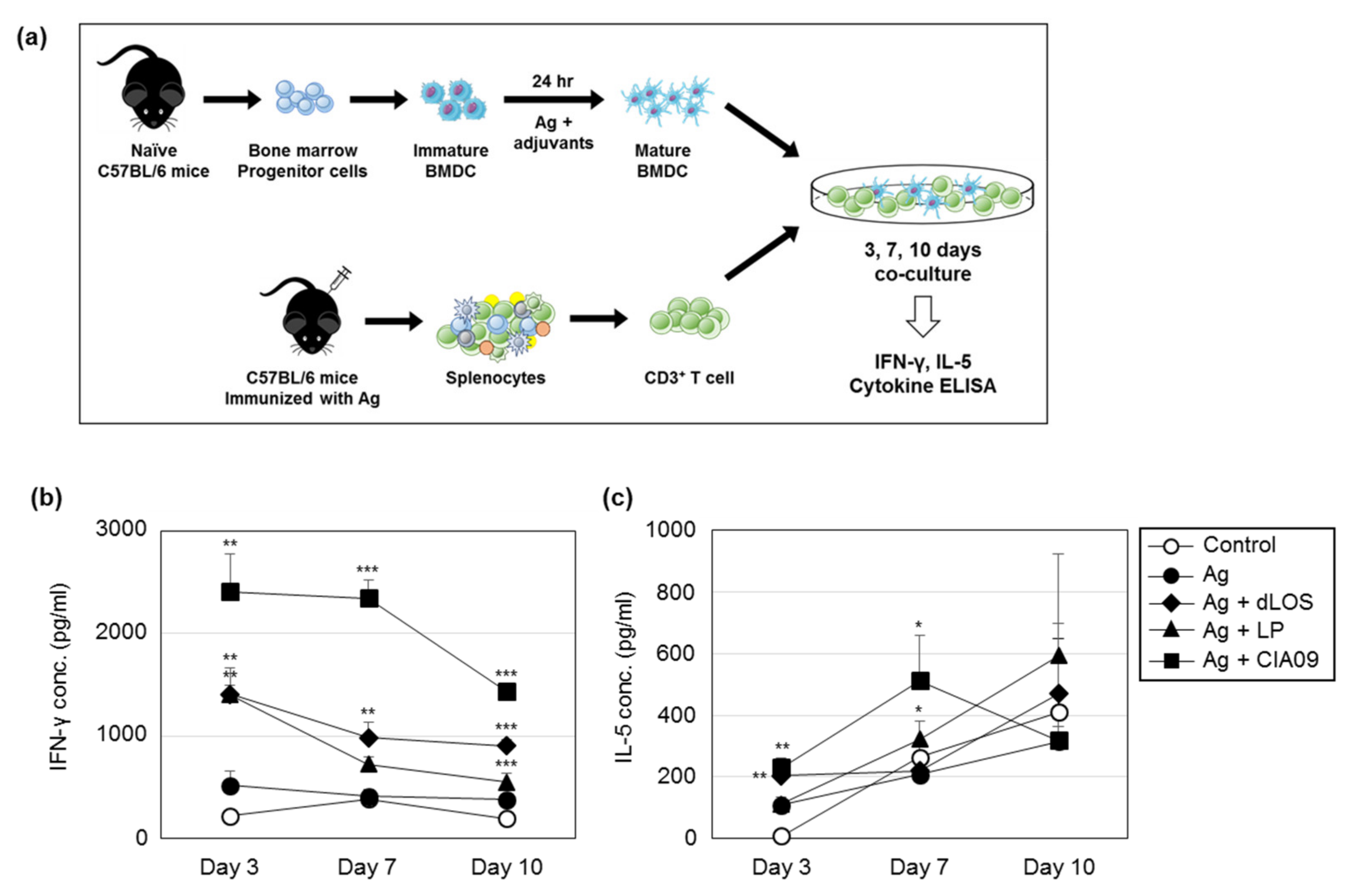
3.8. Efficient Antigen Presentation to T Cells by DCs Pulsed with CIA09-Associated gE
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guy, B. The perfect mix: Recent progress in adjuvant research. Nat. Rev. Microbiol. 2007, 5, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- Di Pasquale, A.; Preiss, S.; Da Silva, F.T.; Garcon, N. Vaccine adjuvants: From 1920 to 2015 and beyond. Vaccines 2015, 3, 320–343. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.M.; Simon, J.K.; Baker, J.R. Applications of nanotechnology for immunology. Nat. Rev. Immunol. 2013, 13, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine adjuvants: Understanding the structure and mechanism of adjuvanticity. Vaccine 2019, 37, 3167–3178. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef] [Green Version]
- De Gregorio, E.; D’Oro, U.; Wack, A. Immunology of TLR-independent vaccine adjuvants. Curr. Opin. Immunol. 2009, 21, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Toussi, D.N.; Massari, P. Immune adjuvant effect of molecularly-defined toll-like receptor ligands. Vaccines 2014, 2, 323–353. [Google Scholar] [CrossRef]
- Bonam, S.R.; Partidos, C.D.; Halmuthur, S.K.M.; Muller, S. An overview of novel adjuvants designed for improving vaccine efficacy. Trends Pharmacol. Sci. 2017, 38, 771–793. [Google Scholar] [CrossRef]
- Bonam, S.R.; Bhunia, D.; Muller, S.; Nerella, S.G.; Alvala, M.; Mahabalarao, S.K.H. Novel trisaccharide based phospholipids as immunomodulators. Int. Immunopharmacol. 2019, 74, e105684. [Google Scholar] [CrossRef] [PubMed]
- Schijns, V.; Fernandez-Tejada, A.; Barjaktarovic, Z.; Bouzalas, I.; Brimnes, J.; Chernysh, S.; Gizurarson, S.; Gursel, I.; Jakopin, Z.; Lawrenz, M.; et al. Modulation of immune responses using adjuvants to facilitate therapeutic vaccination. Immunol. Rev. 2020, 296, 169–190. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; Valiante, N.M. Recent advances in the discovery and delivery of vaccine adjuvants. Nat. Rev. Drug Discov. 2003, 2, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Mount, A.; Koernig, S.; Silva, A.; Drane, D.; Maraskovsky, E.; Morelli, A.B. Combination of adjuvants: The future of vaccine design. Expert Rev. Vaccines 2013, 12, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.T.; Foged, C.; Korsholm, K.S.; Rades, T.; Christensen, D. Liposome-based adjuvants for subunit vaccines: Formulation strategies for subunit antigens and immunostimulators. Pharmaceutics 2016, 8, 7. [Google Scholar] [CrossRef]
- Alving, C.R.; Beck, Z.; Matyas, G.R.; Rao, M. Liposomal adjuvants for human vaccines. Expert Opin. Drug Deliv. 2016, 13, 807–816. [Google Scholar] [CrossRef]
- Didierlaurent, A.M.; Laupeze, B.; Di Pasquale, A.; Hergli, N.; Collignon, C.; Garcon, N. Adjuvant system AS01: Helping to overcome the challenges of modern vaccines. Expert Rev. Vaccines 2017, 16, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisini, R.; Poerio, N.; Mariotti, S.; De Santis, F.; Fraziano, M. The multirole of liposomes in therapy and prevention of infectious diseases. Front. Immunol. 2018, 9, 155. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chen, M.N.; Wang, T. Liposomes used as a vaccine adjuvant-delivery system: From basics to clinical immunization. J. Control Release 2019, 303, 130–150. [Google Scholar] [CrossRef] [PubMed]
- Henriksen-Lacey, M.; Christensen, D.; Bramwell, V.W.; Lindenstrom, T.; Agger, E.M.; Andersen, P.; Perrie, Y. Liposomal cationic charge and antigen adsorption are important properties for the efficient deposition of antigen at the injection site and ability of the vaccine to induce a CMI response. J. Control Release 2010, 145, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhuang, Y.; Xie, X.; Wang, C.; Wang, F.; Zhou, D.; Zeng, J.; Cai, L. The role of surface charge density in cationic liposome-promoted dendritic cell maturation and vaccine-induced immune responses. Nanoscale 2011, 3, 2307–2314. [Google Scholar] [CrossRef] [PubMed]
- Perrie, Y.; Kastner, E.; Kaur, R.; Wilkinson, A.; Ingham, A.J. A case-study investigating the physicochemical characteristics that dictate the function of a liposomal adjuvant. Hum. Vaccin. Immunother. 2013, 9, 1374–1381. [Google Scholar] [CrossRef] [Green Version]
- Vangasseri, D.P.; Cui, Z.R.; Chen, W.H.; Hokey, D.A.; Falo, L.D.; Huang, L. Immunostimulation of dendritic cells by cationic liposomes. Mol. Membr. Biol. 2006, 23, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Korsholm, K.S.; Agger, E.M.; Foged, C.; Christensen, D.; Dietrich, J.; Andersen, C.S.; Geisler, C.; Andersen, P. The adjuvant mechanism of cationic dimethyldioctadecylammonium liposomes. Immunology 2007, 121, 216–226. [Google Scholar] [CrossRef]
- Christensen, D.; Korsholm, K.S.; Andersen, P.; Agger, E.M. Cationic liposomes as vaccine adjuvants. Expert Rev. Vaccines 2011, 10, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, P.; Zhuang, Y.; Li, P.; Jiang, B.L.; Pan, H.; Liu, L.L.; Cai, L.T.; Ma, Y.F. Lymphatic-targeted cationic liposomes: A robust vaccine adjuvant for promoting long-term immunological memory. Vaccine 2014, 32, 5475–5483. [Google Scholar] [CrossRef] [PubMed]
- Perrie, Y.; Frederik, P.M.; Gregoriadis, G. Liposome-mediated DNA vaccination: The effect of vesicle composition. Vaccine 2001, 19, 3301–3310. [Google Scholar] [CrossRef]
- Tada, R.; Suzuki, H.; Takahashi, S.; Negishi, Y.; Kiyono, H.; Kunisawa, J.; Aramaki, Y. Nasal vaccination with pneumococcal surface protein A in combination with cationic liposomes consisting of DOTAP and DC-chol confers antigen-mediated protective immunity against Streptococcus pneumoniae infections in mice. Int. Immunopharmacol. 2018, 61, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Mansury, D.; Ghazvini, K.; Jamehdar, S.A.; Badiee, A.; Tafaghodi, M.; Nikpooe, A.R.; Amini, Y.; Jaafari, M.R. Increasing cellular immune response in liposomal formulations of DOTAP encapsulated by fusion protein Hspx, PPE44, and Esxv, as a potential tuberculosis vaccine candidate. Rep. Biochem. Mol. Biol. 2019, 7, 156–166. [Google Scholar] [PubMed]
- Yan, W.; Chen, W.; Huang, L. Mechanism of adjuvant activity of cationic liposome: Phosphorylation of a MAP kinase, ERK and induction of chemokines. Mol. Immunol. 2007, 44, 3672–3681. [Google Scholar] [CrossRef]
- Christensen, D.; Agger, E.M.; Andreasen, L.V.; Kirby, D.; Andersen, P.; Perrie, Y. Liposome-based cationic adjuvant formulations (CAF): Past, present, and future. J. Liposome Res. 2009, 19, 2–11. [Google Scholar] [CrossRef]
- Agger, E.M.; Rosenkrands, I.; Hansen, J.; Brahimi, K.; Vandahl, B.S.; Aagaard, C.; Werninghaus, K.; Kirschning, C.; Lang, R.; Christensen, D.; et al. Cationic liposomes formulated with synthetic mycobacterial cordfactor (CAF01): A versatile adjuvant for vaccines with different immunological requirements. PLoS ONE 2008, 3, e3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriksen-Lacey, M.; Bramwell, V.W.; Christensen, D.; Agger, E.M.; Andersen, P.; Perrie, Y. Liposomes based on dimethyldioctadecylammonium promote a depot effect and enhance immunogenicity of soluble antigen. J. Control Release 2010, 142, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Davidsen, J.; Rosenkrands, I.; Christensen, D.; Vangala, A.; Kirby, D.; Perrie, Y.; Agger, E.M.; Andersen, P. Characterization of cationic liposomes based on dimethyldioctadecylammonium and synthetic cord factor from M. tuberculosis (trehalose 6,6′-dibehenate)—A novel adjuvant inducing both strong CMI and antibody responses. Biochim. Biophys. Acta 2005, 1718, 22–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aagaard, C.; Hoang, T.; Dietrich, J.; Cardona, P.J.; Izzo, A.; Dolganov, G.; Schoolnik, G.K.; Cassidy, J.P.; Billeskov, R.; Andersen, P. A multistage tuberculosis vaccine that confers efficient protection before and after exposure. Nat. Med. 2011, 17, 189–U224. [Google Scholar] [CrossRef] [PubMed]
- Van Dissel, J.T.; Joosten, S.A.; Hoff, S.T.; Soonawala, D.; Prins, C.; Hokey, D.A.; O’Dee, D.M.; Graves, A.; Thierry-Carstensen, B.; Andreasen, L.V.; et al. A novel liposomal adjuvant system, CAF01, promotes long-lived Mycobacterium tuberculosis-specific T-cell responses in human. Vaccine 2014, 32, 7098–7107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.J.; Ahn, B.Y.; Lee, N.G.; Lee, D.H.; Kim, D.S. A combination of E. coli DNA fragments and modified lipopolysaccharides as a cancer immunotherapy. Vaccine 2006, 24, 5862–5871. [Google Scholar] [CrossRef]
- Han, J.E.; Wui, S.R.; Kim, K.S.; Cho, Y.J.; Cho, W.J.; Lee, N.G. Characterization of the structure and immunostimulatory activity of a vaccine adjuvant, de-O-acylated lipooligosaccharide. PLoS ONE 2014, 9, e85838. [Google Scholar] [CrossRef] [Green Version]
- Wui, S.R.; Kim, H.K.; Han, J.E.; Kim, J.M.; Kim, Y.H.; Chun, J.H.; Cho, Y.J.; Lee, N.G. A combination of the TLR4 agonist CIA05 and alum promotes the immune responses to Bacillus anthracis protective antigen in mice. Int. Immunopharmacol. 2011, 11, 1195–1204. [Google Scholar] [CrossRef]
- Han, J.E.; Wui, S.R.; Park, S.A.; Lee, N.G.; Kim, K.S.; Cho, Y.J.; Kim, H.J.; Kim, H.J. Comparison of the immune responses to the CIA06-adjuvanted human papillomavirus L1 VLP vaccine with those against the licensed HPV vaccine Cervarix™ in mice. Vaccine 2012, 30, 4127–4134. [Google Scholar] [CrossRef]
- Ryu, J.I.; Park, S.A.; Wui, S.R.; Ko, A.; Han, J.E.; Choi, J.A.; Song, M.K.; Kim, K.S.; Cho, Y.J.; Lee, N.G. A de-O-acylated lipooligosaccharide-based adjuvant system promotes antibody and Th1-type immune responses to H1N1 pandemic influenza vaccine in mice. Biomed. Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Ryu, J.I.; Wui, S.R.; Ko, A.; Do, H.T.T.; Lee, Y.J.; Kim, H.J.; Rhee, I.; Park, S.A.; Kim, K.S.; Cho, Y.J.; et al. Increased immunogenicity and protective efficacy of a P. aeruginosa vaccine in mice using an alum and de-O-acylated lipooligosaccharide adjuvant system. J. Microbiol. Biotechnol. 2017, 27, 1539–1548. [Google Scholar] [CrossRef] [PubMed]
- Ko, A.; Wui, S.R.; Ryu, J.I.; Do, H.T.T.; Lee, Y.J.; Lim, S.J.; Rhee, I.; Jung, D.I.; Park, J.A.; Choi, J.A.; et al. Comparison of the adjuvanticity of two adjuvant formulations containing de-O-acylated lipooligosaccharide on Japanese encephalitis vaccine in mice. Arch. Pharm. Res. 2018, 41, 219–228. [Google Scholar] [CrossRef]
- Ko, A.; Wui, S.R.; Ryu, J.I.; Lee, Y.J.; Hien, D.T.T.; Rhee, I.; Shin, S.J.; Park, S.A.; Kim, K.S.; Cho, Y.J.; et al. Potentiation of Th1-type immune responses to Mycobacterium tuberculosis antigens in mice by cationic liposomes combined with de-O-acylated lipooligosaccharide. J. Microbiol. Biotechnol. 2018, 28, 136–144. [Google Scholar] [CrossRef]
- Wui, S.R.; Kim, K.S.; Ryu, J.I.; Ko, A.; Do, H.T.T.; Lee, Y.J.; Kim, H.J.; Lim, S.J.; Park, S.A.; Cho, Y.J.; et al. Efficient induction of cell-mediated immunity to varicella-zoster virus glycoprotein E co-lyophilized with a cationic liposome-based adjuvant in mice. Vaccine 2019, 37, 2131–2141. [Google Scholar] [CrossRef]
- Lee, C.H.; Tsai, C.M. Quantification of bacterial lipopolysaccharides by the purpald assay: Measuring formaldehyde generated from 2-keto-3-deoxyoctonate and heptose at the inner core by periodate oxidation. Anal. Biochem. 1999, 267, 161–168. [Google Scholar] [CrossRef]
- Guermonprez, P.; Valladeau, J.; Zitvogel, L.; Thery, C.; Amigorena, S. Antigen presentation and T cell stimulation by dendritic cells. Annu. Rev. Immunol. 2002, 20, 621–667. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Lindblad, E.B. Aluminium compounds for use in vaccines. Immunol. Cell Biol. 2004, 82, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Mastelic, B.; Ahmed, S.; Egan, W.M.; Del Giudice, G.; Golding, H.; Gust, I.; Neels, P.; Reed, S.G.; Sheets, R.L.; Siegrist, C.A.; et al. Mode of action of adjuvants: Implications for vaccine safety and design. Biologicals 2010, 38, 594–601. [Google Scholar] [CrossRef]
- Batista-Duharte, A.; Lindblad, E.B.; Oviedo-Orta, E. Progress in understanding adjuvant immunotoxicity mechanisms. Toxicol. Lett. 2011, 203, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Hauguel, T.M.; Hackett, C.J. Rationally-designed vaccine adjuvants: Separating efficacy from toxicity. Front. Biosci. 2008, 13, 2806–2813. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wui, S.R.; Ko, A.; Ryu, J.I.; Sim, E.; Lim, S.J.; Park, S.A.; Kim, K.S.; Kim, H.; Youn, H.; Lee, N.G. The Effect of a TLR4 Agonist/Cationic Liposome Adjuvant on Varicella-Zoster Virus Glycoprotein E Vaccine Efficacy: Antigen Presentation, Uptake, and Delivery to Lymph Nodes. Pharmaceutics 2021, 13, 390. https://doi.org/10.3390/pharmaceutics13030390
Wui SR, Ko A, Ryu JI, Sim E, Lim SJ, Park SA, Kim KS, Kim H, Youn H, Lee NG. The Effect of a TLR4 Agonist/Cationic Liposome Adjuvant on Varicella-Zoster Virus Glycoprotein E Vaccine Efficacy: Antigen Presentation, Uptake, and Delivery to Lymph Nodes. Pharmaceutics. 2021; 13(3):390. https://doi.org/10.3390/pharmaceutics13030390
Chicago/Turabian StyleWui, Seo Ri, Ara Ko, Ji In Ryu, Eojin Sim, Soo Jeong Lim, Shin Ae Park, Kwang Sung Kim, Ha Kim, Hyewon Youn, and Na Gyong Lee. 2021. "The Effect of a TLR4 Agonist/Cationic Liposome Adjuvant on Varicella-Zoster Virus Glycoprotein E Vaccine Efficacy: Antigen Presentation, Uptake, and Delivery to Lymph Nodes" Pharmaceutics 13, no. 3: 390. https://doi.org/10.3390/pharmaceutics13030390