
Viral-Vector-Delivered Anti-Angiogenic Therapies to the Eye


Abstract
:1. Introduction
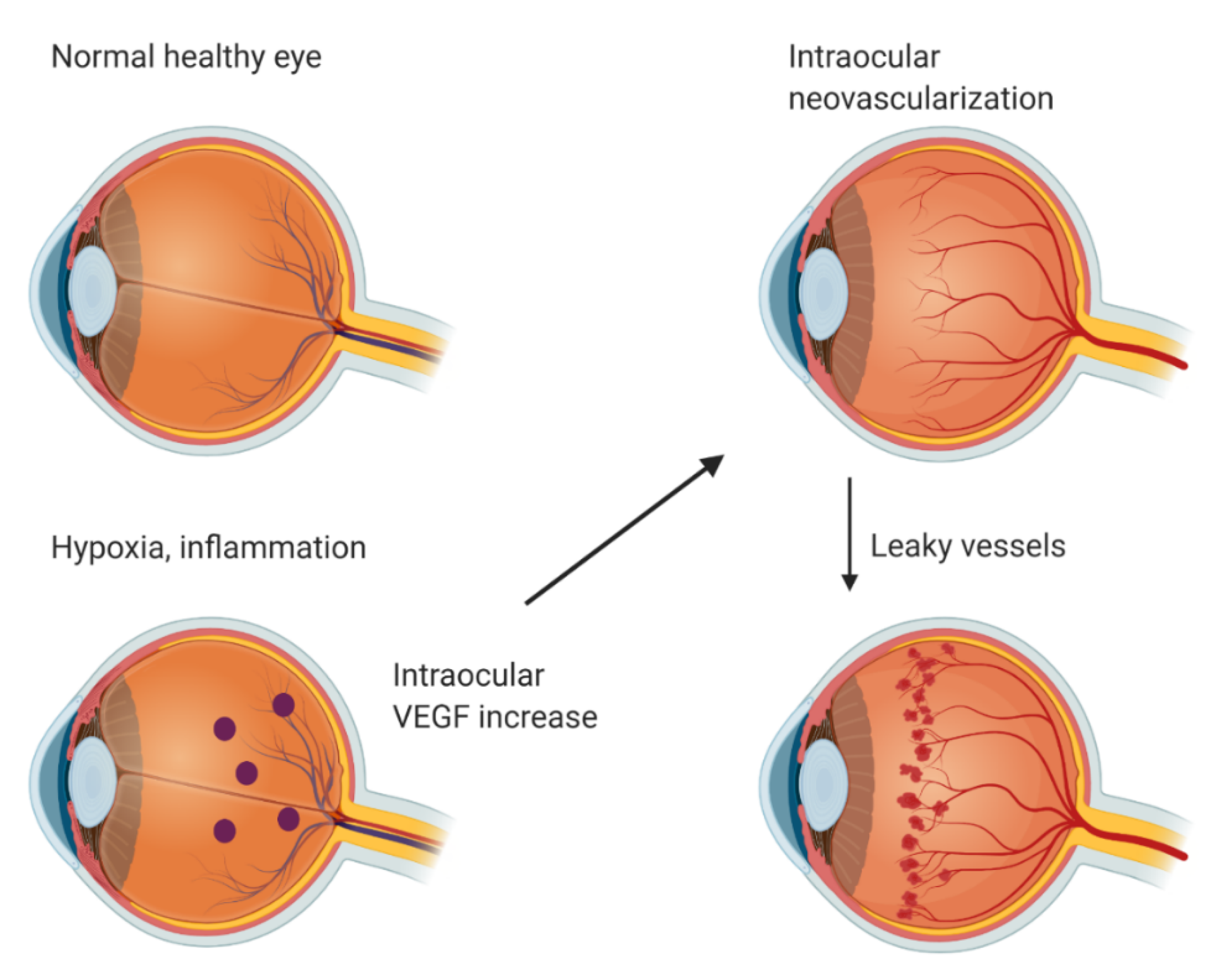
2. Angiogenesis
3. Routes of Administration for Ocular Gene Therapy
4. Gene Therapy and Viral Vectors
5. Preclinical Anti-Angiogenic Studies
5.1. VEGF-Blocking Agents
5.2. Other Pro-Angiogenic Factors
5.3. Preclinical Studies with Genome Editing
5.4. Preclinical Studies with Multigenic Viral Vectors
5.5. Preclinical Studies of Corneal Neovascularization
5.6. Preclinical Studies of Retinopathy of Prematurity
6. Gene Therapy Clinical Trials of Neovascular Ocular Diseases
6.1. AAVCAGsCD59
6.2. ADVM-022
6.3. RGX-314
6.4. Completed Clinical Trials for Ocular Neovascularization
6.5. sFLT-1
6.6. RetinoStat
6.7. PEDF
7. Discussion
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, C.H.; Wang, Z.; Sun, Y.; Chen, J. Animal models of ocular angiogenesis: From development to pathologies. FASEB J. 2017, 31, 4665–4681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.; Ryan, A.M.; Rohan, R.; Amano, S.; Agular, S.; Miller, J.W.; Adamis, A.P. Constitutive Expression of VEGF, VEGFR-1, and VEGFR-2 in Normal Eyes. Investig. Ophthalmol. 2016, 40, 2115–2121. [Google Scholar]
- del Amo, E.M.; Urtti, A. Current and future ophthalmic drug delivery systems. A shift to the posterior segment. Drug Discov. Today 2008, 13, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Wykoff, C.C.; Clark, W.L.; Nielsen, J.S.; Brill, J.V.; Greene, L.S.; Heggen, C.L. Optimizing Anti-VEGF Treatment Outcomes for Patients with Neovascular Age-Related Macular Degeneration. J. Manag. Care Spec. Pharm. 2018, 24, S3–S15. [Google Scholar] [CrossRef] [Green Version]
- Ylä-Herttuala, S.; Bridges, C.; Katz, M.G.; Korpisalo, P. Angiogenic gene therapy in cardiovascular diseases: Dream or vision? Eur. Heart J. 2017, 38, 1365–1371. [Google Scholar]
- Kalesnykas, G.; Kokki, E.; Alasaarela, L.; Lesch, H.P.; Tuulos, T.; Kinnunen, K.; Uusitalo, H.; Airenne, K.; Yla-Herttuala, S. Comparative Study of Adeno-associated Virus, Adenovirus, Bacu lovirus and Lentivirus Vectors for Gene Therapy of the Eyes. Curr. Gene Ther. 2017, 17, 235–247. [Google Scholar] [CrossRef]
- Solinís, M.Á.; Del Pozo-Rodríguez, A.; Apaolaza, P.S.; Rodríguez-Gascón, A. Treatment of ocular disorders by gene therapy. Eur. J. Pharm. Biopharm. 2015, 95, 331–342. [Google Scholar] [CrossRef]
- Ameri, H. Prospect of retinal gene therapy following commercialization of voretigene neparvovec-rzyl for retinal dystrophy mediated by RPE65 mutation. J. Curr. Ophthalmol. 2018, 30, 1–2. [Google Scholar] [CrossRef]
- Hoeben, A.; Landuyt, B.; Highley, M.S.; Wildiers, H.; Van Oosterom, A.T.; De Bruijn, E.A. Vascular endothelial growth factor and angiogenesis. Pharmacol. Rev. 2004, 56, 549–580. [Google Scholar] [CrossRef]
- Ylä-Herttuala, S.; Rissanen, T.T.; Vajanto, I.; Hartikainen, J. Vascular Endothelial Growth Factors. Biology and Current Status of Clinical Applications in Cardiovascular Medicine. J. Am. Coll. Cardiol. 2007, 49, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Vuorio, T.; Jauhiainen, S.; Ylä-Herttuala, S. Pro- and anti-angiogenic therapy and atherosclerosis with special emphasis on vascular endothelial growth factors. Expert Opin. Biol. Ther. 2012, 12, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sene, A.; Chin-Yee, D.; Apte, R.S. Seeing through VEGF: Innate and adaptive immunity in pathological angiogenesis in the eye. Trends Mol. Med. 2015, 21, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Zamil, W.M.; Yassin, S.A. Recent developments in age-related macular degeneration: A review. Clin. Interv. Aging 2017, 12, 1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaughlin, M.D.; Hwang, J.C. Trends in Vitreoretinal Procedures for Medicare Beneficiaries, 2000 to 2014. Ophthalmology 2017, 124, 667–673. [Google Scholar] [CrossRef]
- Deschler, E.K.; Sun, J.K.; Silva, P.S. Side-effects and complications of laser treatment in diabetic retinal disease. Semin. Ophthalmol. 2014, 29, 290–300. [Google Scholar] [CrossRef]
- Mason, J.O.; Yunker, J.J.; Vail, R.; Mcgwin, G. Intravitreal bevacizumab (Avastin) prevention of panretinal photocoagulation-induced complications in patients with severe proliferative diabetic retinopathy. Retina 2008, 28, 1319–1324. [Google Scholar] [CrossRef] [Green Version]
- Fijalkowski, N.; Moshfeghi, D.M. New Laser Technologies for Diabetic Retinopathy. Curr. Ophthalmol. Rep. 2013, 1, 134–143. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Singh, R.P. The role of anti-vascular endothelial growth factor (anti-VEGF) in the management of proliferative diabetic retinopathy. Drugs Context 2018, 7, 212532. [Google Scholar] [CrossRef]
- Avastin|European Medicines Agency. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/avastin#overview-section (accessed on 27 November 2020).
- Patel, S.R.; Berezovsky, D.E.; McCarey, B.E.; Zarnitsyn, V.; Edelhauser, H.F.; Prausnitz, M.R. Targeted administration into the suprachoroidal space using a microneedle for drug delivery to the posterior segment of the eye. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4433–4441. [Google Scholar] [CrossRef]
- Song, L.; Llanga, T.; Conatser, L.M.; Zaric, V.; Gilger, B.C.; Hirsch, M.L. Serotype survey of AAV gene delivery via subconjunctival injection in mice. Gene Ther. 2018, 25, 402–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subrizi, A.; del Amo, E.M.; Korzhikov-Vlakh, V.; Tennikova, T.; Ruponen, M.; Urtti, A. Design principles of ocular drug delivery systems: Importance of drug payload, release rate, and material properties. Drug Discov. Today 2019. [Google Scholar] [CrossRef]
- Challa, P.; Luna, C.; Liton, P.B.; Chamblin, B.S.; Wakefiled, J.K.; Ramabhadran, R.; Epstein, D.L.; Gonzalez, P. Lentiviral mediated gene delivery to the anterior chamber of rodent eyes. Mol. Vis. 2005, 11, 425–430. [Google Scholar] [PubMed]
- Wang, L.; Xiao, R.; Andres-Mateos, E.; Vandenberghe, L.H. Single stranded adeno-associated virus achieves efficient gene transfer to anterior segment in the mouse eye. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Sim, K.S.; Kim, C.Y.; Park, T.K. Transduction Pattern of AAVs in the Trabecular Meshwork and Anterior-Segment Structures in a Rat Model of Ocular Hypertension. Mol. Ther. Methods Clin. Dev. 2019, 14, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, K.; Shen, J.; Hafiz, Z.; Hackett, S.F.; e Silva, R.L.; Khan, M.; Lorenc, V.E.; Chen, D.; Chadha, R.; Zhang, M.; et al. AAV8-vectored suprachoroidal gene transfer produces widespread ocular transgene expression. J. Clin. Investig. 2019, 129, 4901–4911. [Google Scholar] [CrossRef]
- Peng, Y.; Tang, L.; Zhou, Y. Subretinal Injection: A Review on the Novel Route of Therapeutic Delivery for Vitreoretinal Diseases. Ophthalmic Res. 2017, 58, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Ochakovski, G.A.; Bartz-Schmidt, K.U.; Fischer, M.D. Retinal Gene Therapy: Surgical Vector Delivery in the Translation to Clinical Trials. Front. Neurosci. 2017, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Omari, A.; Mahmoud, T.H. Vitrectomy; StatPearls Publishing: Treasure Island, SF, USA, 2020. [Google Scholar]
- Yang, G.S.; Schmidt, M.; Yan, Z.; Lindbloom, J.D.; Harding, T.C.; Donahue, B.A.; Engelhardt, J.F.; Kotin, R.; Davidson, B.L. Virus-Mediated Transduction of Murine Retina with Adeno-Associated Virus: Effects of Viral Capsid and Genome Size. J. Virol. 2002, 76, 7651–7660. [Google Scholar] [CrossRef] [Green Version]
- Petit, L.; Ma, S.; Cheng, S.-Y.; Gao, G.; Punzo, C. Rod Outer Segment Development Influences AAV-Mediated Photoreceptor Transduction After Subretinal Injection. Hum. Gene Ther. 2017. [Google Scholar] [CrossRef]
- Petit, L.; Khanna, H.; Punzo, C. Advances in gene therapy for diseases of the eye. Hum. Gene Ther. 2016, 27, 563–579. [Google Scholar] [CrossRef] [Green Version]
- Harvey, A.R.; Kamphuis, W.; Eggers, R.; Symons, N.A.; Blits, B.; Niclou, S.; Boer, G.J.; Verhaagen, J. Intravitreal injection of adeno-associated viral vectors results in the transduction of different types of retinal neurons in neonatal and adult rats: A comparison with lentiviral vectors. Mol. Cell. Neurosci. 2002, 21, 141–157. [Google Scholar] [CrossRef]
- Hellström, M.; Ruitenberg, M.J.; Pollett, M.A.; Ehlert, E.M.E.; Twisk, J.; Verhaagen, J.; Harvey, A.R. Cellular tropism and transduction properties of seven adeno-associated viral vector serotypes in adult retina after intravitreal injection. Gene Ther. 2009, 16, 521–532. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, T.; Miyake, K.; Asakawa, N.; Miyake, N.; Shimada, T.; Takahashi, H. Direct comparison of administration routes for aav8-mediated ocular gene therapy. Curr. Eye Res. 2013, 38, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Auricchio, A.; Kobinger, G.; Anand, V.; Hildinger, M.; O’Connor, E.; Maguire, A.M.; Wilson, J.M.; Bennett, J. Exchange of surface proteins impacts on viral vector cellular specificity and transduction characteristics: The retina as a model. Hum. Mol. Genet. 2001, 10, 3075–3081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, K.; Gehlbach, P.; Ando, A.; Wahlin, K.; Gunther, V.; McVey, D.; Wei, L.; Campochiaro, P.A. Intraocular adenoviral vector-mediated gene transfer in proliferative retinopathies. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1610–1615. [Google Scholar]
- Ueyama, K.; Mori, K.; Shoji, T.; Omata, H.; Gehlbach, P.L.; Brough, D.E.; Wei, L.L.; Yoneya, S. Ocular localization and transduction by adenoviral vectors are serotype-dependent and can be modified by inclusion of rgd fiber modifications. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Cotrim, A.P.; Baum, B.J. Gene Therapy: Some History, Applications, Problems, and Prospects. Toxicol. Pathol. 2008, 36, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.V.; Rosa da Costa, A.M.; Silva, G.A. Non-viral strategies for ocular gene delivery. Mater. Sci. Eng. C 2017, 77, 1275–1289. [Google Scholar] [CrossRef] [PubMed]
- Rissanen, T.T.; Ylä-Herttuala, S. Current status of cardiovascular gene therapy. Mol. Ther. 2007, 15, 1233–1247. [Google Scholar] [CrossRef] [PubMed]
- Volpers, C.; Kochanek, S. Adenoviral vectors for gene transfer and therapy. J. Gene Med. 2004, 6, S164–S171. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Sasaki, T.; Yano, K.; Sakurai, F.; Kawabata, K.; Kondoh, M.; Hayakawa, T.; Yagi, K.; Mizuguchi, H. Development of a recombinant adenovirus vector production system free of replication-competent adenovirus by utilizing a packaging size limit of the viral genome. Virus Res. 2011, 158, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Kinnunen, K.; Kalesnykas, G.; Mähönen, A.J.; Laidinen, S.; Holma, L.; Heikura, T.; Airenne, K.; Uusitalo, H.; Ylä-Herttuala, S. Baculovirus is an efficient vector for the transduction of the eye: Comparison of baculovirus- and adenovirus-mediated intravitreal vascular endothelial growth factor D gene transfer in the rabbit eye. J. Gene Med. 2009, 11, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.M.; Byrnes, G.A.; Gall, J.G.; Brough, D.E.; Richter King, C.; Wei, L.L. Alternate serotype adenovector provides long-term therapeutic gene expression in the eye. Mol. Vis. 2008, 14, 2535–2546. [Google Scholar] [PubMed]
- Reichel, M.B.; Ali, R.R.; Thrasher, A.J.; Hunt, D.M.; Bhattacharya, S.S.; Baker, D. Immune responses limit adenovirally mediated gene expression in the adult mouse eye. Gene Ther. 1998, 5, 1038–1046. [Google Scholar] [CrossRef] [Green Version]
- Ildefonso, C.J.; Kong, L.; Leen, A.; Chai, S.J.; Petrochelli, V.; Chintagumpala, M.; Hurwitz, M.Y.; Chévez-Barrios, P.; Hurwitz, R.L. Absence of systemic immune response to adenovectors after intraocular administration to children with retinoblastoma. Mol. Ther. 2010, 18, 1885–1890. [Google Scholar] [CrossRef]
- Campochiaro, P.A.; Nguyen, Q.D.; Shah, S.M.; Klein, M.L.; Holz, E.; Frank, R.N.; Saperstein, D.A.; Gupta, A.; Stout, J.T.; Macko, J.; et al. Adenoviral Vector-Delivered Pigment Epithelium-Derived Factor for Neovascular Age-Related Macular Degeneration: Results of a Phase I Clinical Trial. Hum. Gene Ther. 2006, 17, 167–176. [Google Scholar] [CrossRef]
- Deyle, D.R.; Russell, D.W. Adeno-associated virus vector integration. Curr. Opin. Mol. Ther. 2009, 11, 442–447. [Google Scholar]
- Liu, Y.; Fortmann, S.D.; Shen, J.; Wielechowski, E.; Tretiakova, A.; Yoo, S.; Kozarsky, K.; Wang, J.; Wilson, J.M.; Campochiaro, P.A. AAV8-antiVEGFfab Ocular Gene Transfer for Neovascular Age-Related Macular Degeneration. Mol. Ther. 2018, 26, 542–549. [Google Scholar] [CrossRef] [Green Version]
- Testa, F.; Maguire, A.M.; Rossi, S.; Pierce, E.A.; Melillo, P.; Marshall, K.; Banfi, S.; Surace, E.M.; Sun, J.; Acerra, C.; et al. Three-year follow-up after unilateral subretinal delivery of adeno-associated virus in patients with leber congenital amaurosis type 2. Ophthalmology 2013, 120, 1283–1291. [Google Scholar] [CrossRef] [Green Version]
- MacLachlan, T.K.; Lukason, M.; Collins, M.; Munger, R.; Isenberger, E.; Rogers, C.; Malatos, S.; Dufresne, E.; Morris, J.; Calcedo, R.; et al. Preclinical safety evaluation of AAV2-sFLT01 a gene therapy for age-related macular degeneration. Mol. Ther. 2011, 19, 326–334. [Google Scholar] [CrossRef]
- Ye, G.J.; Conlon, T.; Erger, K.; Sonnentag, P.; Sharma, A.K.; Howard, K.; Knop, D.R.; Chulay, J.D. Safety and Biodistribution Evaluation of rAAV2tYF-CB-hRS1, a Recombinant Adeno-Associated Virus Vector Expressing Retinoschisin, in RS1-Deficient Mice. Hum. Gene Ther. Clin. Dev. 2015, 26, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Constable, I.J.; Pierce, C.M.; Lai, C.-M.M.; Magno, A.L.; Degli-Esposti, M.A.; French, M.A.; McAllister, I.L.; Butler, S.; Barone, S.B.; Schwartz, S.D.; et al. Phase 2a Randomized Clinical Trial: Safety and Post Hoc Analysis of Subretinal rAAV.sFLT-1 for Wet Age-related Macular Degeneration. EBioMedicine 2016, 14, 168–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maguire, A.M.; High, K.A.; Auricchio, A.; Wright, J.F.; Pierce, E.A.; Testa, F.; Mingozzi, F.; Bennicelli, J.L.; Ying, G.S.; Rossi, S.; et al. Age-dependent effects of RPE65 gene therapy for Leber’s congenital amaurosis: A phase 1 dose-escalation trial. Lancet 2009, 374, 1597–1605. [Google Scholar] [CrossRef] [Green Version]
- Bennett, J.; Wellman, J.; Marshall, K.A.; McCague, S.; Ashtari, M.; DiStefano-Pappas, J.; Elci, O.U.; Chung, D.C.; Sun, J.; Wright, J.F.; et al. Safety and durability of effect of contralateral-eye administration of AAV2 gene therapy in patients with childhood-onset blindness caused by RPE65 mutations: A follow-on phase 1 trial. Lancet 2016, 388, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Cukras, C.; Wiley, H.E.; Jeffrey, B.G.; Sen, H.N.; Turriff, A.; Zeng, Y.; Vijayasarathy, C.; Marangoni, D.; Ziccardi, L.; Kjellstrom, S.; et al. Retinal AAV8-RS1 Gene Therapy for X-Linked Retinoschisis: Initial Findings from a Phase I/IIa Trial by Intravitreal Delivery. Mol. Ther. 2018, 26, 2282–2294. [Google Scholar] [CrossRef] [Green Version]
- Worgall, S.; Crystal, R.G. Gene Therapy. In Principles of Tissue Engineering, 4th ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 657–686. ISBN 9780123983589. [Google Scholar]
- Vargas, J.E.; Chicaybam, L.; Stein, R.T.; Tanuri, A.; Delgado-Cañedo, A.; Bonamino, M.H. Retroviral vectors and transposons for stable gene therapy: Advances, current challenges and perspectives. J. Transl. Med. 2016, 14, 288. [Google Scholar] [CrossRef] [Green Version]
- Hacein-Bey-Abina, S.; Garrigue, A.; Wang, G.P.; Soulier, J.; Lim, A.; Morillon, E.; Clappier, E.; Caccavelli, L.; Delabesse, E.; Beldjord, K.; et al. Insertional oncogenesis in 4 patients after retrovirus-mediated gene therapy of SCID-X1. J. Clin. Investig. 2008, 9, 3132–3142. [Google Scholar] [CrossRef] [PubMed]
- Askou, A.L.; Benckendorff, J.N.E.; Holmgaard, A.; Storm, T.; Aagaard, L.; Bek, T.; Mikkelsen, J.G.; Corydon, T.J. Suppression of Choroidal Neovascularization in Mice by Subretinal Delivery of Multigenic Lentiviral Vectors Encoding Anti-Angiogenic MicroRNAs. Hum. Gene Ther. Methods 2017, 28, 222–233. [Google Scholar] [CrossRef]
- Bemelmans, A.P.; Bonnel, S.; Houhou, L.; Dufour, N.; Nandrot, E.; Helmlinger, D.; Sarkis, C.; Abitbol, M.; Mallet, J. Retinal cell type expression specificity of HIV-1-derived gene transfer vectors upon subretinal injection in the adult rat: Influence of pseudotyping and promoter. J. Gene Med. 2005, 7, 1367–1374. [Google Scholar] [CrossRef]
- Cheng, L.; Chaidhawangul, S.; Wong-Staal, F.; Gilbert, J.; Poeschla, E.; Toyoguchi, M.; El-Bradey, M.H.; Bergeron-Lynn, G.; Soules, K.A.; Freeman, W.R. Human immunodeficiency virus type 2 (HIV-2) vector-mediated in vivo gene transfer into adult rabbit retina. Curr. Eye Res. 2002, 24, 196–201. [Google Scholar] [CrossRef]
- Ikeda, Y.; Yonemitsu, Y.; Miyazaki, M.; Kohno, R.I.; Murakami, Y.; Murata, T.; Goto, Y.; Tabata, T.; Ueda, Y.; Ono, F.; et al. Acute toxicity study of a simian immunodeficiency virus-based lentiviral vector for retinal gene transfer in nonhuman primates. Hum. Gene Ther. 2009, 20, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Matet, A.; Kostic, C.; Bemelmans, A.-P.; Moulin, A.; Rosolen, S.G.; Martin, S.; Mavilio, F.; Amirjanians, V.; Stieger, K.; Lorenz, B.; et al. Evaluation of tolerance to lentiviral LV-RPE65 gene therapy vector after subretinal delivery in non-human primates. Transl. Res. 2017, 188, 40–57.e4. [Google Scholar] [CrossRef] [Green Version]
- Campochiaro, P.A.; Lauer, A.K.; Sohn, E.H.; Mir, T.A.; Naylor, S.; Anderton, M.C.; Kelleher, M.; Harrop, R.; Ellis, S.; Mitrophanous, K.A. Lentiviral Vector Gene Transfer of Endostatin/Angiostatin for Macular Degeneration (GEM) Study. Hum. Gene Ther. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ono, C.; Okamoto, T.; Abe, T.; Matsuura, Y. Baculovirus as a tool for gene delivery and gene therapy. Viruses 2018, 10, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haeseleer, F.; Imanishi, Y.; Saperstein, D.A.; Palczewski, K. Gene transfer mediated by recombinant baculovirus into mouse eye. Investig. Ophthalmol. Vis. Sci. 2001, 42, 3294–3300. [Google Scholar]
- Reid, C.A.; Nettesheim, E.R.; Connor, T.B.; Lipinski, D.M. Development of an inducible anti-VEGF rAAV gene therapy strategy for the treatment of wet AMD. Sci. Rep. 2018, 8, 11763. [Google Scholar] [CrossRef] [Green Version]
- Pavlova, N.; Kaloudas, D.; Penchovsky, R. Riboswitch distribution, structure, and function in bacteria. Gene 2019, 708, 38–48. [Google Scholar] [CrossRef]
- Grishanin, R.; Vuillemenot, B.; Sharma, P.; Keravala, A.; Greengard, J.; Gelfman, C.; Blumenkrantz, M.; Lawrence, M.; Hu, W.; Kiss, S.; et al. Preclinical Evaluation of ADVM-022, a Novel Gene Therapy Approach to Treating Wet Age-Related Macular Degeneration. Mol. Ther. 2019, 27, 118–129. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.P.; O’Flynn, N.M.J.; Gatherer, M.; McClements, M.E.; Scott, J.A.; MacLaren, R.E.; Goverdhan, S.; Glennie, M.J.; Lotery, A.J. AAV2/8 Anti-angiogenic Gene Therapy Using Single-Chain Antibodies Inhibits Murine Choroidal Neovascularization. Mol. Ther. Methods Clin. Dev. 2019, 13, 86–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.S.; Kim, H.J.; Shin, O.K.; Choi, J.S.; Kim, J.; Cho, Y.H.; Ha, J.; Park, T.K.; Lee, J.Y.; Park, K.; et al. Intravitreal injection of AAV expressing soluble VEGF receptor-1 variant induces anti-VEGF activity and suppresses choroidal neovascularization. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5398–5407. [Google Scholar] [CrossRef]
- Park, T.K.; Lee, S.H.; Choi, J.S.; Nah, S.K.; Kim, H.J.; Park, H.Y.; Lee, H.; Lee, S.H.S.; Park, K. Adeno-Associated Viral Vector-Mediated mTOR Inhibition by Short Hairpin RNA Suppresses Laser-Induced Choroidal Neovascularization. Mol. Ther. Nucleic Acids 2017, 8, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.S.; Chang, H.S.; Kim, J.H.; Kim, H.J.; Choi, J.S.; Chung, S.; Woo, H.N.; Lee, K.J.; Park, K.; Lee, J.Y.; et al. Inhibition of mTOR via an AAV-delivered shRNA tested in a rat oir model as a potential antiangiogenic gene therapy. Investig. Ophthalmol. Vis. Sci. 2020, 61. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.S.; Chang, H.S.; Kim, H.J.; Choi, J.S.; Kim, J.; Kim, J.H.; Woo, H.N.; Nah, S.K.; Jung, S.J.; Lee, J.Y.; et al. Effects of Stuffer DNA on the Suppression of Choroidal Neovascularization by a rAAV Expressing a mTOR-Inhibiting shRNA. Mol. Ther. Methods Clin. Dev. 2019, 14, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Biswal, M.R.; Prentice, H.M.; Smith, G.W.; Zhu, P.; Tong, Y.; Dorey, C.K.; Lewin, A.S.; Blanks, J.C. Cell-specific gene therapy driven by an optimized hypoxia-regulated vector reduces choroidal neovascularization. J. Mol. Med. 2018, 96, 1107–1118. [Google Scholar] [CrossRef]
- Tu, L.; Wang, J.H.; Barathi, V.A.; Prea, S.M.; He, Z.; Lee, J.H.; Bender, J.; King, A.E.; Logan, G.J.; Alexander, I.E.; et al. AAV-mediated gene delivery of the calreticulin anti-angiogenic domain inhibits ocular neovascularization. Angiogenesis 2018, 21, 95–109. [Google Scholar] [CrossRef]
- Sun, P.; Liu, Z. Overexpressing kringle 1 domain of hepatocyte growth factor with adeno-associated virus inhibits the pathological retinal neovascularization in an oxygen-induced retinopathy mouse model. Biochem. Biophys. Res. Commun. 2019, 508, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Zhang, W.; He, T.; Xing, Y. Silencing of galectin-1 inhibits retinal neovascularization and ameliorates retinal hypoxia in a murine model of oxygen-induced ischemic retinopathy. Exp. Eye Res. 2017, 159, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhou, G.; Wu, W.; Duan, Y.; Ma, G.; Song, J.; Xiao, R.; Vandenberghe, L.; Zhang, F.; D’Amore, P.A.; et al. Genome editing abrogates angiogenesis in vivo. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Koo, T.; Park, S.W.; Jo, D.H.; Kim, D.; Kim, J.H.; Cho, H.-Y.; Kim, J.; Kim, J.H.; Kim, J.-S. CRISPR-LbCpf1 prevents choroidal neovascularization in a mouse model of age-related macular degeneration. Nat. Commun. 2018. [Google Scholar] [CrossRef] [PubMed]
- Askou, A.L.; Alsing, S.; Benckendorff, J.N.E.; Holmgaard, A.; Mikkelsen, J.G.; Aagaard, L.; Bek, T.; Corydon, T.J. Suppression of Choroidal Neovascularization by AAV-Based Dual-Acting Antiangiogenic Gene Therapy. Mol. Ther. Nucleic Acids 2019, 16, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, M.L.; Conatser, L.M.; Smith, S.M.; Salmon, J.H.; Wu, J.; Buglak, N.E.; Davis, R.; Gilger, B.C. AAV vector-meditated expression of HLA-G reduces injury-induced corneal vascularization, immune cell infiltration, and fibrosis. Sci. Rep. 2017, 7, 17840. [Google Scholar] [CrossRef] [Green Version]
- Simmons, A.B.; Bretz, C.A.; Wang, H.; Kunz, E.; Hajj, K.; Kennedy, C.; Yang, Z.; Suwanmanee, T.; Kafri, T.; Hartnett, M.E. Gene therapy knockdown of VEGFR2 in retinal endothelial cells to treat retinopathy. Angiogenesis 2018, 21, 751–764. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Wang, H.; Simmons, A.B.; Suwanmanee, T.; Stoddard, G.J.; Kafri, T.; Elizabeth Hartnett, M. Targeted Knockdown of Overexpressed VEGFA or VEGF164 in Müller cells maintains retinal function by triggering different signaling mechanisms. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Abdollahi, I.A.; Hahnfeldt, P.; Maercker, C.; Grö, H.-J.; Debus, J.; Ansorge, W.; Folkman, J.; Hlatky, L. Endostatin’s Antiangiogenic Signaling Network Endostatin, a 20 kDa fragment of collagen XVIII, is a member of a group of endogenous antiangiogenic pro. Mol. Cell 2004, 13, 649–663. [Google Scholar] [CrossRef]
- Mori, K.; Ando, A.; Gehlbach, P.; Nesbitt, D.; Takahashi, K.; Goldsteen, D.; Penn, M.; Chen, C.T.; Mori, K.; Melia, M.; et al. Inhibition of choroidal neovascularization by intravenous injection of adenoviral vectors expressing secretable endostatin. Am. J. Pathol. 2001, 159, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Bee, Y.S.; Ma, Y.L.; Chen, J.; Tsai, P.J.; Sheu, S.J.; Lin, H.C.; Huang, H.; Liu, G.S.; Tai, M.H. Inhibition of experimental choroidal neovascularization by a novel peptide derived from calreticulin anti-angiogenic domain. Int. J. Mol. Sci. 2018, 19, 2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, X.; Yang, S.; Ingle, G.; Zlot, C.; Rangell, L.; Kowalski, J.; Schwall, R.; Ferrara, N.; Gerritsen, M.E. Hepatocyte growth factor enhances vascular endothelial growth factor-induced angiogenesis in vitro and in vivo. Am. J. Pathol. 2001, 158, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Zhen, F.Y.; Gao, Y.; Ji, C.L.; Hai, X.C.; Ching, C.L.; Poon, R.T.; Sheung, T.F.; Luk, J.M.; Kong, H.S.; et al. The kringle 1 domain of hepatocyte growth factor has antiangiogenic and antitumor cell effects on hepatocellular carcinoma. Cancer Res. 2008, 68, 404–414. [Google Scholar] [CrossRef] [Green Version]
- van Beijnum, J.R.; Thijssen, V.L.; Läppchen, T.; Wong, T.J.; Verel, I.; Engbersen, M.; Schulkens, I.A.; Rossin, R.; Grüll, H.; Griffioen, A.W.; et al. A key role for galectin-1 in sprouting angiogenesis revealed by novel rationally designed antibodies. Int. J. Cancer 2016, 139, 824–835. [Google Scholar] [CrossRef]
- Abu El-Asrar, A.M.; Ahmad, A.; Allegaert, E.; Siddiquei, M.M.; Alam, K.; Gikandi, P.W.; De Hertogh, G.; Opdenakker, G. Galectin-1 studies in proliferative diabetic retinopathy. Acta Ophthalmol. 2020, 98, e1–e12. [Google Scholar] [CrossRef]
- Manghwar, H.; Lindsey, K.; Zhang, X.; Jin, S. CRISPR/Cas System: Recent Advances and Future Prospects for Genome Editing. Trends Plant Sci. 2019, 24, 1102–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; Van Der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.; Jin, J.; Hu, Y.; Zhou, K.; Ma, J.X. Overexpression of pigment epithelium-derived factor inhibits retinal inflammation and neovascularization. Am. J. Pathol. 2011, 178, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Carosella, E.D.; Rouas-Freiss, N.; Tronik-Le Roux, D.; Moreau, P.; LeMaoult, J. HLA-G: An Immune Checkpoint Molecule; Advances in Immunology; Academic Press: Cambridge, MA, USA, 2015; Volume 127, pp. 33–144. ISBN 9780128022450. [Google Scholar]
- Le Discorde, M.; Moreau, P.; Sabatier, P.; Legeais, J.M.; Carosella, E.D. Expression of HLA-G in Human Cornea, an Immune-Privileged Tissue. Hum. Immunol. 2003, 64, 1039–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanderVeen, D.K.; Cataltepe, S.U. Anti-vascular endothelial growth factor intravitreal therapy for retinopathy of prematurity. Semin. Perinatol. 2019, 43, 375–380. [Google Scholar] [CrossRef]
- Wiley Database on Gene Therapy Trials Worldwide Gene Therapy Clinical Trials Worldwide. Available online: http://www.abedia.com/wiley/indications.php (accessed on 13 September 2020).
- ClinicalTrials.gov. AAVCAGsCD59 for the Treatment of Wet AMD. Available online: https://clinicaltrials.gov/ct2/show/study/NCT03585556?cond=gene+therapy+eye&draw=6&rank=47 (accessed on 13 September 2020).
- ClinicalTrials.gov. ADVM-022 Intravitreal Gene Therapy for Wet AMD. Available online: https://clinicaltrials.gov/ct2/show/NCT03748784?cond=gene+therapy+eye&draw=3&rank=15 (accessed on 13 September 2020).
- Globe Newswire Adverum Biotechnologies Announces New INFINITY Phase 2 Trial for ADVM-022 in Diabetic Macular Edema, Reports Recent Business Progress and First Quarter 2020 Financial Results. Available online: https://www.globenewswire.com/news-release/2020/05/28/2040528/0/en/Adverum-Biotechnologies-Announces-New-INFINITY-Phase-2-Trial-for-ADVM-022-in-Diabetic-Macular-Edema-Reports-Recent-Business-Progress-and-First-Quarter-2020-Financial-Results.html (accessed on 15 November 2020).
- ClinicalTrials.gov. RGX-314 Gene Therapy for Neovascular AMD Trial. Available online: https://clinicaltrials.gov/ct2/show/NCT03066258?cond=gene+therapy+eye&draw=3&rank=12 (accessed on 13 September 2020).
- ClinicalTrials.gov. Long-term Follow-Up Study of RGX-314. Available online: https://clinicaltrials.gov/ct2/show/NCT03999801?cond=gene+therapy+retina&draw=5&rank=33 (accessed on 13 September 2020).
- ClinicalTrials.gov. RGX-314 Gene Therapy Administered in the Suprachoroidal Space for Participants with Neovascular Age-Related Macular Degeneration (nAMD). Available online: https://clinicaltrials.gov/ct2/show/NCT04514653?cond=gene+therapy%2C+amd&draw=2&rank=2 (accessed on 13 September 2020).
- ClinicalTrials.gov. RGX-314 Gene Therapy Administered in the Suprachoroidal Space for Participants with Diabetic Retinopathy (DR) without Center Involved-Diabetic Macular Edema (CI-DME). Available online: https://clinicaltrials.gov/ct2/show/NCT04567550?term=RGX-314&draw=2&rank=2 (accessed on 12 November 2020).
- Rakoczy, E.P.; Magno, A.L.; Lai, C.M.; Pierce, C.M.; Degli-Esposti, M.A.; Blumenkranz, M.S.; Constable, I.J. Three-Year Follow-Up of Phase 1 and 2a rAAV.sFLT-1 Subretinal Gene Therapy Trials for Exudative Age-Related Macular Degeneration. Am. J. Ophthalmol. 2019, 204, 113–123. [Google Scholar] [CrossRef]
- Heier, J.S.; Kherani, S.; Desai, S.; Dugel, P.; Kaushal, S.; Cheng, S.H.; Delacono, C.; Purvis, A.; Richards, S.; Le-Halpere, A.; et al. Intravitreous injection of AAV2-sFLT01 in patients with advanced neovascular age-related macular degeneration: A phase 1, open-label trial. Lancet 2017, 390, 50–61. [Google Scholar] [CrossRef]
- Farkas, I.; Baranyi, L.; Ishikawa, Y.; Okada, N.; Bohata, C.; Budai, D.; Fukuda, A.; Imai, M.; Okada, H. CD59 blocks not only the insertion of C9 into MAC but inhibits ion channel formation by homologous C5b-8 as well as C5b-9. J. Physiol. 2002, 539, 537–545. [Google Scholar] [CrossRef]
- Hageman, G.S.; Anderson, D.H.; Johnson, L.V.; Hancox, L.S.; Taiber, A.J.; Hardisty, L.I.; Hageman, J.L.; Stockman, H.A.; Borchardt, J.D.; Gehrs, K.M.; et al. A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 7227–7232. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimi, K.B.; Fijalkowski, N.; Cano, M.; Handa, J.T. Decreased membrane complement regulators in the retinal pigmented epithelium contributes to age-related macular degeneration. J. Pathol. 2013, 229, 729–742. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Faber, C.; Falk, M.; Nissen, M.H.; Hviid, T.V.F.; Sørensen, T.L. Altered expression of CD46 and CD59 on leukocytes in neovascular age-related macular degeneration. Am. J. Ophthalmol. 2012, 154, 193–199. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. ADVM-022 Intravitreal Gene Therapy for DME. Available online: https://clinicaltrials.gov/ct2/show/NCT04418427?cond=gene+therapy+retina&draw=3&rank=20 (accessed on 13 September 2020).
- Globe Newswire Adverum Biotechnologies Announces Positive Interim Data from Cohorts 1-4 from OPTIC Phase 1 Trial of ADVM-022 Intravitreal Gene Therapy for wet AMD, Reports Recent Business Progress and Second Quarter 2020 Financial Results. Available online: https://www.globenewswire.com/news-release/2020/08/10/2075903/0/en/Adverum-Biotechnologies-Announces-Positive-Interim-Data-from-Cohorts-1-4-from-OPTIC-Phase-1-Trial-of-ADVM-022-Intravitreal-Gene-Therapy-for-wet-AMD-Reports-Recent-Business-Progress.html (accessed on 12 November 2020).
- Eyewire News Regenxbio Announces Additional Positive Long-Term and Interim Phase 1/2a Trial Update for RGX-314 for the Treatment of Wet AMD. Available online: https://eyewire.news/articles/regenxbio-announces-additional-positive-long-term-and-interim-phase-1-2a-trial-update-for-rgx-314-for-the-treatment-of-wet-amd/ (accessed on 13 September 2020).
- Modern Retina Staff Reports Regenxbio: Dosing of first patient in Phase 2 AAVIATE Trial for wet AMD|Modern Retina. Available online: https://www.modernretina.com/view/regenxbio-dosing-of-first-patient-in-phase-2-aaviate-trial-for-wet-amd (accessed on 12 November 2020).
- Failla, C.M.; Carbo, M.; Morea, V. Positive and negative regulation of angiogenesis by soluble vascular endothelial growth factor receptor-1. Int. J. Mol. Sci. 2018, 19, 1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tombran-Tink, J.; Chader, G.G.; Johnson, L.V. PEDF: A pigment epithelium-derived factor with potent neuronal differentiative activity. Exp. Eye Res. 1991, 53, 411–414. [Google Scholar] [CrossRef] [Green Version]
- Ogata, N.; Nishikawa, M.; Nishimura, T.; Mitsuma, Y.; Matsumura, M. Unbalanced vitreous levels of pigment epithelium-derived factor and vascular endothelial growth factor in diabetic retinopathy. Am. J. Ophthalmol. 2002, 134, 348–353. [Google Scholar] [CrossRef]
- Ogata, N.; Tombran-Tink, J.; Nishikawa, M.; Nishimura, T.; Mitsuma, Y.; Sakamoto, T.; Matsumura, M. Pigment epithelium-derived factor in the vitreous is low in diabetic retinopathy and high in rhegmatogenous retinal detachment. Am. J. Ophthalmol. 2001, 132, 378–382. [Google Scholar] [CrossRef]
- Holekamp, N.M.; Bouck, N.; Volpert, O. Pigment Epithelium-derived Factor Is Deficient in the Vitreous of Patients with Choroidal Neovascularization due to Age-related Macular Degeneration. Am. J. Ophthalmol. 2002, 134, 220–227. [Google Scholar] [CrossRef]
- Ohno-Matsui, K.; Morita, I.; Tombran-Tink, J.; Mrazek, D.; Onodera, M.; Uetama, T.; Hayano, M.; Murota, S.; Mochizuki, M. Novel mechanism for age-related macular degeneration: An equilibrium shift between the angiogenesis factors VEGF and PEDF. J. Cell. Physiol. 2001, 189, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Moore, N.A.; Bracha, P.; Hussain, R.M.; Morral, N.; Ciulla, T.A. Gene therapy for age-related macular degeneration. Expert Opin. Biol. Ther. 2017, 17, 1235–1244. [Google Scholar] [CrossRef]
- Wallace, D.K.; Dean, T.W.; Hartnett, M.E.; Kong, L.; Smith, L.E.; Hubbard, G.B.; McGregor, M.L.; Jordan, C.O.; Mantagos, I.S.; Bell, E.F.; et al. A Dosing Study of Bevacizumab for Retinopathy of Prematurity: Late Recurrences and Additional Treatments. Ophthalmology 2018, 125, 1961–1966. [Google Scholar] [CrossRef]
- Rakoczy, E.P.; Lai, C.-M.; Magno, A.L.; Wikstrom, M.E.; French, M.A.; Pierce, C.M.; Schwartz, S.D.; Blumenkranz, M.S.; Chalberg, T.W.; Degli-Esposti, M.A.; et al. Gene therapy with recombinant adeno-associated vectors for neovascular age-related macular degeneration: 1 year follow-up of a phase 1 randomised clinical trial. Lancet 2015, 386, 2395–2403. [Google Scholar] [CrossRef] [Green Version]
- Veleri, S.; Lazar, C.H.; Chang, B.; Sieving, P.A.; Banin, E.; Swaroop, A. Biology and therapy of inherited retinal degenerative disease: Insights from mouse models. Dis. Model. Mech. 2015, 8, 109–129. [Google Scholar] [CrossRef] [Green Version]
- Yla-Herttuala, S.; Alitalo, K. Gene transfer as a tool to induce therapeutic vascular growth. Nat. Med. 2003, 9, 694–701. [Google Scholar] [CrossRef]
- Spooner, K.; Hong, T.; Wijeyakumar, W.; Chang, A.A. Switching to aflibercept among patients with treatment-resistant neovascular age-related macular degeneration: A systematic review with meta-analysis. Clin. Ophthalmol. 2017, 11, 161–177. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Zhao, J.; Sun, X. Resistance to anti-VEGF therapy in neovascular age-related macular degeneration: A comprehensive review. Drug Des. Devel. Ther. 2016, 10, 1857–1867. [Google Scholar] [PubMed] [Green Version]
- Georgiadis, A.; Duran, Y.; Ribeiro, J.; Abelleira-Hervas, L.; Robbie, S.J.; Sünkel-Laing, B.; Fourali, S.; Gonzalez-Cordero, A.; Cristante, E.; Michaelides, M.; et al. Development of an optimized AAV2/5 gene therapy vector for Leber congenital amaurosis owing to defects in RPE65. Gene Ther. 2016, 23, 857–862. [Google Scholar] [CrossRef]
- Bulaklak, K.; Gersbach, C.A. The once and future gene therapy. Nat. Commun. 2020, 11, 1–4. [Google Scholar] [CrossRef]
- Cabral, T.; DiCarlo, J.E.; Justus, S.; Sengillo, J.D.; Xu, Y.; Tsang, S.H. CRISPR applications in ophthalmologic genome surgery. Curr. Opin. Ophthalmol. 2017, 28, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Single Ascending Dose Study in Participants with LCA10—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT03872479 (accessed on 30 January 2021).
- Fitzpatrick, Z.; Leborgne, C.; Barbon, E.; Masat, E.; Ronzitti, G.; van Wittenberghe, L.; Vignaud, A.; Collaud, F.; Charles, S.; Simon Sola, M.; et al. Influence of Pre-existing Anti-capsid Neutralizing and Binding Antibodies on AAV Vector Transduction. Mol. Ther. Methods Clin. Dev. 2018, 9, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benveniste, O.; Boutin, S.; Monteilhet, V.; Veron, P.; Leborgne, C.; Montus, M.F.; Masurier, C. Prevalence of Serum IgG and Neutralizing Factors Against Adeno-Associated Virus (AAV) Types 1,2,5,6,8, and 9 in the Healthy Population: Implications for Gene Therapy Using AAV Vectors. Hum. Gene Ther. 2010, 21, 704–712. [Google Scholar] [CrossRef]


{kind=link}
| Animalmodel | Vector | Injection Site | Anti-Angiogenic Factor | Indication | Reference |
|---|---|---|---|---|---|
| Laser induced CNV in mouse | rAAV2/2-riboswitch | intravitreal | aflibercept | wet AMD | [70] |
| Laser induced CNV in non-human primate | AAV2.7m8 | intravitreal | codon-optimised cDNA of the aflibercept protein (ADVM-022) | wet AMD | [72] |
| Laser induced CNV in mouse | AAV2/8 | subretinal | anti-VEGF single-chain fragment variable (scFv) antibody | wet AMD | [73] |
| Transgenic mice expressing human VEGF in retina (rho/VEGF mice), rats expressing human VEGF in retina | AAV8 | subretinal, suprachoroidal | humanized antibody fragment that binds human VEGF (anti-VEGFfab) (RGX-314) | wet AMD | [27,51] |
| Laser induced CNV in mouse | AAV2 | intravitreal | soluble variant of VEGFR-1 | wet AMD | [74] |
| Oxygen-induced retinopathy in rat, laser induced CNV in mouse | AAV2 | intravitreal | mTOR shRNA, | wet AMD, ROP, DR | [75,76,77] |
| Laser induced CNV in mouse | scAAV2 | subretinal | endostatin | wet AMD | [78] |
| Oxygen-induced retinopathy in rat, laser induced CNV in mouse | scAAV2 | intravitreal, subretinal | calreticulin anti-angiogenic domain (CAD) | wet AMD, DR | [79] |
| Oxygen-induced retinopathy in mouse | rAAV | intravitreal | Kringle1 domain of hepatocyte growth factor (HGFK1) | wet AMD, DR | [80] |
| Oxygen-induced retinopathy in mouse | Adenovirus GV120 | intravitreal | Galetin-1-RNAinterference | ROP | [81] |
| Laser induced CNV and oxygen-induced retinopathy in mouse, | AAV1-CRISPR/Cas9 | intravitreal | CRISPR/Cas9 targeted to VEGFR 2 | wet AMD, ROP | [82] |
| Laser induced CNV in mouse | AAV9-CRISPR-LbCpf1 | intravitreal | CRISPR-LbCpf1 targeted to VEGF A and HIF1α | wet AMD | [83] |
| Laser induced CNV in mouse | Lentivirus | subretinal | microRNAs targeting VEGF-A | wet AMD | [62] |
| Laser induced CNV in mouse | AAV5 | subretinal | PEDF, multiple miRNAs targeting the VEGFA gene | wet AMD | [84] |
| Corneal chemical burn in a rabbit | AAV8/9 chimeric capsid (8G9) | corneal intrastromal | codon optimized human leucocyte antigen G1+ G5 | Corneal neovascularization | [85] |
| Oxygen-induced retinopathy in rat | Lentivirus | subretinal | shRNAs to VEGFR2 or STAT3 | ROP | [86] |
| Oxygen-induced retinopathy in rat | Lentivirus | subretinal | VEGF-A or VEGF-A164 shRNA | ROP | [87] |
| Clinical Trial | Vector | Injection Site | Anti-Angiogenic Factor | Indication | Reference |
|---|---|---|---|---|---|
| AAVCAGsCD59 (NCT03585556) | AAV2 | intravitreal | soluble CD59 | wet AMD | [102] |
| ADVM-022 (NCT03748784) | AA7 | intravitreal | codon-optimized cDNA of the aflibercept protein | wet AMD | [103] |
| ADVM-022 (NCT04418427) | AA7 | intravitreal | codon-optimized cDNA of the aflibercept protein | diabetic macular edema | [104] |
| RXG-314 (NCT03066258, NCT03999801) | AAV8 | subretinal | soluble anti-VEGF monoclonal antibody fragment related to ranibizumab | wet AMD | [105,106] |
| RXG-314 (NCT04514653) | AAV8 | subrachoroidal | soluble anti-VEGF monoclonal antibody fragment related to ranibizumab | wet AMD | [107] |
| RXG-314 (NCT04567550) | AAV8 | subrachoroidal | soluble anti-VEGF monoclonal antibody fragment related to ranibizumab | diabetic retinopathy | [108] |
| Soluble FLT-1 (NCT01494805) | AAV2 | subretinal | Soluble FLT-1 | wet AMD | [109] |
| Soluble FLT-1 (NCT01494805) | AAV2 | subretinal | Soluble FLT-1 | wet AMD | [55] |
| Soluble FLT-1 (NCT01024998) | AAV2 | intravitreal | Soluble FLT-1 | wet AMD | [110] |
| RetinoStat (NCT01301443, NCT01678872) | lentivirus | subretinal | endostatin and angiostatin | wet AMD | [67] |
| AdPEDF (NCT00109499) | Adenovirus 5 | intravitreal | cDNA for human pigment epithelium-derived factor (PEDF) | wet AMD | [49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koponen, S.; Kokki, E.; Kinnunen, K.; Ylä-Herttuala, S. Viral-Vector-Delivered Anti-Angiogenic Therapies to the Eye. Pharmaceutics 2021, 13, 219. https://doi.org/10.3390/pharmaceutics13020219
Koponen S, Kokki E, Kinnunen K, Ylä-Herttuala S. Viral-Vector-Delivered Anti-Angiogenic Therapies to the Eye. Pharmaceutics. 2021; 13(2):219. https://doi.org/10.3390/pharmaceutics13020219
Chicago/Turabian StyleKoponen, Sanna, Emmi Kokki, Kati Kinnunen, and Seppo Ylä-Herttuala. 2021. "Viral-Vector-Delivered Anti-Angiogenic Therapies to the Eye" Pharmaceutics 13, no. 2: 219. https://doi.org/10.3390/pharmaceutics13020219