
Biomaterial and Therapeutic Approaches for the Manipulation of Macrophage Phenotype in Peripheral and Central Nerve Repair


 , , , and
, , , and
Abstract
:1. Introduction
2. Macrophages in Peripheral Nervous System Injury and Repair
2.1. Overview of Peripheral Nerveous System (PNS) Injury
2.1.1. Distal Defect Wallerian Degeneration
2.1.2. Axonal Regrowth
2.1.3. Restoration of Nerve Function
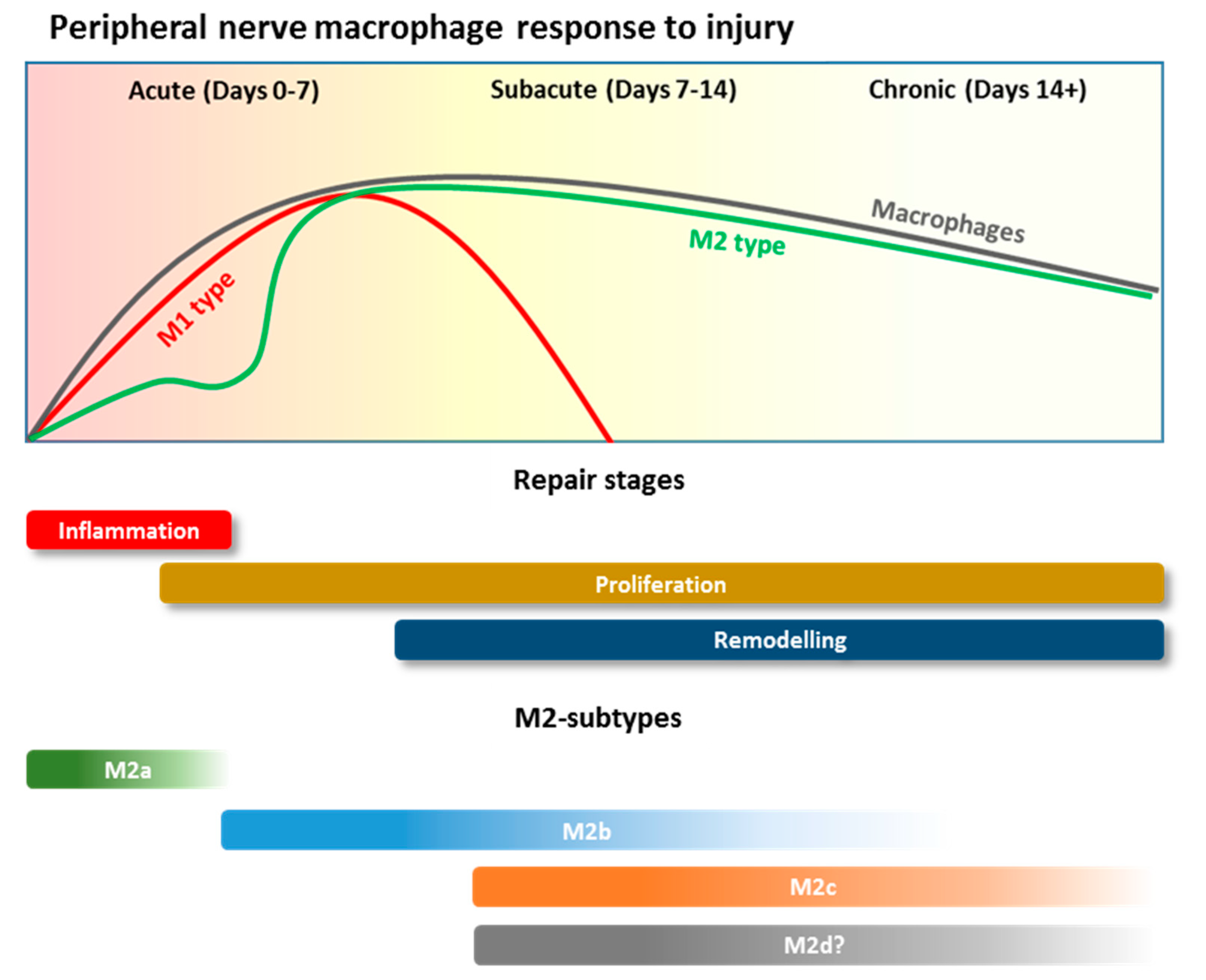
2.2. Macrophage Recruitment during PNI
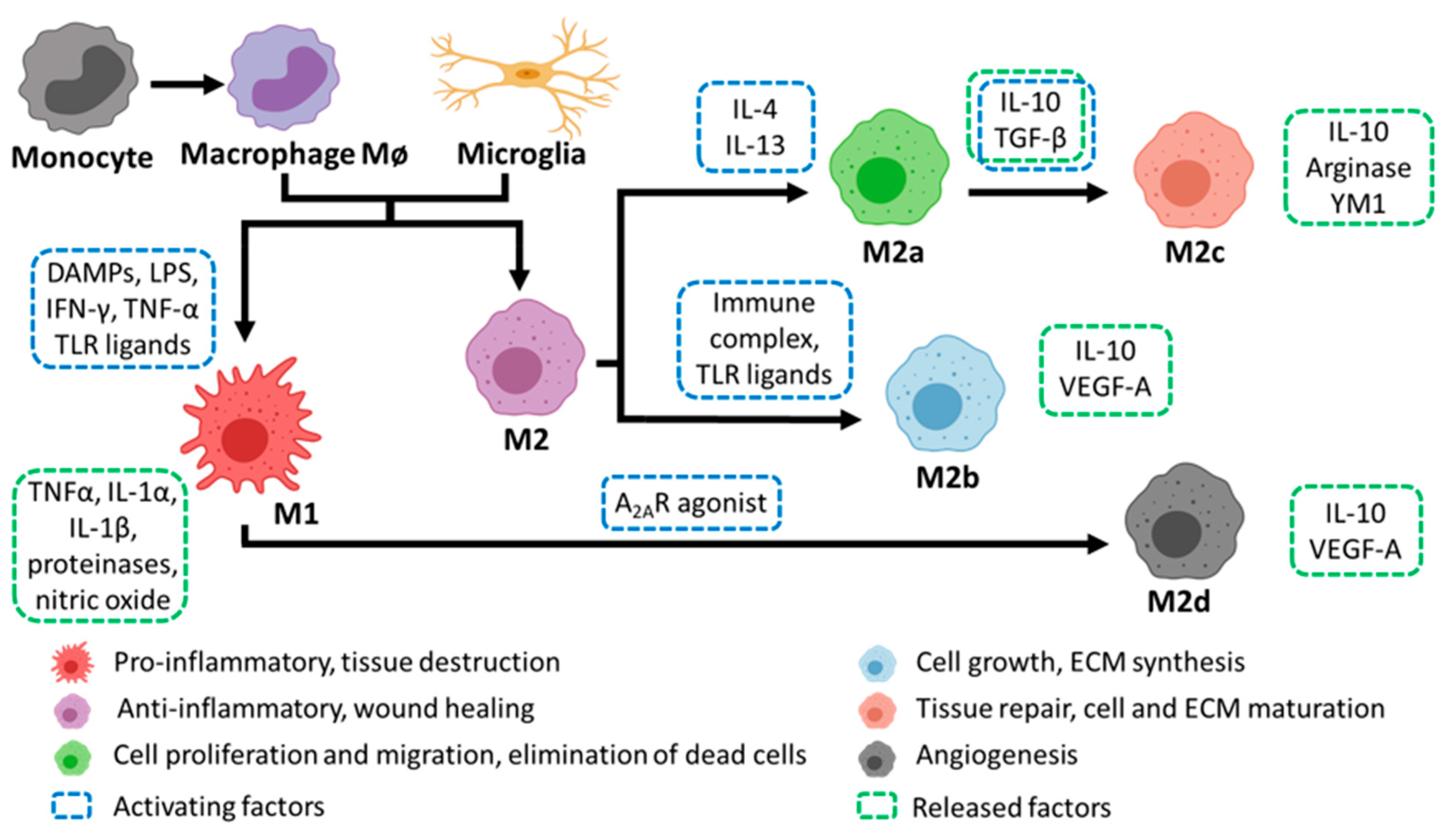
2.3. Macrophage Polarization and Roles during PNI
3. Macrophages in Central Nervous System Damage and Repair
3.1. Overview of Central Nerve System (CNS) Injury
3.1.1. Axon Degeneration
3.1.2. Fibroglial Scar Formation
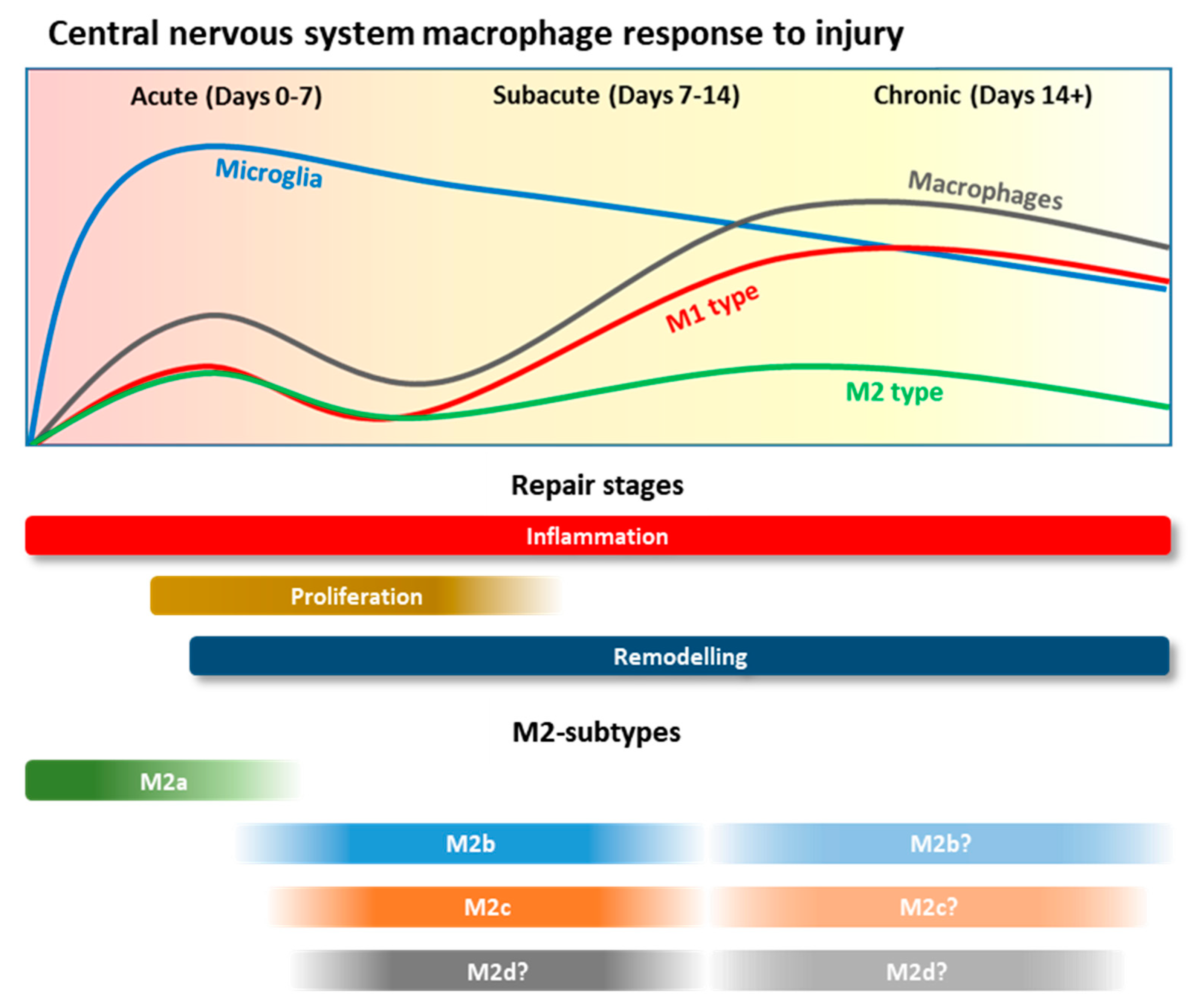
3.2. Macrophage Recruitment after CNS Injury
3.3. Macrophage Polarisation and Roles during CNS Injury
4. Macrophage Response to Current Peripheral Nerve Repair Strategies
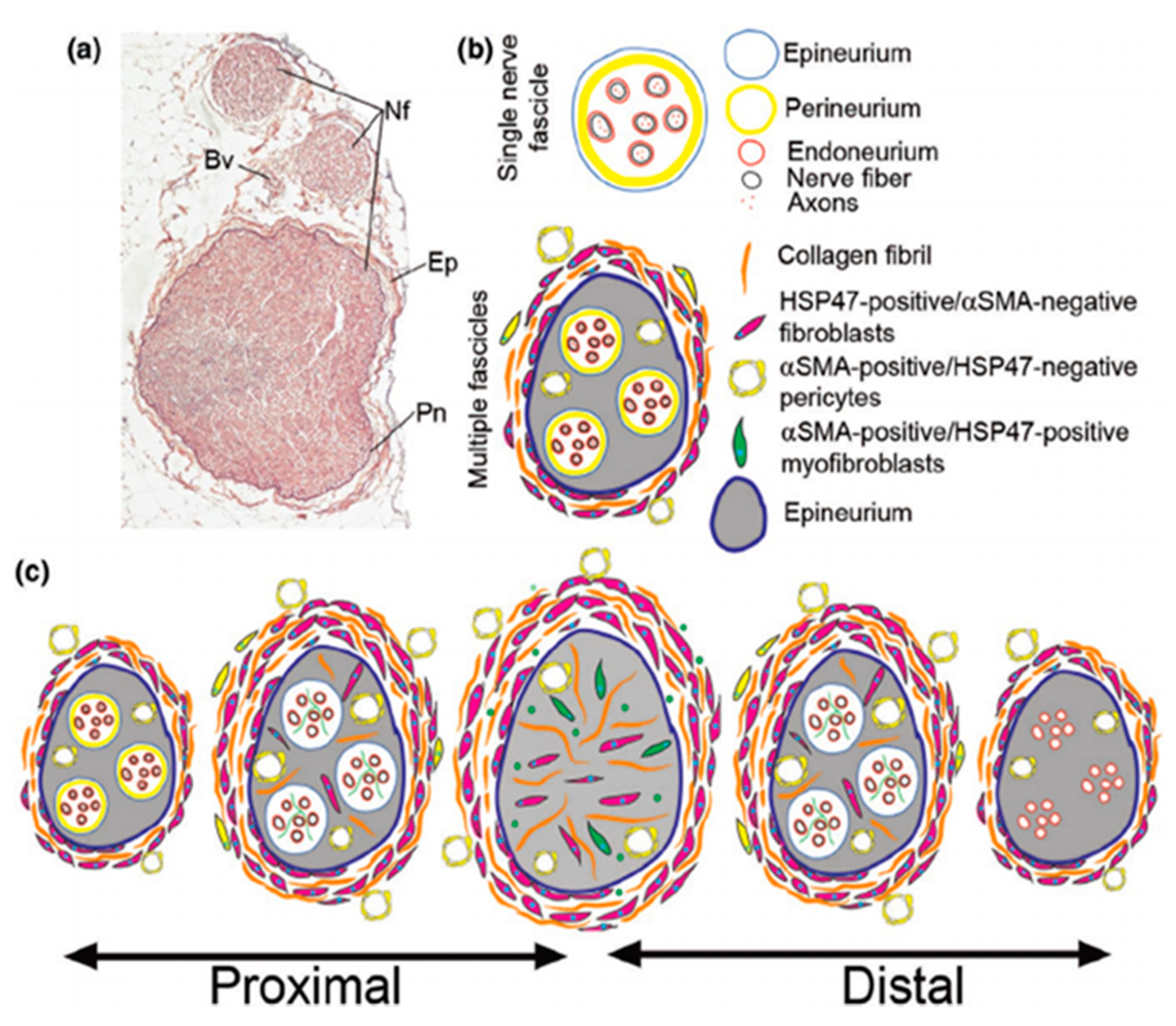
4.1. Auto and Allografts
4.1.1. Decellularized Nerve Grafts
4.1.2. Biomaterial Nerve Guidance Conduits
5. Biomaterial Approaches to Direct Macrophage Phenotype in Peripheral Nervous System Repair
5.1. Biomaterial Composition
5.1.1. Organic
5.1.2. Synthetic Polymers
5.2. Physical or Chemically Modified Biomaterials
5.2.1. Physical Modifications
5.2.2. Chemical Modifications
5.3. Therapeutic Molecule Incorporation
5.3.1. Small Molecule Drugs
5.3.2. Proteins/Macromolecules (Cytokines, Peptides, Antibodies)
5.3.3. Cellular Therapeutics
5.3.4. Gene Therapy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TECHNIQUE/PRODUCT | |||
|---|---|---|---|
| Biological Approach | In vivo | Immune Response | Ref |
| PCL/collagen VI nerve conduit | |||
| Sustained release of Collagen VI | Sciatic nerve in SD rats | Promotes polarization of macrophages towards M2 phenotype | [165] |
| Polysulfone nerve guidance channel | |||
| Local delivery of IL-4 or IFN-γ | * Sciatic nerve in adult Lewis male rats | IL-4 or IFN-γ polarizes macrophages towards M2 and M1 phenotypes, respectively | [198] |
| PCL nanofibrous scaffold | |||
| Local delivery of IL-10 | Sciatic nerve in Wistar rats | IL-10-conjugated PCL scaffold induces M2 polarization | [200] |
| Polysulfone nerve guidance scaffold | |||
| Local delivery of Fractalkine | Sciatic nerve in Lewis rats | Fractalkine promotes the recruitment of anti-inflammatory macrophages | [199] |
| Therapeutical treatment | |||
| Miconazole | * C57BL/6 mice | Suppresses M1 phenotype and induces M2 phenotype | [195] |
| Thrombomodulin | In vivo: sciatic nerve in male SD rats and In vitro: THP-1 cells | Promotes nerve repair by M2 polarization | [196] |
| Vitamin B | Femoral nerve in AO rats | promotes M2 polarization, reduces pro-inflammatory cytokines and increases anti-inflammatory cytokines | [197] |
| ECM/PLGA bridge | |||
| EPI-NCSCs delivery | Sciatic nerve in adult female SD rats | Delivery decreases inflammatory fibroblasts and increases the M2/M1 macrophages ratio | [201] |
| Cell Transplant | |||
| SKPSCs delivery | In vivo: Sciatic nerve in adult Lewis rats; In vitro: unprimed adult macrophages with SKPSC-conditioned medium | SKPSCs enhances expression of arg1 and an increases number of CD206+ macrophages. Nonetheless, these cells were associated with high levels of pro-inflammatory cytokines | [202] |
| Gene therapy—miRNA | |||
| miR-21 antagomir or miR-21 deletion | In vivo: spared nerve injury in mice; In vitro: PECs with exosome-enriched media from sensory neurons | miR-21-5p expression was associated with pro-inflammatory phenotype. Inhibition of miR-21-5p promoted M2 phenotype | [205] |
| EV-miR-23a | antagomir In vivo: spared nerve injury in mice; In vitro: coculture of DRG neurons with mouse macrophages | EV encapsulated miR-23a promoted M1 polarization. Inhibition of miR-23a reduced M1 macrophages numbers and increases M2 macrophages | [206] |
6. Biomaterial Approaches to Direct Macrophage Phenotype in CNS Repair
6.1. Macrophage-Based Clinical Trials in the CNS
6.2. Peripheral Nerve-Based Implants in the Lesioned Cord
6.3. Biomaterial Composition
6.3.1. Organic
6.3.2. Synthetic Polymers
6.4. Physical or Chemically Modified Biomaterials
6.4.1. Physical Modifications
6.4.2. Chemical Modifications
6.5. Therapeutic Molecule Incorporation
6.5.1. Small Molecule Drugs
6.5.2. Proteins/Macromolecules (Cytokines, Peptides, Antibodies, Nucleic Acid Cargoes)
6.5.3. Cellular Therapeutics
7. Future Directions
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Peng, J.; Han, G.H.; Ding, X.; Wei, S.; Gao, G.; Huang, K.; Chang, F.; Wang, Y. Role of macrophages in peripheral nerve injury and repair. Neural Regen. Res. 2019, 14, 1335–1342. [Google Scholar] [CrossRef]
- Chen, P.; Piao, X.; Bonaldo, P. Role of macrophages in Wallerian degeneration and axonal regeneration after peripheral nerve injury. Acta Neuropathol. 2015, 130, 605–618. [Google Scholar] [CrossRef]
- Büttner, R.; Schulz, A.; Reuter, M.; Akula, A.K.; Mindos, T.; Carlstedt, A.; Riecken, L.B.; Baader, S.L.; Bauer, R.; Morrison, H. Inflammaging impairs peripheral nerve maintenance and regeneration. Aging Cell 2018, 17, e12833. [Google Scholar] [CrossRef]
- Mietto, B.S.; Mostacada, K.; Martinez, A.M.B. Neurotrauma and inflammation: CNS and PNS responses. Mediat. Inflamm. 2015, 2015, 251204. [Google Scholar] [CrossRef] [PubMed]
- Villegas, R.; Martinez, N.W.; Lillo, J.; Pihan, P.; Hernandez, D.; Twiss, J.L.; Court, F.A. Calcium Release from Intra-Axonal Endoplasmic Reticulum Leads to Axon Degeneration through Mitochondrial Dysfunction. J. Neurosci. 2014, 34, 7179–7189. [Google Scholar] [CrossRef]
- Zhai, Q.; Wang, J.; Kim, A.; Liu, Q.; Watts, R.; Hoopfer, E.; Mitchison, T.; Luo, L.; He, Z. Involvement of the Ubiquitin-Proteasome System in the Early Stages of Wallerian Degeneration. Neuron 2003, 39, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Stoll, G.; Griffin, J.W.; Li, C.Y.; Trapp, B.D. Wallerian degeneration in the peripheral nervous system: Participation of both Schwann cells and macrophages in myelin degradation. J. Neurocytol. 1989, 18, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Trapp, B.D.; Hauer, P.; Lemke, G. Axonal regulation of myelin protein mRNA levels in actively myelinating Schwann cells. J. Neurosci. 1988, 8, 3515–3521. [Google Scholar] [CrossRef] [Green Version]
- White, F.V.; Toews, A.D.; Goodrum, J.F.; Novicki, D.L.; Bouldin, T.W.; Morell, P. Lipid Metabolism during Early Stages of Wallerian Degeneration in the Rat Sciatic Nerve. J. Neurochem. 1989, 52, 1085–1092. [Google Scholar] [CrossRef]
- Weerasuriya, A.; Mizisin, A.P. The Blood-Nerve Barrier: Structure and Functional Significance. In The Blood-Brain and Other Neural Barriers; Springer: Berlin/Heidelberg, Germany, 2011; pp. 149–173. [Google Scholar] [CrossRef]
- Vargas, M.E.; Barres, B.A. Why is Wallerian degeneration in the CNS so slow? Annu. Rev. Neurosci. 2007, 30, 153–179. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.K. Glial Membranes at the Node of Ranvier Prevent Neurite Outgrowth. Science 2005, 310, 1813–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.-L.; Lassner, F.; Bader, A.; Becker, M.; Walter, G.F.; Berger, A. Cellular activity of resident macrophages during Wallerian degeneration. Microsurgery 2000, 20, 255–261. [Google Scholar] [CrossRef]
- Friede, R.L.; Brück, W. Macrophage functional properties during myelin degradation. Adv. Neurol. 1993, 59, 327–336. [Google Scholar] [PubMed]
- Patton, B.L.; Miner, J.H.; Chiu, A.Y.; Sanes, J.R. Distribution and Function of Laminins in the Neuromuscular System of Developing, Adult, and Mutant Mice. J. Cell Biol. 1997, 139, 1507–1521. [Google Scholar] [CrossRef] [Green Version]
- Jessen, K.R.; Mirsky, R.; Lloyd, A.C. Schwann Cells: Development and Role in Nerve Repair. Cold Spring Harb. Perspect. Biol. 2015, 7, a020487. [Google Scholar] [CrossRef]
- Cattin, A.-L.; Burden, J.J.; Van Emmenis, L.; Mackenzie, F.E.; Hoving, J.J.A.; Calavia, N.G.; Guo, Y.; McLaughlin, M.; Rosenberg, L.H.; Quereda, V.; et al. Macrophage-Induced Blood Vessels Guide Schwann Cell-Mediated Regeneration of Peripheral Nerves. Cell 2015, 162, 1127–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, Q.T.; Sanes, J.R.; Lichtman, J.W. Pre-existing pathways promote precise projection patterns. Nat. Neurosci. 2002, 5, 861–867. [Google Scholar] [CrossRef]
- Gardiner, N.J.; Fernyhough, P.; Tomlinson, D.R.; Mayer, U.; von der Mark, H.; Streuli, C.H. α7 integrin mediates neurite outgrowth of distinct populations of adult sensory neurons. Mol. Cell. Neurosci. 2005, 28, 229–240. [Google Scholar] [CrossRef]
- Jurecka, W.; Ammerer, H.P.; Lassmann, H. Regeneration of a transected peripheral nerve. Acta Neuropathol. 1975, 32, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Martini, R.; Fischer, S.; López-Vales, R.; David, S. Interactions between Schwann cells and macrophages in injury and inherited demyelinating disease. Glia 2008, 56, 1566–1577. [Google Scholar] [CrossRef]
- Seitz, R.J.; Reiners, K.; Himmelmann, F.; Heininger, K.; Hartung, H.-P.; Toyka, K.V. The blood-nerve barrier in Wallerian degeneration: A sequential long-term study. Muscle Nerve 1989, 12, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-L.; Strickland, S. Laminin γ1 is critical for Schwann cell differentiation, axon myelination, and regeneration in the peripheral nerve. J. Cell Biol. 2003, 163, 889–899. [Google Scholar] [CrossRef]
- Fernandez-Valle, C.; Bunge, R.P.; Bunge, M.B. Schwann cells degrade myelin and proliferate in the absence of macrophages: Evidence fromin vitro studies of Wallerian degeneration. J. Neurocytol. 1995, 24, 667–679. [Google Scholar] [CrossRef]
- Chen, P.; Cescon, M.; Zuccolotto, G.; Nobbio, L.; Colombelli, C.; Filaferro, M.; Vitale, G.; Feltri, M.L.; Bonaldo, P. Collagen VI regulates peripheral nerve regeneration by modulating macrophage recruitment and polarization. Acta Neuropathol. 2015, 129, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Rotshenker, S. Wallerian degeneration: The innate-immune response to traumatic nerve injury. J. Neuroinflamm. 2011, 8, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tofaris, G.K.; Patterson, P.H.; Jessen, K.R.; Mirsky, R. Denervated Schwann Cells Attract Macrophages by Secretion of Leukemia Inhibitory Factor (LIF) and Monocyte Chemoattractant Protein-1 in a Process Regulated by Interleukin-6 and LIF. J. Neurosci. 2002, 22, 6696–6703. [Google Scholar] [CrossRef] [PubMed]
- Namikawa, K.; Okamoto, T.; Suzuki, A.; Konishi, H.; Kiyama, H. Pancreatitis-Associated Protein-III Is a Novel Macrophage Chemoattractant Implicated in Nerve Regeneration. J. Neurosci. 2006, 26, 7460–7467. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.D.; DeLeo, F.R. Neutrophil apoptosis and the resolution of infection. Immunol. Res. 2009, 43, 25–61. [Google Scholar] [CrossRef]
- Mueller, M.; Leonhard, C.; Wacker, K.; Ringelstein, E.B.; Okabe, M.; Hickey, W.F.; Kiefer, R. Macrophage Response to Peripheral Nerve Injury: The Quantitative Contribution of Resident and Hematogenous Macrophages. Lab. Investig. 2003, 83, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Rios, R.; Jablonka-Shariff, A.; Broberg, C.; Snyder-Warwick, A.K. Macrophage roles in peripheral nervous system injury and pathology: Allies in neuromuscular junction recovery. Mol. Cell. Neurosci. 2021, 111, 103590. [Google Scholar] [CrossRef]
- Ydens, E.; Amann, L.; Asselbergh, B.; Scott, C.L.; Martens, L.; Sichien, D.; Mossad, O.; Blank, T.; De Prijck, S.; Low, D.; et al. Profiling peripheral nerve macrophages reveals two macrophage subsets with distinct localization, transcriptome and response to injury. Nat. Neurosci. 2020, 23, 676–689. [Google Scholar] [CrossRef]
- Tomlinson, J.E.; Žygelyte, E.; Grenier, J.K.; Edwards, M.G.; Cheetham, J. Temporal changes in macrophage phenotype after peripheral nerve injury. J. Neuroinflamm. 2018, 15, 17–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, W.; Cao, L.; Khanabdali, R.; Kalionis, B.; Tai, X.; Xia, S. The Emerging Role of HMGB1 in Neuropathic Pain: A Potential Therapeutic Target for Neuroinflammation. J. Immunol. Res. 2016, 2016, 6430423. [Google Scholar] [CrossRef] [Green Version]
- Domoto, R.; Sekiguchi, F.; Tsubota, M.; Kawabata, A. Macrophage as a Peripheral Pain Regulator. Cells 2021, 10, 1881. [Google Scholar] [CrossRef]
- Silva, M.T. Macrophage phagocytosis of neutrophils at inflammatory/infectious foci: A cooperative mechanism in the control of infection and infectious inflammation. J. Leukoc. Biol. 2011, 89, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Fertala, J.; Rivlin, M.; Wang, M.L.; Beredjiklian, P.K.; Steplewski, A.; Fertala, A. Collagen-rich deposit formation in the sciatic nerve after injury and surgical repair: A study of collagen-producing cells in a rabbit model. J. Leukoc. Biol. 2020, 10, e01802. [Google Scholar] [CrossRef] [PubMed]
- Laskin, D.L. Macrophages and Inflammatory Mediators in Chemical Toxicity: A Battle of Forces. Chem. Res. Toxicol. 2009, 22, 1376–1385. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Bailey, W.M.; Braun, K.J.; Gensel, J.C. Age decreases macrophage IL-10 expression: Implications for functional recovery and tissue repair in spinal cord injury. Exp. Neurol. 2015, 273, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Nadeau, S.; Filali, M.; Zhang, J.; Kerr, B.J.; Rivest, S.; Soulet, D.; Iwakura, Y.; de Rivero Vaccari, J.P.; Keane, R.W.; Lacroix, S. Functional Recovery after Peripheral Nerve Injury is Dependent on the Pro-Inflammatory Cytokines IL-1β and TNF: Implications for Neuropathic Pain. J. Neurosci. 2011, 31, 12533–12542. [Google Scholar] [CrossRef] [PubMed]
- Stratton, J.; Shah, P. Macrophage polarization in nerve injury: Do Schwann cells play a role? Neural Regen. Res. 2016, 11, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Yoshiba, N.; Edanami, N.; Ohkura, N.; Maekawa, T.; Takahashi, N.; Tohma, A.; Izumi, K.; Maeda, T.; Hosoya, A.; Nakamura, H.; et al. M2 Phenotype Macrophages Colocalize with Schwann Cells in Human Dental Pulp. J. Dent. Res. 2020, 99, 329–338. [Google Scholar] [CrossRef]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Lurier, E.B.; Dalton, D.; Dampier, W.; Raman, P.; Nassiri, S.; Ferraro, N.M.; Rajagopalan, R.; Sarmady, M.; Spiller, K.L. Transcriptome analysis of IL-10-stimulated (M2c) macrophages by next-generation sequencing. Immunobiology 2017, 222, 847–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novak, M.L.; Koh, T.J. Macrophage phenotypes during tissue repair. J. Leukoc. Biol. 2013, 93, 875–881. [Google Scholar] [CrossRef] [Green Version]
- Gensel, J.C.; Zhang, B. Macrophage activation and its role in repair and pathology after spinal cord injury. Brain Res. 2015, 1619, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Painter, M.W.; Lutz, A.B.; Cheng, Y.-C.; Latremoliere, A.; Duong, K.; Miller, C.M.; Posada, S.; Cobos, E.J.; Zhang, A.X.; Wagers, A.J.; et al. Diminished Schwann Cell Repair Responses Underlie Age-Associated Impaired Axonal Regeneration. Neuron 2014, 83, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Moalem, G.; Xu, K.; Yu, L. T lymphocytes play a role in neuropathic pain following peripheral nerve injury in rats. Neuroscience 2004, 129, 767–777. [Google Scholar] [CrossRef]
- Gaudet, A.D.; Popovich, P.G.; Ramer, M.S. Wallerian degeneration: Gaining perspective on inflammatory events after peripheral nerve injury. J. Neuroinflamm. 2011, 8, 110. [Google Scholar] [CrossRef] [Green Version]
- Fawcett, J.W. The Struggle to Make CNS Axons Regenerate: Why Has It Been so Difficult? Neurochem. Res. 2020, 45, 144–158. [Google Scholar] [CrossRef] [Green Version]
- Silver, J. The glial scar is more than just astrocytes. Exp. Neurol. 2016, 286, 147–149. [Google Scholar] [CrossRef]
- Yang, T.; Dai, Y.; Chen, G.; Cui, S. Dissecting the Dual Role of the Glial Scar and Scar-Forming Astrocytes in Spinal Cord Injury. Front. Cell Neurosci. 2020, 14, 78. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, D.J.; Popovich, P.G. Inflammation and its role in neuroprotection, axonal regeneration and functional recovery after spinal cord injury. Exp. Neurol. 2008, 209, 378–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, J.C.; Norenberg, M.D.; Ramsay, D.A.; Dekaban, G.A.; Marcillo, A.E.; Saenz, A.D.; Pasquale-Styles, M.; Dietrich, W.D.; Weaver, L.C. The cellular inflammatory response in human spinal cords after injury. Brain 2006, 129, 3249–3269. [Google Scholar] [CrossRef] [PubMed]
- George, R.; Griffin, J.W. Delayed macrophage responses and myelin clearance during Wallerian degeneration in the central nervous system: The dorsal radiculotomy model. Exp. Neurol. 1994, 129, 225–236. [Google Scholar] [CrossRef]
- Hill, C.E. A view from the ending: Axonal dieback and regeneration following SCI. Neurosci. Lett. 2017, 652, 11–24. [Google Scholar] [CrossRef]
- Evans, T.A.; Barkauskas, D.S.; Myers, J.T.; Hare, E.G.; You, J.Q.; Ransohoff, R.M.; Huang, A.Y.; Silver, J. High-resolution intravital imaging reveals that blood-derived macrophages but not resident microglia facilitate secondary axonal dieback in traumatic spinal cord injury. Exp. Neurol. 2014, 254, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Horn, K.P.; Busch, S.A.; Hawthorne, A.L.; van Rooijen, N.; Silver, J. Another barrier to regeneration in the CNS: Activated macrophages induce extensive retraction of dystrophic axons through direct physical interactions. J. Neurosci. 2008, 28, 9330–9341. [Google Scholar] [CrossRef]
- Busch, S.A.; Horn, K.P.; Silver, D.J.; Silver, J. Overcoming macrophage-mediated axonal dieback following CNS injury. J. Neurosci. 2009, 29, 9967–9976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollis, E.R., II; Zou, Y. Reinduced Wnt signaling limits regenerative potential of sensory axons in the spinal cord following conditioning lesion. Proc. Natl. Acad. Sci. USA 2012, 109, 14663–14668. [Google Scholar] [CrossRef] [Green Version]
- Gensel, J.C.; Wang, Y.; Guan, Z.; Beckwith, K.A.; Braun, K.J.; Wei, P.; McTigue, D.M.; Popovich, P.G. Toll-Like Receptors and Dectin-1, a C-Type Lectin Receptor, Trigger Divergent Functions in CNS Macrophages. J. Neurosci. 2015, 35, 9966–9976. [Google Scholar] [CrossRef] [Green Version]
- McPhail, L.T.; Stirling, D.P.; Tetzlaff, W.; Kwiecien, J.M.; Ramer, M.S. The contribution of activated phagocytes and myelin degeneration to axonal retraction/dieback following spinal cord injury. Eur. J. Neurosci. 2004, 20, 1984–1994. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Kobayakawa, K.; Ohkawa, Y.; Kumamaru, H.; Yokota, K.; Saito, T.; Kijima, K.; Yoshizaki, S.; Harimaya, K.; Nakashima, Y.; et al. Interaction of reactive astrocytes with type I collagen induces astrocytic scar formation through the integrin-N-cadherin pathway after spinal cord injury. Nat. Med. 2017, 23, 818–828. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, T.M.; Wollenberg, A.L.; Kim, J.H.; Ao, Y.; Deming, T.J.; Sofroniew, M.V. Foreign body responses in mouse central nervous system mimic natural wound responses and alter biomaterial functions. Nat. Commun. 2020, 11, 6203. [Google Scholar] [CrossRef] [PubMed]
- Kawano, H.; Kimura-Kuroda, J.; Komuta, Y.; Yoshioka, N.; Li, H.P.; Kawamura, K.; Li, Y.; Raisman, G. Role of the lesion scar in the response to damage and repair of the central nervous system. Cell Tissue Res. 2012, 349, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [Green Version]
- Wanner, I.B.; Anderson, M.A.; Song, B.; Levine, J.; Fernandez, A.; Gray-Thompson, Z.; Ao, Y.; Sofroniew, M.V. Glial scar borders are formed by newly proliferated, elongated astrocytes that interact to corral inflammatory and fibrotic cells via STAT3-dependent mechanisms after spinal cord injury. J. Neurosci. 2013, 33, 12870–12886. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V. Astrocyte barriers to neurotoxic inflammation. Nat. Rev. Neurosci. 2015, 16, 249–263. [Google Scholar] [CrossRef] [Green Version]
- Carbonell, A.L.; Boya, J. Ultrastructural study on meningeal regeneration and meningo-glial relationships after cerebral stab wound in the adult rat. Brain Res. 1988, 439, 337–344. [Google Scholar] [CrossRef]
- Soderblom, C.; Luo, X.; Blumenthal, E.; Bray, E.; Lyapichev, K.; Ramos, J.; Krishnan, V.; Lai-Hsu, C.; Park, K.K.; Tsoulfas, P.; et al. Perivascular fibroblasts form the fibrotic scar after contusive spinal cord injury. J. Neurosci. 2013, 33, 13882–13887. [Google Scholar] [CrossRef] [Green Version]
- Goritz, C.; Dias, D.O.; Tomilin, N.; Barbacid, M.; Shupliakov, O.; Frisen, J. A pericyte origin of spinal cord scar tissue. Science 2011, 333, 238–242. [Google Scholar] [CrossRef]
- Mathewson, A.J.; Berry, M. Observations on the astrocyte response to a cerebral stab wound in adult rats. Brain Res. 1985, 327, 61–69. [Google Scholar] [CrossRef]
- Shearer, M.C.; Fawcett, J.W. The astrocyte/meningeal cell interface--a barrier to successful nerve regeneration? Cell Tissue Res. 2001, 305, 267–273. [Google Scholar] [CrossRef]
- Cregg, J.M.; DePaul, M.A.; Filous, A.R.; Lang, B.T.; Tran, A.; Silver, J. Functional regeneration beyond the glial scar. Exp. Neurol. 2014, 253, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, S.; Bahr, M. Immunocytochemical characterization of reactive optic nerve astrocytes and meningeal cells. Glia 1999, 26, 36–46. [Google Scholar] [CrossRef]
- Davies, S.J.; Goucher, D.R.; Doller, C.; Silver, J. Robust regeneration of adult sensory axons in degenerating white matter of the adult rat spinal cord. J. Neurosci. 1999, 19, 5810–5822. [Google Scholar] [CrossRef] [PubMed]
- Stichel, C.C.; Hermanns, S.; Luhmann, H.J.; Lausberg, F.; Niermann, H.; D’Urso, D.; Servos, G.; Hartwig, H.G.; Muller, H.W. Inhibition of collagen IV deposition promotes regeneration of injured CNS axons. Eur. J. Neurosci. 1999, 11, 632–646. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Soderblom, C.; Krishnan, V.; Ashbaugh, J.; Bethea, J.R.; Lee, J.K. Hematogenous macrophage depletion reduces the fibrotic scar and increases axonal growth after spinal cord injury. Neurobiol. Dis. 2015, 74, 114–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cholas, R.H.; Hsu, H.P.; Spector, M. The reparative response to cross-linked collagen-based scaffolds in a rat spinal cord gap model. Biomaterials 2012, 33, 2050–2059. [Google Scholar] [CrossRef]
- Altinova, H.; Hammes, S.; Palm, M.; Gerardo-Nava, J.; Achenbach, P.; Deumens, R.; Hermans, E.; Fuhrmann, T.; Boecker, A.; van Neerven, S.G.A.; et al. Fibroadhesive scarring of grafted collagen scaffolds interferes with implant-host neural tissue integration and bridging in experimental spinal cord injury. Regen. Biomater. 2019, 6, 75–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witherel, C.E.; Abebayehu, D.; Barker, T.H.; Spiller, K.L. Macrophage and Fibroblast Interactions in Biomaterial-Mediated Fibrosis. Adv. Healthc. Mater. 2019, 8, e1801451. [Google Scholar] [CrossRef]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altinova, H.; Hammes, S.; Palm, M.; Achenbach, P.; Gerardo-Nava, J.; Deumens, R.; Fuhrmann, T.; van Neerven, S.G.A.; Hermans, E.; Weis, J.; et al. Dense fibroadhesive scarring and poor blood vessel-maturation hamper the integration of implanted collagen scaffolds in an experimental model of spinal cord injury. Biomed. Mater. 2020, 15, 015012. [Google Scholar] [CrossRef]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science 2010, 330, 841–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldmann, T.; Wieghofer, P.; Jordao, M.J.; Prutek, F.; Hagemeyer, N.; Frenzel, K.; Amann, L.; Staszewski, O.; Kierdorf, K.; Krueger, M.; et al. Origin, fate and dynamics of macrophages at central nervous system interfaces. Nat. Immunol. 2016, 17, 797–805. [Google Scholar] [CrossRef]
- Hines, D.J.; Hines, R.M.; Mulligan, S.J.; Macvicar, B.A. Microglia processes block the spread of damage in the brain and require functional chloride channels. Glia 2009, 57, 1610–1618. [Google Scholar] [CrossRef]
- Davalos, D.; Grutzendler, J.; Yang, G.; Kim, J.V.; Zuo, Y.; Jung, S.; Littman, D.R.; Dustin, M.L.; Gan, W.B. ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci. 2005, 8, 752–758. [Google Scholar] [CrossRef]
- Haynes, S.E.; Hollopeter, G.; Yang, G.; Kurpius, D.; Dailey, M.E.; Gan, W.B.; Julius, D. The P2Y12 receptor regulates microglial activation by extracellular nucleotides. Nat. Neurosci. 2006, 9, 1512–1519. [Google Scholar] [CrossRef]
- Wu, L.J.; Vadakkan, K.I.; Zhuo, M. ATP-induced chemotaxis of microglial processes requires P2Y receptor-activated initiation of outward potassium currents. Glia 2007, 55, 810–821. [Google Scholar] [CrossRef]
- Ohsawa, K.; Irino, Y.; Sanagi, T.; Nakamura, Y.; Suzuki, E.; Inoue, K.; Kohsaka, S. P2Y12 receptor-mediated integrin-beta1 activation regulates microglial process extension induced by ATP. Glia 2010, 58, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Franke, H.; Sauer, C.; Rudolph, C.; Krugel, U.; Hengstler, J.G.; Illes, P. P2 receptor-mediated stimulation of the PI3-K/Akt-pathway in vivo. Glia 2009, 57, 1031–1045. [Google Scholar] [CrossRef]
- Irino, Y.; Nakamura, Y.; Inoue, K.; Kohsaka, S.; Ohsawa, K. Akt activation is involved in P2Y12 receptor-mediated chemotaxis of microglia. J. Neurosci. Res. 2008, 86, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Ceruti, S.; Villa, G.; Genovese, T.; Mazzon, E.; Longhi, R.; Rosa, P.; Bramanti, P.; Cuzzocrea, S.; Abbracchio, M.P. The P2Y-like receptor GPR17 as a sensor of damage and a new potential target in spinal cord injury. Brain 2009, 132, 2206–2218. [Google Scholar] [CrossRef] [Green Version]
- Dibaj, P.; Nadrigny, F.; Steffens, H.; Scheller, A.; Hirrlinger, J.; Schomburg, E.D.; Neusch, C.; Kirchhoff, F. NO mediates microglial response to acute spinal cord injury under ATP control in vivo. Glia 2010, 58, 1133–1144. [Google Scholar] [CrossRef]
- Drago, F.; Sautiere, P.E.; Le Marrec-Croq, F.; Accorsi, A.; Van Camp, C.; Salzet, M.; Lefebvre, C.; Vizioli, J. Microglia of medicinal leech (Hirudo medicinalis) express a specific activation marker homologous to vertebrate ionized calcium-binding adapter molecule 1 (Iba1/alias aif-1). Dev. Neurobiol. 2014, 74, 987–1001. [Google Scholar] [CrossRef] [PubMed]
- Tahtouh, M.; Croq, F.; Vizioli, J.; Sautiere, P.E.; Van Camp, C.; Salzet, M.; Daha, M.R.; Pestel, J.; Lefebvre, C. Evidence for a novel chemotactic C1q domain-containing factor in the leech nerve cord. Mol. Immunol. 2009, 46, 523–531. [Google Scholar] [CrossRef]
- Pineau, I.; Sun, L.; Bastien, D.; Lacroix, S. Astrocytes initiate inflammation in the injured mouse spinal cord by promoting the entry of neutrophils and inflammatory monocytes in an IL-1 receptor/MyD88-dependent fashion. Brain Behav. Immun. 2010, 24, 540–553. [Google Scholar] [CrossRef]
- Shechter, R.; London, A.; Varol, C.; Raposo, C.; Cusimano, M.; Yovel, G.; Rolls, A.; Mack, M.; Pluchino, S.; Martino, G.; et al. Infiltrating blood-derived macrophages are vital cells playing an anti-inflammatory role in recovery from spinal cord injury in mice. PLoS Med. 2009, 6, e1000113. [Google Scholar] [CrossRef]
- Mawhinney, L.A.; Thawer, S.G.; Lu, W.Y.; Rooijen, N.; Weaver, L.C.; Brown, A.; Dekaban, G.A. Differential detection and distribution of microglial and hematogenous macrophage populations in the injured spinal cord of lys-EGFP-ki transgenic mice. J. Neuropathol. Exp. Neurol. 2012, 71, 180–197. [Google Scholar] [CrossRef] [Green Version]
- Greenhalgh, A.D.; David, S. Differences in the phagocytic response of microglia and peripheral macrophages after spinal cord injury and its effects on cell death. J. Neurosci. 2014, 34, 6316–6322. [Google Scholar] [CrossRef]
- Rolls, A.; Shechter, R.; London, A.; Segev, Y.; Jacob-Hirsch, J.; Amariglio, N.; Rechavi, G.; Schwartz, M. Two faces of chondroitin sulfate proteoglycan in spinal cord repair: A role in microglia/macrophage activation. PLoS Med. 2008, 5, e171. [Google Scholar] [CrossRef]
- Shechter, R.; Raposo, C.; London, A.; Sagi, I.; Schwartz, M. The glial scar-monocyte interplay: A pivotal resolution phase in spinal cord repair. PLoS ONE 2011, 6, e27969. [Google Scholar] [CrossRef]
- Kawashima, H.; Hirose, M.; Hirose, J.; Nagakubo, D.; Plaas, A.H.; Miyasaka, M. Binding of a large chondroitin sulfate/dermatan sulfate proteoglycan, versican, to L-selectin, P-selectin, and CD44. J. Biol. Chem. 2000, 275, 35448–35456. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, T.; Kawashima, H.; Tanaka, T.; Hirose, M.; Toyama-Sorimachi, N.; Matsuzawa, Y.; Miyasaka, M. CD44 binds a chondroitin sulfate proteoglycan, aggrecan. Int. Immunol. 2001, 13, 359–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.L.; Liu, Z.; Shen, J.; Werner, A.; Kreutzberg, G.W.; Raivich, G. Regulation of the cell adhesion molecule CD44 after nerve transection and direct trauma to the mouse brain. J. Comp. Neurol. 2000, 426, 468–492. [Google Scholar] [CrossRef]
- Kloss, C.U.; Werner, A.; Klein, M.A.; Shen, J.; Menuz, K.; Probst, J.C.; Kreutzberg, G.W.; Raivich, G. Integrin family of cell adhesion molecules in the injured brain: Regulation and cellular localization in the normal and regenerating mouse facial motor nucleus. J. Comp. Neurol. 1999, 411, 162–178. [Google Scholar] [CrossRef]
- Kroner, A.; Greenhalgh, A.D.; Zarruk, J.G.; Passos Dos Santos, R.; Gaestel, M.; David, S. TNF and increased intracellular iron alter macrophage polarization to a detrimental M1 phenotype in the injured spinal cord. Neuron 2014, 83, 1098–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longbrake, E.E.; Lai, W.; Ankeny, D.P.; Popovich, P.G. Characterization and modeling of monocyte-derived macrophages after spinal cord injury. J. Neurochem. 2007, 102, 1083–1094. [Google Scholar] [CrossRef]
- Kigerl, K.A.; Popovich, P.G. Toll-like receptors in spinal cord injury. Curr. Top. Microbiol. Immunol. 2009, 336, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Kigerl, K.A.; Gensel, J.C.; Ankeny, D.P.; Alexander, J.K.; Donnelly, D.J.; Popovich, P.G. Identification of two distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in the injured mouse spinal cord. J. Neurosci. 2009, 29, 13435–13444. [Google Scholar] [CrossRef] [Green Version]
- Pineau, I.; Lacroix, S. Proinflammatory cytokine synthesis in the injured mouse spinal cord: Multiphasic expression pattern and identification of the cell types involved. J. Comp. Neurol. 2007, 500, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Blumbergs, P.C.; Jones, N.R.; Manavis, J.; Sarvestani, G.T.; Ghabriel, M.N. Early expression and cellular localization of proinflammatory cytokines interleukin-1beta, interleukin-6, and tumor necrosis factor-alpha in human traumatic spinal cord injury. Spine 2004, 29, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Piccinini, A.M.; Midwood, K.S. DAMPening inflammation by modulating TLR signalling. Mediat. Inflamm. 2010, 2010, 672395. [Google Scholar] [CrossRef] [Green Version]
- Katsumoto, A.; Lu, H.; Miranda, A.S.; Ransohoff, R.M. Ontogeny and functions of central nervous system macrophages. J. Immunol. 2014, 193, 2615–2621. [Google Scholar] [CrossRef]
- Varnum, M.M.; Ikezu, T. The classification of microglial activation phenotypes on neurodegeneration and regeneration in Alzheimer’s disease brain. Arch. Immunol. Ther. Exp. 2012, 60, 251–266. [Google Scholar] [CrossRef]
- Rapalino, O.; Lazarov-Spiegler, O.; Agranov, E.; Velan, G.J.; Yoles, E.; Fraidakis, M.; Solomon, A.; Gepstein, R.; Katz, A.; Belkin, M.; et al. Implantation of stimulated homologous macrophages results in partial recovery of paraplegic rats. Nat. Med. 1998, 4, 814–821. [Google Scholar] [CrossRef]
- Knoller, N.; Auerbach, G.; Fulga, V.; Zelig, G.; Attias, J.; Bakimer, R.; Marder, J.B.; Yoles, E.; Belkin, M.; Schwartz, M.; et al. Clinical experience using incubated autologous macrophages as a treatment for complete spinal cord injury: Phase I study results. J. Neurosurg. Spine 2005, 3, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, G.; Theodorou, D.; Kendirli, A.; Jordao, M.J.C.; Staszewski, O.; Phulphagar, K.; Cantuti-Castelvetri, L.; Dagkalis, A.; Bessis, A.; Simons, M.; et al. Mononuclear phagocytes locally specify and adapt their phenotype in a multiple sclerosis model. Nat. Neurosci. 2018, 21, 1196–1208. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Galli, S.J.; Borregaard, N.; Wynn, T.A. Phenotypic and functional plasticity of cells of innate immunity: Macrophages, mast cells and neutrophils. Nat. Immunol. 2011, 12, 1035–1044. [Google Scholar] [CrossRef] [Green Version]
- Lammertse, D.P.; Jones, L.A.; Charlifue, S.B.; Kirshblum, S.C.; Apple, D.F.; Ragnarsson, K.T.; Falci, S.P.; Heary, R.F.; Choudhri, T.F.; Jenkins, A.L.; et al. Autologous incubated macrophage therapy in acute, complete spinal cord injury: Results of the phase 2 randomized controlled multicenter trial. Spinal Cord 2012, 50, 661–671. [Google Scholar] [CrossRef]
- Yoon, S.H.; Shim, Y.S.; Park, Y.H.; Chung, J.K.; Nam, J.H.; Kim, M.O.; Park, H.C.; Park, S.R.; Min, B.H.; Kim, E.Y.; et al. Complete spinal cord injury treatment using autologous bone marrow cell transplantation and bone marrow stimulation with granulocyte macrophage-colony stimulating factor: Phase I/II clinical trial. Stem Cells 2007, 25, 2066–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.A.; Kumar, S.R.; Narayanan, R.; Arul, K.; Baskaran, M. Autologous bone marrow derived mononuclear cell therapy for spinal cord injury: A phase I/II clinical safety and primary efficacy data. Exp. Clin. Transplant. 2009, 7, 241–248. [Google Scholar]
- London, A.; Itskovich, E.; Benhar, I.; Kalchenko, V.; Mack, M.; Jung, S.; Schwartz, M. Neuroprotection and progenitor cell renewal in the injured adult murine retina requires healing monocyte-derived macrophages. J. Exp. Med. 2011, 208, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J. The primary mechanism of the IL-10-regulated antiinflammatory response is to selectively inhibit transcription. Proc. Natl. Acad. Sci. USA 2005, 102, 8686–8691. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, K.D.; Dreyfus, C.F.; Black, I.B. Brain-derived neurotrophic factor in astrocytes, oligodendrocytes, and microglia/macrophages after spinal cord injury. Neurobiol. Dis. 2000, 7, 574–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Henzl, M.T.; Lorber, B.; Nakazawa, T.; Thomas, T.T.; Jiang, F.; Langer, R.; Benowitz, L.I. Oncomodulin is a macrophage-derived signal for axon regeneration in retinal ganglion cells. Nat. Neurosci. 2006, 9, 843–852. [Google Scholar] [CrossRef]
- Kokaia, Z.; Martino, G.; Schwartz, M.; Lindvall, O. Cross-talk between neural stem cells and immune cells: The key to better brain repair? Nat. Neurosci. 2012, 15, 1078–1087. [Google Scholar] [CrossRef]
- Martino, G. How the brain repairs itself: New therapeutic strategies in inflammatory and degenerative CNS disorders. Lancet Neurol. 2004, 3, 372–378. [Google Scholar] [CrossRef]
- Miron, V.E.; Boyd, A.; Zhao, J.W.; Yuen, T.J.; Ruckh, J.M.; Shadrach, J.L.; van Wijngaarden, P.; Wagers, A.J.; Williams, A.; Franklin, R.J.M.; et al. M2 microglia and macrophages drive oligodendrocyte differentiation during CNS remyelination. Nat. Neurosci. 2013, 16, 1211–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirling, D.P.; Cummins, K.; Wayne Chen, S.R.; Stys, P. Axoplasmic reticulum Ca2+ release causes secondary degeneration of spinal axons. Ann. Neurol. 2014, 75, 220–229. [Google Scholar] [CrossRef]
- Haan, N.; Zhu, B.; Wang, J.; Wei, X.; Song, B. Crosstalk between macrophages and astrocytes affects proliferation, reactive phenotype and inflammatory response, suggesting a role during reactive gliosis following spinal cord injury. J. Neuroinflamm. 2015, 12, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.M.; Gordon, T.; Zochodne, D.W.; Power, H.A. Improving peripheral nerve regeneration: From molecular mechanisms to potential therapeutic targets. Exp. Neurol. 2014, 261, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Houshyar, S.; Bhattacharyya, A.; Shanks, R. Peripheral Nerve Conduit: Materials and Structures. ACS Chem. Neurosci. 2019, 10, 3349–3365. [Google Scholar] [CrossRef]
- Riccio, M.; Marchesini, A.; Pugliese, P.; De Francesco, F. Nerve repair and regeneration: Biological tubulization limits and future perspectives. J. Cell. Physiol. 2019, 234, 3362–3375. [Google Scholar] [CrossRef] [PubMed]
- Terzis, J.K.; Skoulis, T.G.; Soucacos, P.N. Vascularized nerve grafts. A review. Int. Angiol. A J. Int. Union Angiol. 1995, 14, 264–277. [Google Scholar] [CrossRef]
- Vasileiadis, A. Bridging of Peripheral Nerve Defects by Autologous Nerve Grafting Personal Experience. MOJ Orthop Rheumatol. 2016, 5, 00183. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, T.; Mackay, M.; Midha, R. Peripheral Nerve Repair and Grafting Techniques: A Review. Neurol. Med.-Chir. 2000, 40, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Roballo, K.C.S.; Bushman, J. Evaluation of the host immune response and functional recovery in peripheral nerve autografts and allografts. Transplant. Immunol. 2019, 53, 61–71. [Google Scholar] [CrossRef]
- Fansa, H.; Schneider, W.; Keilhoff, G. Revascularization of tissue-engineered nerve grafts and invasion of macrophages. Tissue Eng 2001, 7, 519–524. [Google Scholar] [CrossRef]
- Brooks, D.N.; Weber, R.V.; Chao, J.D.; Rinker, B.D.; Zoldos, J.; Robichaux, M.R.; Ruggeri, S.B.; Anderson, K.A.; Bonatz, E.E.; Wisotsky, S.M.; et al. Processed nerve allografts for peripheral nerve reconstruction: A multicenter study of utilization and outcomes in sensory, mixed, and motor nerve reconstructions. Microsurgery 2012, 32, 1–14. [Google Scholar] [CrossRef]
- Sridharan, R.; Reilly, R.B.; Buckley, C.T. Decellularized grafts with axially aligned channels for peripheral nerve regeneration. J. Mech. Behav. Biomed. Mater. 2015, 41, 124–135. [Google Scholar] [CrossRef]
- Rinker, B.; Zoldos, J.; Weber, R.V.; Ko, J.; Thayer, W.; Greenberg, J.; Leversedge, F.J.; Safa, B.; Buncke, G. Use of Processed Nerve Allografts to Repair Nerve Injuries Greater Than 25 mm in the Hand. Ann. Plast. Surg. 2017, 78, S292–S295. [Google Scholar] [CrossRef]
- Isaacs, J.; Safa, B. A Preliminary Assessment of the Utility of Large-Caliber Processed Nerve Allografts for the Repair of Upper Extremity Nerve Injuries. Hand 2017, 12, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Safa, B.; Jain, S.; Desai, M.J.; Greenberg, J.A.; Niacaris, T.R.; Nydick, J.A.; Leversedge, F.J.; Megee, D.M.; Zoldos, J.; Rinker, B.D.; et al. Peripheral nerve repair throughout the body with processed nerve allografts: Results from a large multicenter study. Microsurgery 2020, 40, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Safa, B.; Buncke, G. Autograft Substitutes: Conduits and Processed Nerve Allografts. Hand Clin. 2016, 32, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Hunter, D.A.; Schellhardt, L.; Jo, S.; Santosa, K.B.; Larson, E.L.; Fuchs, A.G.; Snyder-Warwick, A.K.; Mackinnon, S.E.; Wood, M.D. The accumulation of T cells within acellular nerve allografts is length-dependent and critical for nerve regeneration. Exp. Neurol. 2019, 318, 216–231. [Google Scholar] [CrossRef]
- Kehoe, S.; Zhang, X.F.; Boyd, D. FDA approved guidance conduits and wraps for peripheral nerve injury: A review of materials and efficacy. Injury 2012, 43, 553–572. [Google Scholar] [CrossRef] [PubMed]
- Lackington, W.A.; Ryan, A.J.; O’Brien, F.J. Advances in Nerve Guidance Conduit-Based Therapeutics for Peripheral Nerve Repair. ACS Biomater. Sci. Eng. 2017, 3, 1221–1235. [Google Scholar] [CrossRef] [PubMed]
- Gaudin, R.; Knipfer, C.; Henningsen, A.; Smeets, R.; Heiland, M.; Hadlock, T. Approaches to Peripheral Nerve Repair: Generations of Biomaterial Conduits Yielding to Replacing Autologous Nerve Grafts in Craniomaxillofacial Surgery. BioMed Res. Int. 2016, 2016, 3856262. [Google Scholar] [CrossRef] [Green Version]
- Fornasari, B.E.; Carta, G.; Gambarotta, G.; Raimondo, S. Natural-Based Biomaterials for Peripheral Nerve Injury Repair. Front. Bioeng. Biotechnol. 2020, 8, 554257. [Google Scholar] [CrossRef]
- Pogrel, M.A.; McDonald, A.R.; Kaban, L.B. Gore-Tex tubing as a conduit for repair of lingual and inferior alveolar nerve continuity defects: A preliminary report. J. Oral. Maxillofac. Surg. 1998, 56, 319–321. [Google Scholar] [CrossRef]
- Rbia, N.; Bulstra, L.F.; Saffari, T.M.; Hovius, S.E.R.; Shin, A.Y. Collagen Nerve Conduits and Processed Nerve Allografts for the Reconstruction of Digital Nerve Gaps: A Single-Institution Case Series and Review of the Literature. World Neurosurg. 2019, 127, e1176–e1184. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, Z.; Brooks, P.J.; Barzilay, O.; Fine, N.; Glogauer, M. Macrophages, Foreign Body Giant Cells and Their Response to Implantable Biomaterials. Materials 2015, 8, 5671–5701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, K.E.; García, A.J. Macrophage phenotypes in tissue repair and the foreign body response: Implications for biomaterial-based regenerative medicine strategies. Acta Biomater. 2021, 133, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Chung, L.; Maestas, D.R.; Housseau, F.; Elisseeff, J.H. Key players in the immune response to biomaterial scaffolds for regenerative medicine. Adv. Drug Deliv. Rev. 2017, 114, 184–192. [Google Scholar] [CrossRef]
- Vora, A.R.; Bodell, S.M.; Loescher, A.R.; Smith, K.G.; Robinson, P.P.; Boissonade, F.M. Inflammatory cell accumulation in traumatic neuromas of the human lingual nerve. Arch. Oral Biol. 2007, 52, 74–82. [Google Scholar] [CrossRef]
- Frisén, J.; Risling, M.; Fried, K. Distribution and axonal relations of macrophages in a neuroma. Neuroscience 1993, 55, 1003–1013. [Google Scholar] [CrossRef]
- Meek, M.F.; Den Dunnen, W.F.A. Porosity of the wall of a Neurolac® nerve conduit hampers nerve regeneration. Microsurgery 2009, 29, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Shintani, K.; Uemura, T.; Takamatsu, K.; Yokoi, T.; Onode, E.; Okada, M.; Nakamura, H. Protective effect of biodegradable nerve conduit against peripheral nerve adhesion after neurolysis. J. Neurosurg. JNS 2018, 129, 815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenberg, L.; Stößel, M.; Ronchi, G.; Geuna, S.; Yin, Y.; Mommert, S.; Mårtensson, L.; Metzen, J.; Grothe, C.; Dahlin, L.B. Regeneration of long-distance peripheral nerve defects after delayed reconstruction in healthy and diabetic rats is supported by immunomodulatory chitosan nerve guides. BMC Neurosci. 2017, 18, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Zhang, L.; Wei, Y.; Chen, T.; Ji, X.; Ye, K.; Yu, J.; Tang, B.; Sun, X.; Hu, J. Human hair keratins promote the regeneration of peripheral nerves in a rat sciatic nerve crush model. J. Mater. Sci. Mater. Med. 2019, 30, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, D.; Zhou, L.; Zheng, X.; Hu, Y. Sustained release of collagen VI potentiates sciatic nerve regeneration by modulating macrophage phenotype. Eur. J. Neurosci. 2017, 45, 1258–1267. [Google Scholar] [CrossRef]
- Fearing, B.V.; Van Dyke, M.E. In vitro response of macrophage polarization to a keratin biomaterial. Acta Biomater. 2014, 10, 3136–3144. [Google Scholar] [CrossRef] [PubMed]
- Rowley, A.T.; Nagalla, R.R.; Wang, S.W.; Liu, W.F. Extracellular Matrix-Based Strategies for Immunomodulatory Biomaterials Engineering. Adv. Healthc. Mater. 2019, 8, 1801578. [Google Scholar] [CrossRef]
- Rowley, A.T.; Meli, V.S.; Wu-Woods, N.J.; Chen, E.Y.; Liu, W.F.; Wang, S.-W. Effects of Surface-Bound Collagen-Mimetic Peptides on Macrophage Uptake and Immunomodulation. Front. Bioeng. Biotechnol. 2020, 8, 747. [Google Scholar] [CrossRef]
- Hsieh, J.Y.; Smith, T.D.; Meli, V.S.; Tran, T.N.; Botvinick, E.L.; Liu, W.F. Differential regulation of macrophage inflammatory activation by fibrin and fibrinogen. Acta Biomater. 2017, 47, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Gregory, H.; Phillips, J.B. Materials for peripheral nerve repair constructs: Natural proteins or synthetic polymers? Neurochem. Int. 2021, 143, 104953. [Google Scholar] [CrossRef]
- Nikkhah, M.; Edalat, F.; Manoucheri, S.; Khademhosseini, A. Engineering microscale topographies to control the cell–substrate interface. Biomaterials 2012, 33, 5230–5246. [Google Scholar] [CrossRef] [Green Version]
- Sarhane, K.A.; Ibrahim, Z.; Martin, R.; Krick, K.; Cashman, C.R.; Tuffaha, S.H.; Broyles, J.M.; Prasad, N.; Yao, Z.-C.; Cooney, D.S. Macroporous nanofiber wraps promote axonal regeneration and functional recovery in nerve repair by limiting fibrosis. Acta Biomater. 2019, 88, 332–345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yao, Y.; Duan, Y.; Yu, X.; Shi, H.; Nakkala, J.R.; Zuo, X.; Hong, L.; Mao, Z.; Gao, C. Surface-Anchored Graphene Oxide Nanosheets on Cell-Scale Micropatterned Poly(D,L-lactide-co-caprolactone) Conduits Promote Peripheral Nerve Regeneration. ACS Appl. Mater. Interfaces 2020, 12, 7915–7930. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Ding, F.; Williams, D.F. Neural tissue engineering options for peripheral nerve regeneration. Biomaterials 2014, 35, 6143–6156. [Google Scholar] [CrossRef]
- Sridharan, R.; Cameron, A.R.; Kelly, D.J.; Kearney, C.J.; O’Brien, F.J. Biomaterial based modulation of macrophage polarization: A review and suggested design principles. Mater. Today 2015, 18, 313–325. [Google Scholar] [CrossRef]
- McWhorter, F.Y.; Wang, T.; Nguyen, P.; Chung, T.; Liu, W.F. Modulation of macrophage phenotype by cell shape. Proc. Natl. Acad. Sci. USA 2013, 110, 17253–17258. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Yang, W.; Zhang, K.; Qiu, S.; Xu, J.; Wang, C.; Chai, Y. Nanofiber arrangement regulates peripheral nerve regeneration through differential modulation of macrophage phenotypes. Acta Biomater. 2019, 83, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Bartneck, M.; Schulte, V.A.; Paul, N.E.; Diez, M.; Lensen, M.C.; Zwadlo-Klarwasser, G. Induction of specific macrophage subtypes by defined micro-patterned structures. Acta Biomater. 2010, 6, 3864–3872. [Google Scholar] [CrossRef]
- Blakney, A.K.; Swartzlander, M.D.; Bryant, S.J. The effects of substrate stiffness on the in vitro activation of macrophages and in vivo host response to poly(ethylene glycol)-based hydrogels. J. Biomed. Mater. Res. A 2012, 100, 1375–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridharan, R.; Ryan, E.J.; Kearney, C.J.; Kelly, D.J.; O’Brien, F.J. Macrophage Polarization in Response to Collagen Scaffold Stiffness Is Dependent on Cross-Linking Agent Used To Modulate the Stiffness. ACS Biomater. Sci. Eng. 2019, 5, 544–552. [Google Scholar] [CrossRef]
- Saha, K.; Keung, A.J.; Irwin, E.F.; Li, Y.; Little, L.; Schaffer, D.V.; Healy, K.E. Substrate Modulus Directs Neural Stem Cell Behavior. Biophys. J. 2008, 95, 4426–4438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayal, C.; Moeendarbary, E.; Shipley, R.J.; Phillips, J.B. Mechanical Response of Neural Cells to Physiologically Relevant Stiffness Gradients. Adv. Healthc. Mater. 2020, 9, 1901036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Zhang, L.; Dong, J.; Wang, S. Photocured Biodegradable Polymer Substrates of Varying Stiffness and Microgroove Dimensions for Promoting Nerve Cell Guidance and Differentiation. Langmuir 2012, 28, 12557–12568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Ji, Y.; Zhao, Y.; Liu, Y.; Ding, F.; Gu, X.; Yang, Y. The influence of substrate stiffness on the behavior and functions of Schwann cells in culture. Biomaterials 2012, 33, 6672–6681. [Google Scholar] [CrossRef]
- Rosso, G.; Wehner, D.; Schweitzer, C.; Möllmert, S.; Sock, E.; Guck, J.; Shahin, V. Matrix stiffness mechanosensing modulates the expression and distribution of transcription factors in Schwann cells. Bioeng. Transl. Med. 2021, e10257. [Google Scholar] [CrossRef]
- Friedemann, M.; Kalbitzer, L.; Franz, S.; Moeller, S.; Schnabelrauch, M.; Simon, J.C.; Pompe, T.; Franke, K. Instructing human macrophage polarization by stiffness and glycosaminoglycan functionalization in 3D collagen networks. Adv. Healthc. Mater. 2017, 6, 1600967. [Google Scholar] [CrossRef]
- Boersema, G.S.; Grotenhuis, N.; Bayon, Y.; Lange, J.F.; Bastiaansen-Jenniskens, Y.M. The effect of biomaterials used for tissue regeneration purposes on polarization of macrophages. BioResearch Open Access 2016, 5, 6–14. [Google Scholar] [CrossRef]
- Kočí, Z.; Sridharan, R.; Hibbitts, A.J.; Kneafsey, S.L.; Kearney, C.J.; O’Brien, F.J. The Use of Genipin as an Effective, Biocompatible, Anti-Inflammatory Cross-Linking Method for Nerve Guidance Conduits. Adv. Biosyst. 2020, 4, 1900212. [Google Scholar] [CrossRef]
- Dong, C.; Qiao, F.; Hou, W.; Yang, L.; Lv, Y. Graphene-based conductive fibrous scaffold boosts sciatic nerve regeneration and functional recovery upon electrical stimulation. Appl. Mater. Today 2020, 21, 100870. [Google Scholar] [CrossRef]
- Agarwal, G.; Kumar, N.; Srivastava, A. Highly elastic, electroconductive, immunomodulatory graphene crosslinked collagen cryogel for spinal cord regeneration. Mater. Sci. Eng. C 2021, 118, 111518. [Google Scholar] [CrossRef]
- Qian, Y.; Cheng, Y.; Ouyang, Y.; Yuan, W.-E.; Fan, C. Multilayered spraying and gradient dotting of nanodiamond–polycaprolactone guidance channels for restoration of immune homeostasis. NPG Asia Mater. 2019, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Maleki, M.; Zarezadeh, R.; Nouri, M.; Sadigh, A.R.; Pouremamali, F.; Asemi, Z.; Kafil, H.S.; Alemi, F.; Yousefi, B. Graphene Oxide: A Promising Material for Regenerative Medicine and Tissue Engineering. Biomol. Concepts 2020, 11, 182–200. [Google Scholar] [CrossRef]
- Bei, H.P.; Yang, Y.; Zhang, Q.; Tian, Y.; Luo, X.; Yang, M.; Zhao, X. Graphene-based nanocomposites for neural tissue engineering. Molecules 2019, 24, 658. [Google Scholar] [CrossRef] [Green Version]
- Serrano, M.C.; Feito, M.J.; González Mayorga, A.; Diez-Orejas, R.; Matesanz, M.C.; Portolés, M.T. Response of macrophages and neural cells in contact with reduced graphene oxide microfibers. Biomater. Sci. 2018, 6, 2987–2997. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, X.; Liu, Z.; Han, Q.; Tang, L.; Tian, Z.; Ren, Z.; Rong, C.; Xu, H. Miconazole alleviates peripheral nerve crush injury by mediating a macrophage phenotype change through the NF-κB pathway. Brain Behav. 2019, 9, e01400. [Google Scholar] [CrossRef]
- Huang, T.-C.; Wu, H.-L.; Chen, S.-H.; Wang, Y.-T.; Wu, C.-C. Thrombomodulin facilitates peripheral nerve regeneration through regulating M1/M2 switching. J. Neuroinflamm. 2020, 17, 240. [Google Scholar] [CrossRef] [PubMed]
- Ehmedah, A.; Nedeljkovic, P.; Dacic, S.; Repac, J.; Draskovic Pavlovic, B.; Vucevic, D.; Pekovic, S.; Bozic Nedeljkovic, B. Vitamin B complex treatment attenuates local inflammation after peripheral nerve injury. Molecules 2019, 24, 4615. [Google Scholar] [CrossRef] [Green Version]
- Mokarram, N.; Merchant, A.; Mukhatyar, V.; Patel, G.; Bellamkonda, R.V. Effect of modulating macrophage phenotype on peripheral nerve repair. Biomaterials 2012, 33, 8793–8801. [Google Scholar] [CrossRef] [Green Version]
- Mokarram, N.; Dymanus, K.; Srinivasan, A.; Lyon, J.G.; Tipton, J.; Chu, J.; English, A.W.; Bellamkonda, R.V. Immunoengineering nerve repair. Proc. Natl. Acad. Sci. USA 2017, 114, E5077–E5084. [Google Scholar] [CrossRef] [Green Version]
- Potas, J.R.; Haque, F.; Maclean, F.L.; Nisbet, D.R. Interleukin-10 conjugated electrospun polycaprolactone (PCL) nanofibre scaffolds for promoting alternatively activated (M2) macrophages around the peripheral nerve in vivo. J. Immunol. Methods 2015, 420, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yao, D.; Zhang, J.; Liu, B.; Zhang, L.; Feng, H.; Li, B. The effects of epidermal neural crest stem cells on local inflammation microenvironment in the defected sciatic nerve of rats. Front. Mol. Neurosci. 2017, 10, 133. [Google Scholar] [CrossRef] [Green Version]
- Stratton, J.A.; Shah, P.T.; Kumar, R.; Stykel, M.G.; Shapira, Y.; Grochmal, J.; Guo, G.F.; Biernaskie, J.; Midha, R. The immunomodulatory properties of adult skin-derived precursor S chwann cells: Implications for peripheral nerve injury therapy. Eur. J. Neurosci. 2016, 43, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Liu, Y.; Yang, Y.; Wang, H.; Xu, Y.; Zhang, Z. MSC-derived exosomes-based therapy for peripheral nerve injury: A novel therapeutic strategy. BioMed Res. Int. 2019, 2019, 6458237. [Google Scholar] [CrossRef]
- Qing, L.; Chen, H.; Tang, J.; Jia, X. Exosomes and their microRNA cargo: New players in peripheral nerve regeneration. Neurorehabil. Neural Repair 2018, 32, 765–776. [Google Scholar] [CrossRef] [Green Version]
- Simeoli, R.; Montague, K.; Jones, H.R.; Castaldi, L.; Chambers, D.; Kelleher, J.H.; Vacca, V.; Pitcher, T.; Grist, J.; Al-Ahdal, H. Exosomal cargo including microRNA regulates sensory neuron to macrophage communication after nerve trauma. Nat. Commun. 2017, 8, 1778. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, J.; Wang, X.; Zhang, J.; Xie, C. Extracellular vesicle-encapsulated microRNA-23a from dorsal root ganglia neurons binds to A20 and promotes inflammatory macrophage polarization following peripheral nerve injury. Aging 2021, 13, 6752. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C.; Ma, C.-B.; Yuan, H.-M.; Cao, B.-Y.; Zhu, J.-J. Macrophage-derived microvesicles promote proliferation and migration of Schwann cell on peripheral nerve repair. Biochem. Biophys. Res. Commun. 2015, 468, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Hakim, J.S.; Esmaeili Rad, M.; Grahn, P.J.; Chen, B.K.; Knight, A.M.; Schmeichel, A.M.; Isaq, N.A.; Dadsetan, M.; Yaszemski, M.J.; Windebank, A.J. Positively Charged Oligo[Poly(Ethylene Glycol) Fumarate] Scaffold Implantation Results in a Permissive Lesion Environment after Spinal Cord Injury in Rat. Tissue Eng. Part A 2015, 21, 2099–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Decker, J.T.; Margul, D.J.; Smith, D.R.; Cummings, B.J.; Anderson, A.J.; Shea, L.D. Local Immunomodulation with Anti-inflammatory Cytokine-Encoding Lentivirus Enhances Functional Recovery after Spinal Cord Injury. Mol. Ther. 2018, 26, 1756–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, C.; Sheng, P.; Xie, G.; Zhu, J.; Dong, P.; Quan, D. Fabrication of PLLA nanofibrous multi-channel conduits for neural tissue engineering. J. Control Release 2011, 152 (Suppl. 1), e234–e236. [Google Scholar] [CrossRef]
- Cornelison, R.C.; Gonzalez-Rothi, E.J.; Porvasnik, S.L.; Wellman, S.M.; Park, J.H.; Fuller, D.D.; Schmidt, C.E. Injectable hydrogels of optimized acellular nerve for injection in the injured spinal cord. Biomed. Mater. 2018, 13, 034110. [Google Scholar] [CrossRef]
- Ghuman, H.; Massensini, A.R.; Donnelly, J.; Kim, S.M.; Medberry, C.J.; Badylak, S.F.; Modo, M. ECM hydrogel for the treatment of stroke: Characterization of the host cell infiltrate. Biomaterials 2016, 91, 166–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chedly, J.; Soares, S.; Montembault, A.; von Boxberg, Y.; Veron-Ravaille, M.; Mouffle, C.; Benassy, M.N.; Taxi, J.; David, L.; Nothias, F. Physical chitosan microhydrogels as scaffolds for spinal cord injury restoration and axon regeneration. Biomaterials 2017, 138, 91–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Xie, Y.Y.; Wang, B. Role and prospects of regenerative biomaterials in the repair of spinal cord injury. Neural Regen. Res. 2019, 14, 1352–1363. [Google Scholar] [CrossRef]
- Cao, Z.; Yao, S.; Xiong, Y.; Zhang, Z.; Yang, Y.; He, F.; Zhao, H.; Guo, Y.; Wang, G.; Xie, S.; et al. Directional axonal regrowth induced by an aligned fibrin nanofiber hydrogel contributes to improved motor function recovery in canine L2 spinal cord injury. J. Mater. Sci. Mater. Med. 2020, 31, 40. [Google Scholar] [CrossRef]
- Salatino, J.W.; Ludwig, K.A.; Kozai, T.D.Y.; Purcell, E.K. Glial responses to implanted electrodes in the brain. Nat. Biomed. Eng. 2017, 1, 862–877. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, M.; Sunil, S.; Black, J.; Barkauskas, D.S.; Haung, A.Y.; Miller, R.H.; Selkirk, S.M.; Capadona, J.R. The roles of blood-derived macrophages and resident microglia in the neuroinflammatory response to implanted intracortical microelectrodes. Biomaterials 2014, 35, 8049–8064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caron, I.; Rossi, F.; Papa, S.; Aloe, R.; Sculco, M.; Mauri, E.; Sacchetti, A.; Erba, E.; Panini, N.; Parazzi, V.; et al. A new three dimensional biomimetic hydrogel to deliver factors secreted by human mesenchymal stem cells in spinal cord injury. Biomaterials 2016, 75, 135–147. [Google Scholar] [CrossRef]
- Kelly, S.H.; Shores, L.S.; Votaw, N.L.; Collier, J.H. Biomaterial strategies for generating therapeutic immune responses. Adv. Drug Deliv. Rev. 2017, 114, 3–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, H.; Yokota, K.; Fehlings, M.G. Regeneration of Spinal Cord Connectivity through Stem Cell Transplantation and Biomaterial Scaffolds. Front. Cell Neurosci. 2019, 13, 248. [Google Scholar] [CrossRef] [Green Version]
- Dumont, C.M.; Margul, D.J.; Shea, L.D. Tissue Engineering Approaches to Modulate the Inflammatory Milieu following Spinal Cord Injury. Cells Tissues Organs 2016, 202, 52–66. [Google Scholar] [CrossRef] [Green Version]
- Ham, T.R.; Leipzig, N.D. Biomaterial strategies for limiting the impact of secondary events following spinal cord injury. Biomed. Mater. 2018, 13, 024105. [Google Scholar] [CrossRef]
- Stein, A.; Panjwani, A.; Sison, C.; Rosen, L.; Chugh, R.; Metz, C.; Bank, M.; Bloom, O. Pilot study: Elevated circulating levels of the proinflammatory cytokine macrophage migration inhibitory factor in patients with chronic spinal cord injury. Arch. Phys. Med. Rehabil. 2013, 94, 1498–1507. [Google Scholar] [CrossRef]
- David, S.; Aguayo, A.J. Axonal elongation into peripheral nervous system "bridges" after central nervous system injury in adult rats. Science 1981, 214, 931–933. [Google Scholar] [CrossRef] [PubMed]
- Aguayo, A.J.; David, S.; Bray, G.M. Influences of the glial environment on the elongation of axons after injury: Transplantation studies in adult rodents. J. Exp. Biol. 1981, 95, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.M.; Issa, V.M.K.; Aguayo, A.J. Regeneration of long spinal axons in the rat. J. Neurocytol. 1984, 13, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Côté, M.-P.; Amin, A.A.; Tom, V.J.; Houle, J.D. Peripheral Nerve Grafts Support Regeneration after Spinal Cord Injury. Neurother. J. Am. Soc. Exp. Neurother. 2011, 8, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Cao, Y.; Olson, L. Spinal Cord Repair in Adult Paraplegic Rats: Partial Restoration of Hind Limb Function. Science 1996, 273, 510–513. [Google Scholar] [CrossRef]
- Strasberg, S.R.; Hertl, M.C.; Mackinnon, S.E.; Lee, C.K.; Watanabe, O.; Tarasidis, G.; Hunter, D.A.; Wong, P.Y. Peripheral Nerve Allograft Preservation Improves Regeneration and Decreases Systemic Cyclosporin A Requirements. Exp. Neurol. 1996, 139, 306–316. [Google Scholar] [CrossRef]
- Carriel, V.; Alaminos, M.; Garzón, I.; Campos, A.; Cornelissen, M. Tissue engineering of the peripheral nervous system. Expert Rev. Neurother. 2014, 14, 301–318. [Google Scholar] [CrossRef]
- Chu, T.-H.; Wang, L.; Guo, A.; Chan, V.W.-K.; Wong, C.W.-M.; Wu, W. GDNF-treated acellular nerve graft promotes motoneuron axon regeneration after implantation into cervical root avulsed spinal cord. Neuropathol. Appl. Neurobiol. 2012, 38, 681–695. [Google Scholar] [CrossRef]
- Salgado, A.J.; Sousa, N.; Silva, N.A.; Neves, N.M.; Reis, R.L. Hydrogels for spinal cord injury regeneration. In Natural-Based Polymers for Biomedical Application; Woodhead Publishing: Cambridge, UK, 2008. [Google Scholar]
- Hoban, D.B.; Newland, B.; Moloney, T.C.; Howard, L.; Pandit, A.; Dowd, E. The reduction in immunogenicity of neurotrophin overexpressing stem cells after intra-striatal transplantation by encapsulation in an in situ gelling collagen hydrogel. Biomaterials 2013, 34, 9420–9429. [Google Scholar] [CrossRef]
- Zhong, J.; Chan, A.; Morad, L.; Kornblum, H.I.; Fan, G.; Carmichael, S.T. Hydrogel matrix to support stem cell survival after brain transplantation in stroke. Neurorehabil. Neural Repair 2010, 24, 636–644. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Cao, B.; Feng, M.; Zhou, Q.; Sun, X.; Wu, S.; Jin, S.; Liu, H.; Lianhong, J. Combinated transplantation of neural stem cells and collagen type I promote functional recovery after cerebral ischemia in rats. Anat. Rec. 2010, 293, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Jimenez Hamann, M.C.; Tator, C.H.; Shoichet, M.S. Injectable intrathecal delivery system for localized administration of EGF and FGF-2 to the injured rat spinal cord. Exp. Neurol 2005, 194, 106–119. [Google Scholar] [CrossRef]
- Yoshii, S.; Oka, M.; Shima, M.; Taniguchi, A.; Taki, Y.; Akagi, M. Restoration of function after spinal cord transection using a collagen bridge. J. Biomed. Mater. Res. A 2004, 70, 569–575. [Google Scholar] [CrossRef]
- Yao, L.; Daly, W.; Newland, B.; Yao, S.; Wang, W.; Chen, B.K.; Madigan, N.; Windebank, A.; Pandit, A. Improved axonal regeneration of transected spinal cord mediated by multichannel collagen conduits functionalized with neurotrophin-3 gene. Gene Ther. 2013, 20, 1149–1157. [Google Scholar] [CrossRef]
- Shi, Q.; Gao, W.; Han, X.; Zhu, X.; Sun, J.; Xie, F.; Hou, X.; Yang, H.; Dai, J.; Chen, L. Collagen scaffolds modified with collagen-binding bFGF promotes the neural regeneration in a rat hemisected spinal cord injury model. Sci. China Life Sci. 2014, 57, 232–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Z.; Gao, W.; Yue, B.; Jiang, J.; Gu, Y.; Dai, J.; Chen, L.; Shi, Q. Promotion of neurological recovery in rat spinal cord injury by mesenchymal stem cells loaded on nerve-guided collagen scaffold through increasing alternatively activated macrophage polarization. J. Tissue Eng. Regen. Med. 2018, 12, e1725–e1736. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.Y.; Seo, Y.; Davaa, G.; Kim, H.W.; Kim, S.H.; Hyun, J.K. Decellularized brain matrix enhances macrophage polarization and functional improvements in rat spinal cord injury. Acta Biomater. 2020, 101, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Kumosa, L.S.; Zetterberg, V.; Schouenborg, J. Gelatin promotes rapid restoration of the blood brain barrier after acute brain injury. Acta Biomater. 2018, 65, 137–149. [Google Scholar] [CrossRef]
- Zeng, X.; Zeng, Y.S.; Ma, Y.H.; Lu, L.Y.; Du, B.L.; Zhang, W.; Li, Y.; Chan, W.Y. Bone marrow mesenchymal stem cells in a three-dimensional gelatin sponge scaffold attenuate inflammation, promote angiogenesis, and reduce cavity formation in experimental spinal cord injury. Cell Transplant. 2011, 20, 1881–1899. [Google Scholar] [CrossRef]
- Ma, D.; Zhao, Y.; Huang, L.; Xiao, Z.; Chen, B.; Shi, Y.; Shen, H.; Dai, J. A novel hydrogel-based treatment for complete transection spinal cord injury repair is driven by microglia/macrophages repopulation. Biomaterials 2020, 237, 119830. [Google Scholar] [CrossRef]
- Fawcett, J.W.; Oohashi, T.; Pizzorusso, T. The roles of perineuronal nets and the perinodal extracellular matrix in neuronal function. Nat. Rev. Neurosci. 2019, 20, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Wrobel, M.R.; Sundararaghavan, H.G. Biomaterial Cues to Direct a Pro-regenerative Phenotype in Macrophages and Schwann Cells. Neuroscience 2018, 376, 172–187. [Google Scholar] [CrossRef]
- Stern, R.; Kogan, G.; Jedrzejas, M.J.; Soltes, L. The many ways to cleave hyaluronan. Biotechnol Adv. 2007, 25, 537–557. [Google Scholar] [CrossRef]
- Rayahin, J.E.; Buhrman, J.S.; Zhang, Y.; Koh, T.J.; Gemeinhart, R.A. High and low molecular weight hyaluronic acid differentially influence macrophage activation. ACS Biomater. Sci. Eng. 2015, 1, 481–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Cha, J.; Jang, M.; Kim, P. Hyaluronic acid-based extracellular matrix triggers spontaneous M2-like polarity of monocyte/macrophage. Biomater. Sci. 2019, 7, 2264–2271. [Google Scholar] [CrossRef]
- Khaing, Z.Z.; Milman, B.D.; Vanscoy, J.E.; Seidlits, S.K.; Grill, R.J.; Schmidt, C.E. High molecular weight hyaluronic acid limits astrocyte activation and scar formation after spinal cord injury. J. Neural Eng. 2011, 8, 046033. [Google Scholar] [CrossRef]
- Thompson, R.E.; Pardieck, J.; Smith, L.; Kenny, P.; Crawford, L.; Shoichet, M.; Sakiyama-Elbert, S. Effect of hyaluronic acid hydrogels containing astrocyte-derived extracellular matrix and/or V2a interneurons on histologic outcomes following spinal cord injury. Biomaterials 2018, 162, 208–223. [Google Scholar] [CrossRef]
- Ruppert, S.M.; Hawn, T.R.; Arrigoni, A.; Wight, T.N.; Bollyky, P.L. Tissue integrity signals communicated by high-molecular weight hyaluronan and the resolution of inflammation. Immunol. Res. 2014, 58, 186–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, J.W.; Gilchrist, C.; Fehlings, M.G. High molecular weight hyaluronan reduces lipopolysaccharide mediated microglial activation. J. Neurochem. 2012, 122, 344–355. [Google Scholar] [CrossRef]
- Yasuda, T. Hyaluronan inhibits cytokine production by lipopolysaccharide-stimulated U937 macrophages through down-regulation of NF-kappaB via ICAM-1. Inflamm. Res. 2007, 56, 246–253. [Google Scholar] [CrossRef]
- Li, X.; Zhang, C.; Haggerty, A.E.; Yan, J.; Lan, M.; Seu, M.; Yang, M.; Marlow, M.M.; Maldonado-Lasuncion, I.; Cho, B.; et al. The effect of a nanofiber-hydrogel composite on neural tissue repair and regeneration in the contused spinal cord. Biomaterials 2020, 245, 119978. [Google Scholar] [CrossRef] [PubMed]
- Barros, C.S.; Franco, S.J.; Muller, U. Extracellular matrix: Functions in the nervous system. Cold Spring Harb. Perspect Biol. 2011, 3, a005108. [Google Scholar] [CrossRef] [Green Version]
- Burnside, E.R.; Bradbury, E.J. Manipulating the extracellular matrix and its role in brain and spinal cord plasticity and repair. Neuropathol. Appl. Neurobiol. 2014, 40, 26–59. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Nirwane, A.; Yao, Y. Basement membrane and blood-brain barrier. Stroke Vasc. Neurol. 2019, 4, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, L.W.; Cua, R.; Keough, M.B.; Haylock-Jacobs, S.; Yong, V.W. Pathophysiology of the brain extracellular matrix: A new target for remyelination. Nat. Rev. Neurosci. 2013, 14, 722–729. [Google Scholar] [CrossRef]
- Sikkema, A.H.; Stoffels, J.M.J.; Wang, P.; Basedow, F.J.; Bulsink, R.; Bajramovic, J.J.; Baron, W. Fibronectin aggregates promote features of a classically and alternatively activated phenotype in macrophages. J. Neuroinflammation 2018, 15, 218. [Google Scholar] [CrossRef]
- King, V.R.; Phillips, J.B.; Hunt-Grubbe, H.; Brown, R.; Priestley, J.V. Characterization of non-neuronal elements within fibronectin mats implanted into the damaged adult rat spinal cord. Biomaterials 2006, 27, 485–496. [Google Scholar] [CrossRef]
- Hayashi, H.; Yamada, M.; Kumai, J.; Takagi, N.; Nomizu, M. Biological activities of laminin-111-derived peptide-chitosan matrices in a primary culture of rat cortical neurons. Arch. Biochem. Biophys. 2018, 648, 53–59. [Google Scholar] [CrossRef]
- Lei, W.L.; Xing, S.G.; Deng, C.Y.; Ju, X.C.; Jiang, X.Y.; Luo, Z.G. Laminin/beta1 integrin signal triggers axon formation by promoting microtubule assembly and stabilization. Cell Res. 2012, 22, 954–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, S.; Xu, Q.; Tian, W.; Cui, F.; Cai, Q.; Ma, J.; Lee, I.S. The repair of brain lesion by implantation of hyaluronic acid hydrogels modified with laminin. J. Neurosci. Methods 2005, 148, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Tysseling, V.M.; Sahni, V.; Pashuck, E.T.; Birch, D.; Hebert, A.; Czeisler, C.; Stupp, S.I.; Kessler, J.A. Self-assembling peptide amphiphile promotes plasticity of serotonergic fibers following spinal cord injury. J. Neurosci. Res. 2010, 88, 3161–3170. [Google Scholar] [CrossRef] [Green Version]
- Barroso, M.M.; Freire, E.; Limaverde, G.S.; Rocha, G.M.; Batista, E.J.; Weissmuller, G.; Andrade, L.R.; Coelho-Sampaio, T. Artificial laminin polymers assembled in acidic pH mimic basement membrane organization. J. Biol. Chem. 2008, 283, 11714–11720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menezes, K.; de Menezes, J.R.; Nascimento, M.A.; Santos Rde, S.; Coelho-Sampaio, T. Polylaminin, a polymeric form of laminin, promotes regeneration after spinal cord injury. FASEB J. 2010, 24, 4513–4522. [Google Scholar] [CrossRef]
- Gunther, M.I.; Weidner, N.; Muller, R.; Blesch, A. Cell-seeded alginate hydrogel scaffolds promote directed linear axonal regeneration in the injured rat spinal cord. Acta Biomater. 2015, 27, 140–150. [Google Scholar] [CrossRef]
- Stokols, S.; Tuszynski, M.H. Freeze-dried agarose scaffolds with uniaxial channels stimulate and guide linear axonal growth following spinal cord injury. Biomaterials 2006, 27, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Sung, M.J.; Seo, S.B.; Yoo, S.J.; Lim, W.K.; Kim, H.M. Water-soluble chitosan inhibits the production of pro-inflammatory cytokine in human astrocytoma cells activated by amyloid beta peptide and interleukin-1beta. Neurosci. Lett. 2002, 321, 105–109. [Google Scholar] [CrossRef]
- Khodagholi, F.; Eftekharzadeh, B.; Maghsoudi, N.; Rezaei, P.F. Chitosan prevents oxidative stress-induced amyloid beta formation and cytotoxicity in NT2 neurons: Involvement of transcription factors Nrf2 and NF-kappaB. Mol. Cell Biochem. 2010, 337, 39–51. [Google Scholar] [CrossRef]
- Hong, L.T.A.; Kim, Y.M.; Park, H.H.; Hwang, D.H.; Cui, Y.; Lee, E.M.; Yahn, S.; Lee, J.K.; Song, S.C.; Kim, B.G. An injectable hydrogel enhances tissue repair after spinal cord injury by promoting extracellular matrix remodeling. Nat. Commun. 2017, 8, 533. [Google Scholar] [CrossRef]
- Shin, J.E.; Jung, K.; Kim, M.; Hwang, K.; Lee, H.; Kim, I.S.; Lee, B.H.; Lee, I.S.; Park, K.I. Brain and spinal cord injury repair by implantation of human neural progenitor cells seeded onto polymer scaffolds. Exp. Mol. Med. 2018, 50, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guest, J.D.; Moore, S.W.; Aimetti, A.A.; Kutikov, A.B.; Santamaria, A.J.; Hofstetter, C.P.; Ropper, A.E.; Theodore, N.; Ulich, T.R.; Layer, R.T. Internal decompression of the acutely contused spinal cord: Differential effects of irrigation only versus biodegradable scaffold implantation. Biomaterials 2018, 185, 284–300. [Google Scholar] [CrossRef]
- Pertici, V.; Amendola, J.; Laurin, J.; Gigmes, D.; Madaschi, L.; Carelli, S.; Marqueste, T.; Gorio, A.; Decherchi, P. The use of poly(N-[2-hydroxypropyl]-methacrylamide) hydrogel to repair a T10 spinal cord hemisection in rat: A behavioural, electrophysiological and anatomical examination. ASN Neuro 2013, 5, 149–166. [Google Scholar] [CrossRef]
- Hejcl, A.; Sedy, J.; Kapcalova, M.; Toro, D.A.; Amemori, T.; Lesny, P.; Likavcanova-Masinova, K.; Krumbholcova, E.; Pradny, M.; Michalek, J.; et al. HPMA-RGD hydrogels seeded with mesenchymal stem cells improve functional outcome in chronic spinal cord injury. Stem Cells Dev. 2010, 19, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Kubinova, S.; Horak, D.; Hejcl, A.; Plichta, Z.; Kotek, J.; Proks, V.; Forostyak, S.; Sykova, E. SIKVAV-modified highly superporous PHEMA scaffolds with oriented pores for spinal cord injury repair. J. Tissue Eng. Regen. Med. 2015, 9, 1298–1309. [Google Scholar] [CrossRef]
- Bjugstad, K.B.; Redmond, D.E.; Lampe, K.J.; Kern, D.S.; Sladek, J.R., Jr.; Mahoney, M.J. Biocompatibility of PEG-based hydrogels in primate brain. Cell Transplant. 2008, 17, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Woerly, S.; Fort, S.; Pignot-Paintrand, I.; Cottet, C.; Carcenac, C.; Savasta, M. Development of a sialic acid-containing hydrogel of poly[N-(2-hydroxypropyl) methacrylamide]: Characterization and implantation study. Biomacromolecules 2008, 9, 2329–2337. [Google Scholar] [CrossRef] [PubMed]
- Voges, J.; Lehrke, R.; Kim, D.G.; Lucas, C.; Schroder, R.; Sturm, V.; Stricker, H. Tissue reactions after long-term intracerebral implantation of three different types of biodegradable polylactide rods in the rat. J. Exp. Ther. Oncol. 2002, 2, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Ziemba, K.S.; Smith, G.M.; Jin, Y. Evaluation of cellular organization and axonal regeneration through linear PLA foam implants in acute and chronic spinal cord injury. J. Biomed. Mater. Res. A 2007, 83, 512–520. [Google Scholar] [CrossRef]
- Slotkin, J.R.; Pritchard, C.D.; Luque, B.; Ye, J.; Layer, R.T.; Lawrence, M.S.; O’Shea, T.M.; Roy, R.R.; Zhong, H.; Vollenweider, I.; et al. Biodegradable scaffolds promote tissue remodeling and functional improvement in non-human primates with acute spinal cord injury. Biomaterials 2017, 123, 63–76. [Google Scholar] [CrossRef]
- Sun, X.; Bai, Y.; Zhai, H.; Liu, S.; Zhang, C.; Xu, Y.; Zou, J.; Wang, T.; Chen, S.; Zhu, Q.; et al. Devising micro/nano-architectures in multi-channel nerve conduits towards a pro-regenerative matrix for the repair of spinal cord injury. Acta Biomater. 2019, 86, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, L.M.; Doulames, V.M.; Wang, A.T.; Dubbin, K.; Suhar, R.A.; Kratochvil, M.J.; Medress, Z.A.; Plant, G.W.; Heilshorn, S.C. Designer, injectable gels to prevent transplanted Schwann cell loss during spinal cord injury therapy. Sci. Adv. 2020, 6, eaaz1039. [Google Scholar] [CrossRef] [Green Version]
- Trimaille, T.; Pertici, V.; Gigmes, D. Recent advances in synthetic polymer based hydrogels for spinal cord repair Hydrogels based on synthetic polymers for bone marrow repair. Comptes Rendus Chim. 2016, 19, 157–166. [Google Scholar] [CrossRef]
- Zielinska, A.; Carreiro, F.; Oliveira, A.M.; Neves, A.; Pires, B.; Venkatesh, D.N.; Durazzo, A.; Lucarini, M.; Eder, P.; Silva, A.M.; et al. Polymeric Nanoparticles: Production, Characterization, Toxicology and Ecotoxicology. Molecules 2020, 25, 3731. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Borgens, R.B. Polymer and nano-technology applications for repair and reconstruction of the central nervous system. Exp. Neurol. 2012, 233, 126–144. [Google Scholar] [CrossRef]
- Tukmachev, D.; Forostyak, S.; Koci, Z.; Zaviskova, K.; Vackova, I.; Vyborny, K.; Sandvig, I.; Sandvig, A.; Medberry, C.J.; Badylak, S.F.; et al. Injectable Extracellular Matrix Hydrogels as Scaffolds for Spinal Cord Injury Repair. Tissue Eng. Part A 2016, 22, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Straley, K.S.; Foo, C.W.; Heilshorn, S.C. Biomaterial design strategies for the treatment of spinal cord injuries. J. Neurotrauma 2010, 27, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Fuhrmann, T.; Zhou, Y.; Dalton, P.D. Host reaction to poly(2-hydroxyethyl methacrylate) scaffolds in a small spinal cord injury model. J. Mater. Sci. Mater. Med. 2013, 24, 2001–2011. [Google Scholar] [CrossRef]
- Pertici, V.; Trimaille, T.; Laurin, J.; Felix, M.S.; Marqueste, T.; Pettmann, B.; Chauvin, J.P.; Gigmes, D.; Decherchi, P. Repair of the injured spinal cord by implantation of a synthetic degradable block copolymer in rat. Biomaterials 2014, 35, 6248–6258. [Google Scholar] [CrossRef]
- Rauck, B.M.; Novosat, T.L.; Oudega, M.; Wang, Y. Biocompatibility of a coacervate-based controlled release system for protein delivery to the injured spinal cord. Acta Biomater. 2015, 11, 204–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, C.R.; Serra, T.; Oliveira, M.I.; Planell, J.A.; Barbosa, M.A.; Navarro, M. Impact of 3-D printed PLA- and chitosan-based scaffolds on human monocyte/macrophage responses: Unraveling the effect of 3-D structures on inflammation. Acta Biomater. 2014, 10, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Saino, E.; Focarete, M.L.; Gualandi, C.; Emanuele, E.; Cornaglia, A.I.; Imbriani, M.; Visai, L. Effect of electrospun fiber diameter and alignment on macrophage activation and secretion of proinflammatory cytokines and chemokines. Biomacromolecules 2011, 12, 1900–1911. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; De Laporte, L.; Zelivyanskaya, M.L.; Whittlesey, K.J.; Anderson, A.J.; Cummings, B.J.; Shea, L.D. Multiple channel bridges for spinal cord injury: Cellular characterization of host response. Tissue Eng. Part A 2009, 15, 3283–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuidema, J.M.; Gilbert, R.J.; Gottipati, M.K. Biomaterial Approaches to Modulate Reactive Astroglial Response. Cells Tissues Organs 2018, 205, 372–395. [Google Scholar] [CrossRef]
- Wang, Z.; Nong, J.; Shultz, R.B.; Zhang, Z.; Kim, T.; Tom, V.J.; Ponnappan, R.K.; Zhong, Y. Local delivery of minocycline from metal ion-assisted self-assembled complexes promotes neuroprotection and functional recovery after spinal cord injury. Biomaterials 2017, 112, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Papa, S.; Rossi, F.; Ferrari, R.; Mariani, A.; De Paola, M.; Caron, I.; Fiordaliso, F.; Bisighini, C.; Sammali, E.; Colombo, C.; et al. Selective nanovector mediated treatment of activated proinflammatory microglia/macrophages in spinal cord injury. ACS Nano 2013, 7, 9881–9895. [Google Scholar] [CrossRef]
- Louw, A.M.; Kolar, M.K.; Novikova, L.N.; Kingham, P.J.; Wiberg, M.; Kjems, J.; Novikov, L.N. Chitosan polyplex mediated delivery of miRNA-124 reduces activation of microglial cells in vitro and in rat models of spinal cord injury. Nanomedicine 2016, 12, 643–653. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Young, W. Managing inflammation after spinal cord injury through manipulation of macrophage function. Neural Plast 2013, 2013, 945034. [Google Scholar] [CrossRef] [Green Version]
- Sellers, D.L.; Kim, T.H.; Mount, C.W.; Pun, S.H.; Horner, P.J. Poly(lactic-co-glycolic) acid microspheres encapsulated in Pluronic F-127 prolong hirudin delivery and improve functional recovery from a demyelination lesion. Biomaterials 2014, 35, 8895–8902. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.M.; Kubilius, M.B.; Holland, S.J.; Seidlits, S.K.; Boehler, R.M.; Anderson, A.J.; Cummings, B.J.; Shea, L.D. Channel density and porosity of degradable bridging scaffolds on axon growth after spinal injury. Biomaterials 2013, 34, 2213–2220. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.Y.; Leveque, J.C.; Brumblay, H.; Krebsbach, P.H.; Hollister, S.J.; Lamarca, F. Macro-architectures in spinal cord scaffold implants influence regeneration. J. Neurotrauma 2008, 25, 1027–1037. [Google Scholar] [CrossRef] [Green Version]
- Sussman, E.M.; Halpin, M.C.; Muster, J.; Moon, R.T.; Ratner, B.D. Porous implants modulate healing and induce shifts in local macrophage polarization in the foreign body reaction. Ann. Biomed. Eng. 2014, 42, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Moshayedi, P.; Ng, G.; Kwok, J.C.; Yeo, G.S.; Bryant, C.E.; Fawcett, J.W.; Franze, K.; Guck, J. The relationship between glial cell mechanosensitivity and foreign body reactions in the central nervous system. Biomaterials 2014, 35, 3919–3925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakshi, A.; Fisher, O.; Dagci, T.; Himes, B.T.; Fischer, I.; Lowman, A. Mechanically engineered hydrogel scaffolds for axonal growth and angiogenesis after transplantation in spinal cord injury. J. Neurosurg. Spine 2004, 1, 322–329. [Google Scholar] [CrossRef]
- Liu, S.; Schackel, T.; Weidner, N.; Puttagunta, R. Biomaterial-Supported Cell Transplantation Treatments for Spinal Cord Injury: Challenges and Perspectives. Front. Cell Neurosci. 2017, 11, 430. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Dolado, E.; Gonzalez-Mayorga, A.; Gutierrez, M.C.; Serrano, M.C. Immunomodulatory and angiogenic responses induced by graphene oxide scaffolds in chronic spinal hemisected rats. Biomaterials 2016, 99, 72–81. [Google Scholar] [CrossRef]
- Lopez-Dolado, E.; Gonzalez-Mayorga, A.; Portoles, M.T.; Feito, M.J.; Ferrer, M.L.; Del Monte, F.; Gutierrez, M.C.; Serrano, M.C. Subacute Tissue Response to 3D Graphene Oxide Scaffolds Implanted in the Injured Rat Spinal Cord. Adv. Healthc. Mater. 2015, 4, 1861–1868. [Google Scholar] [CrossRef]
- Margul, D.J.; Park, J.; Boehler, R.M.; Smith, D.R.; Johnson, M.A.; McCreedy, D.A.; He, T.; Ataliwala, A.; Kukushliev, T.V.; Liang, J.; et al. Reducing neuroinflammation by delivery of IL-10 encoding lentivirus from multiple-channel bridges. Bioeng. Transl. Med. 2016, 1, 136–148. [Google Scholar] [CrossRef]
- Smith, D.R.; Dumont, C.M.; Park, J.; Ciciriello, A.J.; Guo, A.; Tatineni, R.; Cummings, B.J.; Anderson, A.J.; Shea, L.D. Polycistronic Delivery of IL-10 and NT-3 Promotes Oligodendrocyte Myelination and Functional Recovery in a Mouse Spinal Cord Injury Model. Tissue Eng. Part A 2020, 26, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.; Joseph, G.; Patel, A.; Patel, S.; Bustin, D.; Mawson, D.; Tuesta, L.M.; Puentes, R.; Ghosh, M.; Pearse, D.D. Suspension matrices for improved Schwann-cell survival after implantation into the injured rat spinal cord. J. Neurotrauma 2010, 27, 789–801. [Google Scholar] [CrossRef]
- Medberry, C.J.; Crapo, P.M.; Siu, B.F.; Carruthers, C.A.; Wolf, M.T.; Nagarkar, S.P.; Agrawal, V.; Jones, K.E.; Kelly, J.; Johnson, S.A.; et al. Hydrogels derived from central nervous system extracellular matrix. Biomaterials 2013, 34, 1033–1040. [Google Scholar] [CrossRef] [Green Version]
- Wolf, M.T.; Dearth, C.L.; Ranallo, C.A.; LoPresti, S.T.; Carey, L.E.; Daly, K.A.; Brown, B.N.; Badylak, S.F. Macrophage polarization in response to ECM coated polypropylene mesh. Biomaterials 2014, 35, 6838–6849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziki, J.L.; Wang, D.S.; Pineda, C.; Sicari, B.M.; Rausch, T.; Badylak, S.F. Solubilized extracellular matrix bioscaffolds derived from diverse source tissues differentially influence macrophage phenotype. J. Biomed. Mater. Res. A 2017, 105, 138–147. [Google Scholar] [CrossRef]
- Brown, B.N.; Valentin, J.E.; Stewart-Akers, A.M.; McCabe, G.P.; Badylak, S.F. Macrophage phenotype and remodeling outcomes in response to biologic scaffolds with and without a cellular component. Biomaterials 2009, 30, 1482–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Trinh, T.H.T.; Yoo, M.; Shin, J.; Lee, H.; Kim, J.; Hwang, E.; Lim, Y.B.; Ryou, C. Self-Assembling Peptides and Their Application in the Treatment of Diseases. Int. J. Mol. Sci. 2019, 20, 5850. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Pi, B.; Wang, H.; Wang, X.-M. Self-assembling peptide nanofiber hydrogels for central nervous system regeneration. Front. Mater. Sci. 2014, 9, 1–13. [Google Scholar] [CrossRef]
- Cigognini, D.; Satta, A.; Colleoni, B.; Silva, D.; Donega, M.; Antonini, S.; Gelain, F. Evaluation of early and late effects into the acute spinal cord injury of an injectable functionalized self-assembling scaffold. PLoS ONE 2011, 6, e19782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cigognini, D.; Silva, D.; Paloppi, S.; Gelain, F. Evaluation of mechanical properties and therapeutic effect of injectable self-assembling hydrogels for spinal cord injury. J. Biomed. Nanotechnol. 2014, 10, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.A. Nanotechnology applications and approaches for neuroregeneration and drug delivery to the central nervous system. Ann. N. Y. Acad. Sci. 2010, 1199, 221–230. [Google Scholar] [CrossRef]
- Saeedi, M.; Eslamifar, M.; Khezri, K.; Dizaj, S.M. Applications of nanotechnology in drug delivery to the central nervous system. Biomed. Pharm. 2019, 111, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Han, M.; Zhou, J.; Zheng, Z.F.; Lu, P.; Wang, J.J.; Wang, J.Q.; Mao, Q.J.; Gao, J.Q.; Ouyang, H.W. Repair of spinal cord injury by inhibition of astrocyte growth and inflammatory factor synthesis through local delivery of flavopiridol in PLGA nanoparticles. Biomaterials 2014, 35, 6585–6594. [Google Scholar] [CrossRef]
- Park, J.; Zhang, Y.; Saito, E.; Gurczynski, S.J.; Moore, B.B.; Cummings, B.J.; Anderson, A.J.; Shea, L.D. Intravascular innate immune cells reprogrammed via intravenous nanoparticles to promote functional recovery after spinal cord injury. Proc. Natl. Acad. Sci. USA 2019, 116, 14947–14954. [Google Scholar] [CrossRef] [Green Version]
- Getts, D.R.; Terry, R.L.; Getts, M.T.; Deffrasnes, C.; Muller, M.; van Vreden, C.; Ashhurst, T.M.; Chami, B.; McCarthy, D.; Wu, H.; et al. Therapeutic inflammatory monocyte modulation using immune-modifying microparticles. Sci. Transl. Med. 2014, 6, 219ra217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, T.; Loomis, K.H.; Pai, S.B.; Karumbaiah, L.; Gaupp, E.; Patil, K.; Patkar, R.; Bellamkonda, R.V. Nanocarrier-mediated inhibition of macrophage migration inhibitory factor attenuates secondary injury after spinal cord injury. ACS Nano 2015, 9, 1492–1505. [Google Scholar] [CrossRef] [PubMed]
- Bin, S.; Zhou, N.; Pan, J.; Pan, F.; Wu, X.F.; Zhou, Z.H. Nano-carrier mediated co-delivery of methyl prednisolone and minocycline for improved post-traumatic spinal cord injury conditions in rats. Drug Dev. Ind. Pharm. 2017, 43, 1033–1041. [Google Scholar] [CrossRef]
- Boehler, R.M.; Kuo, R.; Shin, S.; Goodman, A.G.; Pilecki, M.A.; Gower, R.M.; Leonard, J.N.; Shea, L.D. Lentivirus delivery of IL-10 to promote and sustain macrophage polarization towards an anti-inflammatory phenotype. Biotechnol. Bioeng. 2014, 111, 1210–1221. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.Y.; Fu, E.J.; Patel, P.R.; Hostetler, A.J.; Sawan, H.A.; Moss, K.A.; Hocevar, S.E.; Anderson, A.J.; Chestek, C.A.; Shea, L.D. Lentiviral Interleukin-10 Gene Therapy Preserves Fine Motor Circuitry and Function After a Cervical Spinal Cord Injury in Male and Female Mice. Neurother. J. Am. Soc. Exp. Neurother. 2020, 18, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Nandoe Tewarie, R.S.; Hurtado, A.; Bartels, R.H.; Grotenhuis, A.; Oudega, M. Stem cell-based therapies for spinal cord injury. J. Spinal Cord Med. 2009, 32, 105–114. [Google Scholar] [CrossRef]
- Nagoshi, N.; Tsuji, O.; Nakamura, M.; Okano, H. Cell therapy for spinal cord injury using induced pluripotent stem cells. Regen. Ther. 2019, 11, 75–80. [Google Scholar] [CrossRef]
- Joyce, N.; Annett, G.; Wirthlin, L.; Olson, S.; Bauer, G.; Nolta, J.A. Mesenchymal stem cells for the treatment of neurodegenerative disease. Regen. Med. 2010, 5, 933–946. [Google Scholar] [CrossRef] [Green Version]
- Ohtaki, H.; Ylostalo, J.H.; Foraker, J.E.; Robinson, A.P.; Reger, R.L.; Shioda, S.; Prockop, D.J. Stem/progenitor cells from bone marrow decrease neuronal death in global ischemia by modulation of inflammatory/immune responses. Proc. Natl. Acad. Sci. USA 2008, 105, 14638–14643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Hematti, P. Mesenchymal stem cell-educated macrophages: A novel type of alternatively activated macrophages. Exp. Hematol. 2009, 37, 1445–1453. [Google Scholar] [CrossRef] [Green Version]
- Sykova, E.; Jendelova, P.; Urdzikova, L.; Lesny, P.; Hejcl, A. Bone marrow stem cells and polymer hydrogels--two strategies for spinal cord injury repair. Cell Mol. Neurobiol. 2006, 26, 1113–1129. [Google Scholar] [CrossRef]
- Gao, M.; Lu, P.; Bednark, B.; Lynam, D.; Conner, J.M.; Sakamoto, J.; Tuszynski, M.H. Templated agarose scaffolds for the support of motor axon regeneration into sites of complete spinal cord transection. Biomaterials 2013, 34, 1529–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, H.; Uchida, K.; Guerrero, A.R.; Watanabe, S.; Sugita, D.; Takeura, N.; Yoshida, A.; Long, G.; Wright, K.T.; Johnson, W.E.; et al. Transplantation of mesenchymal stem cells promotes an alternative pathway of macrophage activation and functional recovery after spinal cord injury. J. Neurotrauma 2012, 29, 1614–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritfeld, G.J.; Rauck, B.M.; Novosat, T.L.; Park, D.; Patel, P.; Roos, R.A.; Wang, Y.; Oudega, M. The effect of a polyurethane-based reverse thermal gel on bone marrow stromal cell transplant survival and spinal cord repair. Biomaterials 2014, 35, 1924–1931. [Google Scholar] [CrossRef] [Green Version]
- Ropper, A.E.; Thakor, D.K.; Han, I.; Yu, D.; Zeng, X.; Anderson, J.E.; Aljuboori, Z.; Kim, S.W.; Wang, H.; Sidman, R.L.; et al. Defining recovery neurobiology of injured spinal cord by synthetic matrix-assisted hMSC implantation. Proc. Natl. Acad. Sci. USA 2017, 114, E820–E829. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Ham, T.R.; Neill, N.; Farrag, M.; Mohrman, A.E.; Koenig, A.M.; Leipzig, N.D. A Hydrogel Bridge Incorporating Immobilized Growth Factors and Neural Stem/Progenitor Cells to Treat Spinal Cord Injury. Adv. Healthc. Mater. 2016, 5, 802–812. [Google Scholar] [CrossRef]
- Watanabe, S.; Uchida, K.; Nakajima, H.; Matsuo, H.; Sugita, D.; Yoshida, A.; Honjoh, K.; Johnson, W.E.; Baba, H. Early transplantation of mesenchymal stem cells after spinal cord injury relieves pain hypersensitivity through suppression of pain-related signaling cascades and reduced inflammatory cell recruitment. Stem Cells 2015, 33, 1902–1914. [Google Scholar] [CrossRef] [Green Version]
- Busch, S.A.; Horn, K.P.; Cuascut, F.X.; Hawthorne, A.L.; Bai, L.; Miller, R.H.; Silver, J. Adult NG2+ cells are permissive to neurite outgrowth and stabilize sensory axons during macrophage-induced axonal dieback after spinal cord injury. J. Neurosci. 2010, 30, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Cusimano, M.; Biziato, D.; Brambilla, E.; Donega, M.; Alfaro-Cervello, C.; Snider, S.; Salani, G.; Pucci, F.; Comi, G.; Garcia-Verdugo, J.M.; et al. Transplanted neural stem/precursor cells instruct phagocytes and reduce secondary tissue damage in the injured spinal cord. Brain 2012, 135, 447–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riemann, L.; Younsi, A.; Scherer, M.; Zheng, G.; Skutella, T.; Unterberg, A.W.; Zweckberger, K. Transplantation of Neural Precursor Cells Attenuates Chronic Immune Environment in Cervical Spinal Cord Injury. Front. Neurol. 2018, 9, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, S.; Yasuda, A.; Iwai, H.; Takano, M.; Kobayashi, Y.; Nori, S.; Tsuji, O.; Fujiyoshi, K.; Ebise, H.; Toyama, Y.; et al. Time-dependent changes in the microenvironment of injured spinal cord affects the therapeutic potential of neural stem cell transplantation for spinal cord injury. Mol. Brain 2013, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the glial scar for spinal cord repair. Nat. Commun. 2019, 10, 3879. [Google Scholar] [CrossRef]
- Zárate, S.; Stevnsner, T.; Gredilla, R. Role of Estrogen and Other Sex Hormones in Brain Aging. Neuroprotection and DNA Repair. Front. Aging Neurosci. 2017, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Xie, H.; Wu, J.; Liu, D.; Yao, S. Villainous role of estrogen in macrophage-nerve interaction in endometriosis. Reprod. Biol. Endocrinol. 2018, 16, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schell, J.B.; Crane, C.A.; Smith, M.F., Jr.; Roberts, M.R. Differential ex vivo nitric oxide production by acutely isolated neonatal and adult microglia. J. Neuroimmunol. 2007, 189, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Milligan, C.E.; Levitt, P.; Cunningham, T.J. Brain macrophages and microglia respond differently to lesions of the developing and adult visual system. J. Comp. Neurol. 1991, 314, 136–146. [Google Scholar] [CrossRef]
- Chhor, V.; Moretti, R.; Le Charpentier, T.; Sigaut, S.; Lebon, S.; Schwendimann, L.; Oré, M.V.; Zuiani, C.; Milan, V.; Josserand, J.; et al. Role of microglia in a mouse model of paediatric traumatic brain injury. Brain Behav. Immun. 2017, 63, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Umekawa, T.; Osman, A.M.; Han, W.; Ikeda, T.; Blomgren, K. Resident microglia, rather than blood-derived macrophages, contribute to the earlier and more pronounced inflammatory reaction in the immature compared with the adult hippocampus after hypoxia-ischemia. Glia 2015, 63, 2220–2230. [Google Scholar] [CrossRef] [Green Version]




| TECHNIQUE/PRODUCT | |||
|---|---|---|---|
| Physical Approach | In vitro/In vivo | Immune Response | Ref |
| Micropatterned substrates containing fibronectin and Pluronics F127 | |||
| Micropatterns | BMDMs C57Bl/6J mice | Elongated macrophages on micropatterns express M2 phenotype biomarkers | [176] |
| (P(LLA-CL)) nerve guidance conduit | |||
| Aligned nanofibers | * Sciatic nerve from SD rats | Higher expression of arg1, IL-10 and higher M2/M1 ratio compared to random nanofibers | [177] |
| Microstructured PFPE surfaces | |||
| Micropatterns | PBMCs human macrophages | More M2 macrophages expressed in post patterns than line patterns and greater distance between posts induce a stronger anti-inflammatory effect | [178] |
| PLCL nerve guidance conduit | |||
| Micropatterns and GO nanosheets | * Sciatic nerve in SD rats | Micropatterned and GO-modified conduit promotes M2 phenotype differentiation | [173] |
| PCL macroporous nanofiber wrap | |||
| Pores and fibers size | Sciatic nerve SD rats | Macroporous nanofiber wrap increases IL-10 expression and decreases TNF-α expression and M1/M2 ratio on the injury site | [172] |
| TECHNIQUE/PRODUCT | |||
|---|---|---|---|
| Chemical Approach | In vitro/In vivo | Immune Response | Ref |
| Collagen-based nerve guidance conduit | |||
| Genipin crosslinking | THP-1 cells | Crosslinking of outer conduit reduced M1 polarization | [188] |
| Graphene—based conductive fibrous scaffold | |||
| Graphene modification and electrical stimulation | * Sciatic nerve 4-week-old SD rats | Induced change in macrophages phenotype from M1 to M2 | [189] |
| Graphene crosslinked collagen-based nerve conduit | |||
| Graphene modification | Raw 264.7 macrophages | Macrophages seeded onto graphene crosslinked conduit show high expression of CD163 and CD206 markers | [190] |
| Microporous nanodiamonds/PCL nerve bridge | |||
| Nanodiamond addition | Sciatic nerve in male SD rats | Nanodiamonds induce M2 polarization of macrophages | [191] |
| Biomaterial | Model | Level | Delivery | Duration | Therapeutic | Effect | Ref |
|---|---|---|---|---|---|---|---|
| Peripheral Nerve (Decellularized hydrogel) | SCI Rat (Sprague Dawley) | C3/C4 Unilateral contusion | Injection | 14 days | none | No change M1 CCR7+ cells ↑M2 CD206+ cells | [211] |
| Brain, cord and urinary bladder (Decellularized hydrogel) | SCI Rat (Wistar) | T8 2 mm hemisected excision | Injection | 56 days | none | Spatial differences in CD86+ (M1) and CD206+ (M2) cells ↑ argninase-1 expression | [288] |
| Urinary bladder (Decellularized hydrogel) | Rat (Sprague Dawley) | Middle cerebral artery occlusion Cortex | Injection | 14 days | none | No change CD86+ (M1) cells CD206+ (M2) Present in hydrogel | [212] |
| (Decellularized hydrogel) | Rat (Sprague Dawley) | T9-10 200 kdyne Compression (Moderate- severe injury) | Injection | 56 days | none | ↓ total ED1+ cells ↓ED-1+, CD86+ (M1) cells ↑ ED-1+, Arginase-1+ (M2) cells ↑ Expression CD206, arginase1, and IL-10 | [241] |
| Collagen | SCI Rat (Sprague Dawley) | T7-11 2 mm Lateral hemisected excision | Scaffold | 28 days | MSCs | ↓total CD68+ macrophages ↑ CD206, arginase-1, and IL-10 expression | [240] |
| Gelatin | SCI Rat (Sprague Dawley) | T10-11 | Scaffold | 56 days | MSCs | ↓total CD68+ macrophages ↓ IL-1β and TNF-α expression | [243] |
| HA combined with PCL spun fibers | SCI Rat (Sprague Dawley) | T9 175 kdyne compression (Moderate injury) | Scaffold | 28 days | none | No change CD86+ (M1) cells ↑M2 CD206+ (M2) cells | [255] |
| Agarose/polyethylene glycol/carbomer | SCI (C57BL/6J mice | Aneurysm clip compression | Scaffold | 9 days | MSCs | ↑TNF-α (M1) and 10-fold ↑ arginase-1 expression | [218] |
| Chitosan and water as fragmented physical hydrogel suspension | Rat (Wistar) | Bilateral dorsal over-hemisection | Injection | 56 days | none | ↓ CD86+ (M1) expression ↑M2 CD206+ (M2) expression | [213] |
| Biomaterial | Model | Level | Delivery | Duration | Therapeutic | Effect | Ref |
|---|---|---|---|---|---|---|---|
| Oligo[poly(ethylene glycol) fumarate (OPF) plus Matrigel | SCI Rat (Sprague Dawley) | T9-10 Complete transection | Scaffold | 56 days | Schwann cells | No change CD86+ (M1) cells ↑M2 CD206+ (M2) cells | [208] |
| Poly(glycolic acid) (PGA) | SCI Rat (Sprague Dawley) | T10-11 3 mm Lateral hemisected excision | Scaffold | 56 days | NPCs | ↓CD68+ macrophages ↑M2 CD206+ (M2) cells | [273] |
| PLGA and Poly-L-lysine | SCI Rat (Sprague Dawley) | T10 220 kdyne contusion (Moderate-severe injury) | Scaffold | 84 days | none | ↓CD74 + (M1) cells ↑CD206+ CD163+ and Arginase-1+ (M2) cells | [274] |
| GO | Rat (Wistar) | C6 2 mm lateral hemisected excision | Scaffold | 10/30 days | none | Day 10: No change CD80+ (M1) and CD163+ (M2) cells Day 30: ↓CD86+ (M1) cells Trend ↑ CD163+ (M2) cells | [308,309] |
| PLGA | mice (C57/BL6) | T9-T10 2.25 mm lateral hemisected excision | Scaffold | 30 days | none | No change F4/80+ arginase1- (M1) cells ↑ F4/80+ arginase 1+ (M2) cells | [310] |
| PLGA | mice (C57/BL6) | T9-T10 2 mm lateral hemisected excision | Scaffold | 28 days | none | ↓ CD86, MHCII, iNOS (M1) expression ↑CD206, (Retnla), arginase-1 (M2) expression ↑Hoechst+/F4/80+/Arginase-1+ (M2) cells | [209] |
| PLGA | mice (C57/BL6) | C5 1.15 mm lateral hemisected excision | Scaffold | 84 days | none | ↑F4/80+/Arginase-1+ (M2) cells | [311] |
| Biomaterial | Model | Level | Delivery | Duration | Therapeutic | Effect | Ref |
|---|---|---|---|---|---|---|---|
| Chitosan | SCI Rat (Sprague Dawley) | C3-C4 1–2 mm lateral hemisected excision | Spongostan® | 7 days | None | ↓ ED-1+ macrophages | [299] |
| Minocycline loaded poly-caprolactone nanoparticles | Mice (C57/BL6) | T12 Unilateral stab/ injection | Cord Injection | 15 days | None | ↓ CD68+ macrophages | [298] |
| Flavopiridol loaded PLGA nanoparticles | SCI Rat (Sprague Dawley) | T9-10 Lateral hemisection | Topical delivery | 42 days | None | ↓CD68+ macrophages ↓IL-6 and IL-1β and increased ↑IL-10 expression | [323] |
| Hirudin loaded Pluronic F-127 hydrogels | mice | T8 Contusion with 0.5 mm displacement | Cord injection | 28 days | none | ↓CD68+ macrophages | [301] |
| PLGA nanoparticles | mice (C57/BL6) | T9-10 1.2 mm lateral hemisected excision | PLGA Scaffold | 84 days | none | ↓ iNOS+, CD86+, and MCP-1+(M1) expression ↑ CD206+ and IL-10+ (M2) expression | [324] |
| Minocycline hydrochloride complexed with polysaccharide dextran sulphate | SCI Rat (Sprague Dawley) | C5 200 kdyne contusion (Moderate to severe injury) | Agarose Scaffold and intraperitoneal injection | 42 days | none | ↓CD68+ macrophages ↓ ratio iNOS+, CD68+ (M1) cells to arginase 1+, ↑CD68+ (M2) cells | [297] |
| Chicago sky blue loaded liposomes | SCI Rat (Sprague Dawley) | T9 150 kdyne contusion (Moderate injury) | Intravenous injection | 4 days | none | ↑ CCL2, IL-1β, and iNOS (M1) expression ↑ arginase I and TGF-β (M2) expression ↑ arginase I+, CD68+ (M2) cells | [326] |
| Biomaterial | Model | Level | Delivery | Duration | Therapeutic | Effect | Ref |
|---|---|---|---|---|---|---|---|
| None Cell suspension | SCI Rat (Sprague Dawley) | T9-10 200 kdyne contusion (Moderate to severe injury) | Injection | 35 days | MSCs | ↓ iNOS+, CD16+/32+ (M1) cells ↑ arginase 1+, CD206+(M2) cells | [337] |
| Gelatin sponge scaffolds sheathed with PLGA film | SCI Rat (Sprague Dawley) | T10-11 Full transection | Scaffold | 56 days | MSCs | ↓ CD68+ macrophages ↓ CD68+, IL-1β (M1) cells ↓ CD68+, TNF-α (M2) cells | [243] |
| PLGA | SCI Rat (Sprague Dawley) | T9-10 4 mm lateral hemisected excision | Scaffold | 10 days | MSCs | ↓ CD68+ macrophages ↑CD68+ anginase-1+ (M2) cells | [339] |
| Methacrylamide chitosan hydrogel | Rats (Fischer 344) | T8-9 2.0–2.5 mm lateral hemisected excision | Scaffold | 56 days | NPSCs | ↓ ED-1+ macrophages | [340] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dervan, A.; Franchi, A.; Almeida-Gonzalez, F.R.; Dowling, J.K.; Kwakyi, O.B.; McCoy, C.E.; O’Brien, F.J.; Hibbitts, A. Biomaterial and Therapeutic Approaches for the Manipulation of Macrophage Phenotype in Peripheral and Central Nerve Repair. Pharmaceutics 2021, 13, 2161. https://doi.org/10.3390/pharmaceutics13122161
Dervan A, Franchi A, Almeida-Gonzalez FR, Dowling JK, Kwakyi OB, McCoy CE, O’Brien FJ, Hibbitts A. Biomaterial and Therapeutic Approaches for the Manipulation of Macrophage Phenotype in Peripheral and Central Nerve Repair. Pharmaceutics. 2021; 13(12):2161. https://doi.org/10.3390/pharmaceutics13122161
Chicago/Turabian StyleDervan, Adrian, Antonio Franchi, Francisco R. Almeida-Gonzalez, Jennifer K. Dowling, Ohemaa B. Kwakyi, Claire E. McCoy, Fergal J. O’Brien, and Alan Hibbitts. 2021. "Biomaterial and Therapeutic Approaches for the Manipulation of Macrophage Phenotype in Peripheral and Central Nerve Repair" Pharmaceutics 13, no. 12: 2161. https://doi.org/10.3390/pharmaceutics13122161