
New Therapeutic Approaches for Allergy: A Review of Cell Therapy and Bio- or Nano-Material-Based Strategies



Abstract
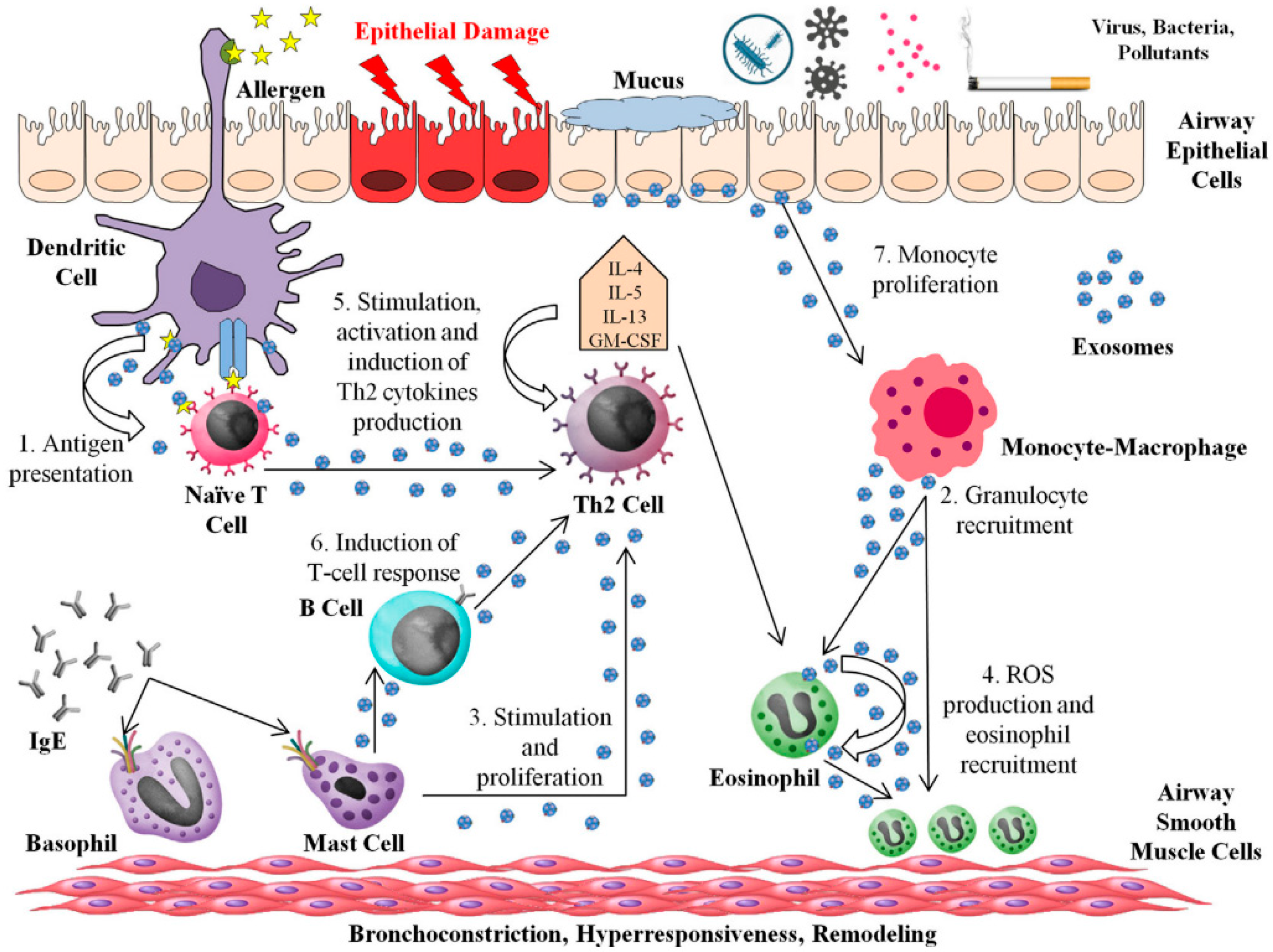
:1. Introduction
2. Cell-Based Therapeutic Actions toward Tolerance State in Allergy
2.1. Key Players Involved in Allergy and Their Use for Cell Therapy
2.1.1. Dendritic Cells (DCs)
2.1.2. B lymphocytes and Immunoglobulin E
2.1.3. Mastocytes
2.1.4. Regulatory T Cells (Tregs)
2.1.5. Exosomes
2.2. Other Cell-Based Therapeutic Actions in Allergy
2.2.1. Mesenchymal Stromal Cells (MSC)
2.2.2. Microbiome
3. Bio- and Nano-Material-Based Strategies for Allergy Therapy
- Using bio- and nano-materials with a direct effect on cells involved in the allergic response.
- Use of bio- and nano-materials as allergen delivery vehicles for immunotherapy.
- Use of bio- and nano-materials as co-delivery systems containing the allergen and immunomodulatory molecules.
3.1. Using Bio- and Nano-Materials with a Direct Effect on Cells Involved in the Allergic Response
3.2. Use of Bio- and Nano-Materials as Allergen Delivery Vehicles for Immunotherapy
3.3. Use of Bio- and Nano-Materials as Co-Delivery Systems Containing the Allergen and Immunomodulatory Molecules
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Lambrecht, B.N.; Hammad, H. The Immunology of Asthma. Nat. Immunol. 2015, 16, 45–56. [Google Scholar] [CrossRef]
- Ozdemir, C.; Akdis, M.; Akdis, C.A. T-Cell Response to Allergens. Chem. Immunol. Allergy 2010, 95, 22–44. [Google Scholar]
- Méndez-Enríquez, E.; Hallgren, J. Mast Cells and Their Progenitors in Allergic Asthma. Front. Immunol. 2019, 10, 821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muraro, A. The European Academy of Allergy and Clinical Immunology (EAACI) Advocacy Manifesto Tackling the Allergy Crisis in Europe—Concerted Policy Action Needed; EAACI—EU Liaison Office: Brussels, Belgium, 2015. [Google Scholar]
- Komlósi, Z.I.; Kovács, N.; Sokolowska, M.; van de Veen, W.; Akdis, M.; Akdis, C.A. Mechanisms of Subcutaneous and Sublingual Aeroallergen Immunotherapy: What Is New? Immunol. Allergy Clin. 2020, 40, 1–14. [Google Scholar] [CrossRef]
- Farokhzad, O.C.; Langer, R. Impact of Nanotechnology on Drug Delivery. ACS Nano 2009, 3, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Kantoff, P.W.; Wooster, R.; Farokhzad, O.C. Cancer Nanomedicine: Progress, Challenges and Opportunities. Nat. Rev. Cancer 2017, 17, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Nakamura, H.; Maeda, H. The EPR Effect: Unique Features of Tumor Blood Vessels for Drug Delivery, Factors Involved, and Limitations and Augmentation of the Effect. Adv. Drug Deliv. Rev. 2011, 63, 136–151. [Google Scholar] [CrossRef]
- Matsumura, Y.; Maeda, H. A New Concept for Macromolecular Therapeutics in Cancer Chemotherapy: Mechanism of Tumoritropic Accumulation of Proteins and the Antitumor Agent Smancs. Cancer Res. 1986, 46, 6387–6392. [Google Scholar]
- Bertrand, N.; Wu, J.; Xu, X.; Kamaly, N.; Farokhzad, O.C. Cancer Nanotechnology: The Impact of Passive and Active Targeting in the Era of Modern Cancer Biology. Adv. Drug Deliv. Rev. 2014, 66, 2–25. [Google Scholar] [CrossRef] [Green Version]
- Sindhwani, S.; Syed, A.M.; Ngai, J.; Kingston, B.R.; Maiorino, L.; Rothschild, J.; MacMillan, P.; Zhang, Y.; Rajesh, N.U.; Hoang, T.; et al. The Entry of Nanoparticles into Solid Tumours. Nat. Mater. 2020, 19, 566–575. [Google Scholar] [CrossRef]
- Shao, K.; Singha, S.; Clemente-Casares, X.; Tsai, S.; Yang, Y.; Santamaria, P. Nanoparticle-Based Immunotherapy for Cancer. ACS Nano 2015, 9, 16–30. [Google Scholar] [CrossRef]
- Feng, X.; Liu, J.; Xu, W.; Li, G.; Ding, J. Tackling Autoimmunity with Nanomedicines. Nanomedicine 2020, 15, 1585–1597. [Google Scholar] [CrossRef] [PubMed]
- Paris, J.L.; Baeza, A. Nano- and Microscale Drug Delivery Approaches for Therapeutic Immunomodulation. ChemNanoMat 2021, 7, 773–788. [Google Scholar] [CrossRef]
- Fontana, F.; Figueiredo, P.; Bauleth-Ramos, T.; Correia, A.; Santos, H.A. Immunostimulation and Immunosuppression: Nanotechnology on the Brink. Small Methods 2018, 2, 1700347. [Google Scholar] [CrossRef]
- Gordon, J.R.; Ma, Y.; Churchman, L.; Gordon, S.A.; Dawicki, W. Regulatory Dendritic Cells for Immunotherapy in Immunologic Diseases. Front. Immunol. 2014, 5, 7. [Google Scholar] [CrossRef]
- Schülke, S. Induction of Interleukin-10 Producing Dendritic Cells As a Tool to Suppress Allergen-Specific T Helper 2 Responses. Front. Immunol. 2018, 9, 455. [Google Scholar] [CrossRef]
- Ness, S.; Lin, S.; Gordon, J.R. Regulatory Dendritic Cells, T Cell Tolerance, and Dendritic Cell Therapy for Immunologic Disease. Front. Immunol. 2021, 12, 633436. [Google Scholar] [CrossRef] [PubMed]
- Koya, T.; Matsuda, H.; Takeda, K.; Matsubara, S.; Miyahara, N.; Balhorn, A.; Dakhama, A.; Gelfand, E.W. IL-10–Treated Dendritic Cells Decrease Airway Hyperresponsiveness and Airway Inflammation in Mice. J. Allergy Clin. Immunol. 2007, 119, 1241–1250. [Google Scholar] [CrossRef]
- Henry, E.; Desmet, C.J.; Garzé, V.; Fiévez, L.; Bedoret, D.; Heirman, C.; Faisca, P.; Jaspar, F.J.; Gosset, P.; Jacquet, A.P.A.; et al. Dendritic Cells Genetically Engineered to Express IL-10 Induce Long-Lasting Antigen-Specific Tolerance in Experimental Asthma. J. Immunol. 2008, 181, 7230–7242. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes-Rius, A.; Desai, A.; Yuen, D.; Johnston, A.P.R.; Voelcker, N.H. Inducing Immune Tolerance with Dendritic Cell-Targeting Nanomedicines. Nat. Nanotechnol. 2021, 16, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Selb, R.; Eckl-Dorna, J.; Neunkirchner, A.; Schmetterer, K.; Marth, K.; Gamper, J.; Jahn-Schmid, B.; Pickl, W.F.; Valenta, R.; Niederberger, V. CD23 Surface Density on B Cells Is Associated with IgE Levels and Determines IgE-Facilitated Allergen Uptake, as Well as Activation of Allergen-Specific T Cells. J. Allergy Clin. Immunol. 2017, 139, 290–299.e4. [Google Scholar] [CrossRef] [Green Version]
- Asrat, S.; Kaur, N.; Liu, X.; Ben, L.-H.; Kajimura, D.; Murphy, A.J.; Sleeman, M.A.; Limnander, A.; Orengo, J.M. Chronic Allergen Exposure Drives Accumulation of Long-Lived IgE Plasma Cells in the Bone Marrow, Giving Rise to Serological Memory. Sci. Immunol. 2020, 5, 43. [Google Scholar] [CrossRef]
- Noh, G.; Lee, J.H. Regulatory B Cells and Allergic Diseases. Allergy. Asthma Immunol. Res. 2011, 3, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Noh, J.; Noh, G.; Kim, H.S.; Kim, R.A.; Choi, W.S. Allergen-Specific Responses of CD19(+)CD5(+)Foxp3(+) Regulatory B Cells (Bregs) and CD4(+)Foxp3(+) Regulatory T Cell (Tregs) in Immune Tolerance of Cow Milk Allergy of Late Eczematous Reactions. Cell. Immunol. 2012, 274, 109–114. [Google Scholar] [CrossRef]
- Braza, F.; Chesne, J.; Durand, M.; Dirou, S.; Brosseau, C.; Mahay, G.; Cheminant, M.A.; Magnan, A.; Brouard, S. A Regulatory CD9+ B-Cell Subset Inhibits HDM-Induced Allergic Airway Inflammation. Allergy 2015, 70, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Wilson, T.M.; Metcalfe, D.D. The Mast Cell and Allergic Diseases: Role in Pathogenesis and Implications for Therapy. Clin. Exp. Allergy 2008, 38, 4–18. [Google Scholar] [CrossRef]
- Xie, G.; Yang, H.; Peng, X.; Lin, L.; Wang, J.; Lin, K.; Cui, Z.; Li, J.; Xiao, H.; Liang, Y.; et al. Mast Cell Exosomes Can Suppress Allergic Reactions by Binding to IgE. J. Allergy Clin. Immunol. 2018, 141, 788–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchan, J.R.; Parker, R. The Two Faces of MiRNA. Science 2007, 318, 1877–1878. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, S.; Powrie, F. Emerging Challenges in Regulatory T Cell Function and Biology. Science 2007, 317, 627–629. [Google Scholar] [CrossRef]
- Feuerer, M.; Hill, J.A.; Mathis, D.; Benoist, C. Foxp3+ Regulatory T Cells: Differentiation, Specification, Subphenotypes. Nat. Immunol. 2009, 10, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 Programs the Development and Function of CD4+CD25+ Regulatory T Cells. Nat. Immunol. 2003, 4, 330–336. [Google Scholar] [CrossRef]
- Noval Rivas, M.; Chatila, T.A. Regulatory T Cells in Allergic Diseases. J. Allergy Clin. Immunol. 2016, 138, 639–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, C.L.; Christie, J.; Ramsdell, F.; Brunkow, M.E.; Ferguson, P.J.; Whitesell, L.; Kelly, T.E.; Saulsbury, F.T.; Chance, P.F.; Ochs, H.D. The Immune Dysregulation, Polyendocrinopathy, Enteropathy, X-Linked Syndrome (IPEX) Is Caused by Mutations of FOXP3. Nat. Genet. 2001, 27, 20–21. [Google Scholar] [CrossRef] [PubMed]
- Baatjes, A.J.; Smith, S.G.; Watson, R.; Howie, K.; Murphy, D.; Larché, M.; Denburg, J.A.; Inman, M.D.; O’Byrne, P.M. T Regulatory Cell Phenotypes in Peripheral Blood and Bronchoalveolar Lavage from Non-Asthmatic and Asthmatic Subjects. Clin. Exp. Allergy 2015, 45, 1654–1662. [Google Scholar] [CrossRef]
- Scherf, W.; Burdach, S.; Hansen, G. Reduced Expression of Transforming Growth Factor Β1 Exacerbates Pathology in an Experimental Asthma Model. Eur. J. Immunol. 2005, 35, 198–206. [Google Scholar] [CrossRef]
- Gri, G.; Piconese, S.; Frossi, B.; Manfroi, V.; Merluzzi, S.; Tripodo, C.; Viola, A.; Odom, S.; Rivera, J.; Colombo, M.P.; et al. CD4+CD25+ Regulatory T Cells Suppress Mast Cell Degranulation and Allergic Responses through OX40-OX40L Interaction. Immunity 2008, 29, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.W.; Hillsamer, P.; Banham, A.H.; Kim, C.H. Cutting Edge: Direct Suppression of B Cells by CD4+CD25+ Regulatory T Cells. J. Immunol. 2005, 175, 4180–4183. [Google Scholar] [CrossRef] [Green Version]
- Meiler, F.; Klunker, S.; Zimmermann, M.; Akdis, C.A.; Akdis, M. Distinct Regulation of IgE, IgG4 and IgA by T Regulatory Cells and Toll-like Receptors. Allergy 2008, 63, 1455–1463. [Google Scholar] [CrossRef]
- Martín-Orozco, E.; Norte-Muñoz, M.; Martínez-García, J. Regulatory T Cells in Allergy and Asthma. Front. Pediatr. 2017, 5, 117. [Google Scholar] [CrossRef] [PubMed]
- Shreffler, W.G.; Wanich, N.; Moloney, M.; Nowak-Wegrzyn, A.; Sampson, H.A. Association of Allergen-Specific Regulatory T Cells with the Onset of Clinical Tolerance to Milk Protein. J. Allergy Clin. Immunol. 2009, 123, 43–52.e7. [Google Scholar] [CrossRef]
- Syed, A.; Garcia, M.A.; Lyu, S.C.; Bucayu, R.; Kohli, A.; Ishida, S.; Berglund, J.P.; Tsai, M.; Maecker, H.; O’Riordan, G.; et al. Peanut Oral Immunotherapy Results in Increased Antigen-Induced Regulatory T-Cell Function and Hypomethylation of Forkhead Box Protein 3 (FOXP3). J. Allergy Clin. Immunol. 2014, 133, 500–510.e11. [Google Scholar] [CrossRef] [Green Version]
- Van Wijk, F.; Wehrens, E.J.M.; Nierkens, S.; Boon, L.; Kasran, A.; Pieters, R.; Knippels, L.M.J. CD4+CD25+ T Cells Regulate the Intensity of Hypersensitivity Responses to Peanut, but Are Not Decisive in the Induction of Oral Sensitization. Clin. Exp. Allergy 2007, 37, 572–581. [Google Scholar] [CrossRef]
- Kearley, J.; Barker, J.E.; Robinson, D.S.; Lloyd, C.M. Resolution of Airway Inflammation and Hyperreactivity after in Vivo Transfer of CD4+CD25+ Regulatory T Cells Is Interleukin 10 Dependent. J. Exp. Med. 2005, 202, 1539–1547. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, H.; Takahashi, K.; Tanaka, H.; Nagai, H.; Inagaki, N. Overcoming Food Allergy through Acquired Tolerance Conferred by Transfer of Tregs in a Murine Model. Allergy 2012, 67, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Lan, Q.; Chen, M.; Chen, H.; Zhu, N.; Zhou, X.; Wang, J.; Fan, H.; Yan, C.-S.; Kuang, J.-L.; et al. Adoptive Transfer of Induced-Treg Cells Effectively Attenuates Murine Airway Allergic Inflammation. PLoS ONE 2012, 7, e40314. [Google Scholar] [CrossRef] [PubMed]
- Leslie, M. Regulatory T Cells Get Their Chance to Shine. Science 2011, 332, 1020–1021. [Google Scholar] [CrossRef] [PubMed]
- Skuljec, J.; Chmielewski, M.; Happle, C.; Habener, A.; Busse, M.; Abken, H.; Hansen, G. Chimeric Antigen Receptor-Redirected Regulatory T Cells Suppress Experimental Allergic Airway Inflammation, a Model of Asthma. Front. Immunol. 2017, 8, 1125. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.G.; Henden, A.S. Infectious Complications of CAR T-Cell Therapy: A Clinical Update. Ther. Adv. Infect. Dis. 2021, 8, 204993612110367. [Google Scholar] [CrossRef]
- Huang, F.; Jia, H.; Zou, Y.; Yao, Y.; Deng, Z. Exosomes: An Important Messenger in the Asthma Inflammatory Microenvironment. J. Int. Med. Res. 2020, 48, 030006052090322. [Google Scholar] [CrossRef] [Green Version]
- Sastre, B.; Cañas, J.A.; Rodrigo-Muñoz, J.M.; del Pozo, V. Novel Modulators of Asthma and Allergy: Exosomes and MicroRNAs. Front. Immunol. 2017, 8, 826. [Google Scholar] [CrossRef]
- Srinivasan, A.; Sundar, I.K. Recent Updates on the Role of Extracellular Vesicles in the Pathogenesis of Allergic Asthma. Extracell. Vesicles Circ. Nucleic Acids 2021, 2, 127–147. [Google Scholar] [CrossRef]
- Östman, S.; Taube, M.; Telemo, E. Tolerosome-Induced Oral Tolerance Is MHC Dependent. Immunology 2005, 116, 464–476. [Google Scholar] [CrossRef]
- Karlsson, M.; Lundin, S.; Dahlgren, U.; Kahu, H.; Pettersson, I.; Telemo, E. “Tolerosomes” Are Produced by Intestinal Epithelial Cells. Eur. J. Immunol. 2001, 31, 2892–2900. [Google Scholar] [CrossRef]
- Almqvist, N.; Lönnqvist, A.; Hultkrantz, S.; Rask, C.; Telemo, E. Serum-Derived Exosomes from Antigen-Fed Mice Prevent Allergic Sensitization in a Model of Allergic Asthma. Immunology 2008, 125, 21–27. [Google Scholar] [CrossRef]
- Pei, W.; Li, X.; Bi, R.; Zhang, X.; Zhong, M.; Yang, H.; Zhang, Y.; Lv, K. Exosome Membrane-Modified M2 Macrophages Targeted Nanomedicine: Treatment for Allergic Asthma. J. Control. Release 2021, 338, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Macias, M.I.; Grande, J.; Moreno, A.; Domínguez, I.; Bornstein, R.; Flores, A.I. Isolation and Characterization of True Mesenchymal Stem Cells Derived from Human Term Decidua Capable of Multilineage Differentiation into All 3 Embryonic Layers. Am. J. Obstet. Gynecol. 2010, 203, 495.e9–495.e23. [Google Scholar] [CrossRef] [PubMed]
- Planat-Benard, V.; Varin, A.; Casteilla, L. MSCs and Inflammatory Cells Crosstalk in Regenerative Medicine: Concerted Actions for Optimized Resolution Driven by Energy Metabolism. Front. Immunol. 2021, 12, 1465. [Google Scholar] [CrossRef]
- Li, H.; Tian, Y.; Xie, L.; Liu, X.; Huang, Z.; Su, W. Mesenchymal Stem Cells in Allergic Diseases: Current Status. Allergol. Int. 2020, 69, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, M.; Sueblinvong, V.; Eisenhauer, P.; Ziats, N.P.; LeClair, L.; Poynter, M.E.; Steele, C.; Rincon, M.; Weiss, D.J. Bone Marrow-Derived Mesenchymal Stromal Cells Inhibit Th2-Mediated Allergic Airways Inflammation in Mice. Stem Cells 2011, 29, 1137–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.-S.; Park, H.-K.; Park, H.-Y.; Jung, J.S.; Jeon, S.-G.; Kim, Y.-K.; Roh, H.J. IFATS Collection: Immunomodulatory Effects of Adipose Tissue-Derived Stem Cells in an Allergic Rhinitis Mouse Model. Stem Cells 2009, 27, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Karp, J.M.; Leng Teo, G.S. Mesenchymal Stem Cell Homing: The Devil Is in the Details. Cell Stem Cell 2009, 4, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Huang, W.; Chen, X.; Lian, Y.; Wang, J.; Cai, C.; Huang, L.; Wang, T.; Ren, J.; Xiang, A.P. CXCR5-Overexpressing Mesenchymal Stromal Cells Exhibit Enhanced Homing and Can Decrease Contact Hypersensitivity. Mol. Ther. 2017, 25, 1434–1447. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Sha, J.; Meng, C.; Zhu, D. Mesenchymal Stem Cell-Based Therapy for Allergic Rhinitis. Stem Cells Int. 2020, 2020, 2367524. [Google Scholar] [CrossRef]
- Baiula, M.; Spampinato, S. Mesenchymal Stem Cell Secretome to Control Inflammation in Allergic Conjunctivitis. Eye Sci. 2015, 30, 140–142. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.-B.; Zhang, H.-Y.; Wang, C.; He, B.-X.; Liu, X.-Q.; Meng, X.-C.; Peng, Y.-Q.; Xu, Z.-B.; Fan, X.-L.; Wu, Z.-J.; et al. Small Extracellular Vesicles Derived from Human Mesenchymal Stromal Cells Prevent Group 2 Innate Lymphoid Cell-dominant Allergic Airway Inflammation through Delivery of MiR-146a-5p. J. Extracell. Vesicles 2020, 9, 1723260. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.M.; Zhuansun, Y.X.; Chen, R.; Lin, L.; Lin, Y.; Li, J.G. Mesenchymal Stem Cell Exosomes Promote Immunosuppression of Regulatory T Cells in Asthma. Exp. Cell Res. 2018, 363, 114–120. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.; McCoy, K.D.; Macpherson, A.J. Use of Axenic Animals in Studying the Adaptation of Mammals to Their Commensal Intestinal Microbiota. Semin. Immunol. 2007, 19, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Ipci, K.; Altıntoprak, N.; Muluk, N.B.; Senturk, M.; Cingi, C. The Possible Mechanisms of the Human Microbiome in Allergic Diseases. Eur. Arch. Oto-Rhino-Laryngol. 2017, 274, 617–626. [Google Scholar] [CrossRef]
- Strachan, D.P. Hay Fever, Hygiene, and Household Size. BMJ Br. Med. J. 1989, 299, 1259–1260. [Google Scholar] [CrossRef] [Green Version]
- Begley, L.; Madapoosi, S.; Opron, K.; Ndum, O.; Baptist, A.; Rysso, K.; Erb-Downward, J.R.; Huang, Y.J. Gut Microbiota Relationships to Lung Function and Adult Asthma Phenotype: A Pilot Study. BMJ Open Respir. Res. 2018, 5, e000324. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Santamarina, A.; Gonzalez, E.G.; Lamas, A.; Mondragon, A.d.C.; Regal, P.; Miranda, J.M. Probiotics as a Possible Strategy for the Prevention and Treatment of Allergies. A Narrative Review. Foods 2021, 10, 701. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Z.; Wu, Y.; Jing, X.; Li, L.; Fang, X. Intestinal Bacteria Encapsulated by Biomaterials Enhance Immunotherapy. Front. Immunol. 2021, 11, 3951. [Google Scholar] [CrossRef]
- Fernández-Rivas, M.; Fernández, S.G.; Nadal, J.A.; de Durana, M.D.A.D.; García, B.E.; González-Mancebo, E.; Martín, S.; Barber, D.; Rico, P.; Tabar, A.I. Randomized Double-Blind, Placebo-Controlled Trial of Sublingual Immunotherapy with a Pru p 3 Quantified Peach Extract. Allergy 2009, 64, 876–883. [Google Scholar] [CrossRef]
- Klimek, L.; Brehler, R.; Hamelmann, E.; Kopp, M.; Ring, J.; Treudler, R.; Jakob, T.; Worm, M.; Pfaar, O. Development of Subcutaneous Allergen Immunotherapy (Part 2): Preventive Aspects and Innovations. Allergo J. Int. 2019, 28, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Andorko, J.I.; Hess, K.L.; Jewell, C.M. Harnessing Biomaterials to Engineer the Lymph Node Microenvironment for Immunity or Tolerance. AAPS J. 2015, 17, 323–338. [Google Scholar] [CrossRef] [Green Version]
- Hotaling, N.A.; Tang, L.; Irvine, D.J.; Babensee, J.E. Biomaterial Strategies for Immunomodulation. Annu. Rev. Biomed. Eng. 2015, 17, 317–349. [Google Scholar] [CrossRef] [Green Version]
- Pohlit, H.; Frey, H.; Saloga, J. Could Allergen-Specific Immunotherapy Benefit from the Use of Nanocarriers? Nanomedicine 2016, 11, 1329–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Felice, G.; Colombo, P. Nanoparticle–Allergen Complexes for Allergen Immunotherapy. Int. J. Nanomed. 2017, 12, 4493–4504. [Google Scholar] [CrossRef] [Green Version]
- Alsaleh, N.B.; Brown, J.M. Engineered Nanomaterials and Type I Allergic Hypersensitivity Reactions. Front. Immunol. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Mutschlechner, S.; Egger, M.; Briza, P.; Wallner, M.; Lackner, P.; Karle, A.; Vogt, A.B.; Fischer, G.F.; Bohle, B.; Ferreira, F. Naturally Processed T Cell–Activating Peptides of the Major Birch Pollen Allergen. J. Allergy Clin. Immunol. 2010, 125, 711–718.e2. [Google Scholar] [CrossRef]
- Blank, F.; Gerber, P.; Rothen-Rutishauser, B.; Sakulkhu, U.; Salaklang, J.; De Peyer, K.; Gehr, P.; Nicod, L.P.; Hofmann, H.; Geiser, T.; et al. Biomedical Nanoparticles Modulate Specific CD4 + T Cell Stimulation by Inhibition of Antigen Processing in Dendritic Cells. Nanotoxicology 2011, 5, 606–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, J.J.; Bateman, H.R.; Stover, A.; Gomez, G.; Norton, S.K.; Zhao, W.; Schwartz, L.B.; Lenk, R.; Kepley, C.L. Fullerene Nanomaterials Inhibit the Allergic Response. J. Immunol. 2007, 179, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Paris, J.L.; Baeza, A.; Vallet-Regí, M. Overcoming the Stability, Toxicity, and Biodegradation Challenges of Tumor Stimuli-Responsive Inorganic Nanoparticles for Delivery of Cancer Therapeutics. Expert Opin. Drug Deliv. 2019, 16, 1095–1112. [Google Scholar] [CrossRef]
- Johnson, L.; Duschl, A.; Himly, M. Nanotechnology-Based Vaccines for Allergen-Specific Immunotherapy: Potentials and Challenges of Conventional and Novel Adjuvants under Research. Vaccines 2020, 8, 237. [Google Scholar] [CrossRef] [PubMed]
- Bookstaver, M.L.; Tsai, S.J.; Bromberg, J.S.; Jewell, C.M. Improving Vaccine and Immunotherapy Design Using Biomaterials. Trends Immunol. 2018, 39, 135–150. [Google Scholar] [CrossRef]
- Gammon, J.M.; Jewell, C.M. Engineering Immune Tolerance with Biomaterials. Adv. Healthc. Mater. 2019, 8, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Smarr, C.B.; Yap, W.T.; Neef, T.P.; Pearson, R.M.; Hunter, Z.N.; Ifergan, I.; Getts, D.R.; Bryce, P.J.; Shea, L.D.; Miller, S.D. Biodegradable Antigen-Associated PLG Nanoparticles Tolerize Th2-Mediated Allergic Airway Inflammation Pre- and Postsensitization. Proc. Natl. Acad. Sci. USA 2016, 113, 5059–5064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratzer, B.; Hofer, S.; Zabel, M.; Pickl, W.F. All the Small Things: How Virus-like Particles and Liposomes Modulate Allergic Immune Responses. Eur. J. Immunol. 2020, 50, 17–32. [Google Scholar] [CrossRef]
- Scheiblhofer, S.; Machado, Y.; Feinle, A.; Thalhamer, J.; Hüsing, N.; Weiss, R. Potential of Nanoparticles for Allergen-Specific Immunotherapy–Use of Silica Nanoparticles as Vaccination Platform. Expert Opin. Drug Deliv. 2016, 13, 1777–1788. [Google Scholar] [CrossRef]
- Soni, S.S.; Rodell, C.B. Polymeric Materials for Immune Engineering: Molecular Interaction to Biomaterial Design. Acta Biomater. 2021, 133, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Pohlit, H.; Bellinghausen, I.; Schömer, M.; Heydenreich, B.; Saloga, J.; Frey, H. Biodegradable PH-Sensitive Poly(Ethylene Glycol) Nanocarriers for Allergen Encapsulation and Controlled Release. Biomacromolecules 2015, 16, 3103–3111. [Google Scholar] [CrossRef] [PubMed]
- Rebouças, J.D.S.; Irache, J.M.; Camacho, A.I.; Esparza, I.; Del Pozo, V.; Sanz, M.L.; Ferrer, M.; Gamazo, C. Development of Poly(Anhydride) Nanoparticles Loaded with Peanut Proteins: The Influence of Preparation Method on the Immunogenic Properties. Eur. J. Pharm. Biopharm. 2012, 82, 241–249. [Google Scholar] [CrossRef]
- Balenga, N.A.B.; Zahedifard, F.; Weiss, R.; Sarbolouki, M.N.; Thalhamer, J.; Rafati, S. Protective Efficiency of Dendrosomes as Novel Nano-Sized Adjuvants for DNA Vaccination against Birch Pollen Allergy. J. Biotechnol. 2006, 124, 602–614. [Google Scholar] [CrossRef]
- Keshavarz Shahbaz, S.; Varasteh, A.R.; Koushki, K.; Ayati, S.H.; Mashayekhi, K.; Sadeghi, M.; Moghadam, M.; Sankian, M. Sublingual Dendritic Cells Targeting by Aptamer: Possible Approach for Improvement of Sublingual Immunotherapy Efficacy. Int. Immunopharmacol. 2020, 85, 106603. [Google Scholar] [CrossRef]
- Rodriguez, M.J.; Ramos-Soriano, J.; Perkins, J.R.; Mascaraque, A.; Torres, M.J.; Gomez, F.; Diaz-Perales, A.; Rojo, J.; Mayorga, C. Glycosylated Nanostructures in Sublingual Immunotherapy Induce Long-Lasting Tolerance in LTP Allergy Mouse Model. Sci. Rep. 2019, 9, 4043. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.J.; Mascaraque, A.; Ramos-Soriano, J.; Torres, M.J.; Perkins, J.R.; Gomez, F.; Garrido-Arandia, M.; Cubells-Baeza, N.; Andreu, D.; Diaz-Perales, A.; et al. Pru p 3-Epitope-Based Sublingual Immunotherapy in a Murine Model for the Treatment of Peach Allergy. Mol. Nutr. Food Res. 2017, 61, 1700110. [Google Scholar] [CrossRef]
- Corthésy, B.; Bioley, G. Therapeutic Intranasal Instillation of Allergen-Loaded Microbubbles Suppresses Experimental Allergic Asthma in Mice. Biomaterials 2017, 142, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Shin, J.U.; Kim, S.H.; Noh, J.Y.; Kim, H.R.; Lee, J.; Chu, H.; Jeong, K.Y.; Park, K.H.; Kim, J.D.; et al. Successful Transdermal Allergen Delivery and Allergen-Specific Immunotherapy Using Biodegradable Microneedle Patches. Biomaterials 2018, 150, 38–48. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, X.; Liu, X.; Kumar, S.; Gochman, G.; Ji, Y.; Liao, Y.-P.; Chang, C.H.; Situ, W.; Lu, J.; et al. Use of Polymeric Nanoparticle Platform Targeting the Liver To Induce Treg-Mediated Antigen-Specific Immune Tolerance in a Pulmonary Allergen Sensitization Model. ACS Nano 2019, 13, 4778–4794. [Google Scholar] [CrossRef]
- McCarthy, D.P.; Yap, J.W.T.; Harp, C.T.; Song, W.K.; Chen, J.; Pearson, R.M.; Miller, S.D.; Shea, L.D. An Antigen-Encapsulating Nanoparticle Platform for TH1/17 Immune Tolerance Therapy. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, C.M.; Murdoch, J.R. Tolerizing Allergic Responses in the Lung. Mucosal Immunol. 2010, 3, 334–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, E.; Gurczynski, S.J.; Kramer, K.R.; Wilke, C.A.; Miller, S.D.; Moore, B.B.; Shea, L.D. Modulating Lung Immune Cells by Pulmonary Delivery of Antigen-Specific Nanoparticles to Treat Autoimmune Disease. Sci. Adv. 2020, 6, 9317–9333. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.M.; Keselowsky, B.G. Combinatorial Drug Delivery Approaches for Immunomodulation. Adv. Drug Deliv. Rev. 2017, 114, 161–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahgordi, S.; Sankian, M.; Yazdani, Y.; Mashayekhi, K.; Hasan Ayati, S.; Sadeghi, M.; Saeidi, M.; Hashemi, M. Immune Responses Modulation by Curcumin and Allergen Encapsulated into PLGA Nanoparticles in Mice Model of Rhinitis Allergic through Sublingual Immunotherapy. Int. Immunopharmacol. 2020, 84, 106525. [Google Scholar] [CrossRef]
- Hong, J.; Xiao, X.; Gao, Q.; Li, S.; Jiang, B.; Sun, X.; Ran, P.; Yang, P. Co-Delivery of Allergen Epitope Fragments and R848 Inhibits Food Allergy by Inducing Tolerogenic Dendritic Cells and Regulatory T Cells. Int. J. Nanomed. 2019, 14, 7053–7064. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Kiran Kumar, M.N.; Wu, M.X. Delivery of Allergen Powder for Safe and Effective Epicutaneous Immunotherapy. J. Allergy Clin. Immunol. 2020, 145, 597–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanagata, N. CpG Oligodeoxynucleotide Nanomedicines for the Prophylaxis or Treatment of Cancers, Infectious Diseases, and Allergies. Int. J. Nanomed. 2017, 12, 515–531. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, X.; Liu, X.; Liao, Y.-P.; Chang, C.H.; Mei, K.-C.; Jiang, J.; Tseng, S.; Gochman, G.; Huang, M.; et al. Antigen- and Epitope-Delivering Nanoparticles Targeting Liver Induce Comparable Immunotolerance in Allergic Airway Disease and Anaphylaxis as Nanoparticle-Delivering Pharmaceuticals. ACS Nano 2021, 15, 1608–1626. [Google Scholar] [CrossRef]
- Emerson, A.E.; Slaby, E.M.; Hiremath, S.C.; Weaver, J.D. Biomaterial-Based Approaches to Engineering Immune Tolerance. Biomater. Sci. 2020, 8, 7014–7032. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bio- or Nano-Material | Experimental Model | Therapeutic Mechanism | Administration Route | Specific Cell Targeting | Cargo | Ref. |
|---|---|---|---|---|---|---|
| PVA-SPIONs | Incubation with CD4+ T cells in vitro | Direct decrease in antigen processing | N/A (in vitro incubation) | No | None | [83] |
| Fullerenes | Passive anaphylaxis mouse model (DNP as hapten allergen) | Direct effect on MC, decreasing IgE-induced release of mediators | Intraperitoneal injection | No | None | [84] |
| Polystyrene and PLGA nanoparticles | Allergic airway inflammation mouse model (OVA as allergen) | Immunotherapy through allergen delivery | Intravenous injection | No | OVA (conjugated or encapsulated) | [89] |
| PEG acetal dimethacrylate nanoparticles | Incubation with DCs co-cultured with T cells in vitro | Immunotherapy through allergen delivery (pH-cleavable carrier) | N/A (in vitro incubation) | No | OVA, grass pollen extract, dust mite allergen. | [93] |
| Poly(anhydride) nanoparticles | Particle administration to non-sensitized mice | Immunotherapy through allergen delivery | Intradermal injection | No | Peanut extract | [94] |
| Dendrosomes | Prophylactic use in mice, prior to sensitization with rBetv1 | Indirect immunotherapy through delivery of plasmid encoding allergen | Footpad injection | No | Plasmid DNA encoding Betv 1a | [95] |
| PLGA nanoparticles | OVA-induced allergic rhinitis mouse model | Immunotherapy through allergen delivery | Sublingual | DC-targeted with aptamer | OVA | [96] |
| Dendrimer | Pru p 3-induced anaphylaxis mouse model | Immunotherapy through allergen delivery | Sublingual | DC-targeted with mannose | Pru p 3 peptide | [97] |
| Gas-filled microbubbles | OVA-induced allergic asthma mouse model | Immunotherapy through allergen delivery | Intranasal | No | OVA | [99] |
| Hyaluronate-based microneedle patches | Atopic dermatitis mouse model | Immunotherapy through allergen delivery | Epicutaneous | No | Der f1 dust-mite allergen | [100] |
| PLGA nanoparticles | OVA-induced allergic airway disease mouse model | Immunotherapy through allergen delivery | Intravenous injection | LSEC-targeted with mannan or peptide | OVA | [101] |
| PLGA nanoparticles | OVA-induced allergic rhinitis mouse model | Immunotherapy through co-delivery of allergen and modulatory molecules | Sublingual | No | Curcumin and OVA | [106] |
| mPEG-PDLLA nanoparticles | OVA-induced food allergy model | Immunotherapy through co-delivery of allergen and modulatory molecules | Oral | No | Peptide IK (OVA fragment) and R848 (TLR-7 ligand) | [107] |
| Dissolving microneedle patches | Peanut allergy mouse model | Immunotherapy through co-delivery of allergen and modulatory molecules | Epicutaneous | No | Peanut allergen, VD3, and CpG oligonucleotide | [108] |
| PLGA nanoparticles | OVA-induced allergic airway disease and OVA-induced anaphylaxis mouse models | Immunotherapy through co-delivery of allergen and modulatory molecules | Intravenous injection | No (comparison with LSEC-targeted without co-delivery) | OVA plus rapamycin or curcumin | [110] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paris, J.L.; de la Torre, P.; Flores, A.I. New Therapeutic Approaches for Allergy: A Review of Cell Therapy and Bio- or Nano-Material-Based Strategies. Pharmaceutics 2021, 13, 2149. https://doi.org/10.3390/pharmaceutics13122149
Paris JL, de la Torre P, Flores AI. New Therapeutic Approaches for Allergy: A Review of Cell Therapy and Bio- or Nano-Material-Based Strategies. Pharmaceutics. 2021; 13(12):2149. https://doi.org/10.3390/pharmaceutics13122149
Chicago/Turabian StyleParis, Juan L., Paz de la Torre, and Ana I. Flores. 2021. "New Therapeutic Approaches for Allergy: A Review of Cell Therapy and Bio- or Nano-Material-Based Strategies" Pharmaceutics 13, no. 12: 2149. https://doi.org/10.3390/pharmaceutics13122149