
Phyto-Phospholipid Conjugated Scorpion Venom Nanovesicles as Promising Carrier That Improves Efficacy of Thymoquinone against Adenocarcinoma Human Alveolar Basal Epithelial Cells

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Design and Optimization of THQ–PL–SV Phytosomes
2.3. Preparation of THQ–PL–SV Nanovesicles
2.3.1. Vesicle Size and Zeta Potential Determination
2.3.2. Optimization of THQ–PL–SV Nanovesicles
2.4. Fourier-Transform Infrared Characterization of the Optimized THQ–PL–SV Nanovesicles
2.5. Cytotoxicity of Optimized THQ–PL–SV
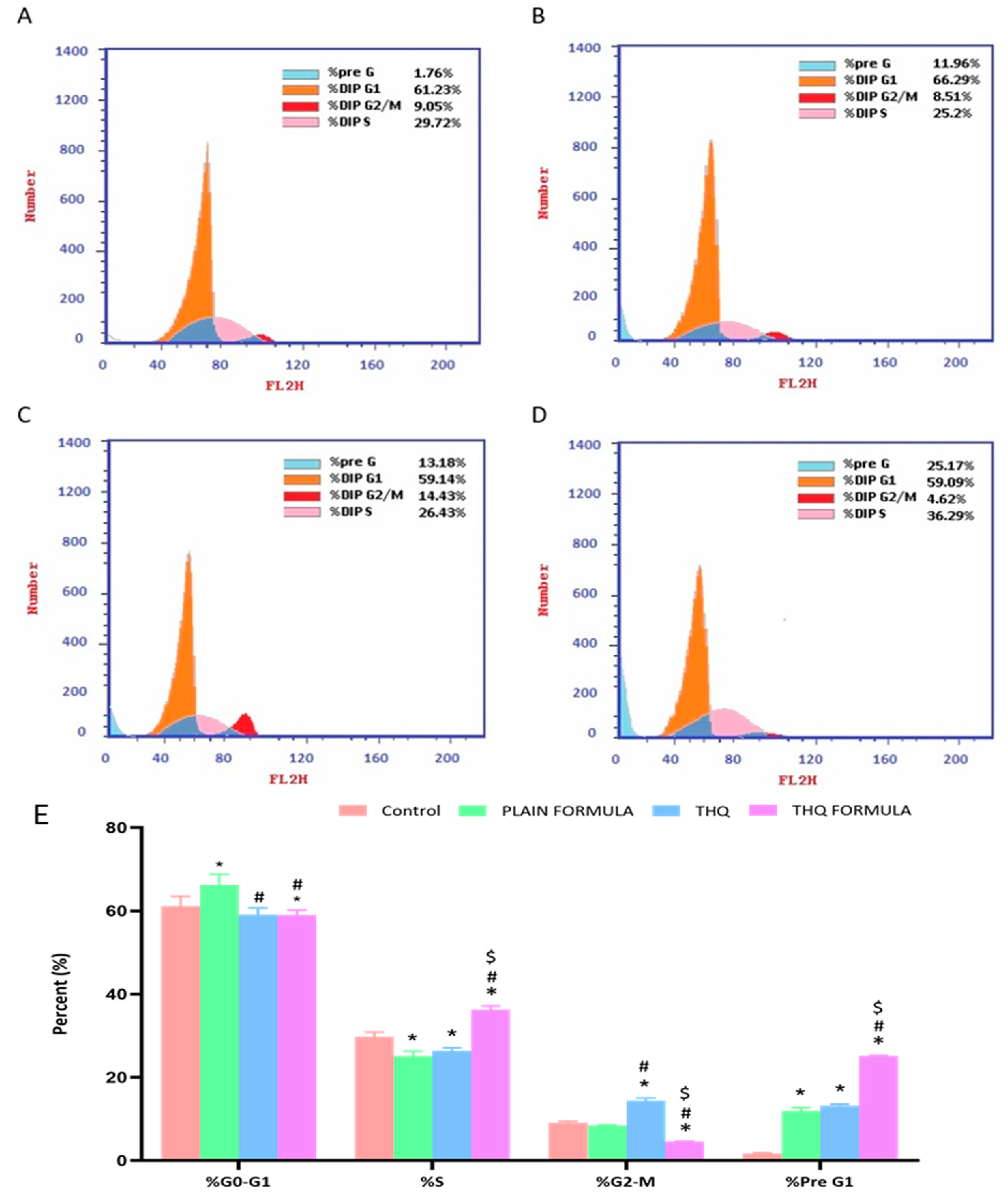
2.5.1. Cell Cycle Analysis
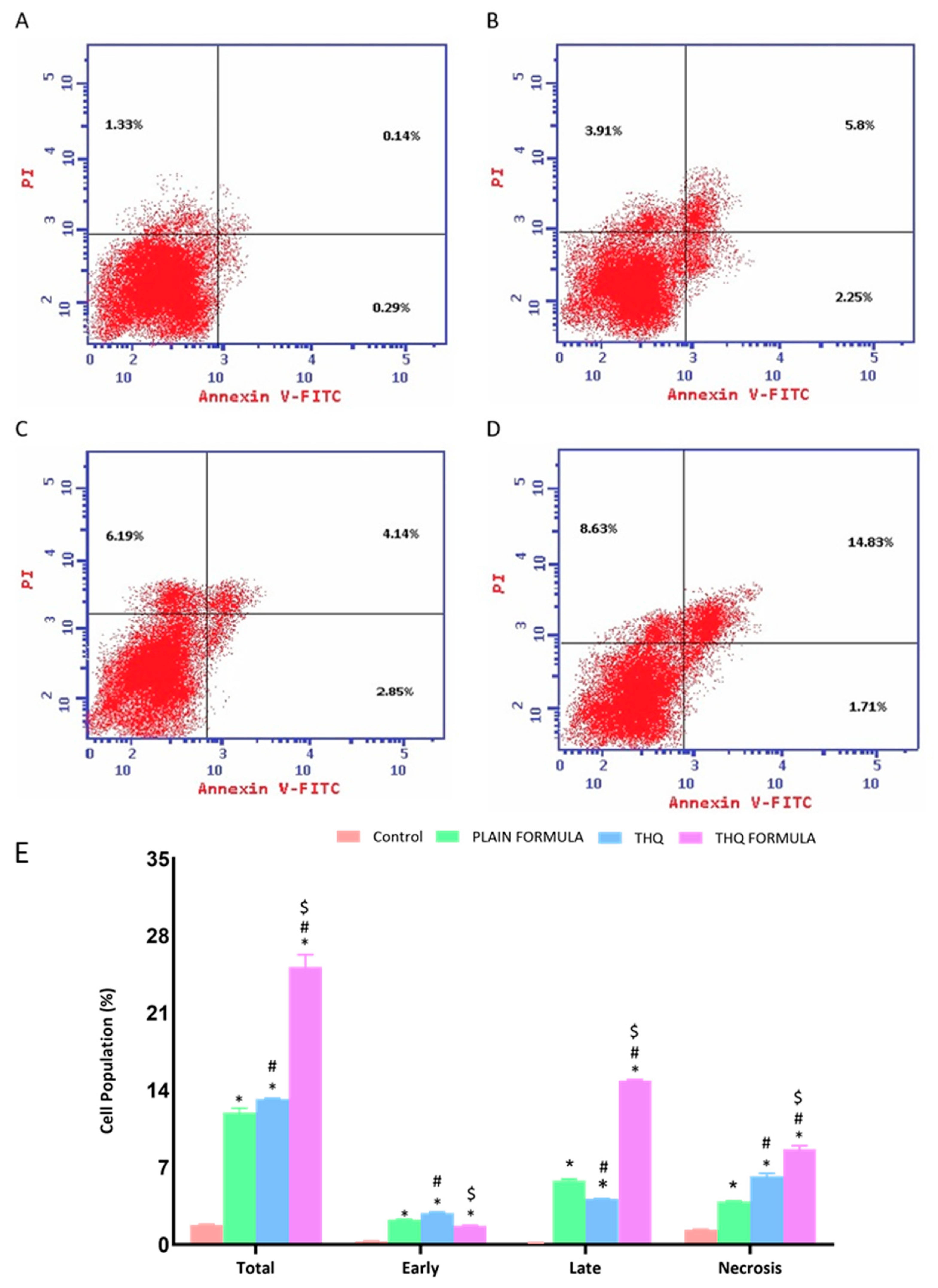
2.5.2. Analysis of Apoptosis by Annexin V Staining
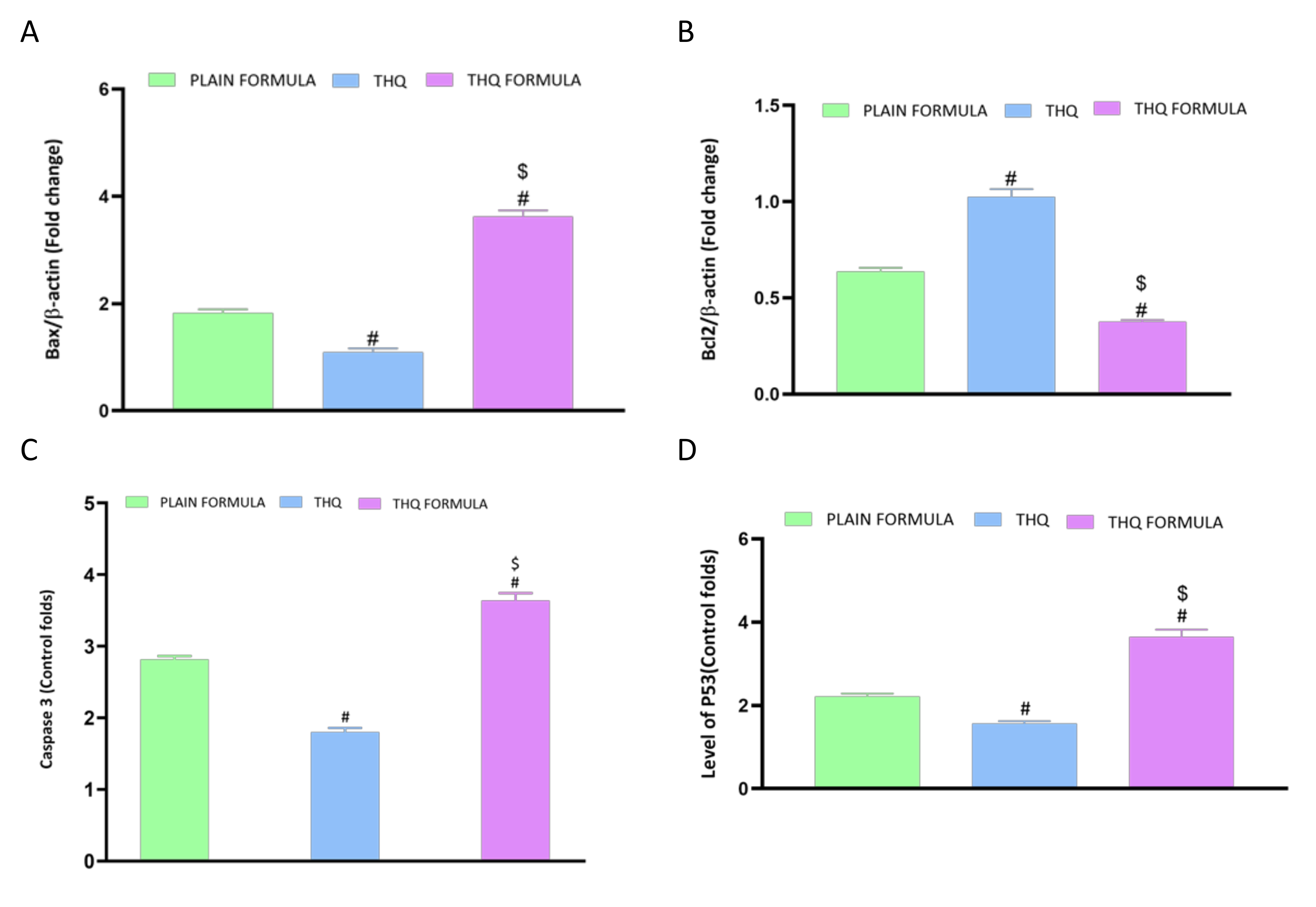
2.5.3. Real-Time Polymerase Chain Reaction (RT-PCR) for Estimation of Bcl-2, Bax, P53, Caspase 3, and TNF-α
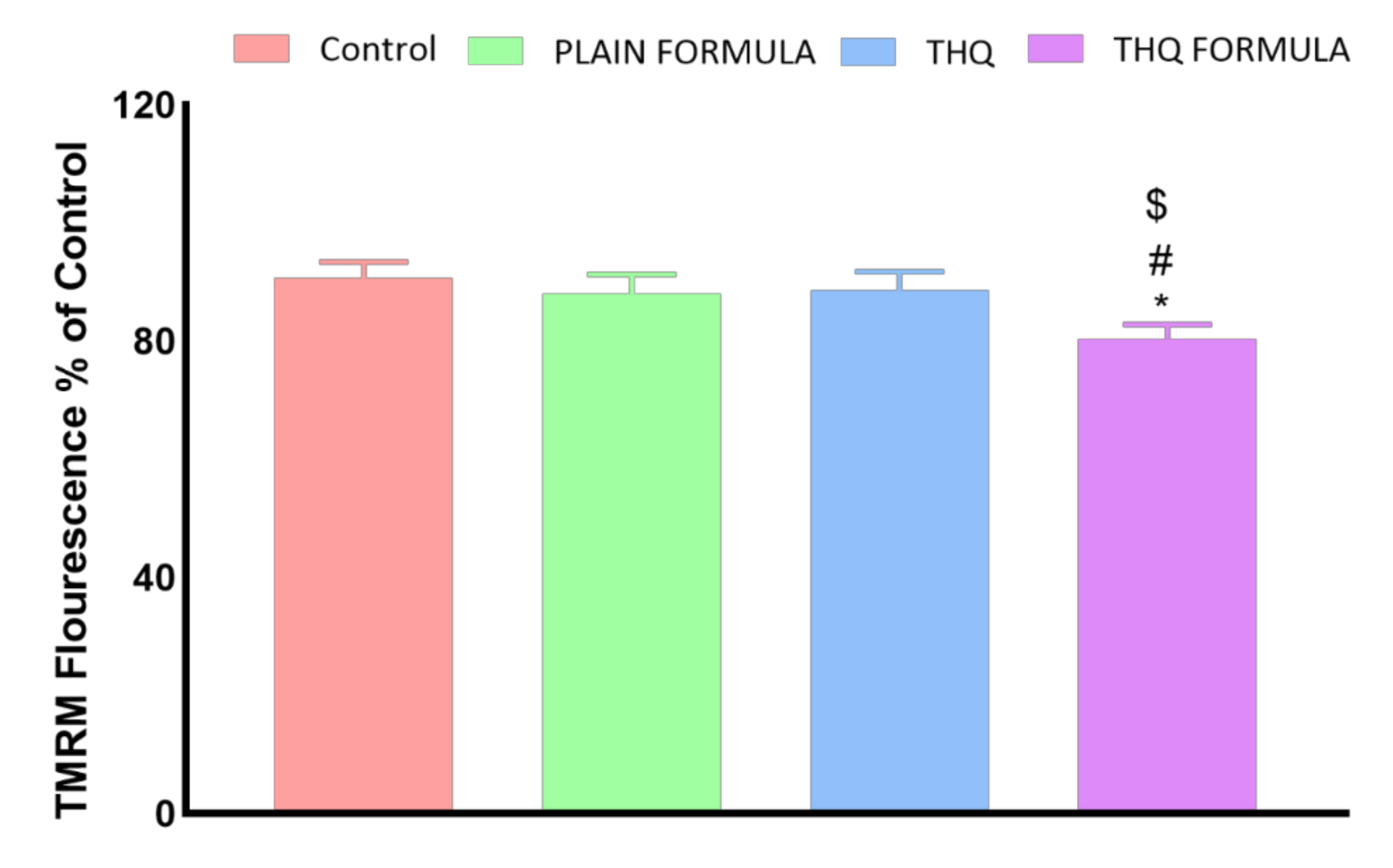
2.5.4. Determination of Mitochondrial Membrane Potential (MMP)
2.6. Statistical Analysis
3. Results
3.1. Experimental Design
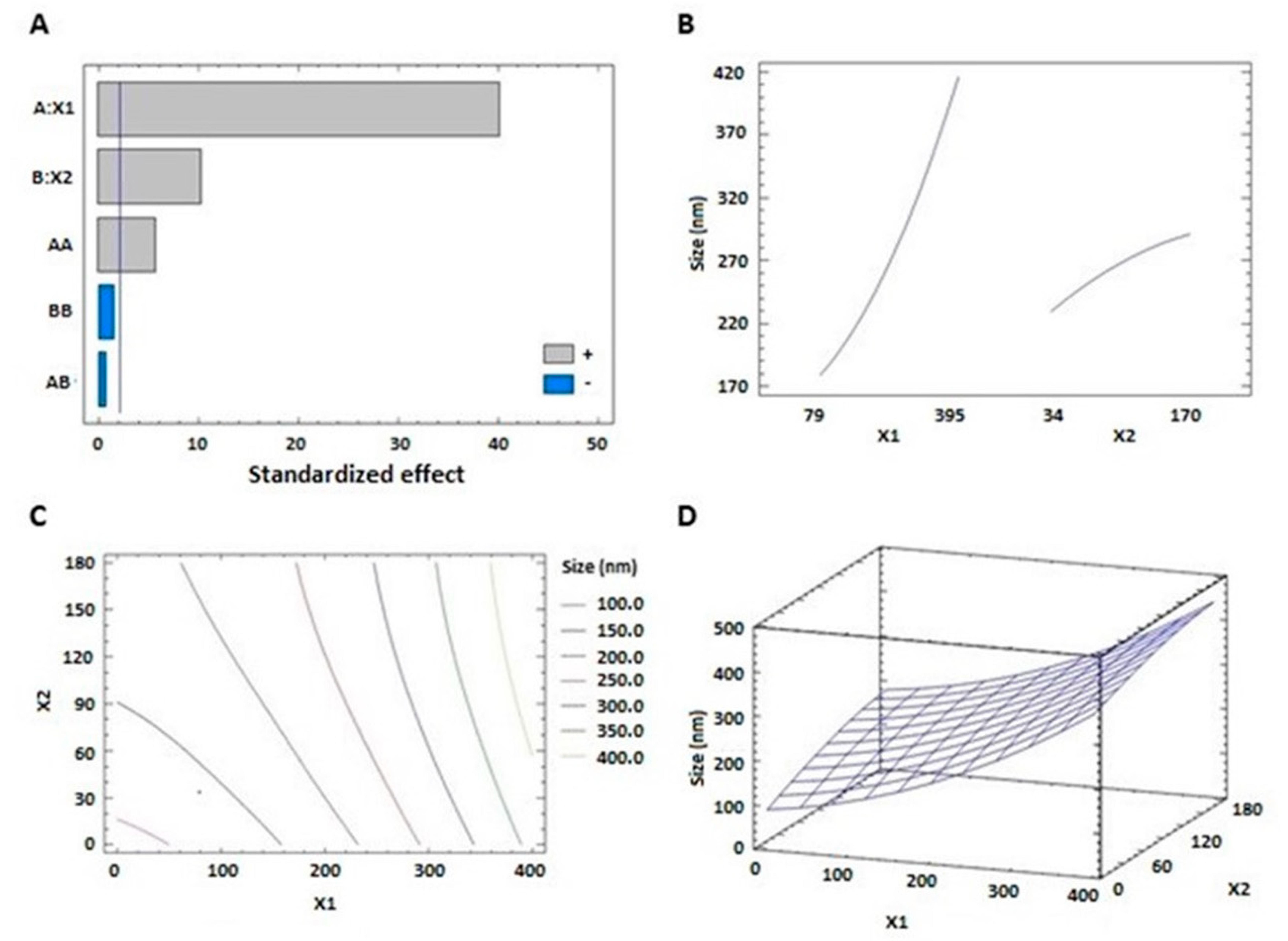
3.1.1. Effect of Variables on Vesicle Size (Y1)
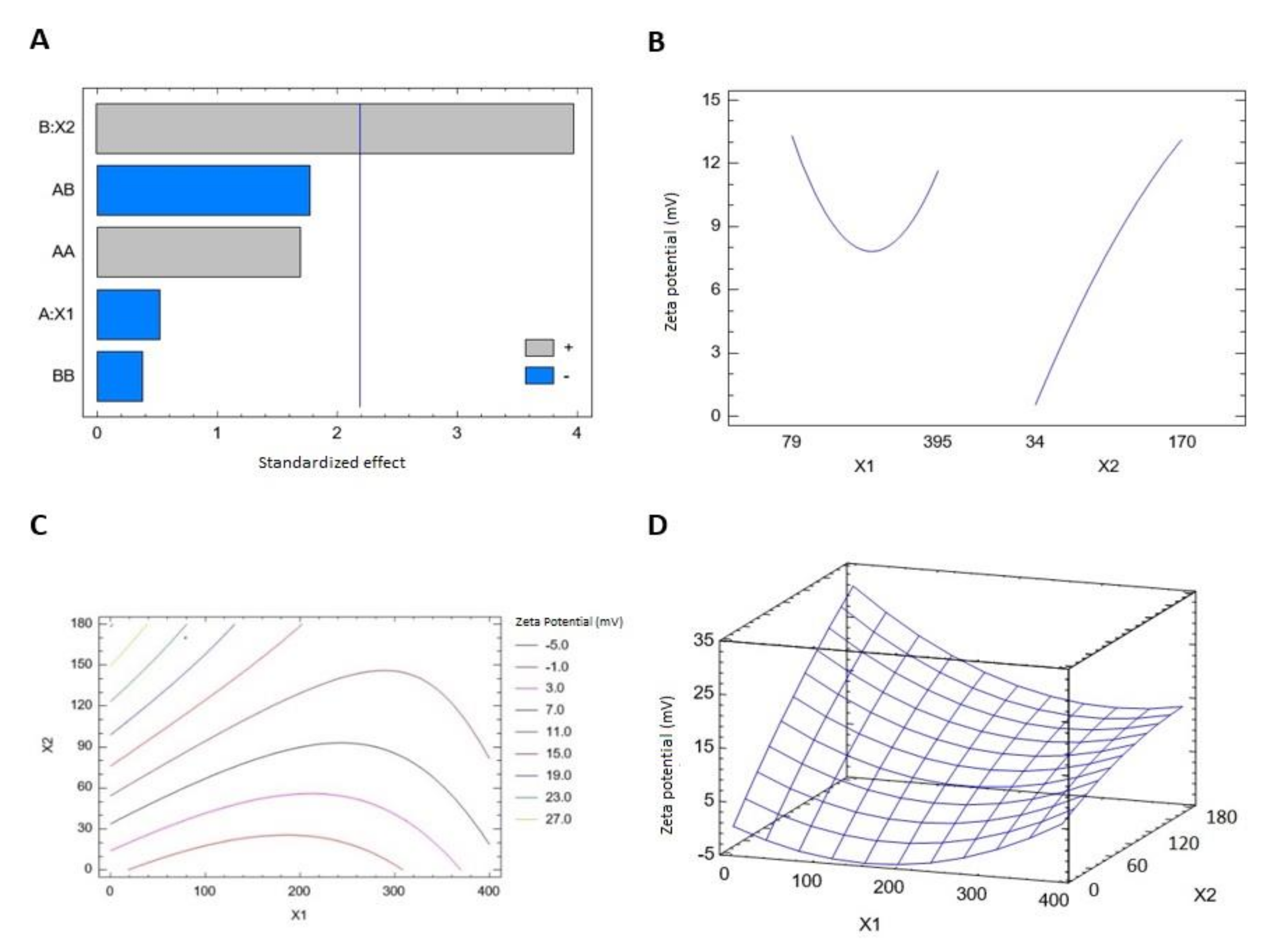
3.1.2. Effect of Variables on Zeta Potential (Y2)
3.1.3. Optimization of THQ–PL–SV Phytosome Formulation
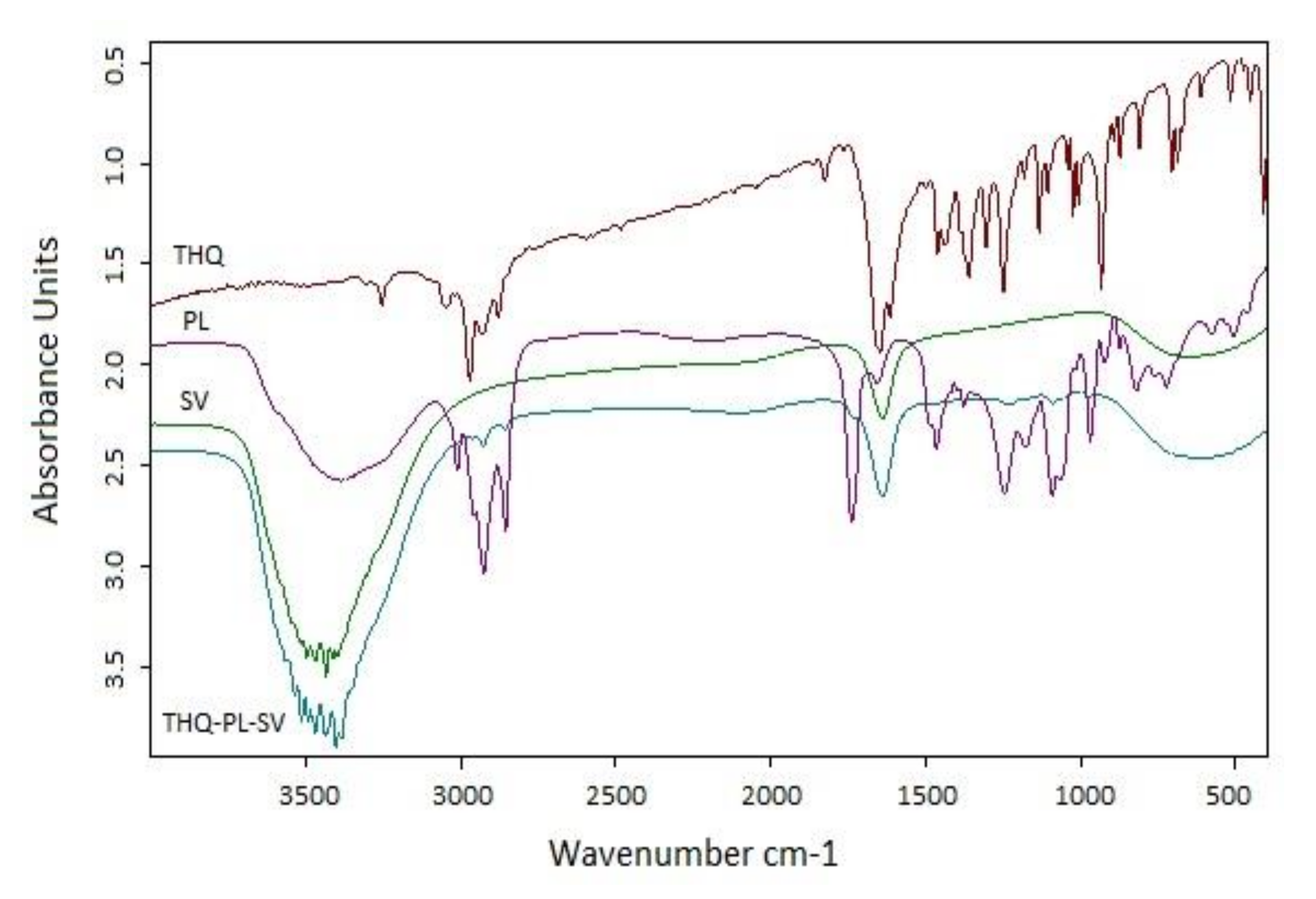
3.2. (FTIR) Characterization of the Optimized THQ–PL–SV Nanovesicles
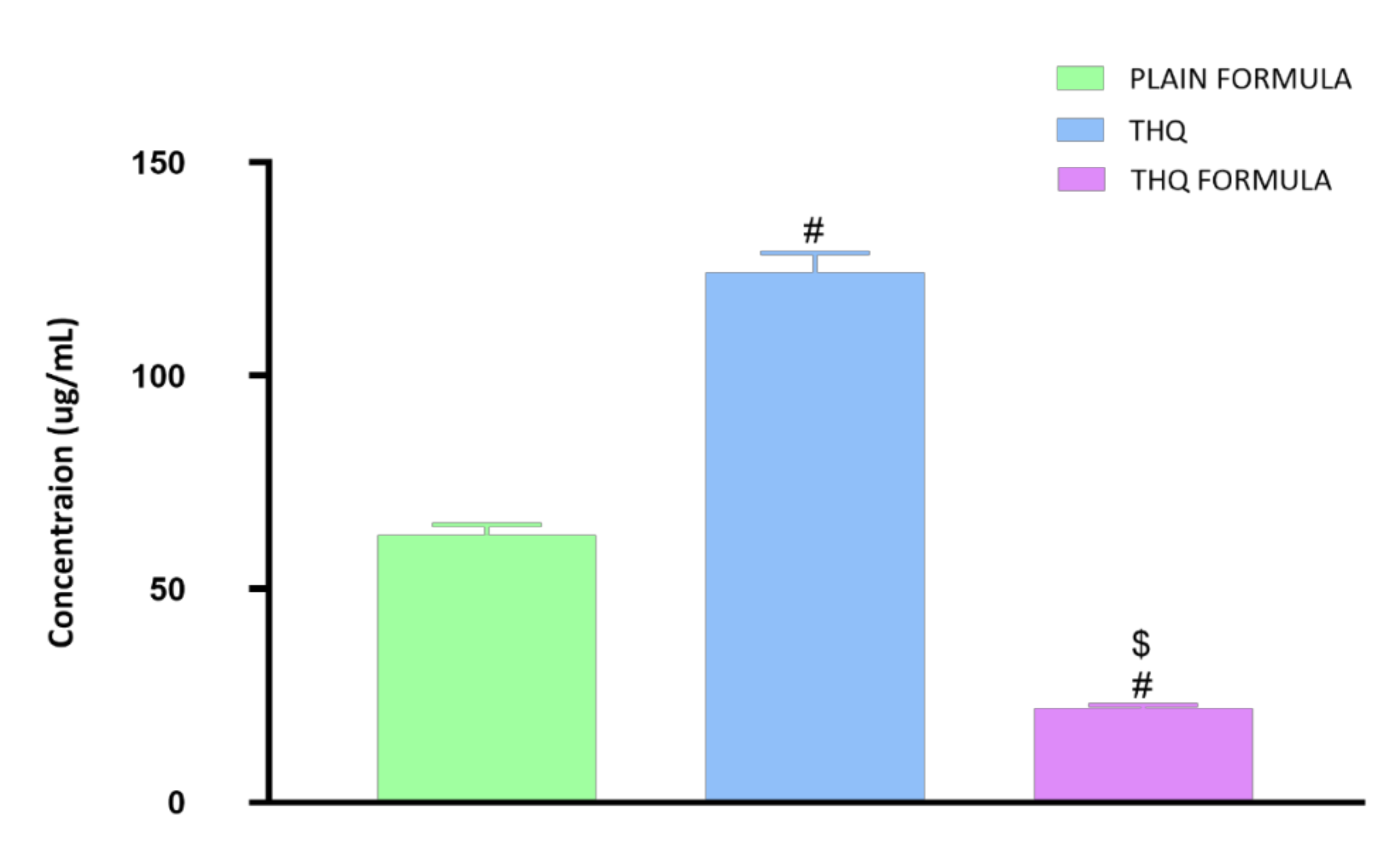
3.3. Determination of IC50 Values
3.4. Cell Cycle Analysis
3.5. Apoptotic Assessment Using Annexin V
3.6. Assessment of mRNA Expression of Caspase-3, Bax, Bcl-2, and P53
3.7. MMP Assessment
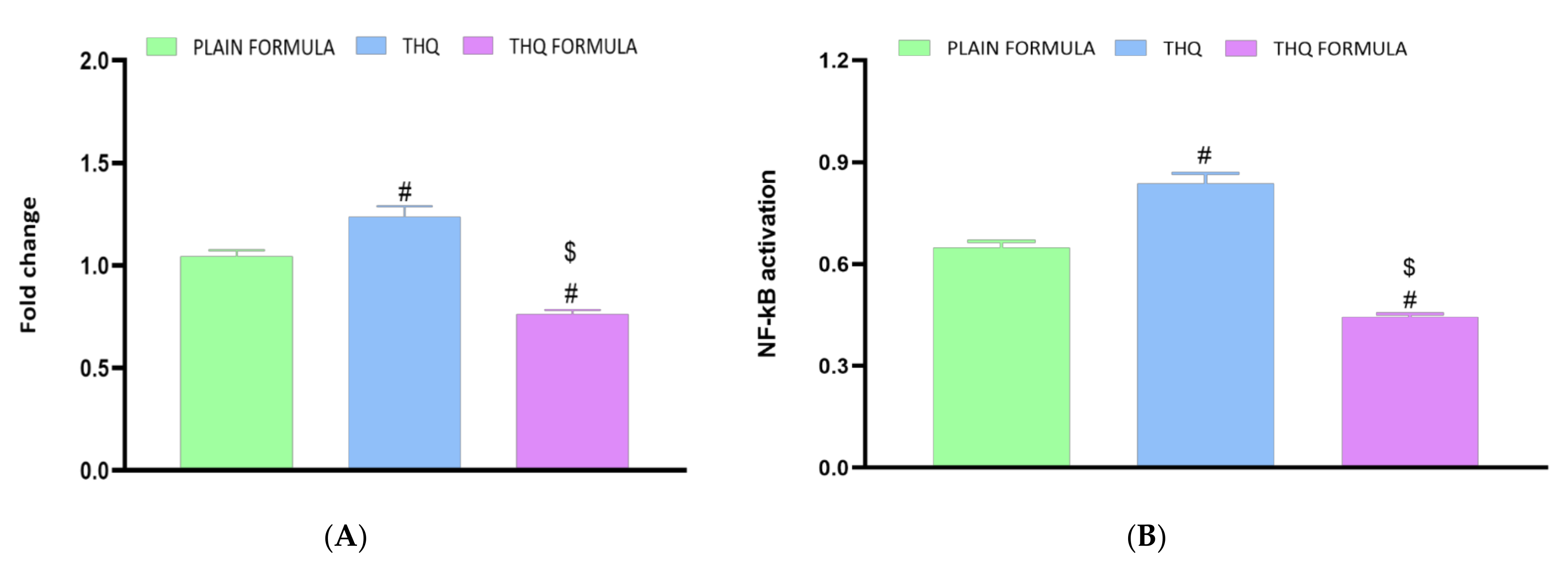
3.8. Assessment of Inflammatory Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Bernhardt, E.B.; Jalal, S.I. Small cell lung cancer. In Cancer Treatment and Research; Springer: New York, NY, USA, 2016; Volume 170, pp. 301–322. [Google Scholar]
- Li, C.; Lu, H. Adenosquamous carcinoma of the lung. Onco. Targets. Ther. 2018, 11, 4829–4835. [Google Scholar] [CrossRef] [Green Version]
- Collins, L.G.; Haines, C.; Perkel, R.; Enck, R.E. Lung Cancer: Diagnosis and Management. Am. Fam. Physician 2007, 75, 56–63. [Google Scholar] [PubMed]
- Daga, A.; Ansari, A.; Patel, S.; Mirza, S.; Rawal, R.; Umrania, V. Current drugs and drug targets in non-small cell lung cancer: Limitations and opportunities. Asian Pac. J. Cancer Prev. 2015, 16, 4147–4156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khader, M.; Eckl, P.M. Thymoquinone: An emerging natural drug with a wide range of medical applications. Iran. J. Basic Med. Sci. 2014, 17, 950–957. [Google Scholar] [PubMed]
- Badary, O.A.; Hamza, M.S.; Tikamdas, R. Thymoquinone: A promising natural compound with potential benefits for COVID-19 prevention and cure. Drug Des. Devel. Ther. 2021, 15, 1819–1833. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.N.; Prajapati, C.P.; Gore, P.R.; Patil, C.R.; Mahajan, U.B.; Sharma, C.; Talla, S.P.; Ojha, S.K. Therapeutic potential and pharmaceutical development of thymoquinone: A multitargeted molecule of natural origin. Front. Pharmacol. 2017, 8, 656. [Google Scholar] [CrossRef] [PubMed]
- Gurung, R.L.; Lim, S.N.; Khaw, A.K.; Soon, J.F.F.; Shenoy, K.; Ali, S.M.; Jayapal, M.; Sethu, S.; Baskar, R.; Prakash Hande, M. Thymoquinone induces telomere shortening, DNA damage and apoptosis in human glioblastoma cells. PLoS ONE 2010, 5, e12124. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.; Kumar, B.N.N.P.; Dey, K.K.; Pal, I.; Parekh, A.; Mandal, M. Molecular targeting of Akt by thymoquinone promotes G1 arrest through translation inhibition of cyclin D1 and induces apoptosis in breast cancer cells. Life Sci. 2013, 93, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Salim, L.Z.A.; Mohan, S.; Othman, R.; Abdelwahab, S.I.; Kamalidehghan, B.; Sheikh, B.Y.; Ibrahim, M.Y. Thymoquinone induces mitochondria-mediated apoptosis in acute lymphoblastic leukaemia in vitro. Molecules 2013, 18, 11219–11240. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, W.N.; Rosli, L.M.B.M.; Doolaanea, A.A. Formulation, cellular uptake and cytotoxicity of thymoquinone-loaded plga nanoparticles in malignant melanoma cancer cells. Int. J. Nanomed. 2020, 15, 8059–8074. [Google Scholar] [CrossRef] [PubMed]
- Alharbi, W.S.; Almughem, F.A.; Almehmady, A.M.; Jarallah, S.J.; Alsharif, W.K.; Alzahrani, N.M.; Alshehri, A.A. Phytosomes as an emerging nanotechnology platform for the topical delivery of bioactive phytochemicals. Pharmaceutics 2021, 13, 1475. [Google Scholar] [CrossRef] [PubMed]
- Saraf, S.; Khan, J.; Alexander, A.; Ajazuddin; Saraf, S. Recent advances and future prospects of phyto-phospholipid complexation technique for improving pharmacokinetic profile of plant actives. J. Control. Release 2013, 168, 50–60. [Google Scholar]
- Elmowafy, M.; Samy, A.; Raslan, M.A.; Salama, A.; Said, R.A.; Abdelaziz, A.E.; El-Eraky, W.; El Awdan, S.; Viitala, T. Enhancement of Bioavailability and Pharmacodynamic Effects of Thymoquinone Via Nanostructured Lipid Carrier (NLC) Formulation. AAPS PharmSciTech 2016, 17, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, H.; Xu, C.; Gu, L. A review: Using nanoparticles to enhance absorption and bioavailability of phenolic phytochemicals. Food Hydrocoll. 2015, 43, 153–164. [Google Scholar] [CrossRef]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Vadhanam, M.V. Bioavailability of phytochemicals and its enhancement by drug delivery systems. Cancer Lett. 2013, 334, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizvi, S.A.A.; Saleh, A.M. Applications of nanoparticle systems in drug delivery technology. Saudi Pharm. J. 2018, 26, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, A.; Sahebkar, A. Opinion Paper: Phytosome: A Fatty Solution for Efficient Formulation of Phytopharmaceuticals. Recent Pat. Drug Deliv. Formul. 2016, 10, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Batzri, S.; Korn, E.D. Interaction of phospholipid vesicles with cells: Endocytosis and fusion as alternate mechanisms for the uptake of lipid-soluble and water-soluble molecules. J. Cell Biol. 1975, 66, 621–634. [Google Scholar] [CrossRef]
- Yang, B.; Dong, Y.; Wang, F.; Zhang, Y. Nanoformulations to enhance the bioavailability and physiological functions of polyphenols. Molecules 2020, 25, 4613. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Knerr, J.M.; Argemi, L.; Bordon, K.C.F.; Pucca, M.B.; Cerni, F.A.; Arantes, E.C.; Çalişkan, F.; Laustsen, A.H. Scorpion venom: Detriments and benefits. Biomedicines 2020, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Solà, M.; Jappe, E.C.; Oscoz, S.; Lauridsen, L.P.; Engmark, M. Biotechnological trends in spider and scorpion antivenom development. Toxins 2016, 8, 226. [Google Scholar] [CrossRef] [Green Version]
- Hauke, T.J.; Herzig, V. Dangerous arachnids—Fake news or reality? Toxicon 2017, 138, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenço, W.R. The evolution and distribution of noxious species of scorpions (Arachnida: Scorpiones). J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petricevich, V.L. Scorpion venom and the inflammatory response. Mediat. Inflamm. 2010, 2010, 903295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, G.F. Venoms as a platform for human drugs: Translating toxins into therapeutics. Expert Opin. Biol. Ther. 2011, 11, 1469–1484. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide therapeutics from venom: Current status and potential. Bioorganic Med. Chem. 2018, 26, 2738–2758. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Vetter, I. Pharmacology and therapeutic potential of venom peptides. Biochem. Pharmacol. 2020, 181, 114207. [Google Scholar] [CrossRef] [PubMed]
- Rapôso, C. Scorpion and spider venoms in cancer treatment: State of the art, challenges, and perspectives. J. Clin. Transl. Res. 2017, 3, 233–249. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, L.; Chiara, P.; Antonio, C.; Calogero, F. Cancer pain managment with a venom of blue scorpion endemic in Cuba, called rhopalurus junceus “escozul”. Open Cancer J. 2012, 5, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Alhakamy, N.A.; Fahmy, U.A.; Badr-Eldin, S.M.; Ahmed, O.A.A.; Asfour, H.Z.; Aldawsari, H.M.; Algandaby, M.M.; Eid, B.G.; Abdel-Naim, A.B.; Awan, Z.A.; et al. Optimized icariin phytosomes exhibit enhanced cytotoxicity and apoptosis-inducing activities in ovarian cancer cells. Pharmaceutics 2020, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Mitić, Ž.; Stolić, A.; Stojanović, S.; Najman, S.; Ignjatović, N.; Nikolić, G.; Trajanović, M. Instrumental methods and techniques for structural and physicochemical characterization of biomaterials and bone tissue: A review. Mater. Sci. Eng. 2017, 79, 930–949. [Google Scholar] [CrossRef]
- Alhakamy, N.A.; Ahmed, O.A.A.; Fahmy, U.A.; Md, S. Development and in vitro evaluation of 2-methoxyestradiol loaded polymeric micelles for enhancing anticancer activities in prostate cancer. Polymers 2021, 13, 884. [Google Scholar] [CrossRef] [PubMed]
- Luna-Vital, D.A.; González de Mejía, E.; Loarca-Piña, G. Selective mechanism of action of dietary peptides from common bean on HCT116 human colorectal cancer cells through loss of mitochondrial membrane potential and DNA damage. J. Funct. Foods 2016, 23, 24–39. [Google Scholar] [CrossRef]
- Alhakamy, N.A.; Badr-Eldin, S.M.; Ahmed, O.A.A.; Asfour, H.Z.; Aldawsari, H.M.; Algandaby, M.M.; Eid, B.G.; Abdel-Naim, A.B.; Awan, Z.A.; Alghaith, A.F.; et al. Piceatannol-loaded emulsomes exhibit enhanced cytostatic and apoptotic activities in colon cancer cells. Antioxidants 2020, 9, 419. [Google Scholar] [CrossRef] [PubMed]
- Awan, Z.A.; Fahmy, U.A.; Badr-eldin, S.M.; Ibrahim, T.S.; Asfour, H.Z.; Al-rabia, M.W.; Alfarsi, A.; Alhakamy, N.A.; Abdulaal, W.H.; Sadoun, H.A.; et al. The enhanced cytotoxic and pro-apoptotic effects of optimized simvastatin-loaded emulsomes on MCF-7 breast cancer cells. Pharmaceutics 2020, 12, 597. [Google Scholar] [CrossRef] [PubMed]
- Ascenso, A.; Pedrosa, T.; Pinho, S.; Pinho, F.; de Oliveira, J.M.P.F.; Cabral Marques, H.; Oliveira, H.; Simões, S.; Santos, C. The Effect of Lycopene Preexposure on UV-B-Irradiated Human Keratinocytes. Oxid. Med. Cell. Longev. 2016, 2016, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Alhakamy, N.A.; Ahmed, O.A.A.; Fahmy, U.A.; Shadab, M. Apamin-conjugated alendronate sodium nanocomplex for management of pancreatic cancer. Pharmaceuticals 2021, 14, 729. [Google Scholar] [CrossRef]
- Hussain, S. Measurement of Nanoparticle-Induced Mitochondrial Membrane Potential Alterations. In Nanotoxicity. Methods in Molecular Biology; Zhang, Q., Ed.; Humana Press: New York, NY, USA, 2019; Volume 1894, pp. 123–131. [Google Scholar]
- Xue, Y.; Chen, Q.; Ding, T.; Sun, J. SiO₂ nanoparticle-induced impairment of mitochondrial energy metabolism in hepatocytes directly and through a Kupffer cell-mediated pathway in vitro. Int. J. Nanomed. 2014, 9, 2891–2903. [Google Scholar]
- Al-Mahallawi, A.M.; Abdelbary, A.A.; Aburahma, M.H. Investigating the potential of employing bilosomes as a novel vesicular carrier for transdermal delivery of tenoxicam. Int. J. Pharm. 2015, 485, 329–340. [Google Scholar] [CrossRef]
- Aldawsari, H.M.; Badr-Eldin, S.M. Enhanced pharmacokinetic performance of dapoxetine hydrochloride via the formulation of instantly-dissolving buccal films with acidic pH modifier and hydrophilic cyclodextrin: Factorial analysis, in vitro and in vivo assessment. J. Adv. Res. 2020, 24, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Shukla, P.; Misra, A.; Mishra, P.R. Interfacial and colloidal properties of emulsified systems: Pharmaceutical and biological perspective. In Colloid and Interface Science in Pharmaceutical Research and Development; Elsevier: Amsterdam, The Netherlands, 2014; pp. 149–172. ISBN 9780444626080. [Google Scholar]
- Yingchoncharoen, P.; Kalinowski, D.S.; Richardson, D.R. Lipid-based drug delivery systems in cancer therapy: What is available and what is yet to come. Pharmacol. Rev. 2016, 68, 701–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.R.; Lin, R.; Li, H.J.; He, W.L.; Du, J.Z.; Wang, J. Strategies to improve tumor penetration of nanomedicines through nanoparticle design. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badr-Eldin, S.M.; Aldawsari, H.M.; Ahmed, O.A.A.; Alhakamy, N.A.; Neamatallah, T.; Okbazghi, S.Z.; Fahmy, U.A. Optimized semisolid self-nanoemulsifying system based on glyceryl behenate: A potential nanoplatform for enhancing antitumor activity of raloxifene hydrochloride in MCF-7 human breast cancer cells. Int. J. Pharm. 2021, 600, 120493. [Google Scholar] [CrossRef] [PubMed]
- Saoji, S.D.; Raut, N.A.; Dhore, P.W.; Borkar, C.D.; Popielarczyk, M.; Dave, V.S. Preparation and Evaluation of Phospholipid-Based Complex of Standardized Centella Extract (SCE) for the Enhanced Delivery of Phytoconstituents. AAPS J. 2016, 18, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, O.A.A.; Badr-Eldin, S.M. Development of an optimized avanafil-loaded invasomal transdermal film: Ex vivo skin permeation and in vivo evaluation. Int. J. Pharm. 2019, 570, 118657. [Google Scholar] [CrossRef] [PubMed]
- Dubey, V.; Mishra, D.; Dutta, T.; Nahar, M.; Saraf, D.K.; Jain, N.K. Dermal and transdermal delivery of an anti-psoriatic agent via ethanolic liposomes. J. Control. Release 2007, 123, 148–154. [Google Scholar] [CrossRef]
- Narayan, R.; Singh, M.; Ranjan, O.P.; Nayak, Y.; Garg, S.; Shavi, G.V.; Nayak, U.Y. Development of risperidone liposomes for brain targeting through intranasal route. Life Sci. 2016, 163, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Mehanna, M.M.; El-Kader, N.A.; Samaha, M.W. Liposomes as potential carriers for ketorolac ophthalmic delivery: Formulation and stability issues. Braz. J. Pharm. Sci. 2017, 53, 1–10. [Google Scholar] [CrossRef]
- Krasnici, S.; Werner, A.; Eichhorn, M.E.; Schmitt-Sody, M.; Pahernik, S.A.; Sauer, B.; Schulze, B.; Teifel, M.; Michaelis, U.; Naujoks, K.; et al. Effect of the surface charge of liposomes on their uptake by angiogenic tumor vessels. Int. J. Cancer 2003, 105, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Zuo, Z.Q.; Du, J.Z.; Wang, Y.C.; Sun, R.; Cao, Z.T.; Ye, X.D.; Wang, J.L.; Leong, K.W.; Wang, J. Surface charge critically affects tumor penetration and therapeutic efficacy of cancer nanomedicines. Nano Today 2016, 11, 133–144. [Google Scholar] [CrossRef]
- Saadat, M.; Zahednezhad, F.; Zakeri-Milani, P.; Heidari, H.R.; Shahbazi-Mojarrad, J.; Valizadeh, H. Drug targeting strategies based on charge dependent uptake of nanoparticles into cancer cells. J. Pharm. Pharm. Sci. 2019, 22, 191–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Yang, F.; Li, F.; Li, Z.; Lang, Y.; Shen, B.; Wu, Y.; Li, W.; Harrison, P.L.; Strong, P.N.; et al. Therapeutic potential of a scorpion venom-derived antimicrobial peptide and its homologs against antibiotic-resistant Gram-positive bacteria. Front. Microbiol. 2018, 9, 1159. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion venom components as potential candidates for drug development. Toxicon 2015, 93, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmari, A.K.; Riyasdeen, A.; Islam, M. Scorpion Venom Causes Apoptosis by Increasing Reactive Oxygen Species and Cell Cycle Arrest in MDA-MB-231 and HCT-8 Cancer Cell Lines. J. Evid. -Based Integr. Med. 2018, 23, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, S.; Padhye, S.; Azmi, A.; Wang, Z.; Philip, P.A.; Kucuk, O.; Sarkar, F.H.; Mohammad, R.M. Review on molecular and therapeutic potential of thymoquinone in cancer. Nutr. Cancer 2010, 62, 938–946. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, L.; Mollica, M.; Re, A.T.; Wu, S.; Zuo, L. Nitric oxide in cancer metastasis. Cancer Lett. 2014, 353, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahmy, U.A.; Badr-Eldin, S.M.; Ahmed, O.A.A.; Aldawsari, H.M.; Tima, S.; Asfour, H.Z.; Al-Rabia, M.W.; Negm, A.A.; Sultan, M.H.; Madkhali, O.A.A.; et al. Intranasal niosomal in situ gel as a promising approach for enhancing flibanserin bioavailability and brain delivery: In vitro optimization and ex vivo/in vivo evaluation. Pharmaceutics 2020, 12, 485. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, O.A.A.; Fahmy, U.A.; Bakhaidar, R.; El-Moselhy, M.A.; Okbazghi, S.Z.; Ahmed, A.S.F.; Hammad, A.S.A.; Alhakamy, N.A. Omega-3 self-nanoemulsion role in gastroprotection against indomethacin-induced gastric injury in rats. Pharmaceutics 2020, 12, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T. Thymoquinone-induced antitumor and apoptosis in human lung adenocarcinoma cells. J. Cell. Physiol. 2019, 234, 10421–10431. [Google Scholar] [CrossRef] [PubMed]
- Wirries, A.; Breyer, S.; Quint, K.; Schobert, R.; Ocker, M. Thymoquinone hydrazone derivatives cause cell cycle arrest in p53-competent colorectal cancer cells. Exp. Ther. Med. 2010, 1, 369–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czarnomysy, R.; Surażyński, A.; Muszynska, A.; Gornowicz, A.; Bielawska, A.; Bielawski, K. A novel series of pyrazole-platinum(II) complexes as potential anti-cancer agents that induce cell cycle arrest and apoptosis in breast cancer cells. J. Enzym. Inhib. Med. Chem. 2018, 33, 1006–1023. [Google Scholar] [CrossRef]
- Park, E.J.; Chauhan, A.K.; Min, K.J.; Park, D.C.; Kwon, T.K. Thymoquinone induces apoptosis through downregulation of c-FLIP and Bcl-2 in Renal carcinoma Caki cells. Oncol. Rep. 2016, 36, 2261–2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gali-Muhtasib, H.U.; Abou Kheir, W.G.; Kheir, L.A.; Darwiche, N.; Crooks, P.A. Molecular pathway for thymoquinone-induced cell-cycle arrest and apoptosis in neoplastic keratinocytes. Anticancer. Drugs 2004, 15, 389–399. [Google Scholar] [CrossRef]
- Alhakamy, N.A.; Badr-Eldin, S.M.; Fahmy, U.A.; Alruwaili, N.K.; Awan, Z.A.; Caruso, G.; Alfaleh, M.A.; Alaofi, A.L.; Arif, F.O.; Ahmed, O.A.A.; et al. Thymoquinone-loaded soy-phospholipid-based phytosomes exhibit anticancer potential against human lung cancer cells. Pharmaceutics 2020, 12, 761. [Google Scholar] [CrossRef] [PubMed]
- Naseri, M.H.; Mahdavi, M.; Davoodi, J.; Tackallou, H.S.; Goudarzvand, M.; Neishabouri, S.H. Up regulation of Bax and down regulation of Bcl2 during 3-NC mediated apoptosis in human cancer cells. Cancer Cell Int. 2015, 15, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, V.C.L.; Cash, H.L.; Morse, J.L.; Lu, S.; Zhitkovich, A. S-phase sensing of DNA-protein crosslinks triggers TopBP1-independent ATR activation and p53-mediated cell death by formaldehyde. Cell Cycle 2012, 11, 2526–2537. [Google Scholar] [CrossRef]
- Wang, B.; Xiao, Z.; Ko, H.L.; Ren, E.C. The p53 response element and transcriptional repression. Cell Cycle 2010, 9, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Costantini, P.; Jacotot, E.; Decaudin, D.; Kroemer, G. Mitochondrion as a Novel Target of Anticancer Chemotherapy. JNCI J. Natl. Cancer Inst. 2000, 92, 1042–1053. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of reactive oxygen species in cancer progression: Molecular mechanisms and recent advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szondy, Z.; Sarang, Z.; Kiss, B.; Garabuczi, É.; Köröskényi, K. Anti-inflammatory mechanisms triggered by apoptotic cells during their clearance. Front. Immunol. 2017, 8, 909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cvetanovic, M.; Ucker, D.S. Innate Immune Discrimination of Apoptotic Cells: Repression of Proinflammatory Macrophage Transcription Is Coupled Directly to Specific Recognition. J. Immunol. 2004, 172, 880–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | Levels | ||
|---|---|---|---|
| (−1) | (0) | (+1) | |
| X1: PL concentration (mg) | 79 | 237 | 395 |
| X2: SV concentration (mg) | 34 | 102 | 170 |
| Responses | Desirability constraint | ||
| Y1: Particle size (nm) | Minimize | ||
| Y2: Zeta potential (mV) | Maximize | ||
| Run | X1 | X2 | Y1 | Y2 |
|---|---|---|---|---|
| PL Concentration | SV Concentration | Vesicle Size * ± SD | Zeta Potential * ± SD | |
| (mg) | (mg) | (nm) | (mV) | |
| 1 | 79.0 | 170.0 | 198.8 ± 9.8 | 26.8 ± 7.4 |
| 2 | 395.0 | 34.0 | 388.0 ± 13.6 | 2.7 ± 4.1 |
| 3 | 395.0 | 102.0 | 412.4 ± 24.1 | 17.7 ± 8.7 |
| 4 | 395.0 | 170.0 | 435.6 ± 19.3 | 11.8 ± 4.6 |
| 5 | 79.0 | 34.0 | 143.7 ± 14.6 | 4.4 ± 3.9 |
| 6 | 79.0 | 102.0 | 176.6 ± 17.9 | 6.7 ± 3.7 |
| 7 | 237.0 | 34.0 | 212.5 ± 13.5 | 3.8 ± 4.7 |
| 8 | 237.0 | 170.0 | 298.8 ± 23.3 | 9.8 ± 5.2 |
| 9 | 237.0 | 102.0 | 274.5 ± 19.9 | 7.9 ± 6.7 |
| 10 | 79.0 | 170.0 | 199.7 ± 21.1 | 27.1 ± 5.4 |
| 11 | 395.0 | 34.0 | 389.7 ± 26.4 | 2.9 ± 7.3 |
| 12 | 395.0 | 102.0 | 410.3 ± 28.7 | 18.9 ± 5.7 |
| 13 | 395.0 | 170.0 | 430.5 ± 23.1 | 11.8 ± 3.4 |
| 14 | 79.0 | 34.0 | 145.8 ± 12.5 | 4.1 ± 5.3 |
| 15 | 79.0 | 102.0 | 174.6 ± 23.8 | 6.6 ± 4.3 |
| 16 | 237.0 | 34.0 | 214.7 ± 18.9 | 3.8 ± 4.7 |
| 17 | 237.0 | 170.0 | 298.4 ± 22.2 | 9.9 ± 5.1 |
| 18 | 237.0 | 102.0 | 276.8 ± 24.3 | 7.8 ± 3.6 |
| Responses | R2 (%) | Adjusted R2 (%) | Standard Error of Est. | PRESS | Significant Terms |
|---|---|---|---|---|---|
| Y1: Vesicle size (nm) | 99.3799 | 99.1216 | 10.2385 | 102.10 | X1, X2, |
| Y2: Zeta potential (mV) | 66.8896 | 53.0935 | 5.47816 | 3.46 | X2 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| A:X1 | 169782 | 1 | 169782. | 1619.63 | <0.0001 |
| B:X2 | 11243.7 | 1 | 11243.7 | 107.26 | <0.0001 |
| AA | 3485.13 | 1 | 3485.13 | 33.25 | 0.0001 |
| AB | 52.8392 | 1 | 52.8392 | 0.50 | 0.4925 |
| BB | 246.804 | 1 | 246.804 | 2.35 | 0.1532 |
| blocks | 0.0102722 | 1 | 0.0102722 | 0.00 | 0.9923 |
| Total error | 1153.1 | 11 | 104.828 | ||
| Total (corr.) | 185964. | 17 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| A:X1 | 8.1675 | 1 | 8.1675 | 0.27 | 0.6122 |
| B:X2 | 475.021 | 1 | 475.021 | 15.83 | 0.0022 |
| AA | 85.5625 | 1 | 85.5625 | 2.85 | 0.1194 |
| AB | 93.845 | 1 | 93.845 | 3.13 | 0.1047 |
| BB | 4.2025 | 1 | 4.2025 | 0.14 | 0.7154 |
| blocks | 0.0938889 | 1 | 0.0938889 | 0.00 | 0.9564 |
| Total error | 330.113 | 11 | 30.0103 | ||
| Total (corr.) | 997.005 | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asfour, H.Z.; Fahmy, U.A.; Alharbi, W.S.; Almehmady, A.M.; Alamoudi, A.J.; Tima, S.; Mansouri, R.A.; Omar, U.M.; Ahmed, O.A.A.; Zakai, S.A.; et al. Phyto-Phospholipid Conjugated Scorpion Venom Nanovesicles as Promising Carrier That Improves Efficacy of Thymoquinone against Adenocarcinoma Human Alveolar Basal Epithelial Cells. Pharmaceutics 2021, 13, 2144. https://doi.org/10.3390/pharmaceutics13122144
Asfour HZ, Fahmy UA, Alharbi WS, Almehmady AM, Alamoudi AJ, Tima S, Mansouri RA, Omar UM, Ahmed OAA, Zakai SA, et al. Phyto-Phospholipid Conjugated Scorpion Venom Nanovesicles as Promising Carrier That Improves Efficacy of Thymoquinone against Adenocarcinoma Human Alveolar Basal Epithelial Cells. Pharmaceutics. 2021; 13(12):2144. https://doi.org/10.3390/pharmaceutics13122144
Chicago/Turabian StyleAsfour, Hani Z., Usama A. Fahmy, Waleed S. Alharbi, Alshaimaa M. Almehmady, Abdulmohsin J. Alamoudi, Singkome Tima, Rasha A. Mansouri, Ulfat M. Omar, Osama A. A. Ahmed, Shadi A. Zakai, and et al. 2021. "Phyto-Phospholipid Conjugated Scorpion Venom Nanovesicles as Promising Carrier That Improves Efficacy of Thymoquinone against Adenocarcinoma Human Alveolar Basal Epithelial Cells" Pharmaceutics 13, no. 12: 2144. https://doi.org/10.3390/pharmaceutics13122144