
Novel Fluorinated Spermine and Small Molecule PEI to Deliver Anti-PD-L1 and Anti-VEGF siRNA for Highly Efficient Tumor Therapy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Cell Culture
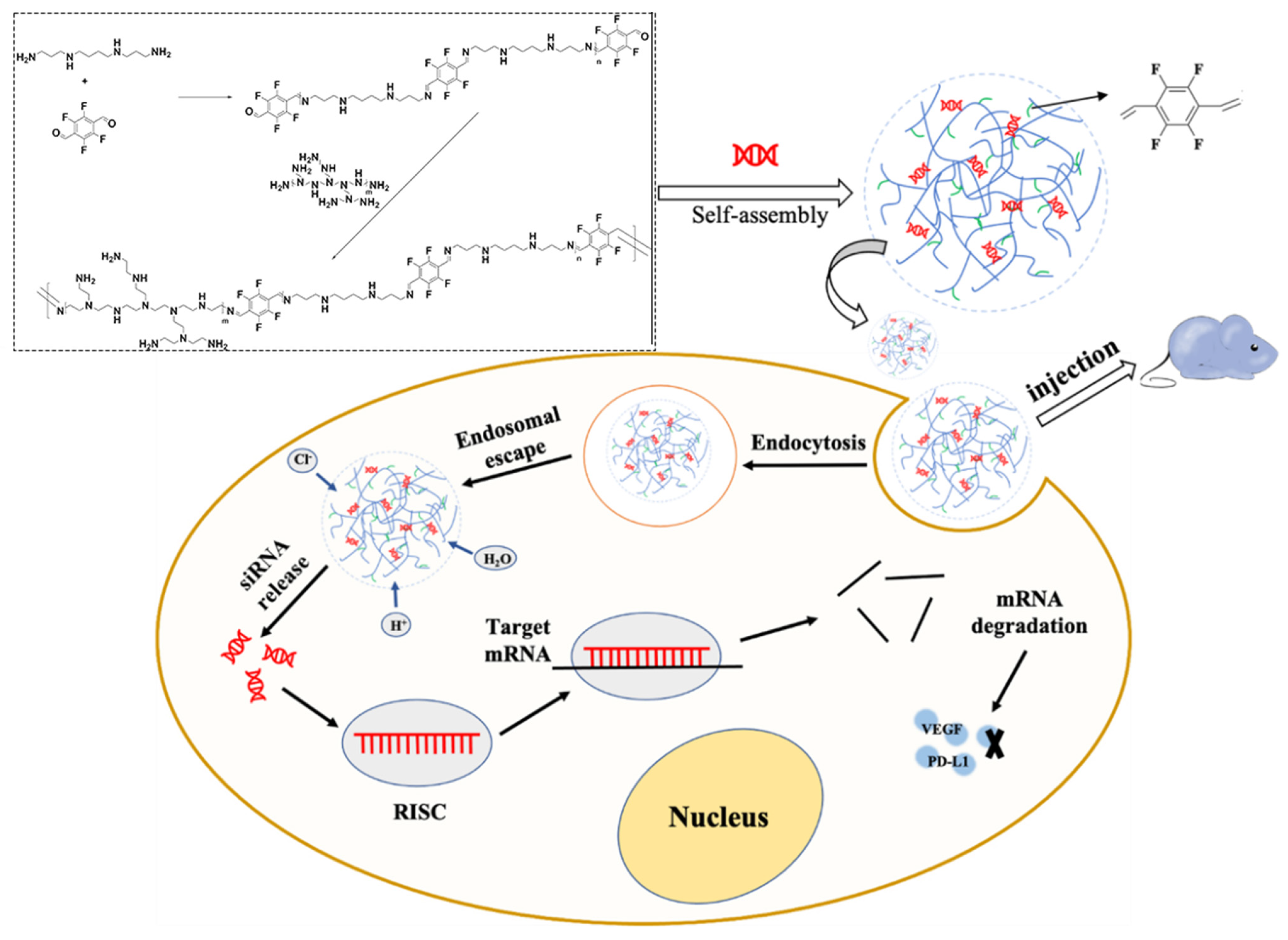
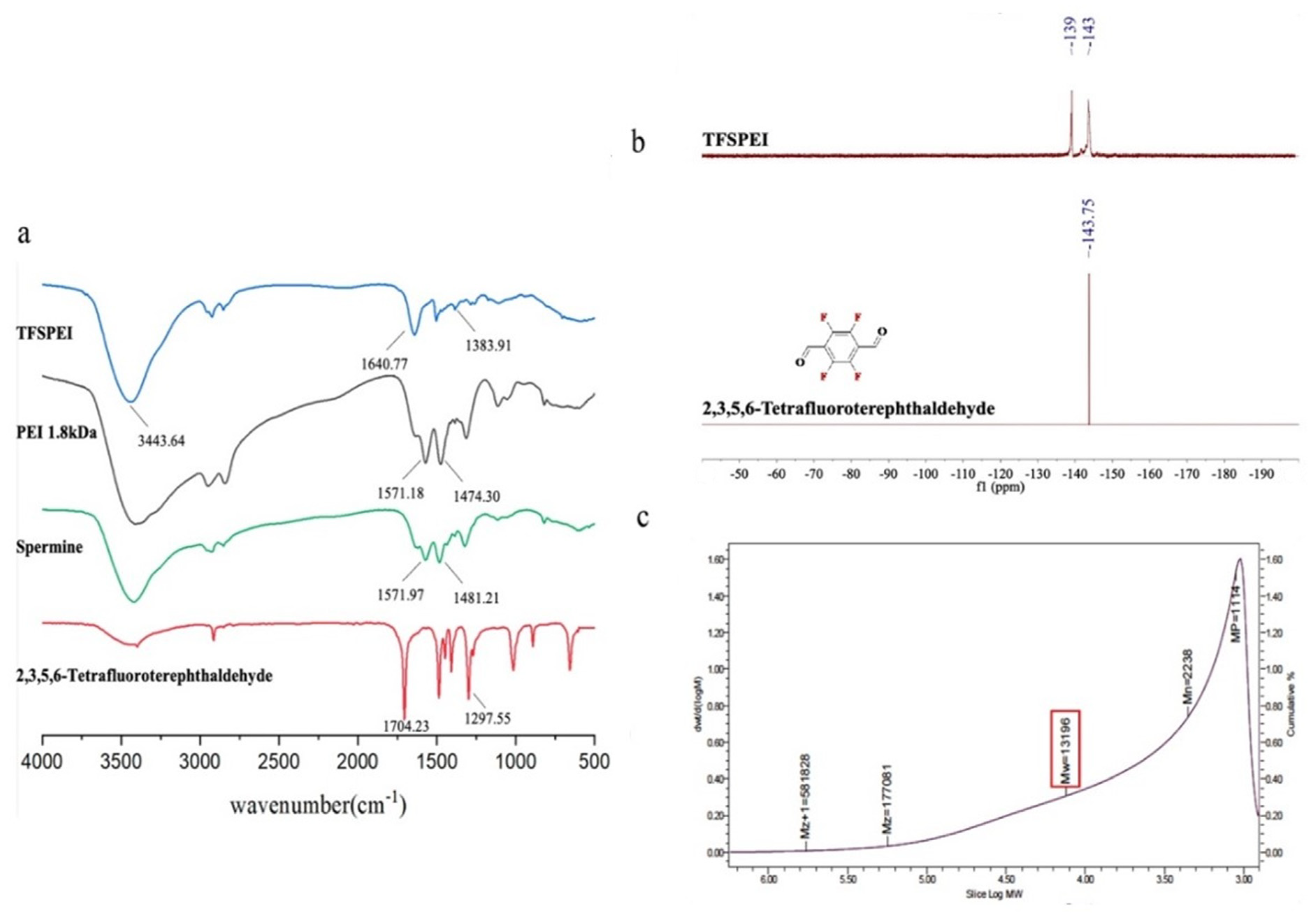
2.2.2. Synthesis and Characterization of TFSPEI
2.2.3. Preparation of TFSPEI Polyplex
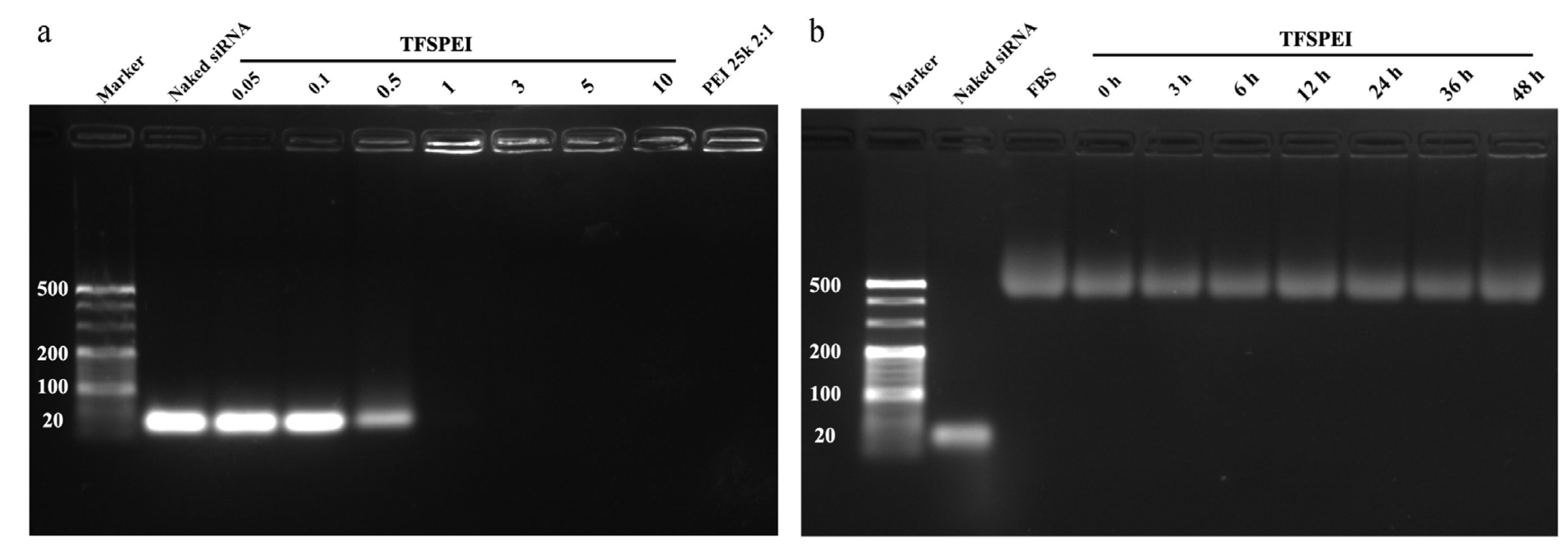
2.2.4. Agarose Gel Electrophoresis
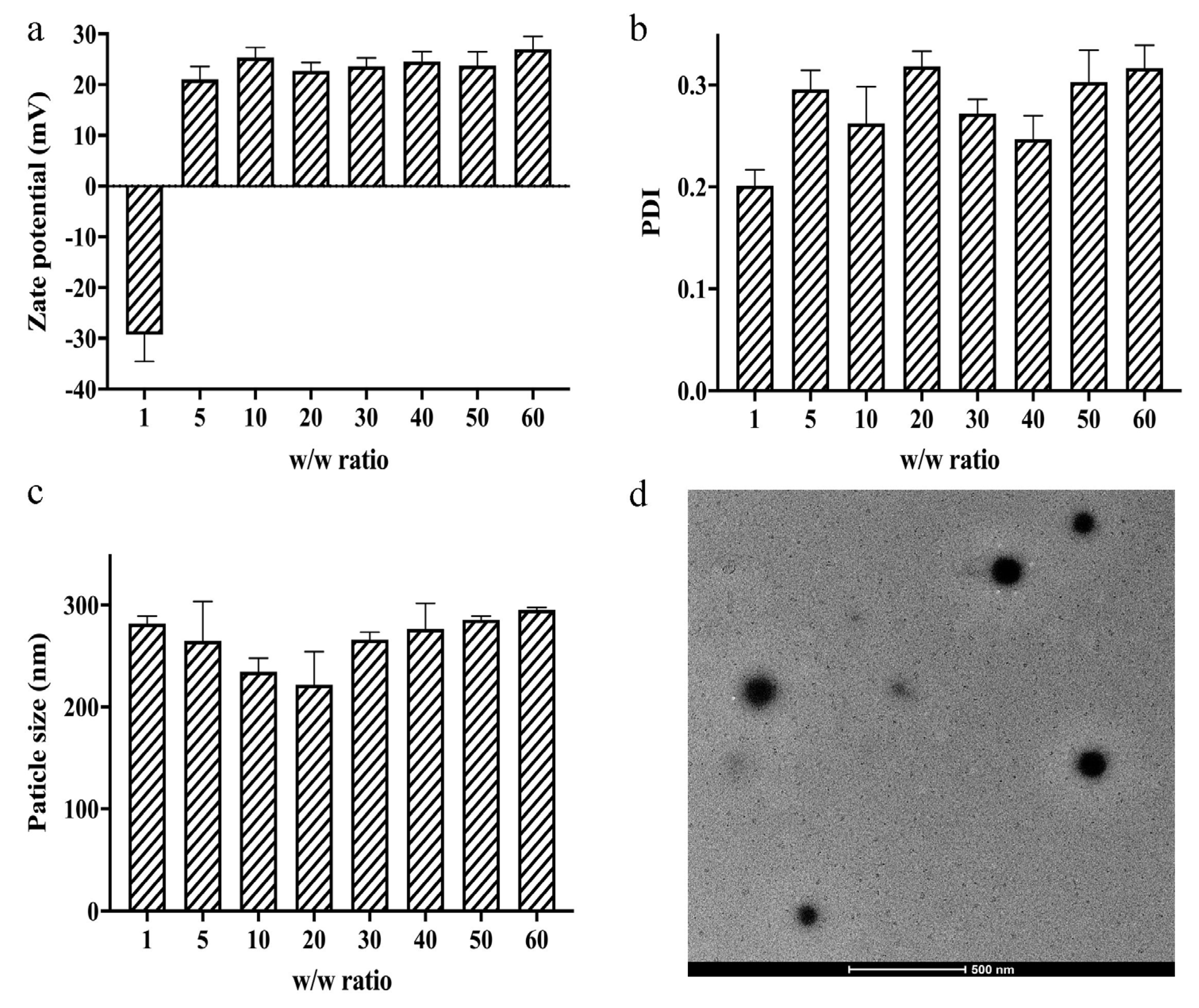
2.2.5. Particle size, Zeta Potential, and Morphology Measurements
2.2.6. In Vitro Cytotoxicity
2.2.7. In Vitro Endocytosis Efficiency
2.2.8. Intracellular Uptake
2.2.9. In Vitro Gene Silencing Efficiency
2.2.10. In Vivo Anti-Tumor Treatment
2.2.11. In Vivo Anti-Tumor Effects
2.2.12. In Vivo Cytotoxicity
2.2.13. Statistical Analysis
3. Results and Discussion
3.1. Characterization of TFSPEI
3.2. AGE and Serum Stability
3.3. Zeta Potential, Particle Size, and Morphology Measurements
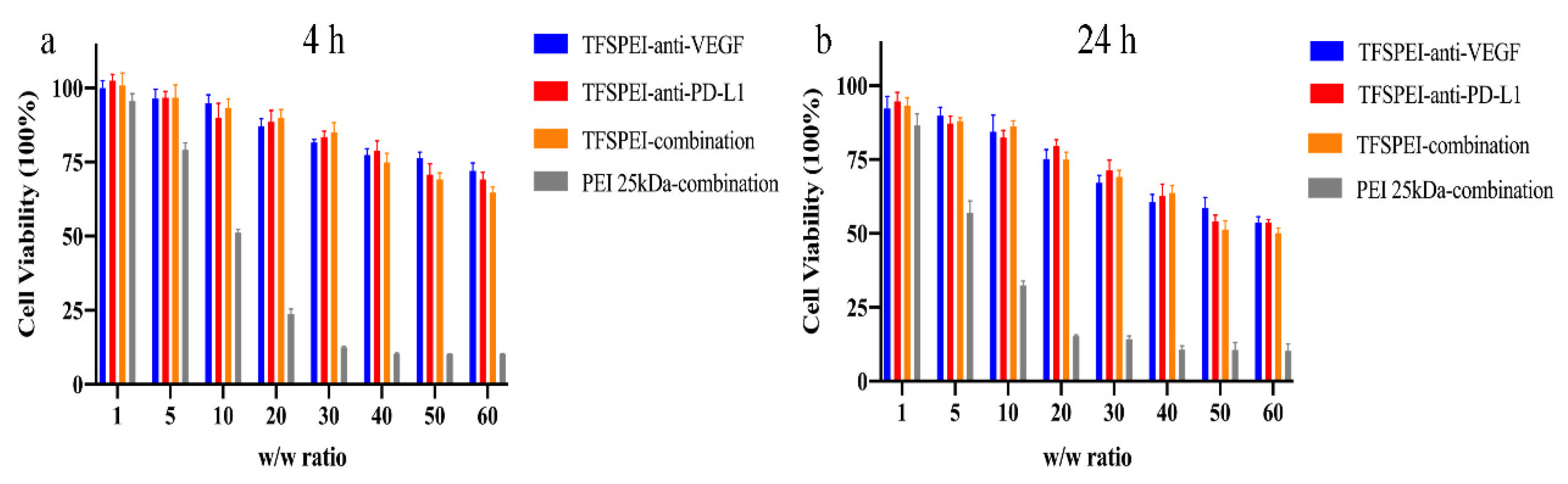
3.4. In Vitro Cytotoxicity
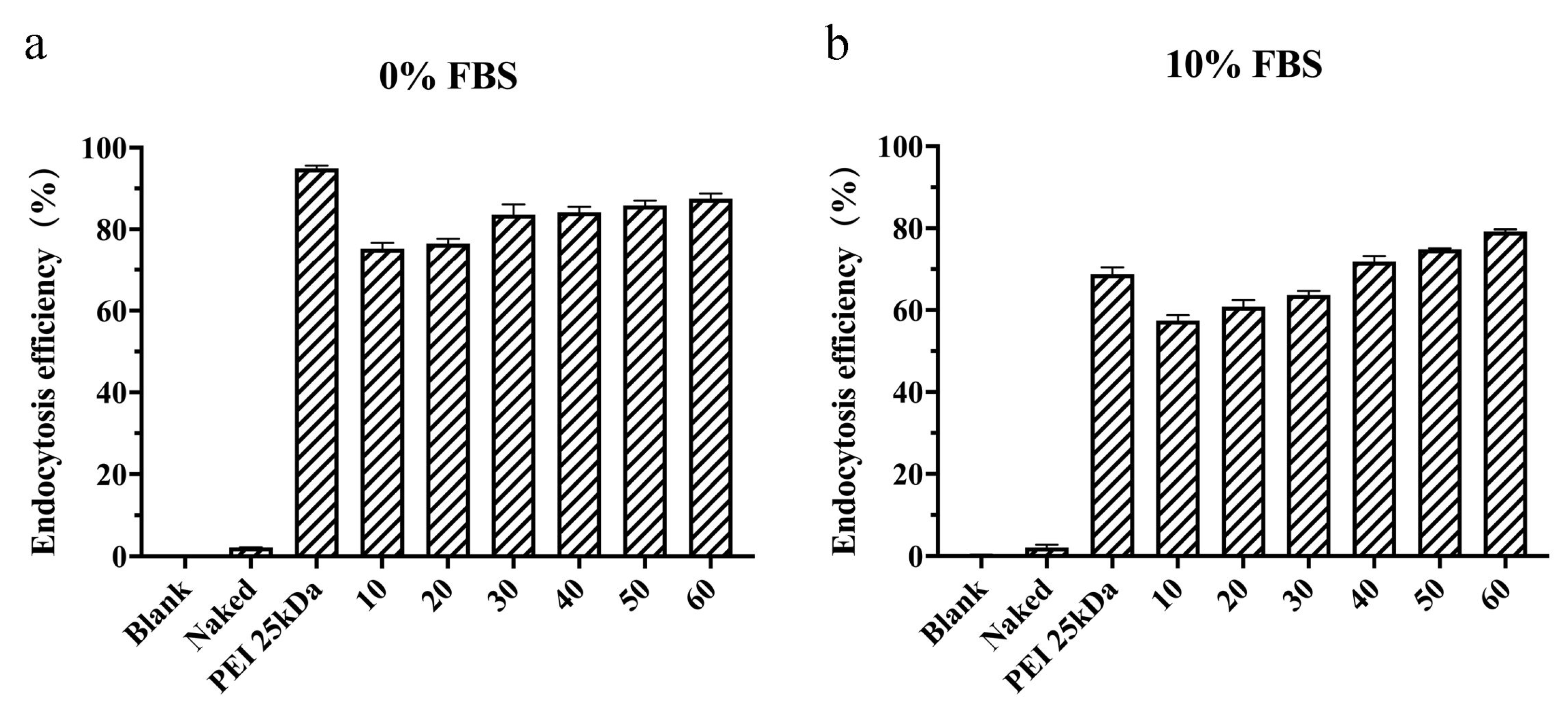
3.5. In Vitro Endocytosis Efficiency
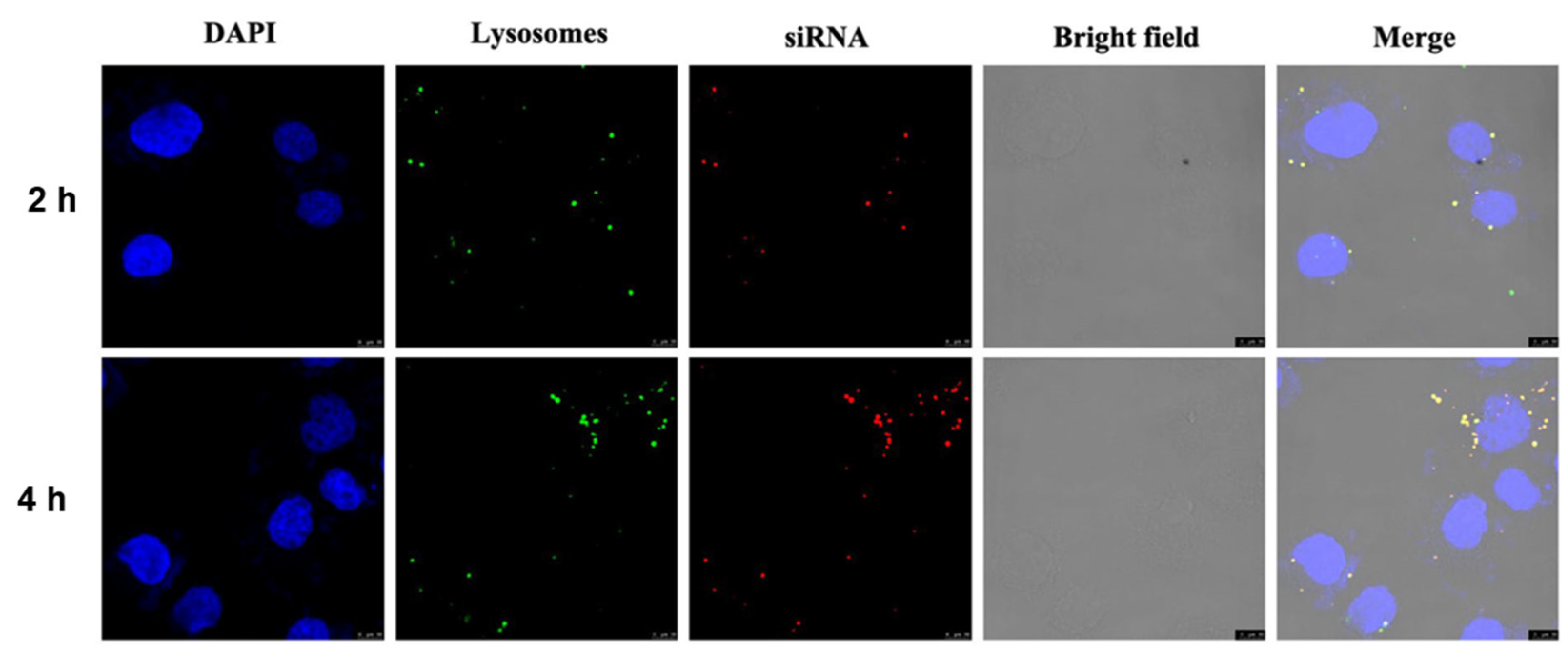
3.6. Intracellular Uptake
3.7. In Vitro Gene Silencing Efficiency
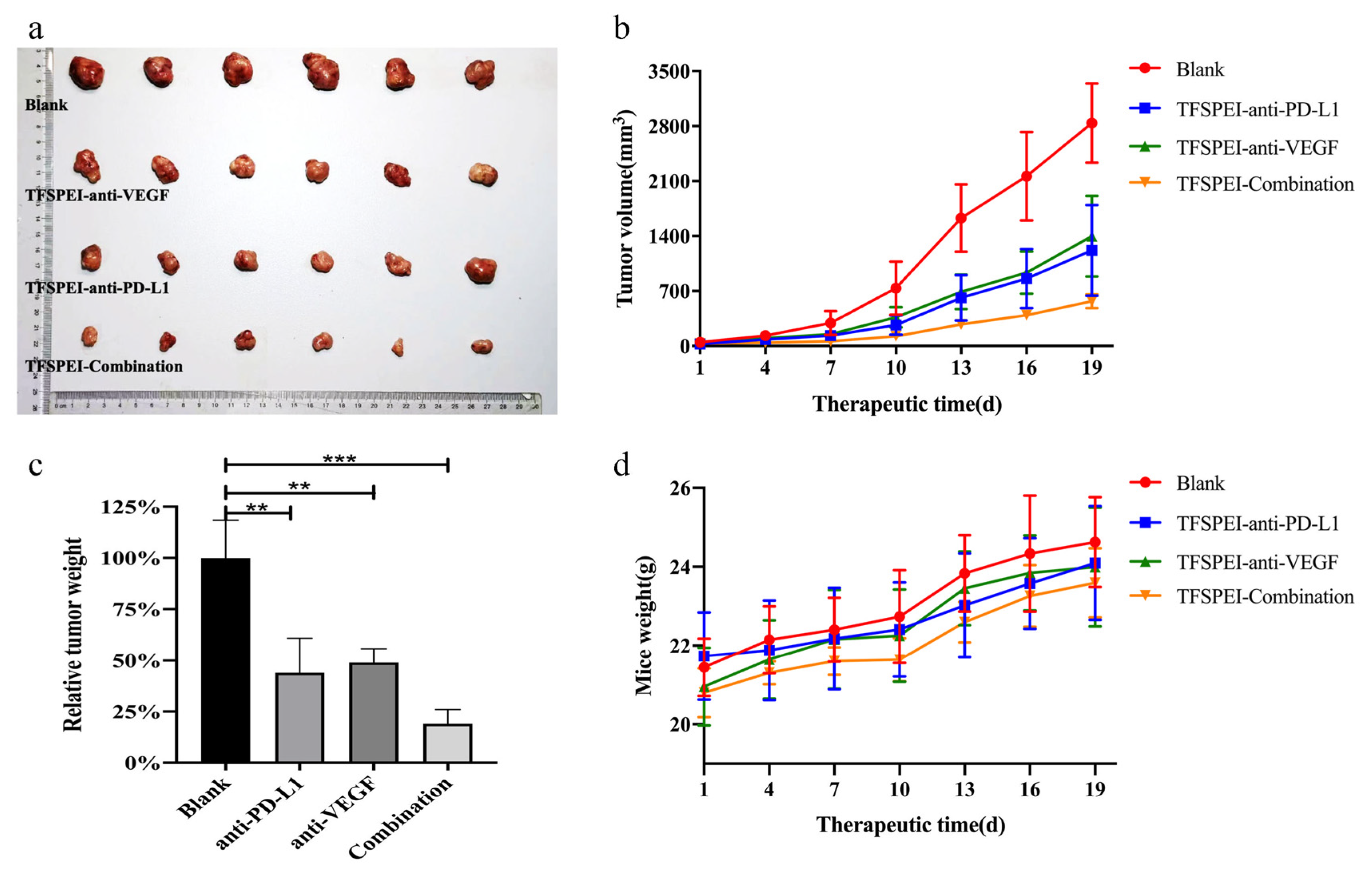
3.8. In Vivo Anti-Tumor Effects
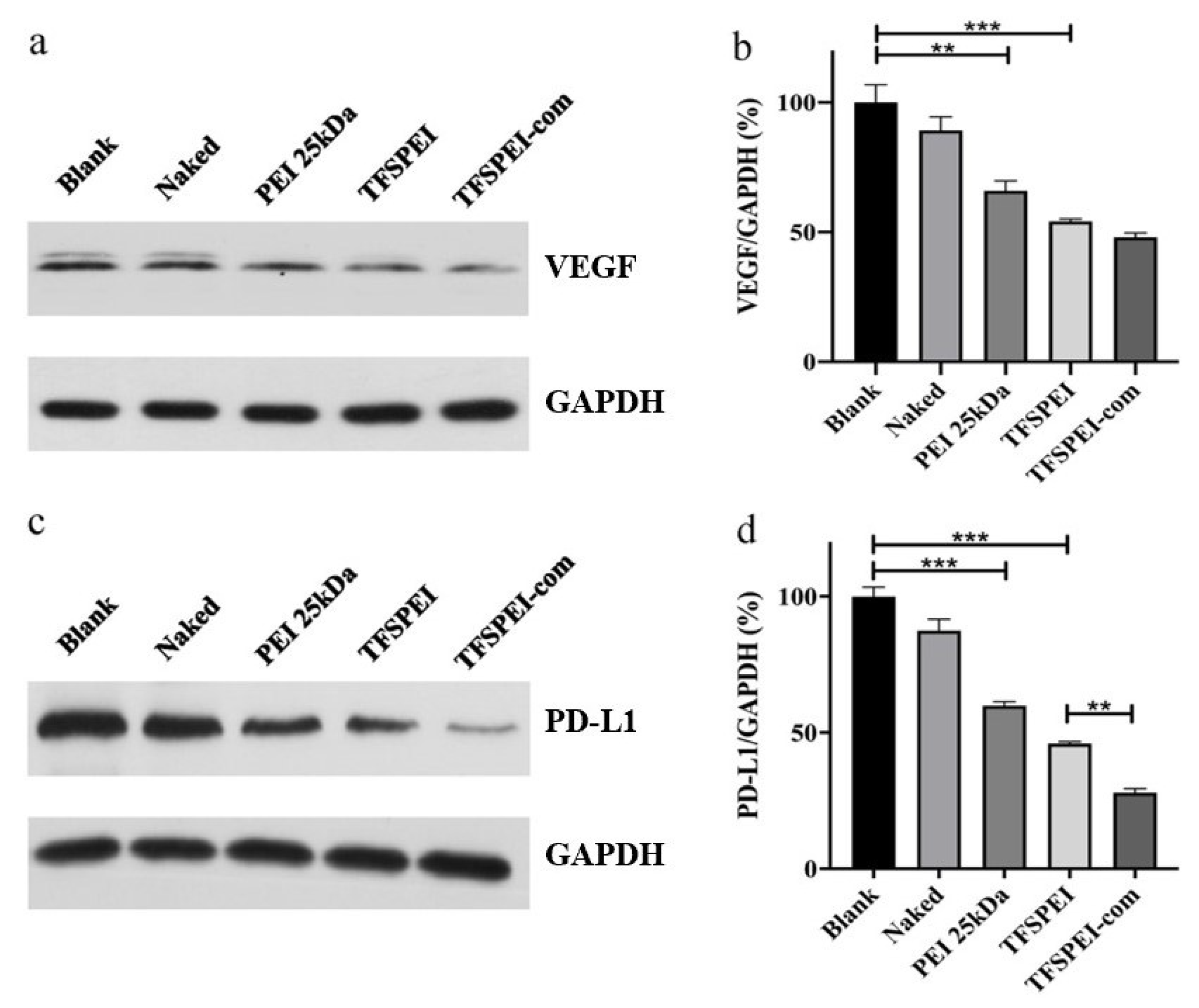
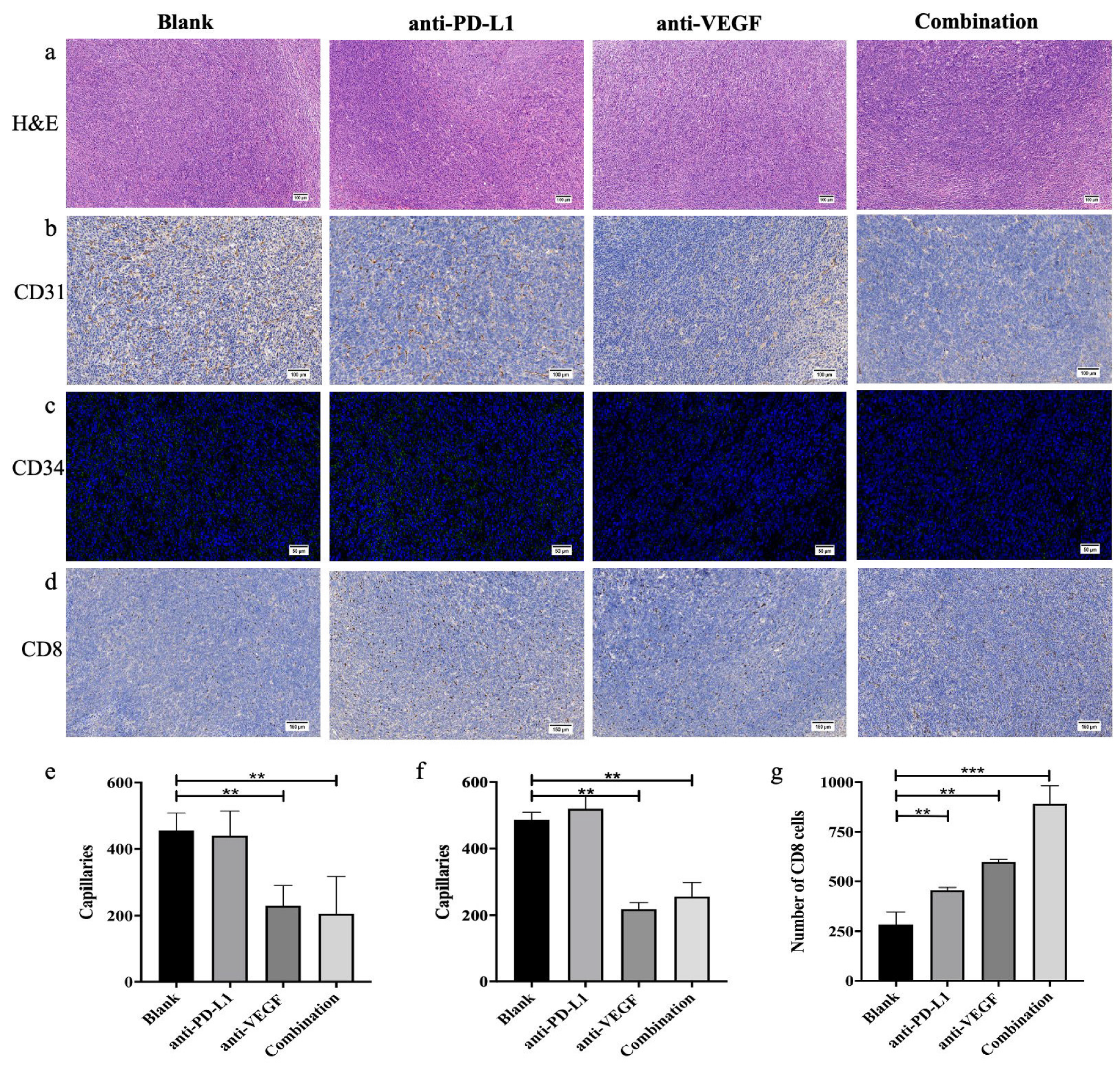
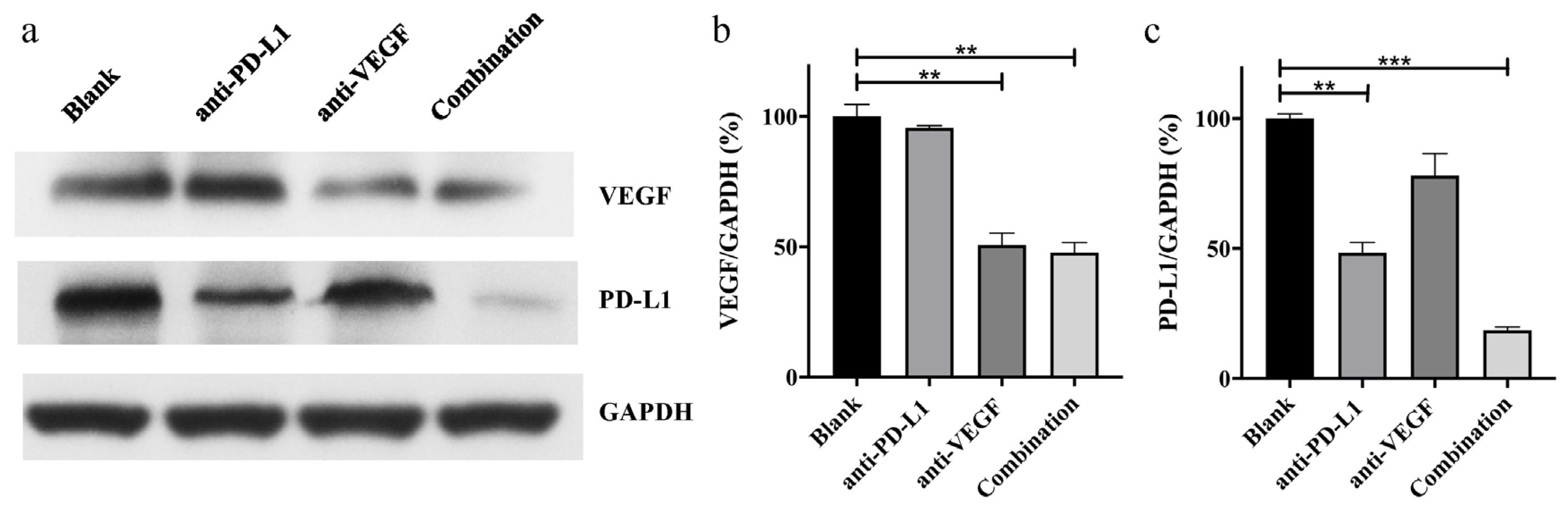
3.9. In Vivo Anti-Tumor Effect Analysis
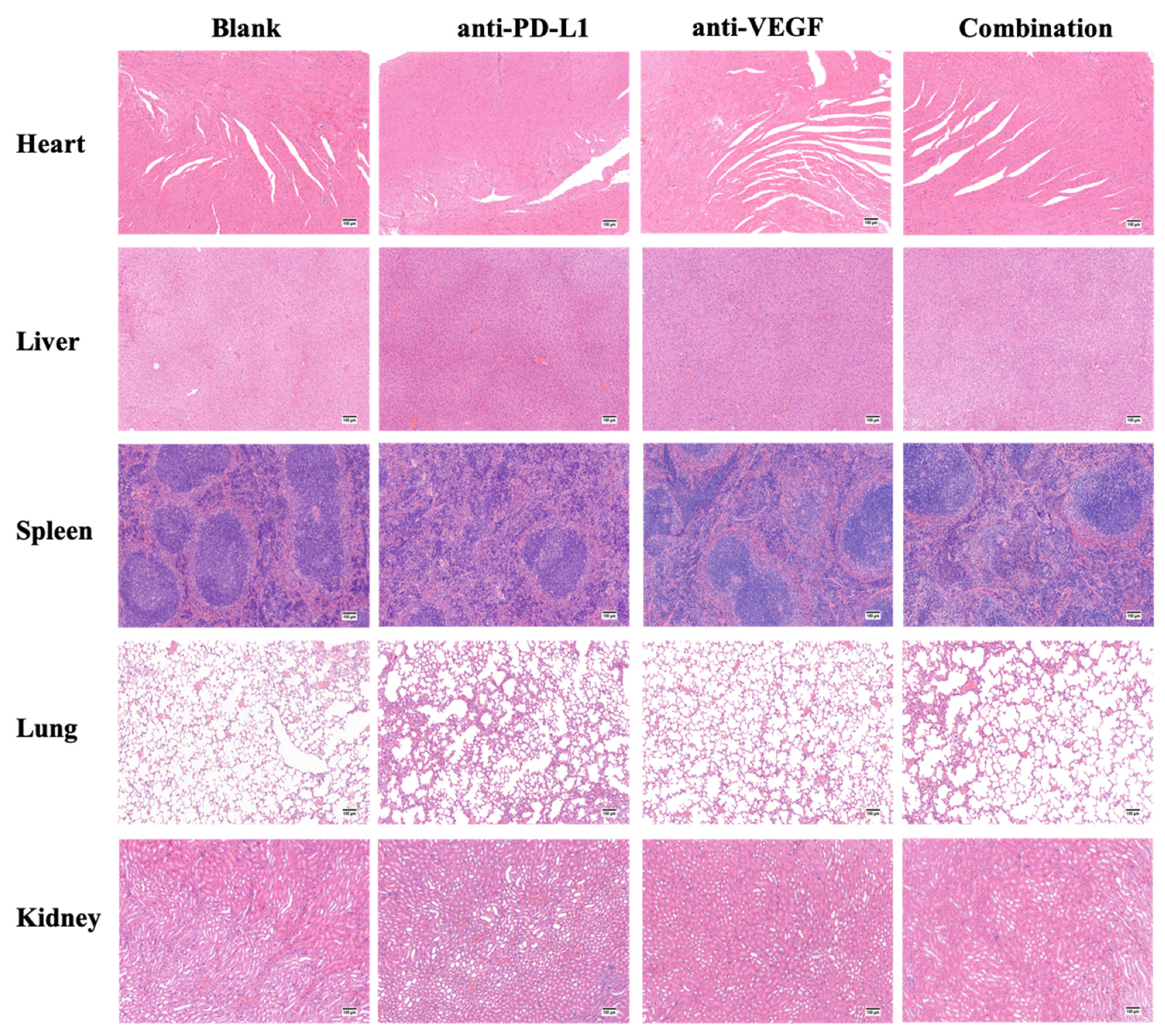
3.10. In Vivo Cytotoxicity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef]
- Rask-Andersen, M.; Masuram, S.; Schioth, H.B. The druggable genome: Evaluation of drug targets in clinical trials suggests major shifts in molecular class and indication. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Ginn, S.L.; Amaya, A.K.; Alexander, I.E.; Edelstein, M.; Abedi, M.R. Gene therapy clinical trials worldwide to 2017: An update. J. Gene Med. 2018, 20, e3015. [Google Scholar] [CrossRef]
- Dunbar, C.E.; High, K.A.; Joung, J.K.; Kohn, D.B.; Ozawa, K.; Sadelain, M. Gene therapy comes of age. Science 2018, 359, eaan4672. [Google Scholar] [CrossRef] [Green Version]
- Gutknecht, M.F.; Seaman, M.E.; Ning, B.; Cornejo, D.A.; Mugler, E.; Antkowiak, P.F.; Moskaluk, C.A.; Hu, S.; Epstein, F.H.; Kelly, K.A. Identification of the S100 fused-type protein hornerin as a regulator of tumor vascularity. Nat. Commun. 2017, 8, 552. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.G.; Jang, M.; Kim, Y.; Leem, G.; Shin, E.C. VEGF-A drives TOX-dependent T cell exhaustion in anti–PD-1–resistant microsatellite stable colorectal cancers. Sci. Immunol. 2019, 4, eaay0555. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Li, L.; Yang, T.; Fan, X.; Wu, G. Combined application of anti-VEGF and anti-EGFR attenuates the growth and angiogenesis of colorectal cancer mainly through suppressing AKT and ERK signaling in mice model. BMC Cancer 2016, 16, 791. [Google Scholar] [CrossRef] [Green Version]
- Eisermann, K.; Fraizer, G. The androgen receptor and VEGF: Mechanisms of androgen-regulated angiogenesis in prostate cancer. Cancers 2017, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Weddell, J.C.; Imoukhuede, P.I. Computational systems biology for the VEGF family in angiogenesis. In Encyclopedia of Cardiovascular Research and Medicine; Elsevier: Amsterdam, The Netherlands, 2018; pp. 659–676. [Google Scholar]
- Meder, L.; Schuldt, P.; Thelen, M.; Schmitt, A.; Dietlein, F.; Klein, S.; Borchmann, S.; Wennhold, K.; Vlasic, I.; Oberbeck, S. Combined VEGF and PD-L1 blockade displays synergistic treatment effects in an autochthonous mouse model of small cell lung cancer. Cancer Res. 2018, 78, 4270–4281. [Google Scholar] [CrossRef] [Green Version]
- Einstein, D.J.; Mcdermott, D.F. Combined blockade of vascular endothelial growth factor and programmed death 1 pathways in advanced kidney cancer. Clin. Adv. Hematol. Oncol. 2017, 15, 478–488. [Google Scholar] [PubMed]
- Voron, T.; Colussi, O.; Marcheteau, E.; Pernot, S.; Terme, M. VEGF-A modulates expression of inhibitory checkpoints on CD8+ T cells in tumors. J. Exp. Med. 2015, 212, 139–148. [Google Scholar] [CrossRef]
- Allen, E.; Jabouille, A.; Rivera, L.B.; Lodewijckx, I.; Missiaen, R.; Steri, V.; Feyen, K.; Tawney, J.; Hanahan, D.; Michael, I.P. Combined antiangiogenic and anti–PD-L1 therapy stimulates tumor immunity through HEV formation. Sci. Transl. Med. 2017, 9, eaak9679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oussa, N.A.; Dahmani, A.; Gomis, M.; Richaud, M.; Andreev, E.; Navab-Daneshmand, A.R.; Taillefer, J.; Carli, C.; Boulet, S.; Sabbagh, L.; et al. VEGF requires the receptor NRP-1 to lnhibit lipopolysaccharide-dependent dendritic cell maturation. J. Immunol. 2016, 197, 3927–3935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.R.; Maute, R.L.; Dulken, B.W.; Hutter, G.; George, B.M.; Mccracken, M.N.; Gupta, R.; Tsai, J.M.; Sinha, R.; Corey, D. PD-1 expression by tumour-associated macrophages inhibits phagocytosis and tumour immunity. Nature 2017, 545, 495–499. [Google Scholar] [CrossRef]
- Akhmetzyanova, I.; Drabczyk, M.; Neff, C.P.; Gibbert, K.; Dietze, K.K.; Werner, T.; Liu, J.; Chen, L.; Lang, K.S.; Palmer, B.E.; et al. PD-L1 expression on retrovirus-infected cells mediates immune escape from CD8+ T cell killing. PLoS Pathog. 2015, 11, e1005224. [Google Scholar]
- Garris, C.S.; Arlauckas, S.P.; Kohler, R.H.; Trefny, M.P.; Garren, S.; Piot, C.; Engblom, C.; Pfirschke, C.; Siwicki, M.; Gungabeesoon, J.; et al. Successful anti-PD-1 cancer immunotherapy requires T cell-dendritic cell crosstalk involving the cytokines IFN-gamma and IL-12. Immunity 2018, 49, 1148–1161.e1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spranger, S.; Spaapen, R.M.; Zha, Y.; Williams, J.; Meng, Y.; Ha, T.T.; Gajewski, T.F. Up-regulation of PD-L1, IDO, and T(regs) in the melanoma tumor microenvironment is driven by CD8(+) T cells. Sci. Transl. Med. 2013, 5, 200ra116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.; Choi, J.; Yang, M.J.; Hong, S.P.; Lee, C.K.; Kubota, Y.; Lim, D.S.; Koh, G.Y. A MST1–FOXO1 cascade establishes endothelial tip cell polarity and facilitates sprouting angiogenesis. Nat. Commun. 2019, 10, 838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duxbury, M.S.; Matros, E.; Ito, H.; Zinner, M.J.; Ashley, S.W.; Whang, E.E. Systemic siRNA-mediated gene silencing: A new approach to targeted therapy of cancer. Ann. Surg. 2004, 240, 667–674, discussion 675–666. [Google Scholar] [CrossRef]
- Wen, Y.; Meng, W.S. Recent in vivo evidences of particle-based delivery of small-interfering RNA (siRNA) into solid tumors. J. Pharm. Innov. 2014, 9, 158–173. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, J.C.; Kowalski, P.S.; Anderson, D.G. Advances in the delivery of RNA therapeutics: From concept to clinical reality. Genome Med. 2017, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.J. Therapeutic siRNA: State of the art. Signal Transduct. Target. Ther. 2020, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Malcolm, D.W.; Benoit, D.S.W. Controlled and sustained delivery of siRNA/NPs from hydrogels expedites bone fracture healing. Biomaterials 2017, 139, 127–138. [Google Scholar] [CrossRef]
- Reis, L.O.; Pereira, T.C.; Favaro, W.J.; Cagnon, V.H.A.; Lopes-Cendes, I.; Ferreira, U. Experimental animal model and RNA interference: A promising association for bladder cancer research. World J. Urol. 2009, 27, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Liu, X.; Zhu, D.; Wang, Y.; Zhang, Z.; Zhou, X.; Qiu, N.; Chen, X.; Shen, Y. Nonviral cancer gene therapy: Delivery cascade and vector nanoproperty integration. Adv. Drug Deliv. Rev. 2017, 115, 115–154. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Mi, C.L.; Cao, X.X.; Wang, T.Y. Progress of cationic gene delivery reagents for non-viral vector. Appl. Microbiol. Biotechnol. 2021, 105, 525–538. [Google Scholar] [CrossRef]
- Pandey, A.P.; Sawant, K.K. Polyethylenimine: A versatile, multifunctional non-viral vector for nucleic acid delivery. Mater. Sci. Eng. C 2016, 68, 904–918. [Google Scholar] [CrossRef]
- Wang, X.; Niu, D.; Hu, C.; Li, P. Polyethyleneimine-based nanocarriers for gene delivery. Curr. Pharm. Des. 2015, 21, 6140–6156. [Google Scholar] [CrossRef]
- Beyerle, A.; Irmler, M.; Beckers, J.; Kissel, T.; Stoeger, T. Toxicity pathway focused gene expression profiling of PEI-based polymers for pulmonary applications. Mol. Pharm. 2010, 7, 727–737. [Google Scholar] [CrossRef]
- Kwon, J.Y. Before and after endosomal escape: Roles of stimuli-converting siRNA/polymer interactions in determining gene silencing efficiency. Acc. Chem. Res. 2012, 45, 1077–1088. [Google Scholar] [CrossRef]
- Benjaminsen, R.V.; Mattebjerg, M.A.; Henriksen, J.R.; Moghimi, S.M.; Andresen, T.L. The possible “proton sponge ” effect of polyethylenimine (PEI) does not include change in lysosomal pH. Mol. Ther. 2013, 21, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Zhu, X.; Guo, Q.; Wang, Y.; Zuo, T.; Luo, F.; Qian, Z. Self-assembled mPEG-PCL-g-PEI micelles for simultaneous codelivery of chemotherapeutic drugs and DNA: Synthesis and characterization in vitro. Int. J. Nanomed. 2012, 7, 1749–1759. [Google Scholar]
- Shi, J.; Xiao, Z.; Votruba, A.R.; Vilos, C.; Farokhzad, O.C. Differentially charged hollow core/shell lipid-polymer-lipid hybrid nanoparticles for small interfering RNA delivery. Angew. Chem. Int. Ed. Engl. 2011, 50, 7027–7031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurtovenko, A.A. Molecular-level insight into the interactions of DNA/polycation complexes with model cell membranes. J. Phys. Chem. B 2019, 123, 6505–6514. [Google Scholar] [CrossRef]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in oligonucleotide drug delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef]
- Setten, R.L.; Rossi, J.J.; Han, S.P. The current state and future directions of RNAi-based therapeutics. Nat. Rev. Drug Discov. 2019, 19, 291. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Liu, H.; Li, L.; Cheng, Y. A fluorinated dendrimer achieves excellent gene transfection efficacy at extremely low nitrogen to phosphorus ratios. Nat. Commun. 2014, 5, 3053. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Lv, J.; Zhuang, Q.; Yang, Z.; Liu, Z. A general strategy towards personalized nanovaccines based on fluoropolymers for post-surgical cancer immunotherapy. Nat. Nanotechnol. 2020, 15, 1–10. [Google Scholar] [CrossRef]
- Xiao, Q.; Rubien, J.D.; Wang, Z.; Reed, E.H.; Hammer, D.A.; Sahoo, D.; Heiney, P.A.; Yadavalli, S.S.; Goulian, M.; Wilner, S.E.; et al. Self-sorting and coassembly of fluorinated, hydrogenated, and hybrid janus dendrimers into dendrimersomes. J. Am. Chem. Soc. 2016, 138, 12655–12663. [Google Scholar] [CrossRef] [Green Version]
- Gillis, E.P.; Eastman, K.J.; Hill, M.D.; Donnelly, D.J.; Meanwell, N.A. Applications of fluorine in medicinal chemistry. J. Med. Chem. 2015, 58, 8315–8359. [Google Scholar] [CrossRef]
- Kukowska-Latallo, J.F.; Chen, C.; Raczka, E.; Qunintana, A.; Rymaszewski, M.; Baker, J.R. Intravascular and endobronchial DNA delivery to murine lung tissue using a novel, nonviral vector. Hum. Gene Ther. 2000, 11, 1385–1395. [Google Scholar] [CrossRef]
- Wang, L.H.; Wu, D.C.; Xu, H.X.; You, Y.Z. High DNA-binding affinity and gene-transfection efficacy of bioreducible cationic nanomicelles with a fluorinated core. Angew. Chem. Int. Ed. Engl. 2016, 55, 755–759. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, W.; Yang, Q.; Niu, H.; Liu, Q.; Liu, Y.; Gao, M.; Xu, M.; Xu, A.; Liu, S. Preliminary investigation on cytotoxicity of fluorinated polymer nanoparticles. J. Environ. Sci. 2018, 69, 217–226. [Google Scholar] [CrossRef]
- Zhang, T.; Huang, Y.; Ma, X.; Gong, N.; Liu, X.; Liu, L.; Ye, X.; Hu, B.; Li, C.; Tian, J.H.; et al. Fluorinated oligoethylenimine nanoassemblies for efficient siRNA-mediated gene silencing in serum-containing media by effective endosomal escape. Nano Lett. 2018, 18, 6301–6311. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cheng, Y. The effect of fluorination on the transfection efficacy of surface-engineered dendrimers. Biomaterials 2014, 35, 6603–6613. [Google Scholar] [CrossRef] [PubMed]
- Duan, F.; Hu, M.; Guo, C.; Song, Y.; Wang, M.; He, L.; Zhang, Z.; Pettinari, R.; Zhou, L. Chromium-based metal-organic framework embedded with cobalt phthalocyanine for the sensitively impedimetric cytosensing of colorectal cancer (CT26) cells and cell imaging. Chem. Eng. J. 2020, 398. [Google Scholar] [CrossRef]
- Midoux, P.; Breuzard, G.; Gomez, P.J.; Pichon, C. Polymer-based gene delivery: A current review on the uptake and intracellular trafficking of polyplexes. Curr. Gene Ther. 2008, 8, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Lertkiatmongkol, P.; Liao, D.; Mei, H.; Hu, Y.; Newman, P.J. Endothelial functions of platelet/endothelial cell adhesion molecule-1 (CD31). Curr. Opin. Hematol. 2016, 23, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ravin, S.S.; Reik, A.; Liu, P.Q.; Li, L.; Wu, X.; Su, L.; Raley, C.; Theobald, N.; Choi, U.; Song, A.H. Targeted gene addition in human CD34(+) hematopoietic cells for correction of X-linked chronic granulomatous disease. Nat. Biotechnol. 2016, 34, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, G.; Stockinger, H.; Balderas, R.; van Zelm, M.C.; Zola, H.; Hart, D.; Engel, P. Nomenclature of CD molecules from the tenth human leucocyte differentiation antigen workshop. Clin. Transl. Immunol. 2016, 5, e57. [Google Scholar] [CrossRef] [Green Version]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Yuan, Z.; Jin, Y.; Zhang, W.; Yuan, W.-E. Novel Fluorinated Spermine and Small Molecule PEI to Deliver Anti-PD-L1 and Anti-VEGF siRNA for Highly Efficient Tumor Therapy. Pharmaceutics 2021, 13, 2058. https://doi.org/10.3390/pharmaceutics13122058
Zhang Y, Yuan Z, Jin Y, Zhang W, Yuan W-E. Novel Fluorinated Spermine and Small Molecule PEI to Deliver Anti-PD-L1 and Anti-VEGF siRNA for Highly Efficient Tumor Therapy. Pharmaceutics. 2021; 13(12):2058. https://doi.org/10.3390/pharmaceutics13122058
Chicago/Turabian StyleZhang, Yihui, Zihan Yuan, Yi Jin, Wenkai Zhang, and Wei-En Yuan. 2021. "Novel Fluorinated Spermine and Small Molecule PEI to Deliver Anti-PD-L1 and Anti-VEGF siRNA for Highly Efficient Tumor Therapy" Pharmaceutics 13, no. 12: 2058. https://doi.org/10.3390/pharmaceutics13122058