
Carotenoids from Persimmon (Diospyros kaki Thunb.) Byproducts Exert Photoprotective, Antioxidative and Microbial Anti-Adhesive Effects on HaCaT

 , , and
, , and 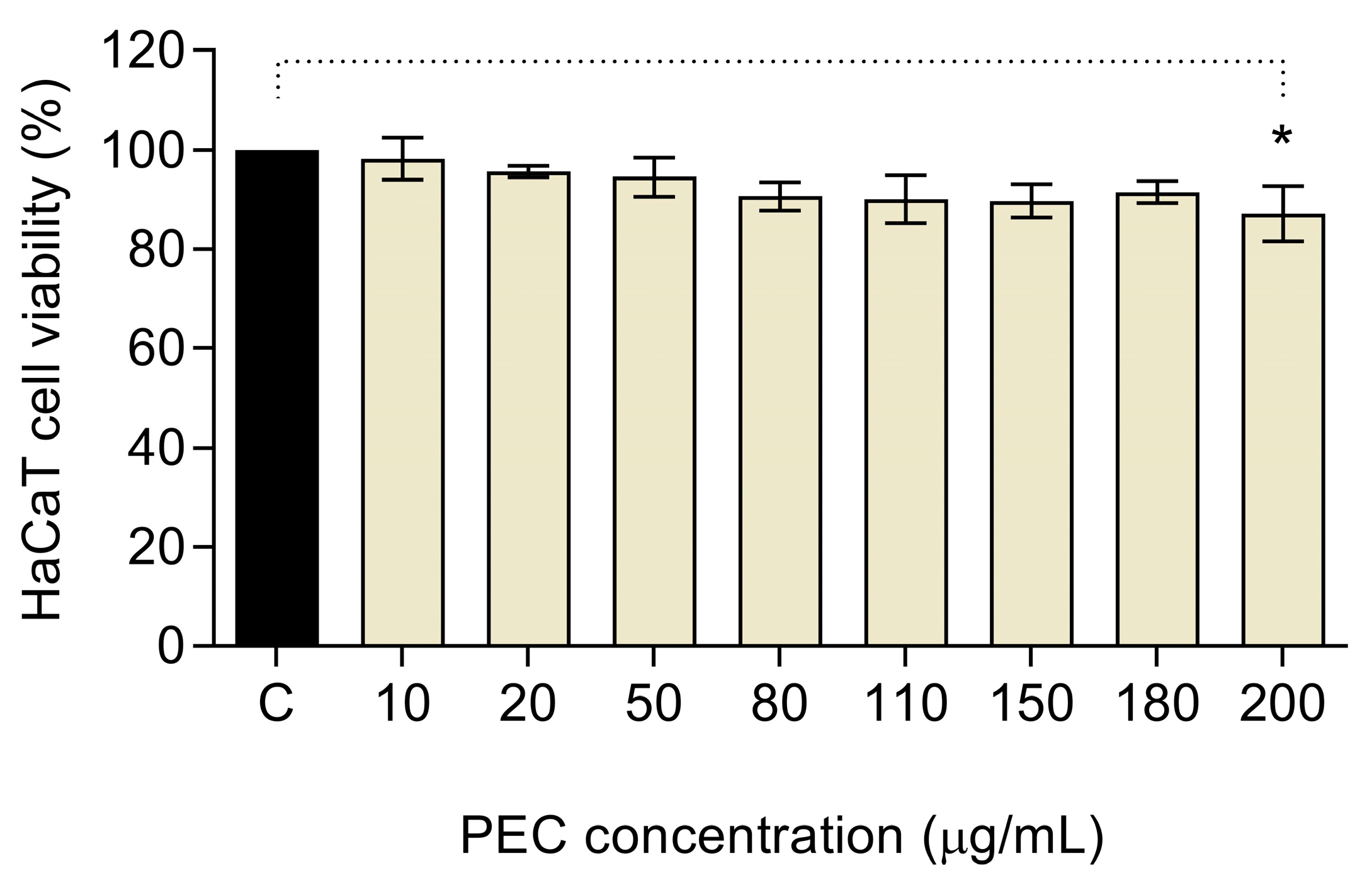{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and Carotenoid-Rich Acetone Extraction
2.3. Cellular Assays
2.3.1. Culture of HaCaT Cell Line
2.3.2. Cytotoxicity Assay
2.3.3. In Vitro UV Photoprotective Assay
2.3.4. Intracellular ROS Scavenging Activity
2.4. Microbial Assays
2.4.1. Microbiology Culture Media and Reagents
2.4.2. Microbial Strains and Inoculum Preparation
2.4.3. Antimicrobial Assay
2.4.4. Inhibitory Microbial Adhesion to HaCaT Cell Monolayers
2.5. In Vitro Permeation Test
2.5.1. PEC Preparation
2.5.2. Franz Diffusion Cell System
2.5.3. High Performance Liquid Chromatography and Tandem Mass Spectrometry (HPLC-PDA-MS/MS) Analysis
2.6. Statistical Analysis
3. Results
3.1. Cytotoxic Activity of PEC in HaCaT Cells
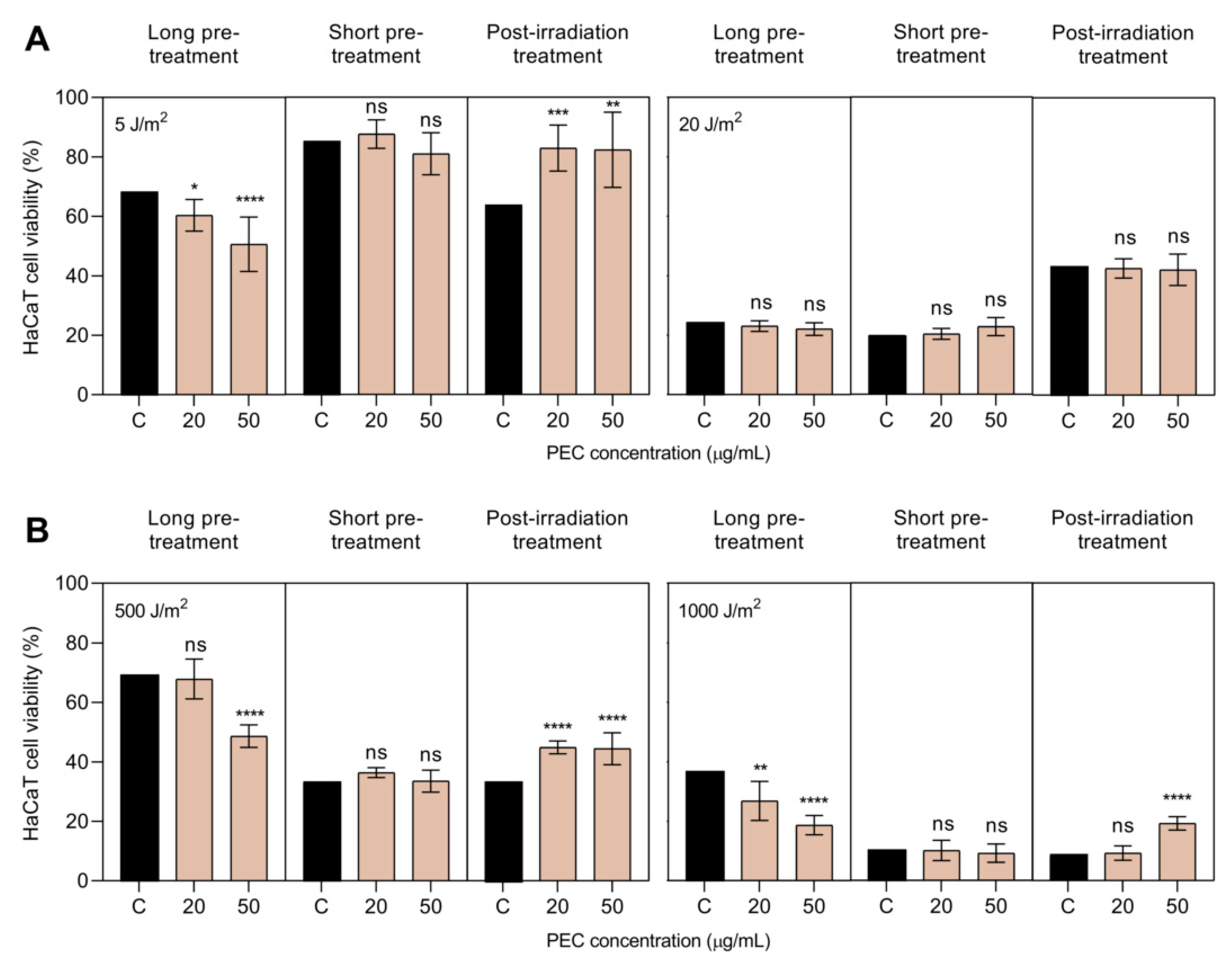
3.2. Photoprotective Activity of PEC against UV-A and UV-B Irradiation
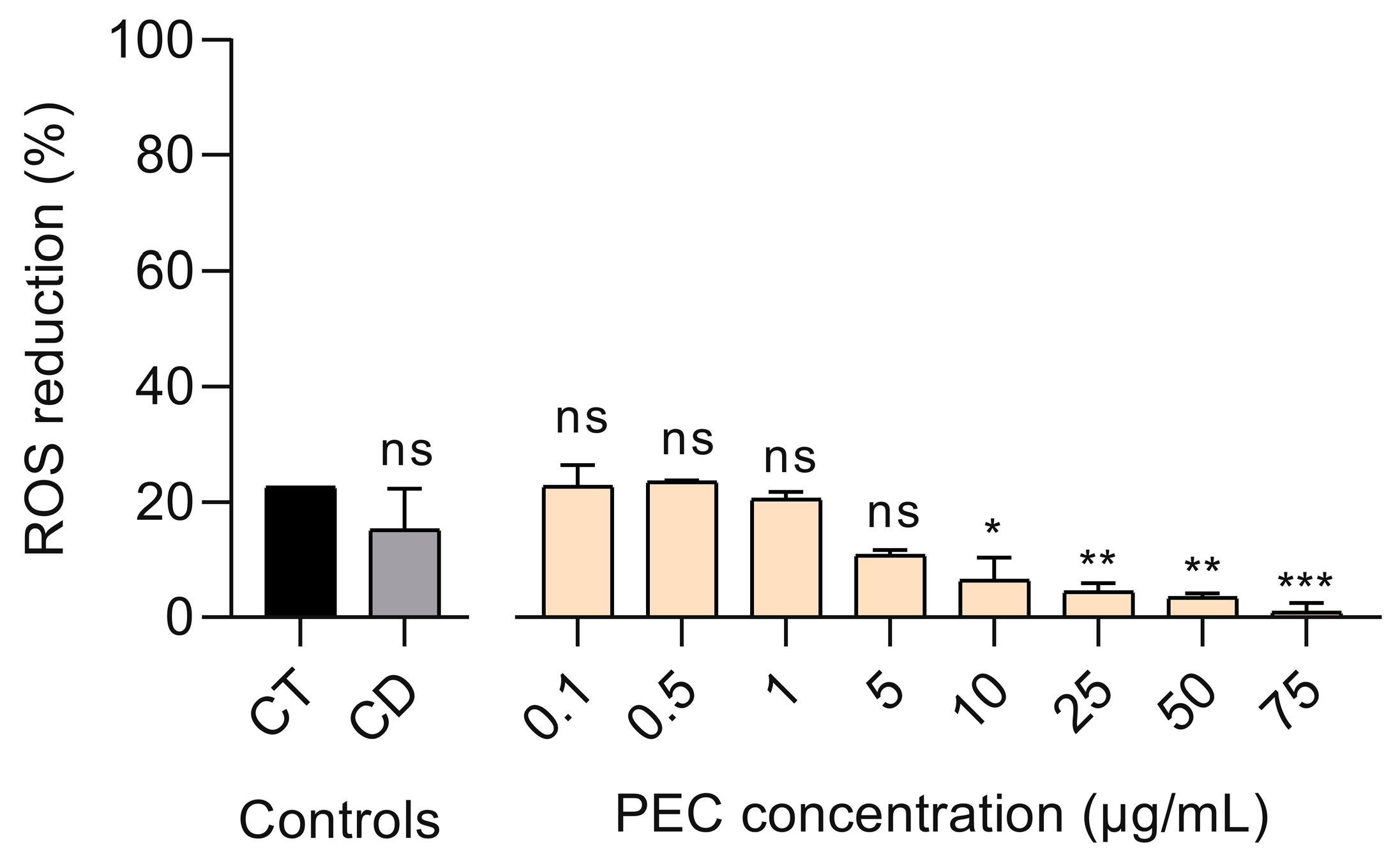
3.3. Effect of PEC on UV-B-Induced ROS Production in Treated HaCaT Cells
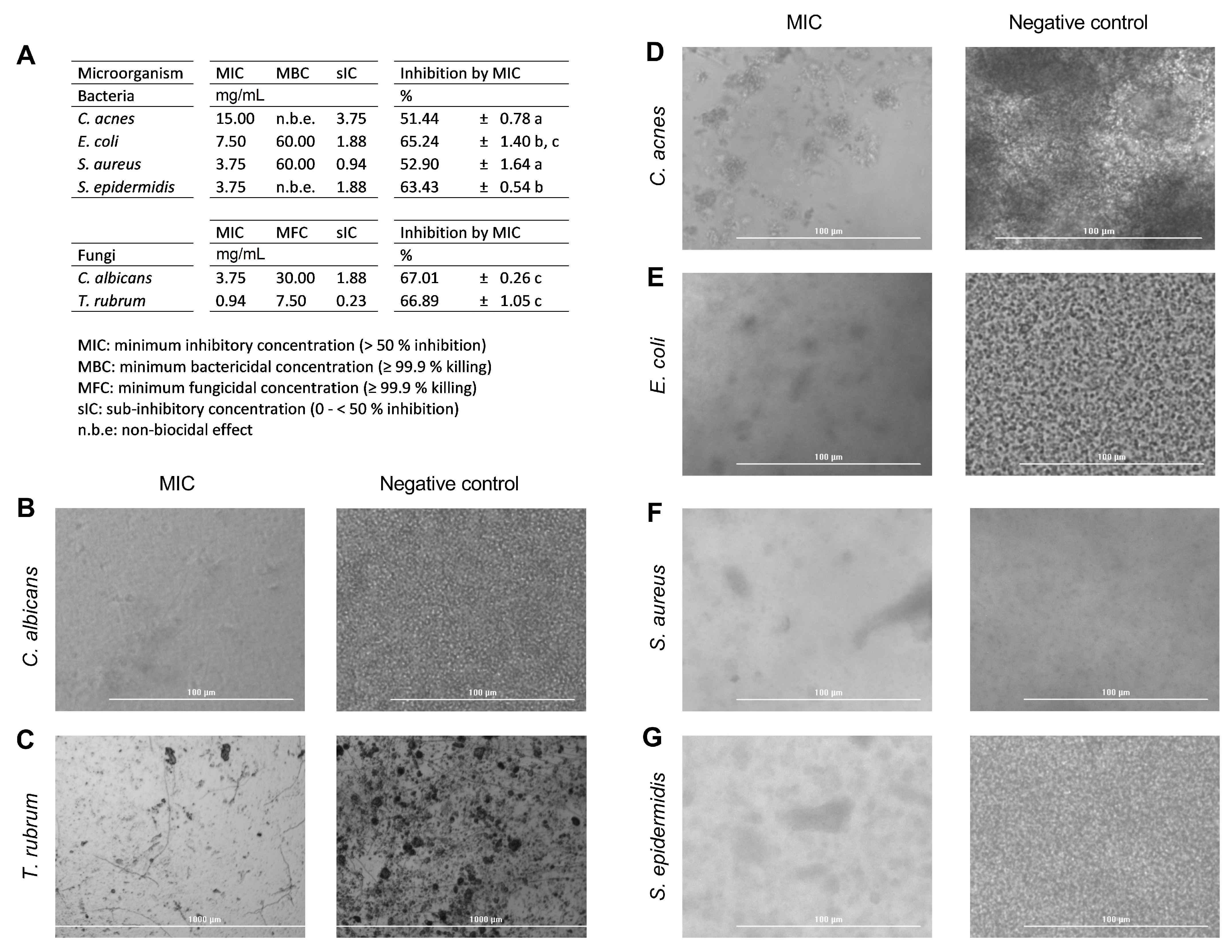
3.4. Antimicrobial Activity Results
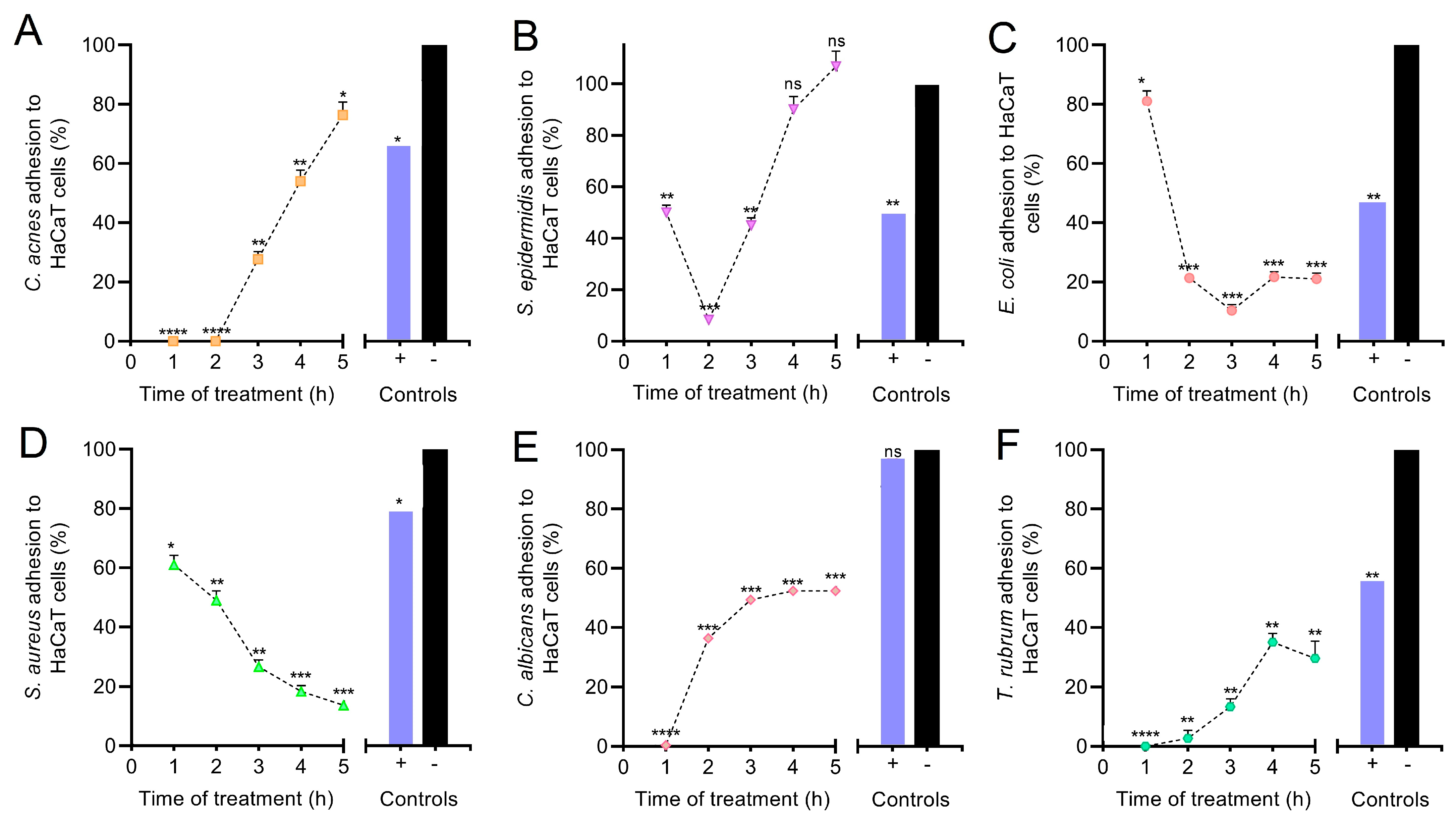
3.5. Anti-Adhesive Effect of PEC on HaCaT Monolayer Cells
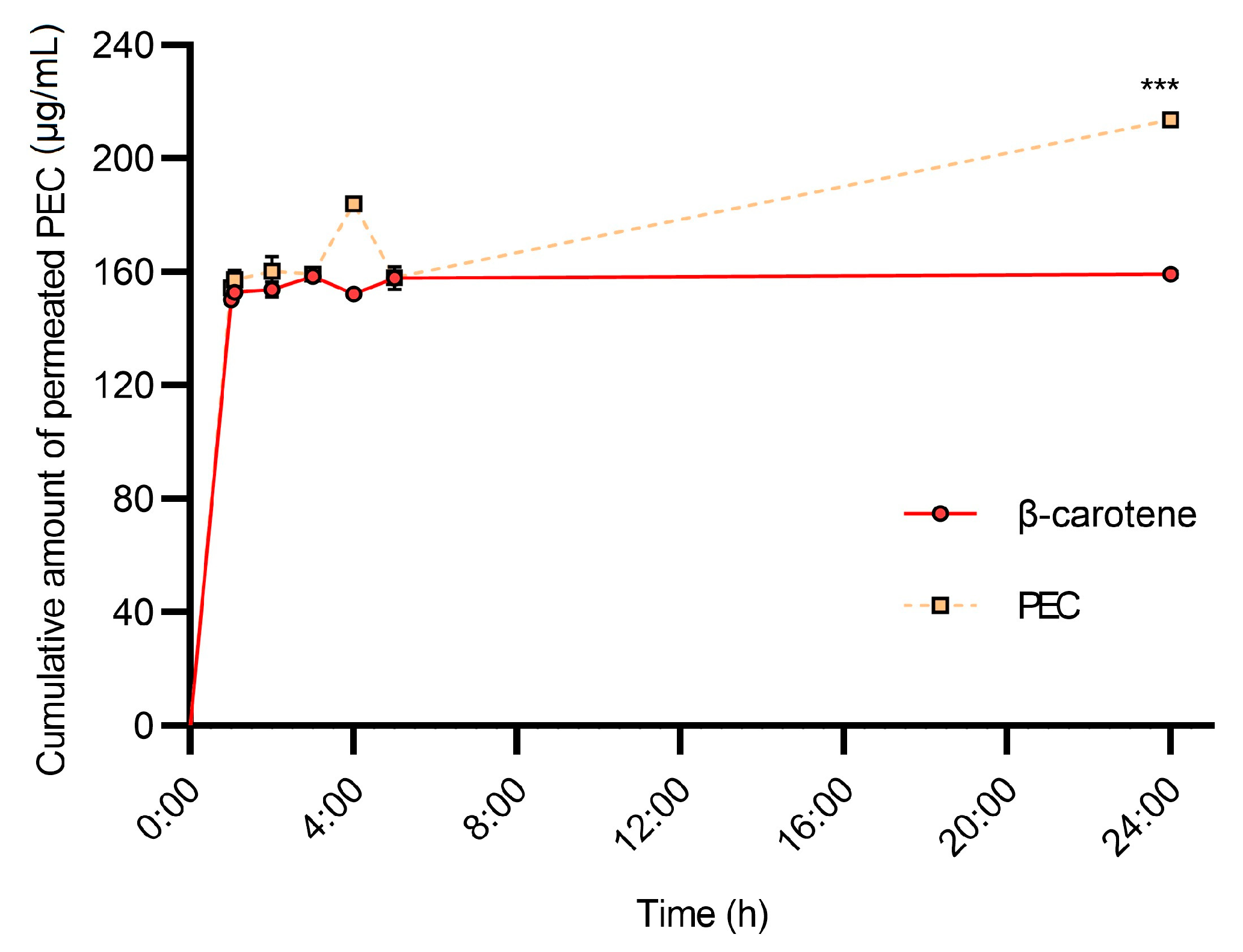
3.6. HPLC-PDA-MS/MS Analysis of Permeated Samples
4. Discussion
4.1. UV Radiation and ROS Production
4.2. Microbiology
4.3. In Vitro Permeation of PEC Carotenoids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woodby, B.; Penta, K.; Pecorelli, A.; Lila, M.A.; Valacchi, G. Skin Health from the Inside Out. Annu. Rev. Food Sci. Technol. 2020, 11, 235–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meléndez-Martínez, A.J.; Vicario, I.M.; Francisco, H. Importancia nutricional de los pigmentos carotenoides. Arch. Lat. Nutr. 2004, 54, 149–155. [Google Scholar]
- Zerres, S.; Stahl, W. Carotenoids in human skin. Biochim. Biophys. Acta Bba–Mol. Cell Biol. Lipids 2020, 1865, 158588. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Pal, A.; Singh, D.; Ansari, K.M. Topical application of Nexrutine inhibits ultraviolet B-induced cutaneous inflammatory responses in SKH-1 hairless mouse. Photodermatol. Photoimmunol. Photomed. 2017, 34, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baccarin, T.; Mitjans, M.; Ramos, D.; Lemos-Senna, E.; Vinardell, M.P. Photoprotection by Punica granatum seed oil nanoemulsion entrapping polyphenol-rich ethyl acetate fraction against UVB-induced DNA damage in human keratinocyte (HaCaT) cell line. J. Photochem. Photobiol. B Biol. 2015, 153, 127–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, M.; Fukunaga, A.; Washio, K.; Taguchi, K.; Oda, Y.; Ogura, K.; Nishigori, C. Anti-Inflammatory Role of Langerhans Cells and Apoptotic Keratinocytes in Ultraviolet-B–Induced Cutaneous Inflammation. J. Immunol. 2017, 199, 2937–2947. [Google Scholar] [CrossRef] [Green Version]
- Nichols, J.A.; Katiyar, S.K. Skin photoprotection by natural polyphenols: Anti-inflammatory, antioxidant and DNA repair mechanisms. Arch. Derm. Res. 2009, 302, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Nishigori, C. Cellular aspects of photocarcinogenesis. Photochem. Photobiol. Sci. 2006, 5, 208–214. [Google Scholar] [CrossRef]
- Center, S.S. UV Light. Available online: http://solar-center.stanford.edu/about/uvlight.html (accessed on 3 May 2021).
- Psotova, J.; Svobodova, A.; Kolarova, H.; Walterova, D. Photoprotective properties of Prunella vulgaris and rosmarinic acid on human keratinocytes. J. Photochem. Photobiol. B Biol. 2006, 84, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.-L.; Fang, J.-Y.; Chen, M.; Wu, C.-J.; Huang, C.-C.; Hung, C.-F. Chrysin Protects Epidermal Keratinocytes from UVA- and UVB-Induced Damage. J. Agric. Food Chem. 2011, 59, 8391–8400. [Google Scholar] [CrossRef]
- Balić, A.; Vlašić, D.; Mokos, M.; Marinović, B. The Role of the Skin Barrier in Periorificial Dermatitis. Acta Derm. Croat. 2019, 27, 169–179. [Google Scholar]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Golmohammadzadeh, S.; Imani, F.; Hosseinzadeh, H.; Jaafari, M.R. Preparation, Characterization and Evaluation of Sun Protective and Moisturizing Effects of Nanoliposomes Containing Safranal. Iran. J. Basic Med. Sci. 2011, 14, 521–533. [Google Scholar] [PubMed]
- Oh, J.; Kim, J.H.; Park, J.G.; Yi, Y.-S.; Park, K.W.; Rho, H.S.; Lee, M.-S.; Yoo, J.W.; Kang, S.-H.; Hong, Y.D.; et al. Radical Scavenging Activity-Based and AP-1-Targeted Anti-Inflammatory Effects of Lutein in Macrophage-Like and Skin Keratinocytic Cells. Mediat. Inflamm. 2013, 2013, 787042. [Google Scholar] [CrossRef]
- Svobodová, A.; Psotová, J.; Walterová, D. Natural phenolics in the prevention of UV-induced skin damage. A review. Biomed. Pap. 2003, 147, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Reis-Mansur, M.C.P.P.; Cardoso-Rurr, J.S.; Silva, J.V.M.A.; De Souza, G.R.; Cardoso, V.D.S.; Mansoldo, F.; Pinheiro, Y.; Schultz, J.; Balottin, L.B.L.; Da Silva, A.J.R.; et al. Carotenoids from UV-resistant Antarctic Microbacterium sp. LEMMJ01. Sci. Rep. 2019, 9, 9554. [Google Scholar] [CrossRef] [PubMed]
- Abney, K.R.; Kopsell, D.; Sams, C.E.; Zivanovic, S.; Kopsell, D.E. UV-B Radiation Impacts Shoot Tissue Pigment Composition in Allium fistulosum L. Cultigens. Sci. World J. 2013, 2013, 513867. [Google Scholar] [CrossRef] [Green Version]
- Becatti, E.; Petroni, K.; Giuntini, D.; Castagna, A.; Calvenzani, V.; Serra, G.; Mensuali-Sodi, A.; Tonelli, C.; Ranieri, A. Solar UV−B Radiation Influences Carotenoid Accumulation of Tomato Fruit through Both Ethylene-Dependent and -Independent Mechanisms. J. Agric. Food Chem. 2009, 57, 10979–10989. [Google Scholar] [CrossRef]
- Biswas, D.K.; Ma, B.-L.; Xu, H.; Li, Y.; Jiang, G. Lutein-mediated photoprotection of photosynthetic machinery in Arabidopsis thaliana exposed to chronic low ultraviolet-B radiation. J. Plant Physiol. 2020, 248, 153160. [Google Scholar] [CrossRef]
- Assumpção, C.F.; Assis, R.Q.; Poletto, V.S.H.; Castagna, A.; Ranieri, A.; Neugart, S.; Flôres, S.H.; Rios, A.O. Application of supplemental UV-B radiation in pre-harvest to enhance health-promoting compounds accumulation in green and red lettuce. J. Food Process. Preserv. 2019, 43, e14213. [Google Scholar] [CrossRef]
- Ascenso, A.; Pedrosa, T.; Pinho, S.; Pinho, F.; De Oliveira, J.M.P.F.; Marques, H.; Oliveira, H.; Simões, S.; Santos, C. The Effect of Lycopene Preexposure on UV-B-Irradiated Human Keratinocytes. Oxidative Med. Cell. Longev. 2016, 2016, 8214631. [Google Scholar] [CrossRef] [PubMed]
- Santocono, M.; Zurria, M.; Berrettini, M.; Fedeli, D.; Falcioni, G. Influence of astaxanthin, zeaxanthin and lutein on DNA damage and repair in UVA-irradiated cells. J. Photochem. Photobiol. B Biol. 2006, 85, 205–215. [Google Scholar] [CrossRef]
- Tominaga, K.; Hongo, N.; Fujishita, M.; Takahashi, Y.; Adachi, Y. Protective effects of astaxanthin on skin deterioration. J. Clin. Biochem. Nutr. 2017, 61, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grice, J.E.; Zhang, Q.; Roberts, M.S. Chemical structure—Skin transport relationships. In Toxicology of the Skin, 1st ed.; Monteiro-Riviere, N.A., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 55–68. [Google Scholar]
- Ghodratizadeh, S.; Kanbak, G.; Beyramzadeh, M.; Dikmen, Z.G.; Memarzadeh, S.; Habibian, R. Effect of carotenoid β-cryptoxanthin on cellular and humoral immune response in rabbit. Veter-Res. Commun. 2013, 38, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Gea-Botella, S.; Agulló, L.; Martí, N.; Martínez-Madrid, M.; Lizama, V.; Martín-Bermudo, F.; Berná, G.; Saura, D.; Valero, M. Carotenoids from persimmon juice processing. Food Res. Int. 2021, 141, 109882. [Google Scholar] [CrossRef]
- Salazar-Bermeo, J.; Moreno-Chamba, B.; Martínez-Madrid, M.C.; Saura, D.; Valero, M.; Martí, N. Potential of Persimmon Dietary Fiber Obtained from Byproducts as Antioxidant, Prebiotic and Modulating Agent of the Intestinal Epithelial Barrier Function. Antioxidants 2021, 10, 1668. [Google Scholar] [CrossRef]
- Rodriguez-Luna, A.; Ávila-Román, J.; Oliveira, H.; Motilva, V.; Talero, E. Fucoxanthin and Rosmarinic Acid Combination Has Anti-Inflammatory Effects through Regulation of NLRP3 Inflammasome in UVB-Exposed HaCaT Keratinocytes. Mar. Drugs 2019, 17, 451. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, M.T.; Silva, A.C.G.; Nascimento, T.L.; A Diniz, D.G.; Valadares, M.C.; Lima, E.M. Protective effect of sucupira oil nanoemulsion against oxidative stress in UVA-irradiated HaCaT cells. J. Pharm. Pharm. 2019, 71, 1532–1543. [Google Scholar] [CrossRef]
- Ravi, H.; Kurrey, N.; Manabe, Y.; Sugawara, T.; Baskaran, V. Polymeric chitosan-glycolipid nanocarriers for an effective delivery of marine carotenoid fucoxanthin for induction of apoptosis in human colon cancer cells (Caco-2 cells). Mater. Sci. Eng. C 2018, 91, 785–795. [Google Scholar] [CrossRef]
- Jiménez-Zamora, A.; Pastoriza, S.; Rufián-Henares, J.Á. Revalorization of coffee by-products. Prebiotic, antimicrobial and antioxidant properties. LWT Food Sci. Technol. 2015, 61, 12–18. [Google Scholar] [CrossRef]
- Sardi, J.D.C.O.; Polaquini, C.R.; Freires, I.A.; Galvao, L.C.D.C.; Lazarini, J.G.; Torrezan, G.S.; Regasini, L.O.; Rosalen, P.L. Antibacterial activity of diacetylcurcumin against Staphylococcus aureus results in decreased biofilm and cellular adhesion. J. Med Microbiol. 2017, 66, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Ashtikar, M.; Matthäus, C.; Schmitt, M.; Krafft, C.; Fahr, A.; Popp, J. Non-invasive depth profile imaging of the stratum corneum using confocal Raman microscopy: First insights into the method. Eur. J. Pharm. Sci. 2013, 50, 601–608. [Google Scholar] [CrossRef]
- Ng, S.-F.; Rouse, J.J.; Sanderson, F.D.; Meidan, V.; Eccleston, G.M. Validation of a Static Franz Diffusion Cell System for In Vitro Permeation Studies. Aaps Pharmscitech 2010, 11, 1432–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopec, R.; Schweiggert, R.M.; Riedl, K.; Carle, R.; Schwartz, S.J. Comparison of high-performance liquid chromatography/tandem mass spectrometry and high-performance liquid chromatography/photo-diode array detection for the quantitation of carotenoids, retinyl esters, α-tocopherol and phylloquinone in chylomicron-rich fr. Rapid Commun. Mass Spectrom. 2013, 27, 1393–1402. [Google Scholar] [CrossRef] [Green Version]
- Mercadante, A.Z.; Rodrigues, D.B.; Petry, F.C.; Mariutti, L. Carotenoid esters in foods–A review and practical directions on analysis and occurrence. Food Res. Int. 2017, 99, 830–850. [Google Scholar] [CrossRef] [PubMed]
- Rivera, S.M.; Christou, P.; I Garayoa, R.C. Identification of carotenoids using mass spectrometry. Mass Spectrom. Rev. 2014, 33, 353–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélez, S.M.R. Guide for Carotenoid Identification in Biological Samples. J. Nat. Prod. 2016, 79, 1473–1484. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Sato, M.; Okamoto, M.; Masuda, J.; Yamaki, S.; Tamari, M.; Tanokashira, Y.; Kishimoto, S.; Ohmiya, A.; Abe, T.; et al. Metabolome-based discrimination of chrysanthemum cultivars for the efficient generation of flower color variations in mutation breeding. Metabolomics 2019, 15, 118. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, D.; Raffa, S.; Flori, E.; Aspite, N.; Briganti, S.; Cardinali, G.; Torrisi, M.R.; Picardo, M. Keratinocyte growth factor down-regulates intracellular ROS production induced by UVB. J. Derm. Sci. 2009, 54, 106–113. [Google Scholar] [CrossRef]
- Rohmah, M.; Raharjo, S.; Hidayat, C.; Martien, R. Application of Response Surface Methodology for the Optimization of β-Carotene-Loaded Nanostructured Lipid Carrier from Mixtures of Palm Stearin and Palm Olein. J. Am. Oil Chem. Soc. 2019, 97, 213–223. [Google Scholar] [CrossRef]
- Godar, D.E. UV Doses Worldwide. Photochem. Photobiol. 2005, 81, 736–749. [Google Scholar] [CrossRef] [PubMed]
- Sajo, M.E.J.; Kim, C.-S.; Kim, S.-K.; Shim, K.Y.; Kang, T.-Y.; Lee, K.-J. Antioxidant and Anti-Inflammatory Effects of Shungite against Ultraviolet B Irradiation-Induced Skin Damage in Hairless Mice. Oxidative Med. Cell. Longev. 2017, 2017, 7340143. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Hu, J.Y.; Wang, S.Q. The role of antioxidants in photoprotection: A critical review. J. Am. Acad. Derm. 2012, 67, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Vostálová, J.; Zdařilová, A.; Svobodová, A. Prunella vulgaris extract and rosmarinic acid prevent UVB-induced DNA damage and oxidative stress in HaCaT keratinocytes. Arch. Derm. Res. 2010, 302, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.L.; Howland, M.C.; Szmodis, A.W.; Li, Q.; Daemen, L.L.; Parikh, A.; Majewski, J. Early Stages of Oxidative Stress-Induced Membrane Permeabilization: A Neutron Reflectometry Study. J. Am. Chem. Soc. 2009, 131, 3631–3638. [Google Scholar] [CrossRef] [Green Version]
- Yeo, S.-K.; Liong, M.-T. Effects and applications of sub-lethal ultrasound, electroporation and UV radiations in bioprocessing. Ann. Microbiol. 2013, 63, 813–824. [Google Scholar] [CrossRef]
- Fisher, A.B. Peroxiredoxin 6 in the repair of peroxidized cell membranes and cell signaling. Arch. Biochem. Biophys. 2017, 617, 68–83. [Google Scholar] [CrossRef] [Green Version]
- Lohan, S.B.; Vitt, K.; Scholz, P.; Keck, C.; Meinke, M.C. ROS production and glutathione response in keratinocytes after application of β-carotene and VIS/NIR irradiation. Chem.-Biol. Interact. 2018, 280, 1–7. [Google Scholar] [CrossRef]
- Karpiński, T.M.; Adamczak, A. Fucoxanthin—An Antibacterial Carotenoid. Antioxidants 2019, 8, 239. [Google Scholar] [CrossRef] [Green Version]
- Keceli, T.M.; Erginkaya, Z.; Turkkan, E.; Kaya, Ü. Antioxidant and Antibacterial Effects of Carotenoids Extracted from Rhodotorula glutinis Strains. Asian J. Chem. 2013, 25, 42–46. [Google Scholar] [CrossRef]
- Manimala, M.R.A.; Murugesan, R. In vitro antioxidant and antimicrobial activity of carotenoid pigment extracted from Sporobolomyces sp. isolated from natural source. J. Appl. Nat. Sci. 2014, 6, 649–653. [Google Scholar] [CrossRef]
- Vargas-Sinisterra, A.F.; Ramírez-Castrillón, M. Yeast carotenoids: Production and activity as antimicrobial biomolecule. Arch. Microbiol. 2021, 203, 873–888. [Google Scholar] [CrossRef] [PubMed]
- Hraishawi, R.M.; Abdul-Razak, A.S.; Al-Hayder, M.N.; Al-wafi, H. Investigation the antimicrobial and antioxidant activity of lycopene extraction from Solanum Lycopersicum. Eur. Asian J. Biosci. 2020, 14, 5305–5310. [Google Scholar]
- Maretti, E.; Leo, E.; Rustichelli, C.; Truzzi, E.; Siligardi, C.; Iannuccelli, V. In vivo β-carotene skin permeation modulated by Nanostructured Lipid Carriers. Int. J. Pharm. 2021, 597, 120322. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gea-Botella, S.; Moreno-Chamba, B.; de la Casa, L.; Salazar-Bermeo, J.; Martí, N.; Martínez-Madrid, M.C.; Valero, M.; Saura, D. Carotenoids from Persimmon (Diospyros kaki Thunb.) Byproducts Exert Photoprotective, Antioxidative and Microbial Anti-Adhesive Effects on HaCaT. Pharmaceutics 2021, 13, 1898. https://doi.org/10.3390/pharmaceutics13111898
Gea-Botella S, Moreno-Chamba B, de la Casa L, Salazar-Bermeo J, Martí N, Martínez-Madrid MC, Valero M, Saura D. Carotenoids from Persimmon (Diospyros kaki Thunb.) Byproducts Exert Photoprotective, Antioxidative and Microbial Anti-Adhesive Effects on HaCaT. Pharmaceutics. 2021; 13(11):1898. https://doi.org/10.3390/pharmaceutics13111898
Chicago/Turabian StyleGea-Botella, Sara, Bryan Moreno-Chamba, Laura de la Casa, Julio Salazar-Bermeo, Nuria Martí, María Concepción Martínez-Madrid, Manuel Valero, and Domingo Saura. 2021. "Carotenoids from Persimmon (Diospyros kaki Thunb.) Byproducts Exert Photoprotective, Antioxidative and Microbial Anti-Adhesive Effects on HaCaT" Pharmaceutics 13, no. 11: 1898. https://doi.org/10.3390/pharmaceutics13111898