The Positive and Negative Outcome of Morphine and Disulfiram Subacute Co-Administration in Rats in the Absence of Ethanol Challenge

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Drugs and Reagents
2.2. Animals
- (1)
- DSF at a dose of 25 mg/kg, i.p.
- (2)
- DSF at a dose of 50 mg/kg, i.p.
- (3)
- DSF at a dose of 100 mg/kg, i.p.
- (4)
- DSF (25 mg/kg) + MRF (10 mg/kg), i.p.
- (5)
- DSF (50 mg/kg) + MRF (10 mg/kg), i.p.
- (6)
- DSF (100 mg/kg) + MRF (10 mg/kg), i.p.
- (7)
- MRF (10 mg/kg), i.p.
- (8)
- 0.1% methylcellulose (vehicle), i.p.
- (1)
- DSF at a dose of 100 mg/kg, i.p.
- (2)
- DSF (100 mg/kg) + MRF (10 mg/kg), i.p.
- (3)
- MRF (10 mg/kg), i.p.
- (4)
- 0.1% methylcellulose (vehicle), i.p.
- (1)
- DSF at a dose of 100 mg/kg, i.p.
- (2)
- DSF (100 mg/kg) + MRF (10 mg/kg), i.p.
- (3)
- MRF (10 mg/kg), i.p.
- (4)
- 0.1% methylcellulose (vehicle), i.p.
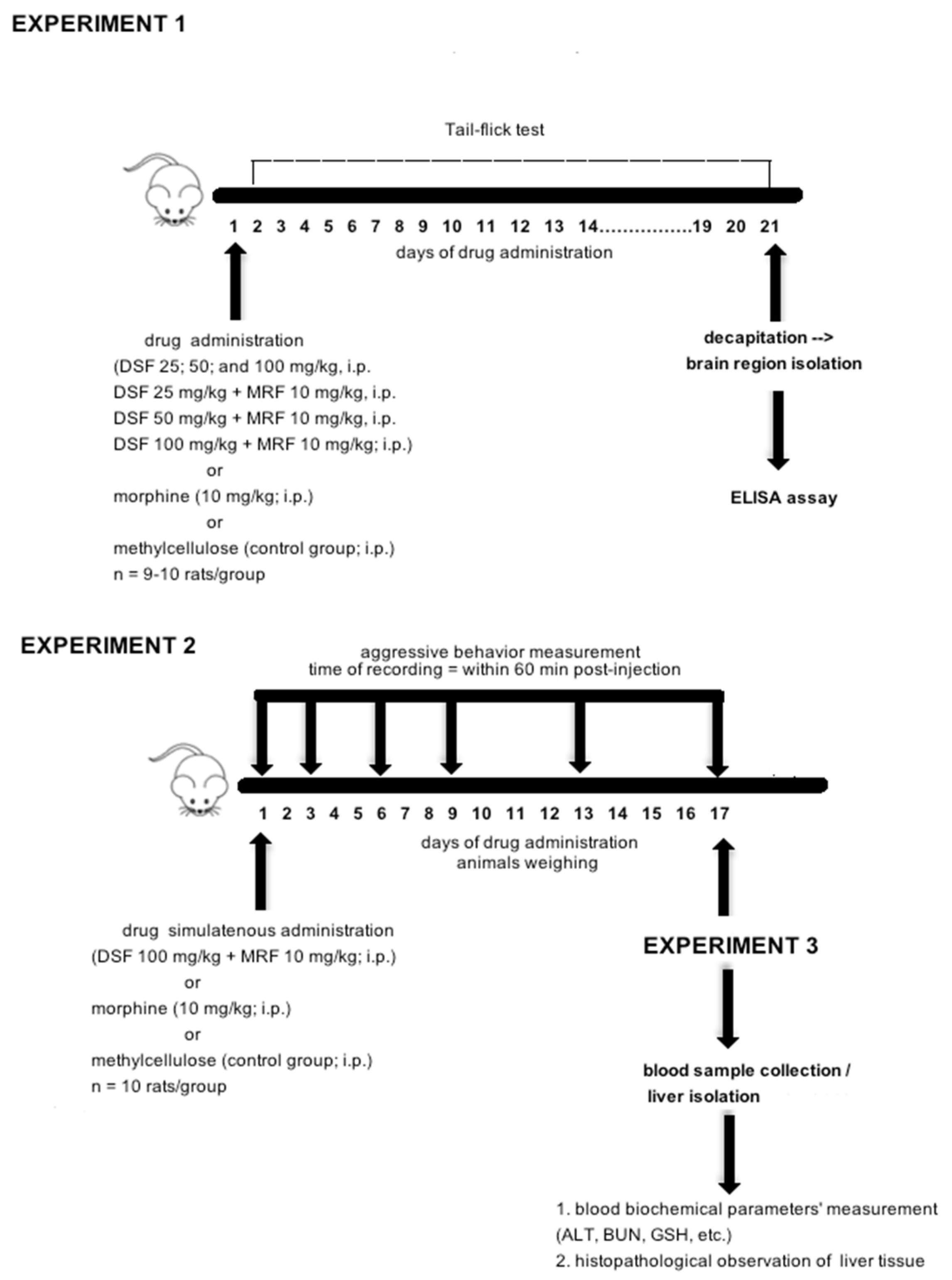
2.3. Experimental Design and Procedure Used
2.3.1. Experiment 1: Effect of Repeated Simultaneous Administration of Disulfiram and Morphine on Thermally Evoked Pain and Biochemical Markers of Morphine-Induced Tolerance
Tail Flick Test
Enzyme-Linked Immunosorbent Assay (ELISA)
Tissue Preparation
2.3.2. Experiment 2: Effect of Disulfiram and Morphine Subacute Co-Administration on Aggressive Behavior
2.3.3. Experiment 3: Examination of DSF-Induced Toxicity
Colorimetric Analysis
Histopathological Studies
2.4. Statistical Analysis
3. Results
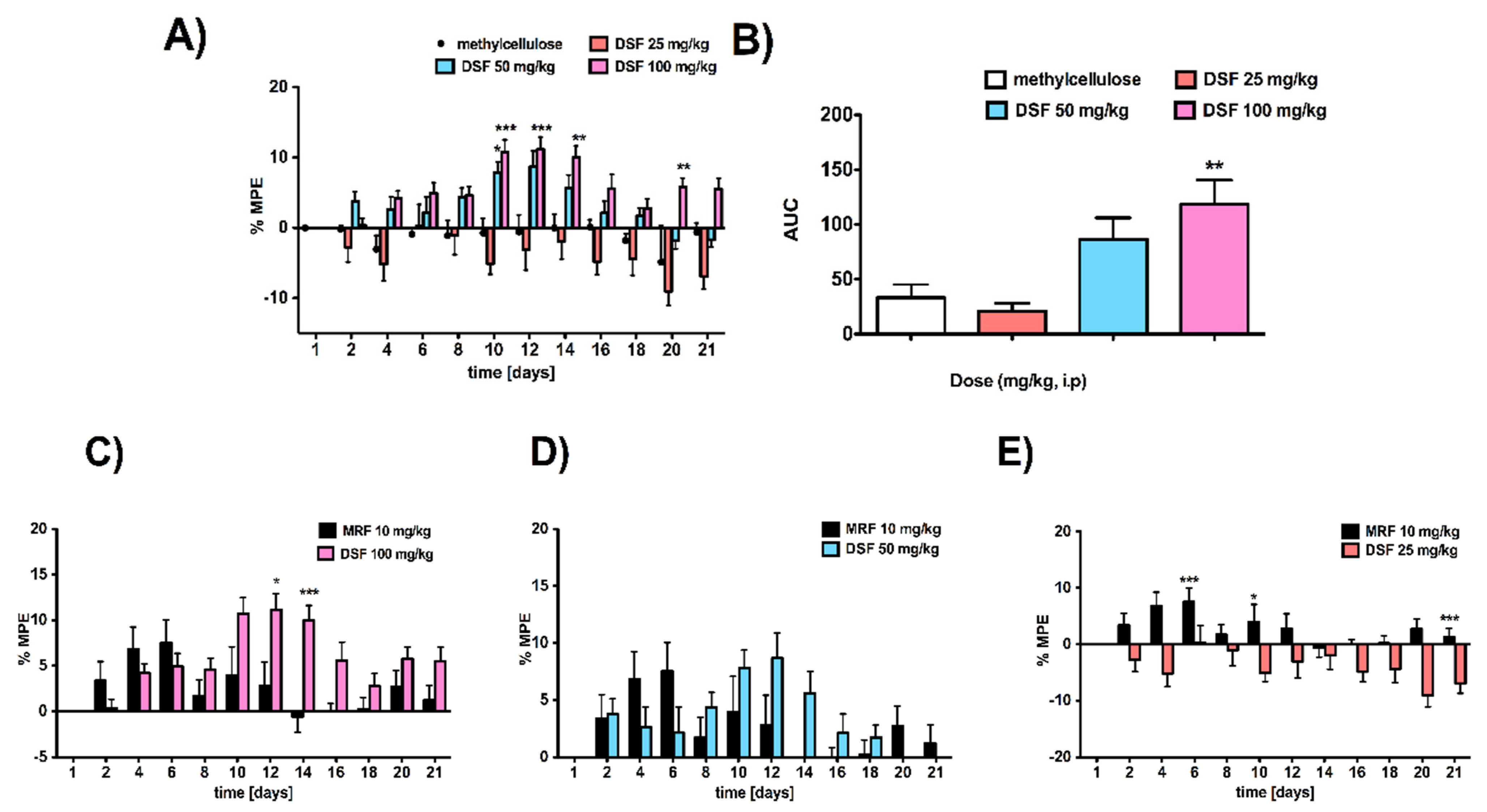
3.1. Rats Subacutely Exposed to an Escalating Dose of Disulfiram Alone Were Less Sensitive to Thermally Induced Pain Compared to Vehicle-Treated Animals
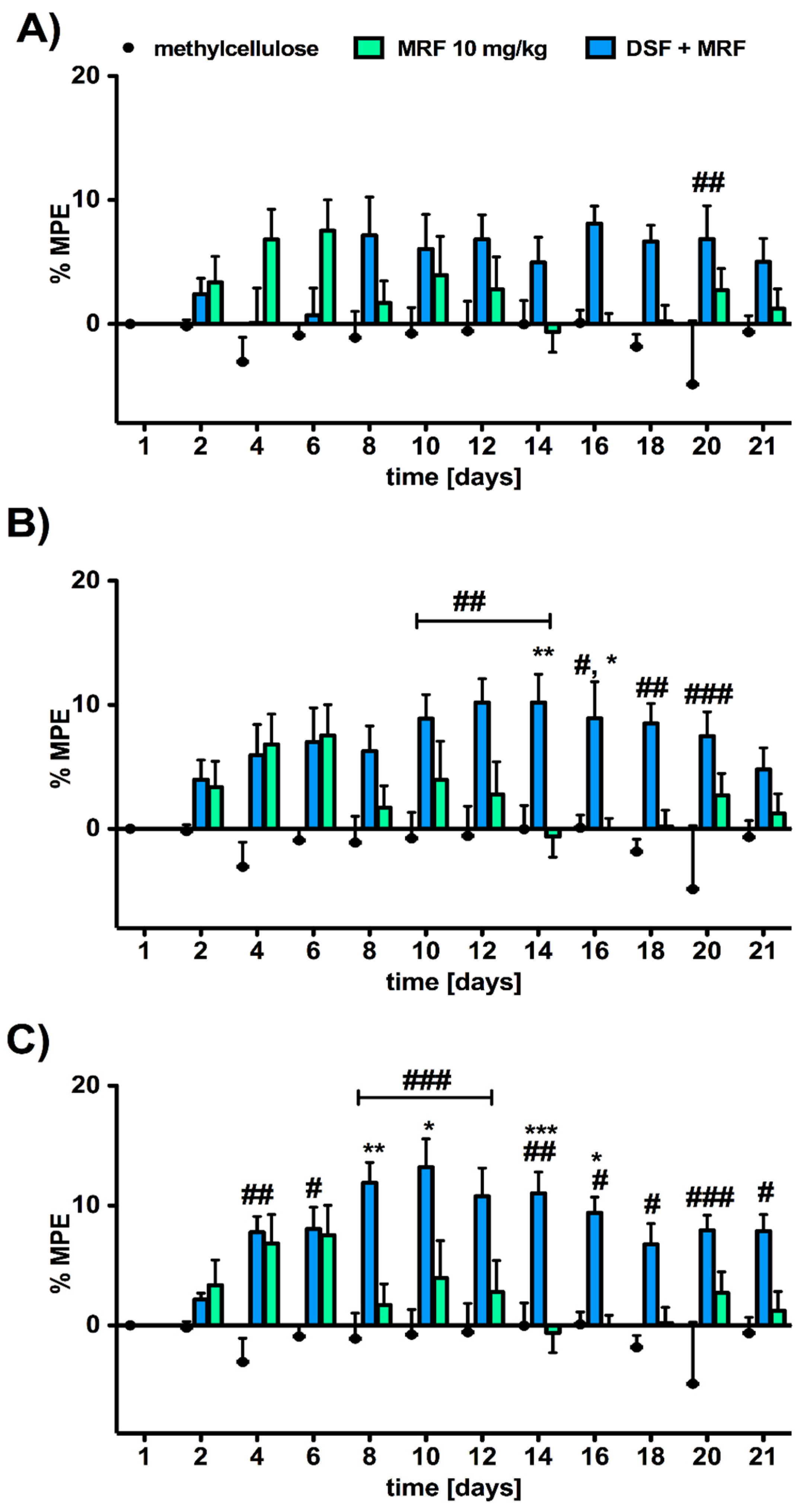
3.2. Analgesic Response to Morphine Was Affected by the Addition of Disulfiram in a Dose-Dependent Manner
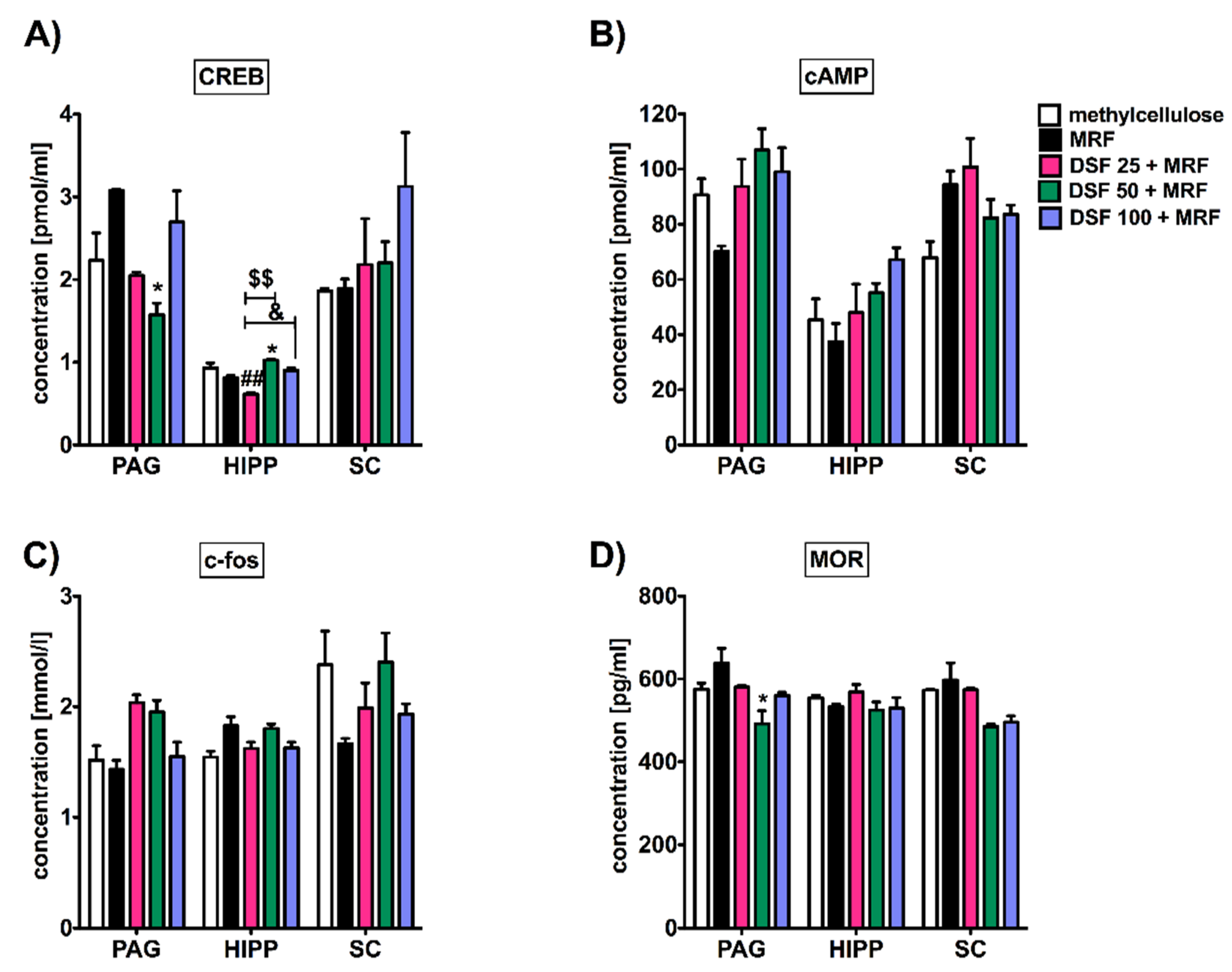
3.3. Subacute Administration of Disulfiram and Morphine Did Not Alter c-fos, cAMP, and Mu-Opioid Receptor Levels except Transcription Factor (CREB) Protein in the Periaqueductal Gray (PAG), Hippocampus, and Spinal Cord
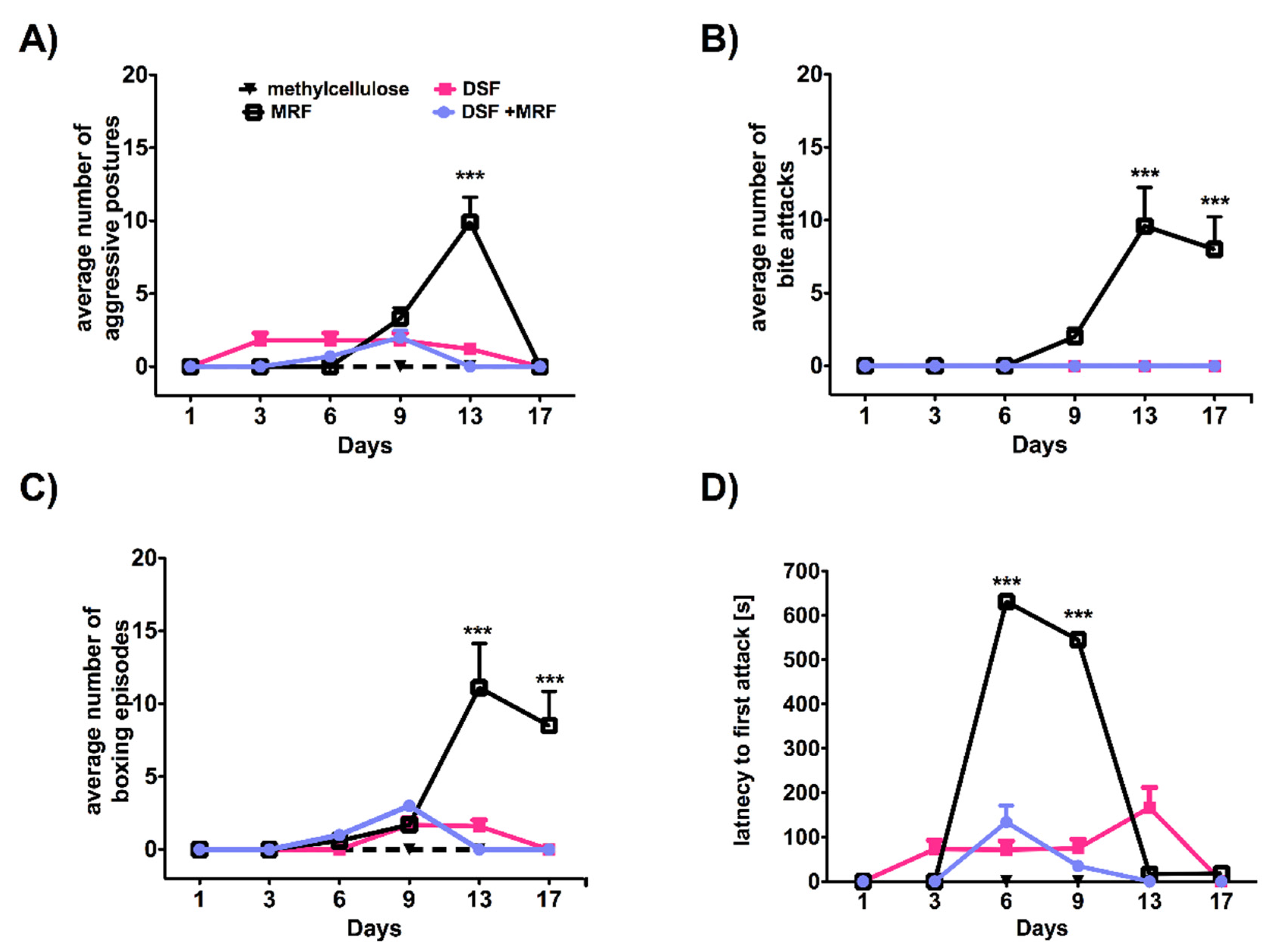
3.4. Repeated Simultaneous Administration of Disulfiram and Morphine at the Highest Dose Tested Affected Morphine-Induced Aggressive Response in Rats
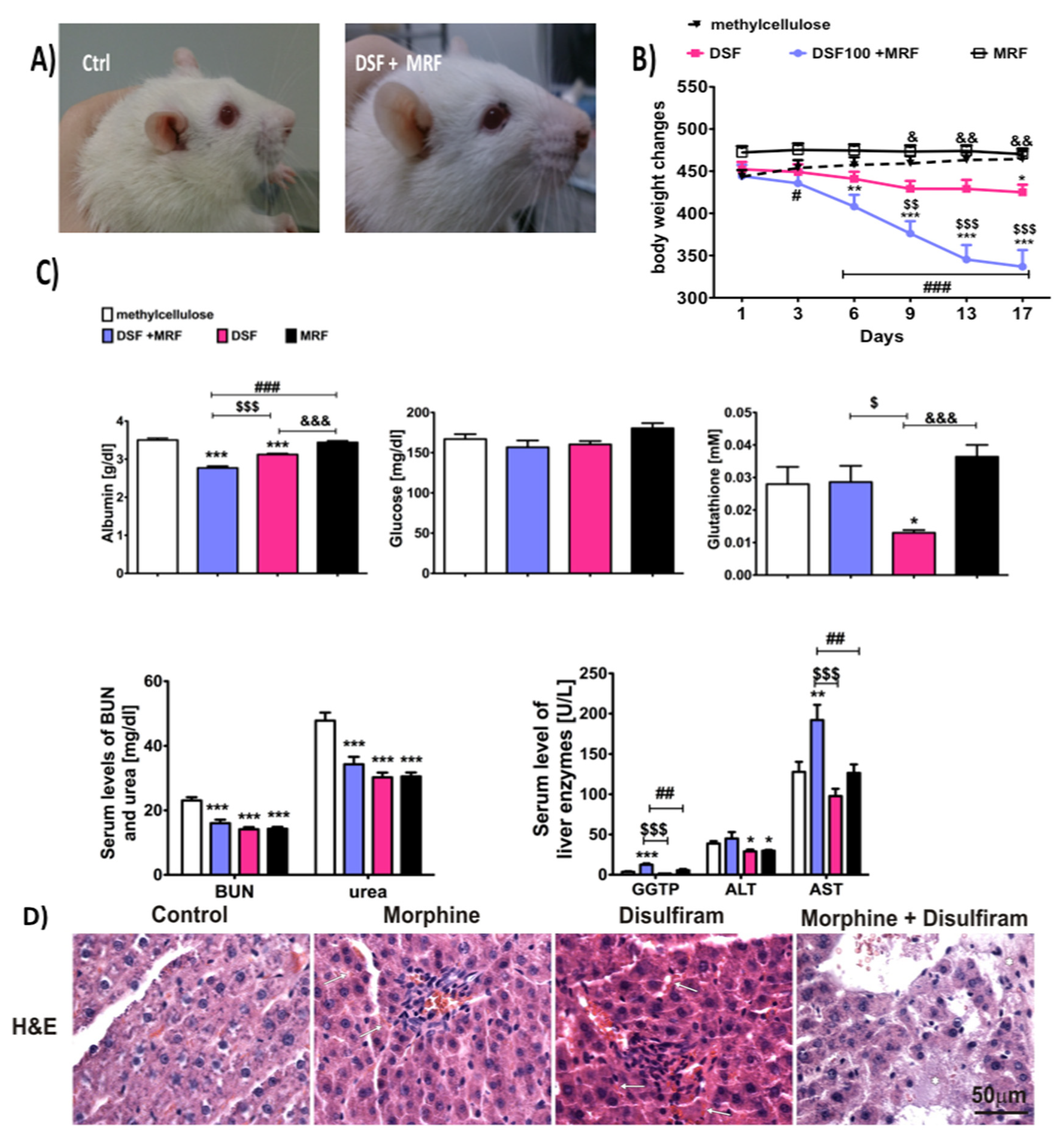
3.5. Morphine and Disulfiram Co-Administration Significantly Augmented Disulfiram-Induced Hepatic Failure
4. Discussion
4.1. Disulfiram Pain-Relieving Effect and Its Impact on Analgesia and Morphine Tolerance Development
4.2. Disulfiram Impact on Morphine-Induced Tolerance Markers
4.3. Disulfiram Impact on Morphine-Induced Aggressive Response
4.4. The Concomitant Therapy and Disulfiram-Induced Hepatic Failure
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whelan, P.J.; Remski, K. Buprenorphine vs. methadone treatment: A review of evidence in both developed and developing worlds. J. Neurosci. Rural. Pract. 2012, 3, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Amador, E.; Gazdar, A. Sudden death during disulfiram–alcohol reaction. Q. J. Stud. Alcohol 1967, 28, 649–654. [Google Scholar] [CrossRef]
- Gessner, P.K.; Gessner, T. Relevant physical and chemical properties. In Disulfiram and Its Metabolite, Diethyldithio Carbamate; Genner, P.K., Gedddner, T., Eds.; Springer Science and Business Media: Cornwall, UK, 1992; pp. 7–9. [Google Scholar]
- Gaval-Cruz, M.; Weinshenker, D. Mechanism of disulfiram-induced cocaine abstinence: Antabuse and cocaine relaps. Mol. Interv. 2009, 9, 175–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, K.; Ziedonis, D.; O’Malley, S.; McCance-Katez, E.; Gordon, L.; Rounsaville, B. Pharmacologic interventions for alcohol-and-cocaine-abusing individuals. Am. J. Addict. 1993, 2, 77–79. [Google Scholar] [CrossRef]
- Grassi, M.C.; Cioce, A.M.; Giudici, F.D.; Antonilli, L.; Nencini, P. Short-term efficacy of disulfiram or naltrexone in reducing positive urinanalysis for both cocaine and cocaethylene in cocaine abusers: A pilot study. Pharmacol. Res. 2007, 55, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Maj, J.; Przegalinski, E.; Wielosz, M. Disulfiram and the drug-induced effects on motility. J. Pharm. Pharmacol. 1968, 20, 247–248. [Google Scholar] [CrossRef]
- Nich, C.; McCance-Katz, E.F.; Petrakis, I.L.; Cubells, J.F.; Rounsaville, B.J.; Carroll, K.M. Sex differences in cocaine-dependent individuals‘ response to disulfiram treatment. Addict. Behav. 2004, 29, 1123–1128. [Google Scholar] [CrossRef] [Green Version]
- de Corde, A.; Krząścik, P.; Wolińska, R.; Kleczkowska, P.; Filip, M.; Bujalska-Zadrożny, M. Disulfiram attenuates morphine or methadone withdrawal syndrome in mice. Behav. Pharmacol. 2018, 25, 393–399. [Google Scholar] [CrossRef]
- Reuter, J.; Raedler, T.; Rose, M.; Hand, I.; Glasher, J.; Buchel, C. Pathological gambling is linked to reduced activation of the mesolimbic reward system. Nat. Neurosci. 2005, 8, 147–148. [Google Scholar] [CrossRef]
- Lacoursiere, R.B.; Swatek, R. Adverse interaction between disulfiram and marijuana: A case report. Am. J. Psychiatry 1983, 140, 242–244. [Google Scholar]
- Nestler, E.J. Common molecular and cellular substrates of addiction and memory. Neurobiol. Learn. Mem. 2002, 78, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.T.; Christie, M.J.; Manzoni, O. Cellular and synaptic adaptations mediating opioid dependence. Physiol. Rev. 2001, 81, 299–343. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Pu, X.P. Chronic morphine administration induces over-expression of aldolase C with reduction of CREB phosphorylation in the mouse hippocampus. Eur. J. Pharmacol. 2009, 609, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Major, L.F.; Lerner, P.; Ballenger, J.C.; Brown, G.L.; Goodwin, F.K.; Lovenberg, W. Dopamine-beta-hydroxylase in the cerebrospinal fluid: Relationship to disulfiram-induced psychosis. Biol. Psychiatry 1979, 14, 337–344. [Google Scholar] [PubMed]
- Paradisi, R.; Grossi, G.; Pintore, A.; Venturoli, S.; Porcu, E.; Capelli, M. Evidence for a pathological reduction in brain dopamine metabolism in idiopathic hyperprolactinemia. Acta Endocrinol. 1991, 125, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.P.; Cooper, D.A.; Schank, J.R.; Lyle, M.A.; Gaval-Cruz, M.; Ogbonmwan, Y.E. Disulfiram attenuates drug-primed reinstatement of cocaine seeking via inhibition of dopamine β-hydroxylase. Neuropsychopharmacol 2010, 35, 2440–2449. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Grunberg, N.E.; Mueller, G.P. Disulfiram causes sustained behavioral and biochemical effects in rats. Pharmacol. Biochem. Behav. 1997, 56, 409–415. [Google Scholar] [CrossRef]
- Tran, A.T.; Rison, R.A.; Beydoun, S.R. Disulfiram neuroapthy: Two case reports. J. Med. Case Rep. 2016, 10, 72. [Google Scholar] [CrossRef] [Green Version]
- Bradley, W.G.; Hewer, R.L. Peripheral neuropathy due to disulfiram. Br. Med. J. 1966, 2, 449–450. [Google Scholar] [CrossRef] [Green Version]
- Gardner-Thorpe, C.; Benjamin, S. Peripheral neuropathy after disulfiram administration. J. Neurol. Neurosurg. Psychiat. 1971, 34, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchand, J.E.; Hershman, K.; Kumar, M.S.A.; Tjompson, M.L.; Kream, R.M. Disulfiram administration affects substance P-like immunoreactive and monoaminergic neural systems in rodent brain. J. Biol. Chem. 1990, 265, 264–273. [Google Scholar] [PubMed]
- Bilecki, W.; Przewłocki, R. Effect of opioids on Ca2+/cAMP responsive element binding protein. Acta Neurobiol. Exp. Wars 2000, 60, 557–567. [Google Scholar] [PubMed]
- Center for Substance Abuse Treatment. Incorporating Alcohol Pharmacotherapies Into Medical Practice. Treatment Improvement Protocol (TIP) Series 49. HHS Publication No. (SMA) 09-4380; Substance Abuse and Mental Health Services Administration: Rockville, MD, USA, 2009. [Google Scholar]
- Berman, M.; Taylor, S.; Merged, B. Morphine and human aggression. Addict. Behav. 1993, 18, 263–268. [Google Scholar] [CrossRef]
- Kantak, K.; Miczek, K. Social, motor, and autonomic signs of morphine withdrawal; differential sensitivities to catecholaminergic drugs. Psychopharmacology 1998, 96, 468–476. [Google Scholar] [CrossRef]
- Kanui, T.I.; Hole, K. Morphine induces aggression but not analgesia in the naked mole-rat (Heterocephalus glaber). Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1990, 96, 131–133. [Google Scholar] [CrossRef]
- Sukhotina, I.A. Morphine withdrawal-facilitated aggression is attenuatedby morphine-conditioned stimuli. Pharmacol. Biochem. Behav. 2001, 68, 93–98. [Google Scholar] [CrossRef]
- Garrat, M.; Brooks, R.C. A genetic reduction in antioxidant function causes elevated aggression in mice. J. Exp. Biol. 2015, 218 (Pt. 2), 223–227. [Google Scholar] [CrossRef] [Green Version]
- Constantini, D.; Carere, C.; Caramaschi, D.; Koolhaas, J.M. Aggressive and non-aggressive personalities differ in oxidative status in selected lines of mice (Mus musculus). Biol. Lett. 2008, 4, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Dvoto, P.; Flore, G.; Saba, P.; Cadeddu, R.; Gessa, G.L. Disulfiram stimulates dopamine release from noradrenergic terminals and potentiates cocaine-induced dopamine release in the prefrontal cortex. Psychopharmacology 2012, 219, 1153–1164. [Google Scholar] [CrossRef]
- Sheel-Kruger, J.; Randrup, A. Stereotyped hyperactivity behavior produced by dopamine in the absence of noradrenaline. Life Sci. 1967, 6, 1389. [Google Scholar] [CrossRef]
- Reis, D.J. The relationship between brain norepinephrine and aggressive behavior. Res. Publ. Ass. Res. Nerv. Ment. Dis. 1972, 50, 266. [Google Scholar] [PubMed]
- Rabkin, J.M.; Corless, C.L.; Orloff, S.L.; Benner, K.G.; Flora, K.D.; Rosen, H.R.; Olyaei, A.J. Liver transplantation for disulfiram-induced hepatic failure. Am. J. Gastroenterol. 1998, 93, 830–831. [Google Scholar] [CrossRef] [PubMed]
- Watts, T.E.; Pandey, R.A.; Vancil, T.J. Fatal fulminant hepatic failure related to the use of disulfiram. J. Ark. Med. Soc. 2014, 110, 280–283. [Google Scholar]
- Nagendra, S.N.; Shetty, K.T.; Rao, K.M.; Rao, B.S. Effect of disulfiram administration on rat brain glutathione metabolism. Alcohol 1994, 11, 7–10. [Google Scholar] [CrossRef]
- Ohno, Y.; Hirota, K.; Kawanishi, T.; Takanaka, A. Loss of viability after disulfiram treatment without preceding depletion of intracellular GSH. J. Toxicol. Sci. 1990, 15, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Dong, H.; Thompson, D.C.; Shertzer, H.G.; Nebert, D.W.; Vasiliou, V. Glutathione defense mechanism in liver injury: Insights from animal models. Food Chem. Toxicol. 2013, 60, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Swiderska-Kołacz, G.; Parka, B.; Kołątaj, A.; Klusek, J. Wpływ morfiny i skopolaminy na zawartość glutationu i aktywność enzymów glutationowych w tkankach myszy. Med. Wet. 2004, 60, 793. [Google Scholar]
- Kong, A.N.; Owuor, E.; Yu, R.; Hebbar, V.; Chen, C.; Hu, R. Induction of xenobiotic enzymes by the MAP kinase pathway and the antioxidant or electrophile response element (ARE/EpRE). Drug Metab. Rev. 2001, 33, 255–271. [Google Scholar] [CrossRef]
- Atici, S.; Cinel, I.; Cinel, L.; Doruk, N.; Eskandrai, G.; Oral, U. Liver and kidney toxicity in chronic use of opioids: An experimental long term treatment model. J. Biosci. 2005, 30, 245–252. [Google Scholar] [CrossRef]
- Jalili, C.; Makalani, F.; Roshankhah, S.; Sohrabi, K.; Salahshoor, M.R. Protective effect of resveratrol against morphine damage to kidneys of mice. Int. J. Morphol. 2017, 35, 1409–1415. [Google Scholar] [CrossRef] [Green Version]
- Khairnar, S.I.; Mahajan, U.B.; Patil, K.R.; Patel, H.M.; Shinde, S.D.; Goyal, S.N.; Belemkar, S.; Ojha, S.; Patil, C.R. Disulfiram and it copper chetlate attenuate cisplatin-induced acute nephrotoxicity in rats via reduction of oxidative stress and inflammation. Biol. Trace Elem. Res. 2020, 193, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Chahkandi, M.; Askari, N.; Asadikaram, G. The effect of acute and chronic morphine on some blood biochemical parameters in an inflammation condition in gonadectomized male rats. Addict. Health 2015, 7, 130–139. [Google Scholar] [PubMed]
- Nagi, N.; Murao, T.; Okamoto, N.; Ito, Y. Disulfiram reduces elevated blood glucose levels in Otsuka Long-Evans Tokushima Fatty (OLETF) rats, a model of type 2 diabetes. J. Oleo Sci. 2009, 58, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagamatsu, K.; Ohno, Y.; Ikebuchi, H.; Takahashi, A.; Terao, T.; Takanaka, A. Morphine metabolism in isolated rat hepatocytes and its implications for hepatotoxicity. Biochem. Pharmacol. 1986, 35, 3543–3548. [Google Scholar] [CrossRef]
- Skoulis, N.P.; James, R.C.; Harbison, R.D.; Roberts, S.M. Depression of hepatic glutathione by opioid analgesic drugs in mice. Toxicol. Appl. Pharmacol. 1989, 99, 139–147. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frączek, K.; Kowalczyk, A.; Pekala, M.; Kasarello, K.; Sygitowicz, G.; Sulejczak, D.; Zaremba, M.; Konop, M.; Frankowska, M.; Filip, M.; et al. The Positive and Negative Outcome of Morphine and Disulfiram Subacute Co-Administration in Rats in the Absence of Ethanol Challenge. Pharmaceutics 2021, 13, 29. https://doi.org/10.3390/pharmaceutics13010029
Frączek K, Kowalczyk A, Pekala M, Kasarello K, Sygitowicz G, Sulejczak D, Zaremba M, Konop M, Frankowska M, Filip M, et al. The Positive and Negative Outcome of Morphine and Disulfiram Subacute Co-Administration in Rats in the Absence of Ethanol Challenge. Pharmaceutics. 2021; 13(1):29. https://doi.org/10.3390/pharmaceutics13010029
Chicago/Turabian StyleFrączek, Karolina, Agnieszka Kowalczyk, Martyna Pekala, Kaja Kasarello, Grażyna Sygitowicz, Dorota Sulejczak, Malgorzata Zaremba, Marek Konop, Malgorzata Frankowska, Malgorzata Filip, and et al. 2021. "The Positive and Negative Outcome of Morphine and Disulfiram Subacute Co-Administration in Rats in the Absence of Ethanol Challenge" Pharmaceutics 13, no. 1: 29. https://doi.org/10.3390/pharmaceutics13010029