Non-Viral Targeted Nucleic Acid Delivery: Apply Sequences for Optimization


Abstract
:1. Introduction
2. Sequence-Defined Macromolecular Carriers
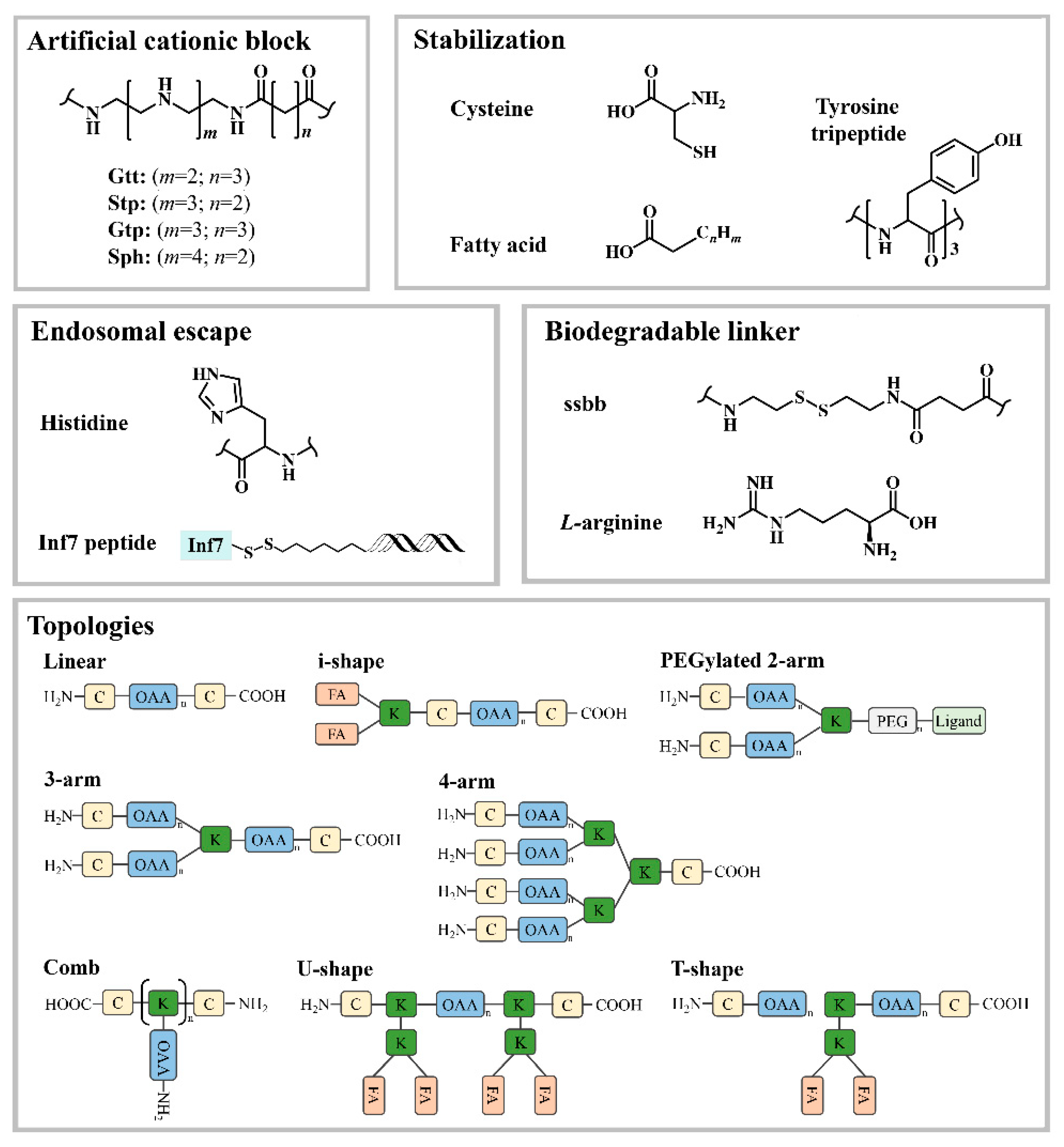
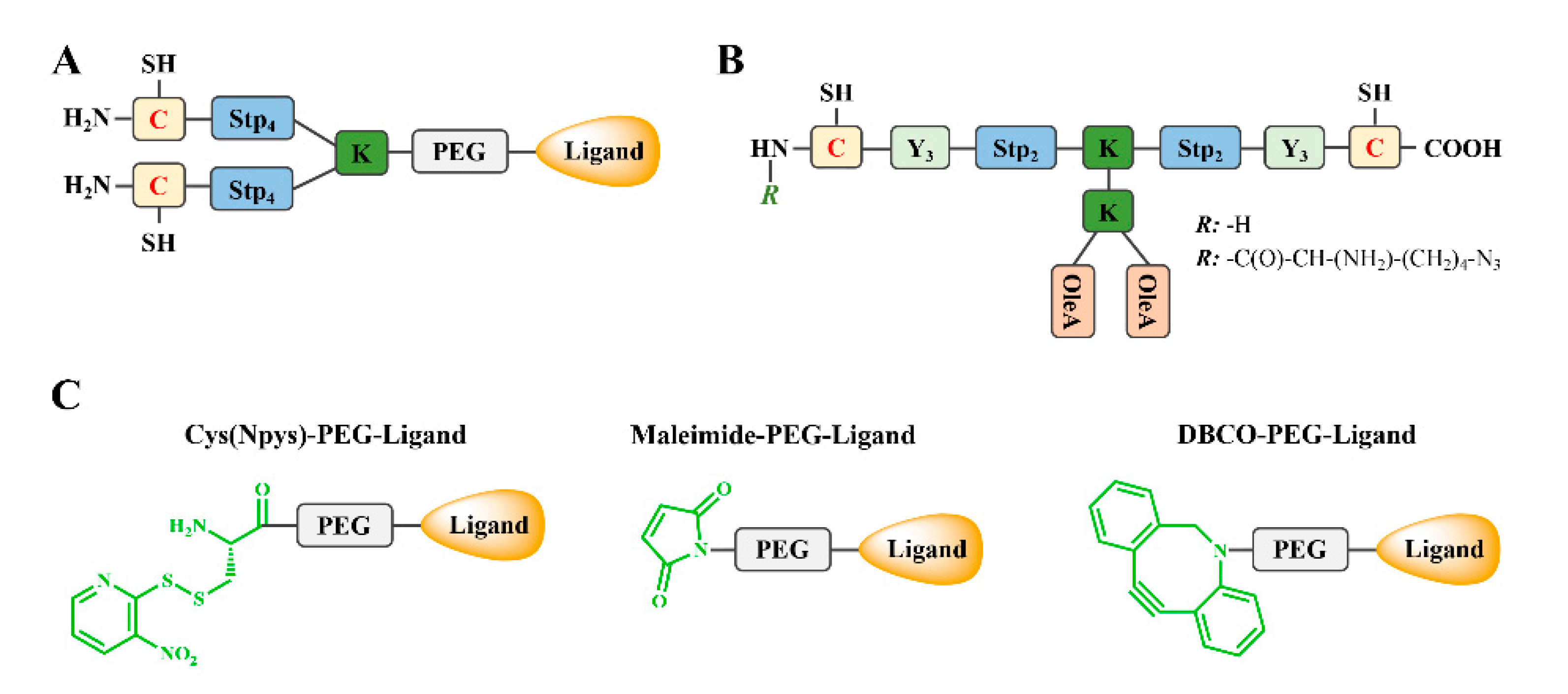
2.1. Lysine-Based Sequence-Defined Peptides
2.2. Cell-Penetrating Peptides (CPPs)
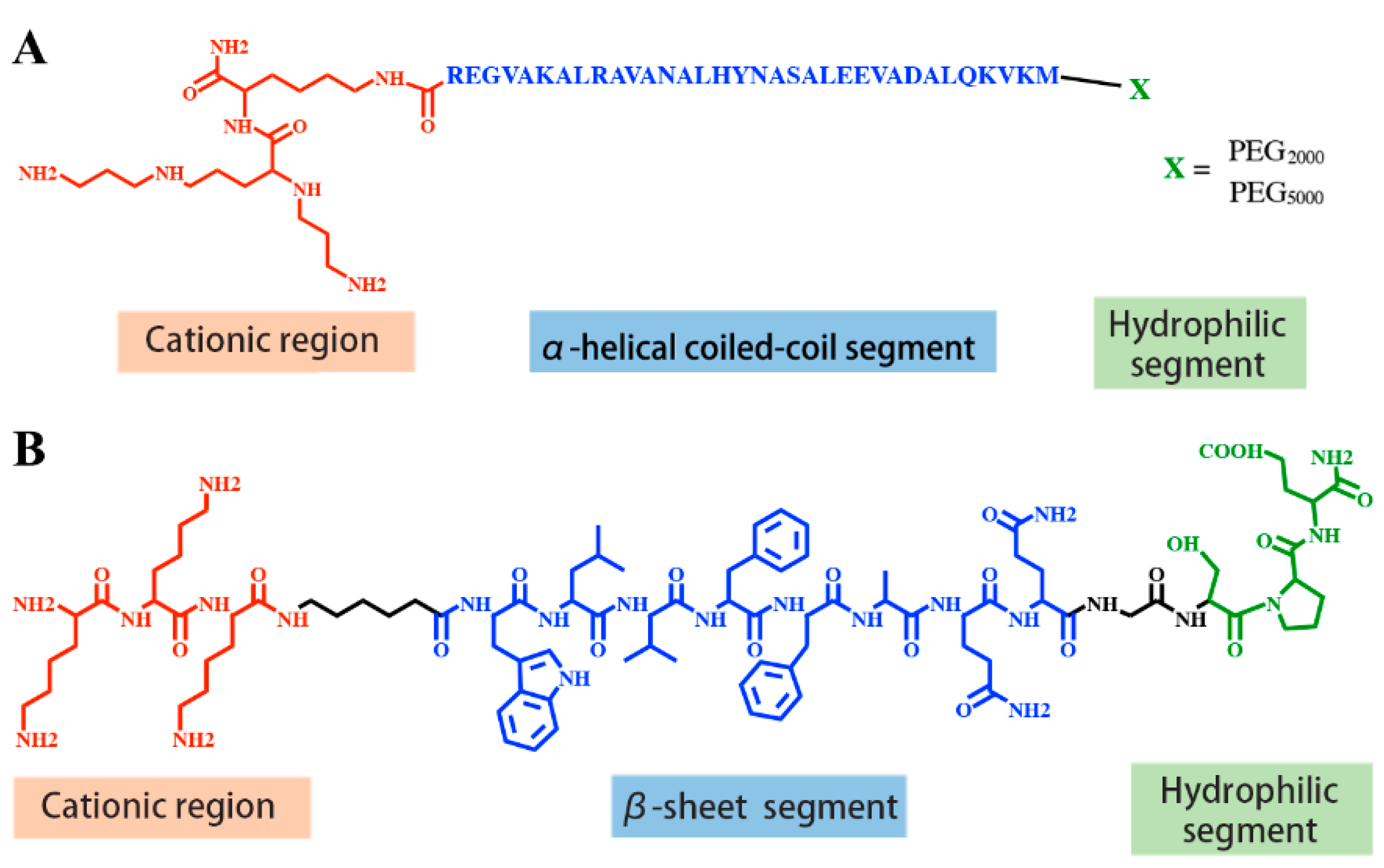
2.3. Peptides with Supramolecular Self-Assembly Domains
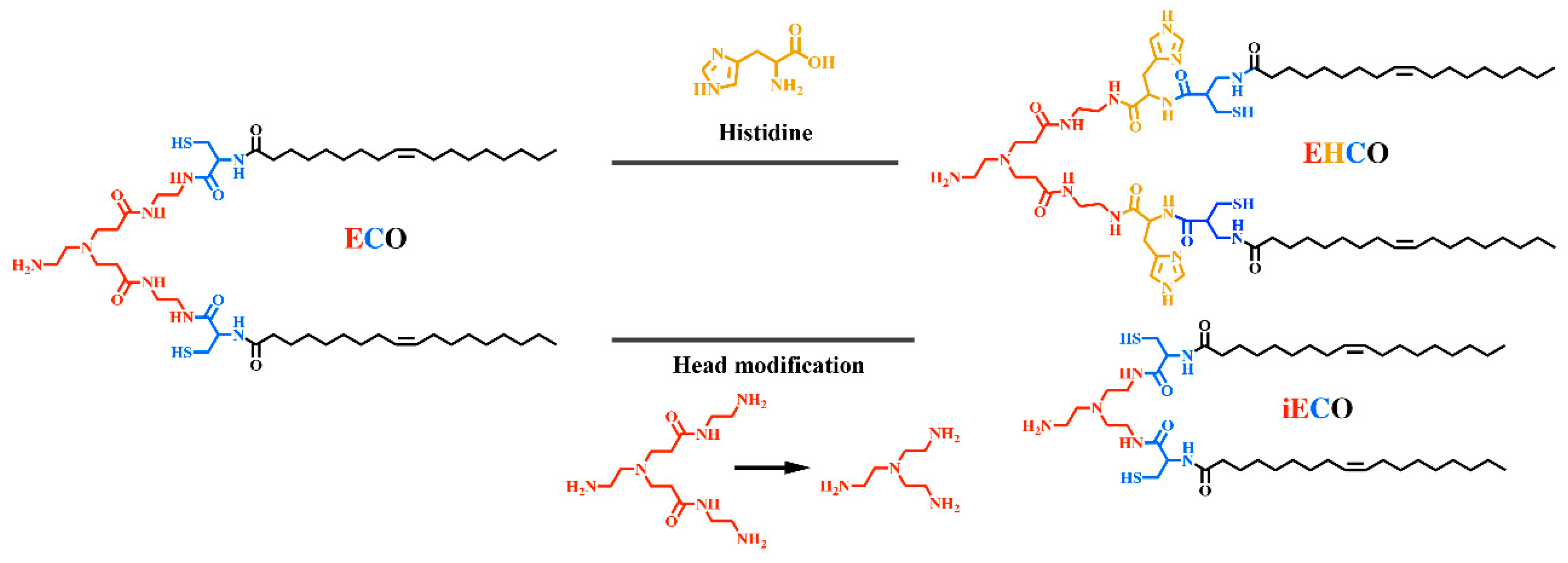
2.4. Lipo-Peptides
2.5. Oligoaminoamines (OAAs)
3. Optimizing Carriers for Different Types of Therapeutic Nucleic Acids
3.1. Stable Nanoparticle Formation Is Important for siRNA Carriers
3.2. Other Nucleic Acid Cargos Including Cas9/sgRNA or PMO
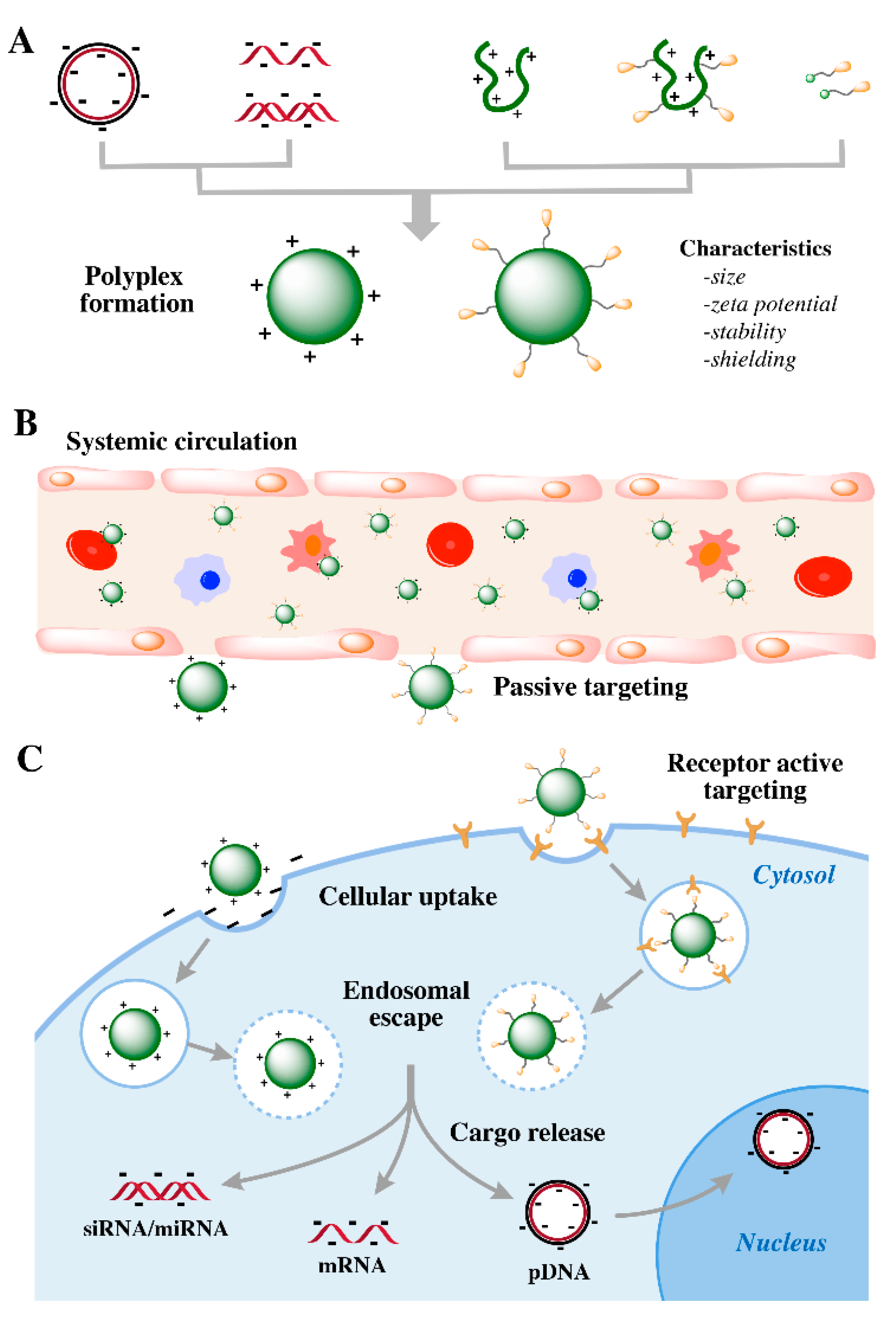
4. Pharmacological Barriers for In Vivo Delivery
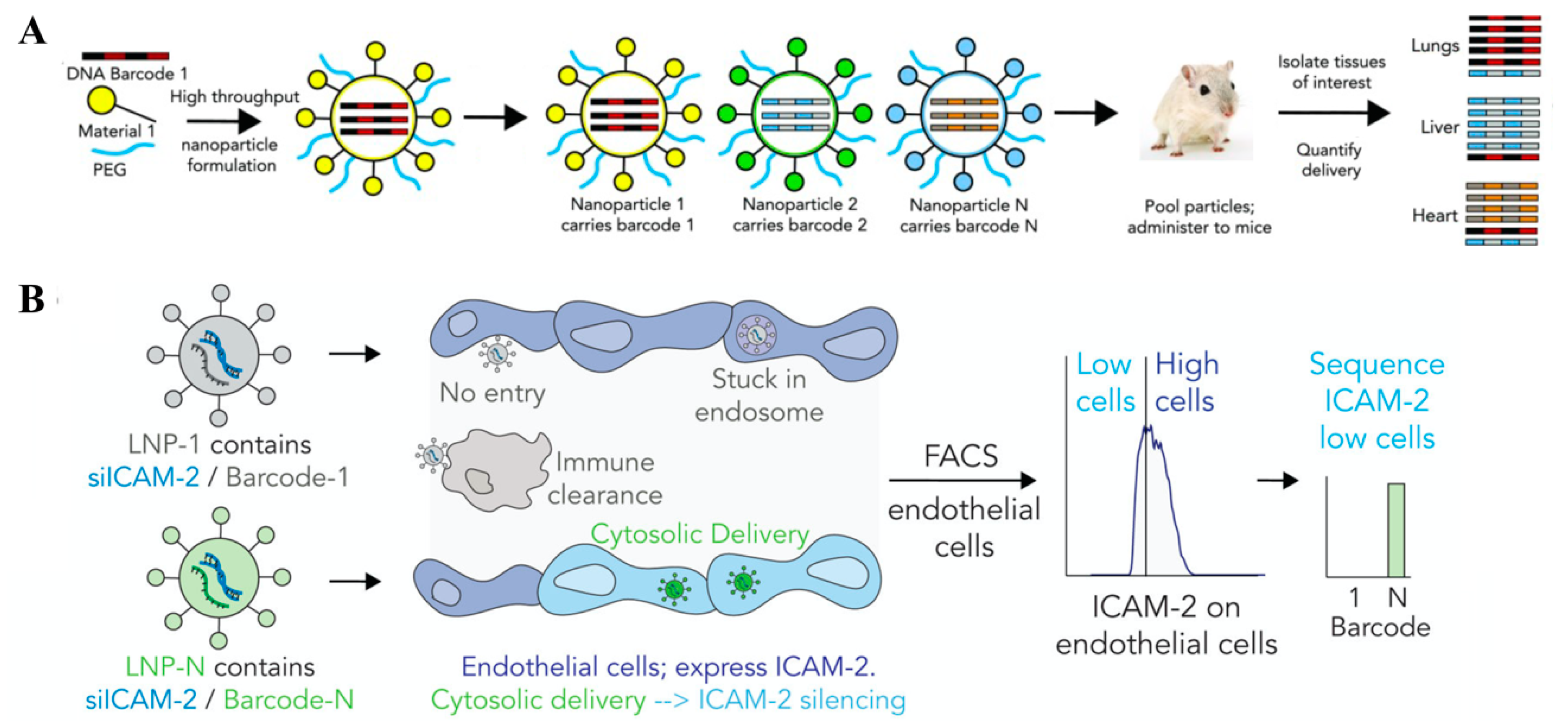
5. Barcoding—A New Mode to Apply Sequences for Finding In Vivo Nanocarriers
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ASGPR | Asialoglycoprotein receptor |
| ASOs | Antisense oligonucleotides |
| Cas9 | CRISPR-associated protein 9 |
| cLNPs | cLNP constrained lipid nanoparticles |
| cMBP2 | c-Met-binding peptide |
| CPPs | cell-penetrating peptides |
| CRC | Cysteine–arginine–cysteine |
| DBCO | Dibenzocyclooctyne amine |
| ddPCR | digital droplet PCR |
| DSB | double-strand break |
| DTNB | 5,5′-Dithio-bis(2-nitrobenzoic acid) |
| ECO | 1-aminoethyl)iminobis[N-(oleicylcysteinyl-1-amino-ethyl)propionamide] |
| EG5 | Eglin-5 |
| EHCO | (1-aminoethyl) imino bis [N-(oleoyl cysteinyl histinyl-1-aminoethyl) propionamide] |
| EGFR | Epidermal growth factor receptor |
| EPR | Enhanced permeation and retention |
| ESC | Enhanced stabilization chemistry |
| Fmoc | Fluorenyl methoxycarbonyl |
| FolA | Folic acid |
| FR | Folate receptor |
| FRET | Fluorescence resonance energy transfer |
| GSH | Glutathione |
| INF7 | glutamic acid-enriched analogue of the influenza hemagglutinin membrane protein HA2 |
| LNPs | Lipid nanoparticles |
| LPEI | linear PEI |
| mRNA | Messager RNA |
| MTX | Methotrexate |
| NIS | Sodium iodide symporter |
| NLS | Nuclear localization signal |
| NPC | Nuclear pore complex |
| OAAs | Oligoaminoamides |
| OPSS | ortho-pyridyl disulfide |
| PAs | Peptide amphiphiles |
| pDNA | Plasmid DNA |
| PEG | Polyethylene glycol |
| PEI | Polyethylenimine |
| pHPMA | poly(N-(2-hydroxypropyl)methacrylamide) |
| pING4 | pDNA encoding inhibitor of growth 4 |
| PMOs | Phosphorodiamidate morpholino oligonucleotides |
| PT | Pretubulysin |
| RAFT | Reversible addition-fragmentation chain transfer |
| RISC | RNA-induced silencing complex |
| RNAi | RNA interference |
| RNP | Ribonucleoprotein |
| SCOs | splice-correcting oligonucleotides |
| sgRNA | single guide RNA |
| siRNA | Small interfering RNA |
| Stp | Succinoyltetraethylene-pentamine |
| Sph | Succinoyl-pentaethylene hexamine |
| SPS | Solid phase supported synthesis |
| SSBB | Disulfide building block = succinoyl-cystamine |
| tBoc | t-butyloxycarbonyl |
| TCPs | Targeted combinatorial polyplexes |
| TLPs | Targeted lipopolyplexes |
| TNBC | triple-negative breast cancer |
| Tf | Transferrin |
| TfR | Transferrin receptor |
| TTR | Transthyretin |
References
- Davis, M.E.; Zuckerman, J.E.; Choi, C.H.J.; Seligson, D.; Tolcher, A.; Alabi, C.A.; Yen, Y.; Heidel, J.D.; Ribas, A. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nature 2010, 464, 1067–1070. [Google Scholar] [CrossRef] [PubMed]
- Tabernero, J.; Shapiro, G.I.; LoRusso, P.M.; Cervantes, A.; Schwartz, G.K.; Weiss, G.J.; Paz-Ares, L.; Cho, D.C.; Infante, J.R.; Alsina, M. First-in-humans trial of an RNA interference therapeutic targeting VEGF and KSP in cancer patients with liver involvement. Cancer Discov. 2013, 3, 406–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.D.; Yang, C.-C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L. Patisiran, an RNAi therapeutic, for hereditary transthyretin amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef]
- Chen, Y.; Gao, D.-Y.; Huang, L. In vivo delivery of miRNAs for cancer therapy: Challenges and strategies. Adv. Drug Deliv. Rev. 2015, 81, 128–141. [Google Scholar] [CrossRef] [Green Version]
- Havens, M.A.; Hastings, M.L. Splice-switching antisense oligonucleotides as therapeutic drugs. Nucleic Acids Res. 2016, 44, 6549–6563. [Google Scholar] [CrossRef]
- Lu, Q.L.; Mann, C.J.; Lou, F.; Bou-Gharios, G.; Morris, G.E.; Xue, S.-A.; Fletcher, S.; Partridge, T.A.; Wilton, S.D. Functional amounts of dystrophin produced by skipping the mutated exon in the mdx dystrophic mouse. Nat. Med. 2003, 9, 1009–1014. [Google Scholar] [CrossRef]
- Ottesen, E.W. ISS-N1 makes the first FDA-approved drug for spinal muscular atrophy. Transl. Neurosci. 2017, 8, 1–6. [Google Scholar] [CrossRef]
- Levin, A.A. Treating disease at the RNA level with oligonucleotides. N. Engl. J. Med. 2019, 380, 57–70. [Google Scholar] [CrossRef]
- Yin, H.; Song, C.-Q.; Dorkin, J.R.; Zhu, L.J.; Li, Y.; Wu, Q.; Park, A.; Yang, J.; Suresh, S.; Bizhanova, A. Therapeutic genome editing by combined viral and non-viral delivery of CRISPR system components in vivo. Nat. Biotechnol. 2016, 34, 328–333. [Google Scholar] [CrossRef]
- Ginn, S.L.; Amaya, A.K.; Alexander, I.E.; Edelstein, M.; Abedi, M.R. Gene therapy clinical trials worldwide to 2017: An update. J. Gene Med. 2018, 20, e3015. [Google Scholar] [CrossRef]
- Durymanov, M.; Reineke, J. Non-viral delivery of nucleic acids: Insight into mechanisms of overcoming intracellular barriers. Front. Pharmacol. 2018, 9, 971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lächelt, U.; Wagner, E. Nucleic acid therapeutics using polyplexes: A journey of 50 years (and beyond). Chem. Rev. 2015, 115, 11043–11078. [Google Scholar] [CrossRef] [PubMed]
- Hager, S.; Wagner, E. Bioresponsive polyplexes–chemically programmed for nucleic acid delivery. Expert Opin. Drug Deliv. 2018, 15, 1067–1083. [Google Scholar] [CrossRef] [PubMed]
- Felgner, P.; Barenholz, Y.; Behr, J.; Cheng, S.; Cullis, P.; Huang, L.; Jessee, J.; Seymour, L.; Szoka, F.; Thierry, A. Nomenclature for synthetic gene delivery systems. Hum. Gene Ther. 1997, 8, 511–512. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Liang, H.; Jin, R.; Nie, Y. Virus-inspired and mimetic designs in non-viral gene delivery. J. Gene Med. 2019, 21, e3090. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Miao, L.; Satterlee, A.; Huang, L. Delivery of oligonucleotides with lipid nanoparticles. Adv. Drug Deliv. Rev. 2015, 87, 68–80. [Google Scholar] [CrossRef] [Green Version]
- Cabral, H.; Miyata, K.; Osada, K.; Kataoka, K. Block copolymer micelles in nanomedicine applications. Chem. Rev. 2018, 118, 6844–6892. [Google Scholar] [CrossRef] [Green Version]
- Bono, N.; Ponti, F.; Mantovani, D.; Candiani, G. Non-Viral in Vitro Gene Delivery: It is Now Time to Set the Bar! Pharmaceutics 2020, 12, 183. [Google Scholar] [CrossRef] [Green Version]
- Wahane, A.; Waghmode, A.; Kapphahn, A.; Dhuri, K.; Gupta, A.; Bahal, R. Role of Lipid-Based and Polymer-Based Non-Viral Vectors in Nucleic Acid Delivery for Next-Generation Gene Therapy. Molecules 2020, 25, 2866. [Google Scholar] [CrossRef]
- Yu, H.; Zou, Y.; Wang, Y.; Huang, X.; Huang, G.; Sumer, B.D.; Boothman, D.A.; Gao, J. Overcoming endosomal barrier by amphotericin B-loaded dual pH-responsive PDMA-b-PDPA micelleplexes for siRNA delivery. ACS Nano 2011, 5, 9246–9255. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Saeed, M.; Zhou, Y.; Yang, L.; Wang, D.; Yu, H. Non-viral gene delivery for cancer immunotherapy. J. Gene Med. 2019, 21, e3092. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Wan, T.; Wang, H.; Zhang, S.; Ping, Y.; Cheng, Y. A boronic acid–rich dendrimer with robust and unprecedented efficiency for cytosolic protein delivery and CRISPR-Cas9 gene editing. Sci. Adv. 2019, 5, eaaw8922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wan, T.; Chen, Y.; Chen, Y.; Sun, H.; Cao, T.; Songyang, Z.; Tang, G.; Wu, C.; Ping, Y. Cationic polymer-mediated CRISPR/Cas9 plasmid delivery for genome editing. Macromol. Rapid Commun. 2019, 40, 1800068. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bai, H.; Wang, H.; Shen, Y.; Tang, G.; Ping, Y. Reactive oxygen species (ROS)-responsive nanomedicine for RNAi-based cancer therapy. Nanoscale 2018, 10, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Lutz, J.-F. Sequence-controlled polymerizations: The next Holy Grail in polymer science? Polym. Chem. 2010, 1, 55–62. [Google Scholar] [CrossRef]
- Lutz, J.-F.; Ouchi, M.; Liu, D.R.; Sawamoto, M. Sequence-controlled polymers. Science 2013, 341. [Google Scholar] [CrossRef]
- Hartmann, L.; Häfele, S.; Peschka-Süss, R.; Antonietti, M.; Börner, H.G. Sequence positioning of disulfide linkages to program the degradation of monodisperse poly (amidoamines). Macromolecules 2007, 40, 7771–7776. [Google Scholar] [CrossRef]
- Boyer, C.; Bulmus, V.; Davis, T.P.; Ladmiral, V.; Liu, J.; Perrier, S. Bioapplications of RAFT polymerization. Chem. Rev. 2009, 109, 5402–5436. [Google Scholar] [CrossRef]
- Jiang, X.; Lok, M.C.; Hennink, W.E. Degradable-brushed pHEMA–pDMAEMA synthesized via ATRP and click chemistry for gene delivery. Bioconj. Chem. 2007, 18, 2077–2084. [Google Scholar] [CrossRef]
- Cheng, Y.; Yumul, R.C.; Pun, S.H. Virus-inspired polymer for efficient in vitro and in vivo gene delivery. Angew. Chem. Int. Ed. 2016, 55, 12013–12017. [Google Scholar] [CrossRef]
- Nuhn, L.; Kaps, L.; Diken, M.; Schuppan, D.; Zentel, R. Reductive decationizable block copolymers for stimuli-responsive mRNA delivery. Macromol. Rapid Commun. 2016, 37, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Plank, C.; Tang, M.X.; Wolfe, A.R.; Szoka, F.C. Branched cationic peptides for gene delivery: Role of type and number of cationic residues in formation and in vitro activity of DNA polyplexes. Hum. Gene Ther. 1999, 10, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Read, M.L.; Singh, S.; Ahmed, Z.; Stevenson, M.; Briggs, S.S.; Oupicky, D.; Barrett, L.B.; Spice, R.; Kendall, M.; Berry, M. A versatile reducible polycation-based system for efficient delivery of a broad range of nucleic acids. Nucleic Acids Res. 2005, 33, e86. [Google Scholar] [CrossRef] [Green Version]
- Plank, C.; Zatloukal, K.; Cotten, M.; Mechtler, K.; Wagner, E. Gene transfer into hepatocytes using asialoglycoprotein receptor mediated endocytosis of DNA complexed with an artificial tetra-antennary galactose ligand. Bioconj. Chem. 1992, 3, 533–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuberg, P.; Wagner, A.; Remy, J.-S.; Kichler, A. Design and evaluation of ionizable peptide amphiphiles for siRNA delivery. Int. J. Pharm. 2019, 566, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Ohsaki, M.; Okuda, T.; Wada, A.; Hirayama, T.; Niidome, T.; Aoyagi, H. In vitro gene transfection using dendritic poly(l-lysine). Bioconj. Chem. 2002, 13, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, M.S.; Collard, W.T.; Adami, R.C.; McKenzie, D.L.; Rice, K.G. Peptide-mediated gene delivery: Influence of peptide structure on gene expression. Bioconj. Chem. 1997, 8, 81–88. [Google Scholar] [CrossRef]
- McKenzie, D.L.; Kwok, K.Y.; Rice, K.G. A potent new class of reductively activated peptide gene delivery agents. J. Biol. Chem. 2000, 275, 9970–9977. [Google Scholar] [CrossRef] [Green Version]
- McKenzie, D.L.; Smiley, E.; Kwok, K.Y.; Rice, K.G. Low molecular weight disulfide cross-linking peptides as nonviral gene delivery carriers. Bioconj. Chem. 2000, 11, 901–909. [Google Scholar] [CrossRef]
- Kadlecova, Z.; Baldi, L.; Hacker, D.; Wurm, F.M.; Klok, H.-A. Comparative study on the in vitro cytotoxicity of linear, dendritic, and hyperbranched polylysine analogues. Biomacromolecules 2012, 13, 3127–3137. [Google Scholar] [CrossRef]
- Kawano, T.; Okuda, T.; Aoyagi, H.; Niidome, T. Long circulation of intravenously administered plasmid DNA delivered with dendritic poly(l-lysine) in the blood flow. J. Control. Release 2004, 99, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Kawakami, S.; Akimoto, N.; Niidome, T.; Yamashita, F.; Hashida, M. PEGylated lysine dendrimers for tumor-selective targeting after intravenous injection in tumor-bearing mice. J. Control. Release 2006, 116, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Kawakami, S.; Maeie, T.; Niidome, T.; Yamashita, F.; Hashida, M. Biodistribution characteristics of amino acid dendrimers and their PEGylated derivatives after intravenous administration. J. Control. Release 2006, 114, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Sugiyama, A.; Niidome, T.; Aoyagi, H. Characters of dendritic poly(l-lysine) analogues with the terminal lysines replaced with arginines and histidines as gene carriers in vitro. Biomaterials 2004, 25, 537–544. [Google Scholar] [CrossRef]
- Chen, Q.-R.; Zhang, L.; Luther, P.W.; Mixson, A.J. Optimal transfection with the HK polymer depends on its degree of branching and the pH of endocytic vesicles. Nucleic Acids Res. 2002, 30, 1338–1345. [Google Scholar] [CrossRef]
- Leng, Q.; Mixson, A.J. Modified branched peptides with a histidine-rich tail enhance in vitro gene transfection. Nucleic Acids Res. 2005, 33, e40. [Google Scholar] [CrossRef] [Green Version]
- Leng, Q.; Scaria, P.; Zhu, J.; Ambulos, N.; Campbell, P.; Mixson, A.J. Highly branched HK peptides are effective carriers of siRNA. J. Gene Med. 2005, 7, 977–986. [Google Scholar] [CrossRef]
- Leng, Q.; Mixson, A.J. Small interfering RNA targeting Raf-1 inhibits tumor growth in vitro and in vivo. Cancer Gene Ther. 2005, 12, 682–690. [Google Scholar] [CrossRef] [Green Version]
- Leng, Q.; Chou, S.T.; Scaria, P.V.; Woodle, M.C.; Mixson, A.J. Increased tumor distribution and expression of histidine-rich plasmid polyplexes. J. Gene Med. 2014, 16, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Leng, Q.; Mixson, A.J. The neuropilin-1 receptor mediates enhanced tumor delivery of H2K polyplexes. J. Gene Med. 2016, 18, 134–144. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, L.; Stass, S.; Mixson, A. Co-polymer of histidine and lysine markedly enhances transfection efficiency of liposomes. Gene Ther. 2000, 7, 1698–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.-R.; Zhang, L.; Stass, S.A.; Mixson, A.J. Branched co-polymers of histidine and lysine are efficient carriers of plasmids. Nucleic Acids Res. 2001, 29, 1334–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leng, Q.; Scaria, P.; Lu, P.; Woodle, M.C.; Mixson, A.J. Systemic delivery of HK Raf-1 siRNA polyplexes inhibits MDA-MB-435 xenografts. Cancer Gene Ther. 2008, 15, 485–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, S.-T.; Leng, Q.; Scaria, P.; Kahn, J.D.; Tricoli, L.J.; Woodle, M.; Mixson, A.J. Surface-modified HK: siRNA nanoplexes with enhanced pharmacokinetics and tumor growth inhibition. Biomacromolecules 2013, 14, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Takeda, K.; Ezawa, R.; Ishii, J.; Ogino, C.; Kondo, A. A display of pH-sensitive fusogenic GALA peptide facilitates endosomal escape from a Bio-nanocapsule via an endocytic uptake pathway. J. Nanobiotechnol. 2014, 12, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleal, K.; He, L.; Watson, P.D.; Jones, A.T. Endocytosis, intracellular traffic and fate of cell penetrating peptide based conjugates and nanoparticles. Curr. Pharm. Des. 2013, 19, 2878–2894. [Google Scholar] [CrossRef]
- Subbarao, N.K.; Parente, R.A.; Szoka, F.C.; Nadasdi, L.; Pongracz, K. The pH-dependent bilayer destabilization by an amphipathic peptide. Biochemistry 1987, 26, 2964–2972. [Google Scholar] [CrossRef]
- Wyman, T.B.; Nicol, F.; Zelphati, O.; Scaria, P.; Plank, C.; Szoka, F.C. Design, synthesis, and characterization of a cationic peptide that binds to nucleic acids and permeabilizes bilayers. Biochemistry 1997, 36, 3008–3017. [Google Scholar] [CrossRef]
- McCarthy, H.O.; McCaffrey, J.; McCrudden, C.M.; Zholobenko, A.; Ali, A.A.; McBride, J.W.; Massey, A.S.; Pentlavalli, S.; Chen, K.-H.; Cole, G. Development and characterization of self-assembling nanoparticles using a bio-inspired amphipathic peptide for gene delivery. J. Control. Release 2014, 189, 141–149. [Google Scholar] [CrossRef]
- Bennett, R.; Yakkundi, A.; McKeen, H.D.; McClements, L.; McKeogh, T.J.; McCrudden, C.M.; Arthur, K.; Robson, T.; McCarthy, H.O. RALA-mediated delivery of FKBPL nucleic acid therapeutics. Nanomedicine 2015, 10, 2989–3001. [Google Scholar] [CrossRef] [Green Version]
- Udhayakumar, V.K.; De Beuckelaer, A.; McCaffrey, J.; McCrudden, C.M.; Kirschman, J.L.; Vanover, D.; Van Hoecke, L.; Roose, K.; Deswarte, K.; De Geest, B.G. Arginine-rich peptide-based mRNA nanocomplexes efficiently instigate cytotoxic t cell immunity dependent on the amphipathic organization of the peptide. Adv. Healthc. Mater. 2017, 6, 1601412. [Google Scholar] [CrossRef] [PubMed]
- Mäe, M.; Andaloussi, S.E.; Lundin, P.; Oskolkov, N.; Johansson, H.J.; Guterstam, P.; Langel, Ü. A stearylated CPP for delivery of splice correcting oligonucleotides using a non-covalent co-incubation strategy. J. Control. Release 2009, 134, 221–227. [Google Scholar] [CrossRef]
- Lehto, T.; Simonson, O.E.; Mäger, I.; Ezzat, K.; Sork, H.; Copolovici, D.-M.; Viola, J.R.; Zaghloul, E.M.; Lundin, P.; Moreno, P.M. A peptide-based vector for efficient gene transfer in vitro and in vivo. Mol. Ther. 2011, 19, 1457–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EL Andaloussi, S.; Lehto, T.; Mäger, I.; Rosenthal-Aizman, K.; Oprea, I.I.; Simonson, O.E.; Sork, H.; Ezzat, K.; Copolovici, D.M.; Kurrikoff, K. Design of a peptide-based vector, PepFect6, for efficient delivery of siRNA in cell culture and systemically in vivo. Nucleic Acids Res. 2011, 39, 3972–3987. [Google Scholar] [CrossRef] [Green Version]
- Veiman, K.L.; Mäger, I.; Ezzat, K.; Margus, H.; Lehto, T.; Langel, K.; Kurrikoff, K.; Arukuusk, P.; Suhorutšenko, J.; Padari, K.; et al. PepFect14 peptide vector for efficient gene delivery in cell cultures. Mol. Pharm. 2013, 10, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Oskolkov, N.; Arukuusk, P.; Copolovici, D.-M.; Lindberg, S.; Margus, H.; Padari, K.; Pooga, M.; Langel, Ü. NickFects, phosphorylated derivatives of transportan 10 for cellular delivery of oligonucleotides. Int. J. Pept. Res. Ther. 2011, 17, 147. [Google Scholar] [CrossRef]
- Arukuusk, P.; Pärnaste, L.; Margus, H.; Eriksson, N.K.; Vasconcelos, L.; Padari, K.; Pooga, M.; Langel, U. Differential endosomal pathways for radically modified peptide vectors. Bioconj. Chem. 2013, 24, 1721–1732. [Google Scholar] [CrossRef]
- Arukuusk, P.; Pärnaste, L.; Oskolkov, N.; Copolovici, D.M.; Margus, H.; Padari, K.; Möll, K.; Maslovskaja, J.; Tegova, R.; Kivi, G.; et al. New generation of efficient peptide-based vectors, NickFects, for the delivery of nucleic acids. Biochim. Biophys. Acta 2013, 1828, 1365–1373. [Google Scholar] [CrossRef] [Green Version]
- Freimann, K.; Arukuusk, P.; Kurrikoff, K.; Vasconcelos, L.D.F.; Veiman, K.L.; Uusna, J.; Margus, H.; Garcia-Sosa, A.T.; Pooga, M.; Langel, Ü. Optimization of in vivo DNA delivery with NickFect peptide vectors. J. Control. Release 2016, 241, 135–143. [Google Scholar] [CrossRef]
- Fominaya, J.; Gasset, M.; García, R.; Roncal, F.; Pablo Albar, J.; Bernad, A. An optimized amphiphilic cationic peptide as an efficient non-viral gene delivery vector. J. Gene Med. 2000, 2, 455–464. [Google Scholar] [CrossRef]
- Ezzat, K.; EL Andaloussi, S.; Zaghloul, E.M.; Lehto, T.; Lindberg, S.; Moreno, P.M.; Viola, J.R.; Magdy, T.; Abdo, R.; Guterstam, P. PepFect 14, a novel cell-penetrating peptide for oligonucleotide delivery in solution and as solid formulation. Nucleic Acids Res. 2011, 39, 5284–5298. [Google Scholar] [CrossRef] [PubMed]
- Ezzat, K.; Zaghloul, E.M.; Andaloussi, S.E.; Lehto, T.; El-Sayed, R.; Magdy, T.; Smith, C.E.; Langel, Ü. Solid formulation of cell-penetrating peptide nanocomplexes with siRNA and their stability in simulated gastric conditions. J. Control. Release 2012, 162, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hamley, I.W. Peptide nanotubes. Angew. Chem. Int. Ed. 2014, 53, 6866–6881. [Google Scholar] [CrossRef] [PubMed]
- Hartgerink, J.D.; Beniash, E.; Stupp, S.I. Self-assembly and mineralization of peptide-amphiphile nanofibers. Science 2001, 294, 1684–1688. [Google Scholar] [CrossRef] [Green Version]
- Hartgerink, J.D.; Beniash, E.; Stupp, S.I. Peptide-amphiphile nanofibers: A versatile scaffold for the preparation of self-assembling materials. Proc. Natl. Acad. Sci. USA 2002, 99, 5133–5138. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Webber, M.J.; Stupp, S.I. Self-assembly of peptide amphiphiles: From molecules to nanostructures to biomaterials. Biopolymers 2010, 94, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hendricks, M.P.; Sato, K.; Palmer, L.C.; Stupp, S.I. Supramolecular assembly of peptide amphiphiles. Acc. Chem. Res. 2017, 50, 2440–2448. [Google Scholar] [CrossRef]
- Da Silva, R.M.; Van Der Zwaag, D.; Albertazzi, L.; Lee, S.S.; Meijer, E.; Stupp, S.I. Super-resolution microscopy reveals structural diversity in molecular exchange among peptide amphiphile nanofibres. Nat. Commun. 2016, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.b.; Lee, E.; Yoon, Y.R.; Lee, M.S.; Lee, M. Filamentous artificial virus from a self-assembled discrete nanoribbon. Angew. Chem. Int. Ed. 2008, 47, 4525–4528. [Google Scholar] [CrossRef]
- Ni, R.; Chau, Y. Structural mimics of viruses through peptide/DNA co-assembly. J. Am. Chem. Soc. 2014, 136, 17902–17905. [Google Scholar] [CrossRef]
- Ruff, Y.; Moyer, T.; Newcomb, C.J.; Demeler, B.; Stupp, S.I. Precision templating with DNA of a virus-like particle with peptide nanostructures. J. Am. Chem. Soc. 2013, 135, 6211–6219. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garcia, A.; Kraft, D.J.; Janssen, A.F.; Bomans, P.H.; Sommerdijk, N.A.; Thies-Weesie, D.M.; Favretto, M.E.; Brock, R.; De Wolf, F.A.; Werten, M.W. Design and self-assembly of simple coat proteins for artificial viruses. Nat. Nanotechnol. 2014, 9, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Jekhmane, S.; De Haas, R.; Paulino da Silva Filho, O.; van Asbeck, A.H.; Favretto, M.E.; Hernandez Garcia, A.; Brock, R.; De Vries, R. Virus-like particles of mRNA with artificial minimal coat proteins: Particle formation, stability, and transfection efficiency. Nucleic Acid Ther. 2017, 27, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Ni, R.; Chau, Y. Tuning the inter-nanofibril interaction to regulate the morphology and function of peptide/DNA co-assembled viral mimics. Angew. Chem. Int. Ed. 2017, 56, 9356–9360. [Google Scholar] [CrossRef] [PubMed]
- Ni, R.; Chau, Y. Nanoassembly of Oligopeptides and DNA Mimics the Sequential Disassembly of a Spherical Virus. Angew. Chem. Int. Ed. 2020, 59, 3578–3584. [Google Scholar] [CrossRef]
- Serrano, C.M.; Freeman, R.; Godbe, J.; Lewis, J.A.; Stupp, S.I. DNA-Peptide Amphiphile Nanofibers Enhance Aptamer Function. ACS Appl. Bio Mater. 2019, 2, 2955–2963. [Google Scholar] [CrossRef]
- Ryter, J.M.; Schultz, S.C. Molecular basis of double-stranded RNA-protein interactions: Structure of a dsRNA-binding domain complexed with dsRNA. EMBO J. 1998, 17, 7505–7513. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Bergenfeld, I.; Arora, P.S.; Canary, J.W. Reversible redox reconfiguration of secondary structures in a designed peptide. Angew. Chem. Int. Ed. 2012, 124, 12265–12267. [Google Scholar] [CrossRef]
- Hernandez-Garcia, A.; Álvarez, Z.; Simkin, D.; Madhan, A.; Pariset, E.; Tantakitti, F.; de Vargas-Dorantes, O.J.; Lee, S.S.; Kiskinis, E.; Stupp, S.I. Peptide–siRNA Supramolecular Particles for Neural Cell Transfection. Adv. Sci. 2019, 6, 1801458. [Google Scholar] [CrossRef] [Green Version]
- Malamas, A.S.; Gujrati, M.; Kummitha, C.M.; Xu, R.; Lu, Z.R. Design and evaluation of new pH-sensitive amphiphilic cationic lipids for siRNA delivery. J. Control. Release 2013, 171, 296–307. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.L.; Ramusovic, S.; Nguyen, T.; Lu, Z.R. Novel polymerizable surfactants with pH-sensitive amphiphilicity and cell membrane disruption for efficient siRNA delivery. Bioconj. Chem. 2007, 18, 2169–2177. [Google Scholar] [CrossRef]
- Wang, X.L.; Jensen, R.; Lu, Z.R. A novel environment-sensitive biodegradable polydisulfide with protonatable pendants for nucleic acid delivery. J. Control. Release 2007, 120, 250–258. [Google Scholar] [CrossRef]
- Xu, R.; Wang, X.L.; Lu, Z.R. New amphiphilic carriers forming pH-sensitive nanoparticles for nucleic acid delivery. Langmuir 2010, 26, 13874–13882. [Google Scholar] [CrossRef]
- Sun, D.; Sahu, B.; Gao, S.; Schur, R.M.; Vaidya, A.M.; Maeda, A.; Palczewski, K.; Lu, Z.R. Targeted Multifunctional Lipid ECO Plasmid DNA Nanoparticles as Efficient Non-viral Gene Therapy for Leber’s Congenital Amaurosis. Mol. Ther. Nucleic Acids 2017, 7, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Schur, R.M.; Sears, A.E.; Gao, S.-Q.; Sun, W.; Naderi, A.; Kern, T.; Palczewski, K.; Lu, Z.-R. Stable Retinoid Analogue Targeted Dual pH-Sensitive Smart Lipid ECO/pDNA Nanoparticles for Specific Gene Delivery in the Retinal Pigment Epithelium. ACS Appl. Bio Mater. 2020, 3, 3078–3086. [Google Scholar] [CrossRef]
- Sun, D.; Schur, R.M.; Sears, A.E.; Gao, S.Q.; Vaidya, A.; Sun, W.; Maeda, A.; Kern, T.; Palczewski, K.; Lu, Z.R. Non-viral Gene Therapy for Stargardt Disease with ECO/pRHO-ABCA4 Self-Assembled Nanoparticles. Mol. Ther. 2020, 28, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Parvani, J.G.; Gujrati, M.D.; Mack, M.A.; Schiemann, W.P.; Lu, Z.R. Silencing β3 Integrin by Targeted ECO/siRNA Nanoparticles Inhibits EMT and Metastasis of Triple-Negative Breast Cancer. Cancer Res. 2015, 75, 2316–2325. [Google Scholar] [CrossRef] [Green Version]
- Vaidya, A.M.; Sun, Z.; Ayat, N.; Schilb, A.; Liu, X.; Jiang, H.; Sun, D.; Scheidt, J.; Qian, V.; He, S. Systemic delivery of tumor-targeting siRNA nanoparticles against an oncogenic LncRNA facilitates effective triple-negative breast cancer therapy. Bioconj. Chem. 2019, 30, 907–919. [Google Scholar] [CrossRef]
- Hartmann, L.; Krause, E.; Antonietti, M.; Börner, H.G. Solid-phase supported polymer synthesis of sequence-defined, multifunctional poly(amidoamines). Biomacromolecules 2006, 7, 1239–1244. [Google Scholar] [CrossRef]
- Hartmann, L.; Häfele, S.; Peschka-Süss, R.; Antonietti, M.; Börner, H.G. Tailor-made poly(amidoamine)s for controlled complexation and condensation of DNA. Chem. A Eur. J. 2008, 14, 2025–2033. [Google Scholar] [CrossRef]
- Hill, S.A.; Gerke, C.; Hartmann, L. Recent Developments in Solid-Phase Strategies towards Synthetic, Sequence-Defined Macromolecules. Chem. Asian J. 2018, 13, 3611–3622. [Google Scholar] [CrossRef] [PubMed]
- Salcher, E.E.; Kos, P.; Fröhlich, T.; Badgujar, N.; Scheible, M.; Wagner, E. Sequence-defined four-arm oligo(ethanamino)amides for pDNA and siRNA delivery: Impact of building blocks on efficacy. J. Control. Release 2012, 164, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Schaffert, D.; Badgujar, N.; Wagner, E. Novel Fmoc-polyamino acids for solid-phase synthesis of defined polyamidoamines. Org. Lett. 2011, 13, 1586–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boussif, O.; Lezoualc’h, F.; Zanta, M.A.; Mergny, M.D.; Scherman, D.; Demeneix, B.; Behr, J.-P. A versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo: Polyethylenimine. Proc. Natl. Acad. Sci. USA 1995, 92, 7297–7301. [Google Scholar] [CrossRef] [Green Version]
- Wightman, L.; Kircheis, R.; Rössler, V.; Carotta, S.; Ruzicka, R.; Kursa, M.; Wagner, E. Different behavior of branched and linear polyethylenimine for gene delivery in vitro and in vivo. J. Gene Med. 2001, 3, 362–372. [Google Scholar] [CrossRef]
- Scholz, C.; Kos, P.; Leclercq, L.; Jin, X.; Cottet, H.; Wagner, E. Correlation of length of linear oligo(ethanamino) amides with gene transfer and cytotoxicity. ChemMedChem 2014, 9, 2104–2110. [Google Scholar] [CrossRef]
- Schaffert, D.; Troiber, C.; Salcher, E.E.; Fröhlich, T.; Martin, I.; Badgujar, N.; Dohmen, C.; Edinger, D.; Kläger, R.; Maiwald, G.; et al. Solid-phase synthesis of sequence-defined T-, i-, and U-shape polymers for pDNA and siRNA delivery. Angew. Chem. Int. Ed. 2011, 50, 8986–8989. [Google Scholar] [CrossRef]
- Fröhlich, T.; Edinger, D.; Kläger, R.; Troiber, C.; Salcher, E.; Badgujar, N.; Martin, I.; Schaffert, D.; Cengizeroglu, A.; Hadwiger, P.; et al. Structure-activity relationships of siRNA carriers based on sequence-defined oligo (ethane amino) amides. J. Control. Release 2012, 160, 532–541. [Google Scholar] [CrossRef]
- Lächelt, U.; Kos, P.; Mickler, F.M.; Herrmann, A.; Salcher, E.E.; Rödl, W.; Badgujar, N.; Bräuchle, C.; Wagner, E. Fine-tuning of proton sponges by precise diaminoethanes and histidines in pDNA polyplexes. Nanomedicine 2014, 10, 35–44. [Google Scholar] [CrossRef]
- Scholz, C.; Kos, P.; Wagner, E. Comb-like oligoaminoethane carriers: Change in topology improves pDNA delivery. Bioconj. Chem. 2014, 25, 251–261. [Google Scholar] [CrossRef]
- Schaffert, D.; Troiber, C.; Wagner, E. New sequence-defined polyaminoamides with tailored endosomolytic properties for plasmid DNA delivery. Bioconj. Chem. 2012, 23, 1157–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troiber, C.; Edinger, D.; Kos, P.; Schreiner, L.; Kläger, R.; Herrmann, A.; Wagner, E. Stabilizing effect of tyrosine trimers on pDNA and siRNA polyplexes. Biomaterials 2013, 34, 1624–1633. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Müller, K.; Krhac Levacic, A.; Kos, P.; Lächelt, U.; Wagner, E. Combinatorial Optimization of Sequence-Defined Oligo(ethanamino)amides for Folate Receptor-Targeted pDNA and siRNA Delivery. Bioconj. Chem. 2016, 27, 647–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urnauer, S.; Morys, S.; Krhac Levacic, A.; Müller, A.M.; Schug, C.; Schmohl, K.A.; Schwenk, N.; Zach, C.; Carlsen, J.; Bartenstein, P.; et al. Sequence-defined cMET/HGFR-targeted Polymers as Gene Delivery Vehicles for the Theranostic Sodium Iodide Symporter (NIS) Gene. Mol. Ther. 2016, 24, 1395–1404. [Google Scholar] [CrossRef] [Green Version]
- Kos, P.; Lächelt, U.; Herrmann, A.; Mickler, F.M.; Döblinger, M.; He, D.; Krhač Levačić, A.; Morys, S.; Bräuchle, C.; Wagner, E. Histidine-rich stabilized polyplexes for cMet-directed tumor-targeted gene transfer. Nanoscale 2015, 7, 5350–5362. [Google Scholar] [CrossRef] [Green Version]
- Morys, S.; Krhac Levacic, A.; Urnauer, S.; Kempter, S.; Kern, S.; Rädler, J.O.; Spitzweg, C.; Lächelt, U.; Wagner, E. Influence of Defined Hydrophilic Blocks within Oligoaminoamide Copolymers: Compaction versus Shielding of pDNA Nanoparticles. Polymers 2017, 9, 142. [Google Scholar] [CrossRef]
- Hatakeyama, H.; Akita, H.; Harashima, H. A multifunctional envelope type nano device (MEND) for gene delivery to tumours based on the EPR effect: A strategy for overcoming the PEG dilemma. Adv. Drug Deliv. Rev. 2011, 63, 152–160. [Google Scholar] [CrossRef]
- Walker, G.F.; Fella, C.; Pelisek, J.; Fahrmeir, J.; Boeckle, S.; Ogris, M.; Wagner, E. Toward synthetic viruses: Endosomal pH-triggered deshielding of targeted polyplexes greatly enhances gene transfer in vitro and in vivo. Mol. Ther. 2005, 11, 418–425. [Google Scholar] [CrossRef]
- Knorr, V.; Allmendinger, L.; Walker, G.F.; Paintner, F.F.; Wagner, E. An acetal-based PEGylation reagent for pH-sensitive shielding of DNA polyplexes. Bioconj. Chem. 2007, 18, 1218–1225. [Google Scholar] [CrossRef]
- Williford, J.-M.; Archang, M.M.; Minn, I.; Ren, Y.; Wo, M.; Vandermark, J.; Fisher, P.B.; Pomper, M.G.; Mao, H.-Q. Critical length of PEG grafts on lPEI/DNA nanoparticles for efficient in vivo delivery. ACS Biomater. Sci. Eng. 2016, 2, 567–578. [Google Scholar] [CrossRef]
- Klein, P.M.; Reinhard, S.; Lee, D.-J.; Müller, K.; Ponader, D.; Hartmann, L.; Wagner, E. Precise redox-sensitive cleavage sites for improved bioactivity of siRNA lipopolyplexes. Nanoscale 2016, 8, 18098–18104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhard, S.; Wang, Y.; Dengler, S.; Wagner, E. Precise Enzymatic Cleavage Sites for Improved Bioactivity of siRNA Lipo-Polyplexes. Bioconj. Chem. 2018, 29, 3649–3657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoy, S.M. Patisiran: First global approval. Drugs 2018, 78, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J. Givosiran: First Approval. Drugs 2020, 80, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.A.; Castanotto, D. FDA-approved oligonucleotide therapies in 2017. Mol. Ther. 2017, 25, 1069–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, P.M.; Müller, K.; Gutmann, C.; Kos, P.; Levacic, A.K.; Edinger, D.; Höhn, M.; Leroux, J.-C.; Gauthier, M.A.; Wagner, E. Twin disulfides as opportunity for improving stability and transfection efficiency of oligoaminoethane polyplexes. J. Control. Release 2015, 205, 109–119. [Google Scholar] [CrossRef]
- Luo, J.; Höhn, M.; Reinhard, S.; Loy, D.M.; Klein, P.M.; Wagner, E. IL4-Receptor-Targeted Dual Antitumoral Apoptotic Peptide—siRNA Conjugate Lipoplexes. Adv. Funct. Mater. 2019, 29, 1900697. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, J.; Truebenbach, I.; Reinhard, S.; Klein, P.M.; Höhn, M.; Kern, S.; Morys, S.; Loy, D.M.; Wagner, E.; et al. Double Click-Functionalized siRNA Polyplexes for Gene Silencing in Epidermal Growth Factor Receptor-Positive Tumor Cells. ACS Biomater. Sci. Eng. 2020, 6, 1074–1089. [Google Scholar] [CrossRef]
- Zarei, A.; Razban, V.; Hosseini, S.E.; Tabei, S.M.B. Creating cell and animal models of human disease by genome editing using CRISPR/Cas9. J. Gene Med. 2019, 21, e3082. [Google Scholar] [CrossRef]
- Xu, X.; Wan, T.; Xin, H.; Li, D.; Pan, H.; Wu, J.; Ping, Y. Delivery of CRISPR/Cas9 for therapeutic genome editing. J. Gene Med. 2019, 21, e3107. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.K.; Kwon, K.; Ryu, J.S.; Lee, H.N.; Park, C.; Chung, H.J. Nonviral genome editing based on a polymer-derivatized CRISPR nanocomplex for targeting bacterial pathogens and antibiotic resistance. Bioconj. Chem. 2017, 28, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-X.; Zhang, Y.; Yin, H. Genome editing with mRNA encoding ZFN, TALEN, and Cas9. Mol. Ther. 2019, 27, 735–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Sun, Z.; Jiang, H.; Vaidya, A.M.; Xin, R.; Ayat, N.R.; Schilb, A.L.; Qiao, P.L.; Han, Z.; Naderi, A. Synthesis and evaluation of pH-sensitive multifunctional lipids for efficient delivery of CRISPR/Cas9 in gene editing. Bioconj. Chem. 2018, 30, 667–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, J.; Lin, Y.; Krhac Levacic, A.; Al Danaf, N.; Peng, L.; Höhn, M.; Lamb, D.C.; Wagner, E.; Lächelt, U. Delivery of Cas9/sgRNA ribonucleoprotein complexes via hydroxystearyl oligoamino amides. Bioconj. Chem. 2020, 31, 729–742. [Google Scholar] [CrossRef]
- Kuhn, J.; Klein, P.M.; Al Danaf, N.; Nordin, J.Z.; Reinhard, S.; Loy, D.M.; Höhn, M.; El Andaloussi, S.; Lamb, D.C.; Wagner, E. Supramolecular Assembly of Aminoethylene-Lipopeptide PMO Conjugates into RNA Splice-Switching Nanomicelles. Adv. Funct. Mater. 2019, 29, 1906432. [Google Scholar] [CrossRef]
- Abuchowski, A.; McCoy, J.R.; Palczuk, N.C.; van Es, T.; Davis, F.F. Effect of covalent attachment of polyethylene glycol on immunogenicity and circulating life of bovine liver catalase. J. Biol. Chem. 1977, 252, 3582–3586. [Google Scholar]
- Jiang, G.; Park, K.; Kim, J.; Kim, K.S.; Oh, E.J.; Kang, H.; Han, S.E.; Oh, Y.K.; Park, T.G.; Kwang Hahn, S. Hyaluronic acid–polyethyleneimine conjugate for target specific intracellular delivery of siRNA. Biopolymers 2008, 89, 635–642. [Google Scholar] [CrossRef]
- Kursa, M.; Walker, G.F.; Roessler, V.; Ogris, M.; Roedl, W.; Kircheis, R.; Wagner, E. Novel shielded transferrin−polyethylene glycol−polyethylenimine/DNA complexes for systemic tumor-targeted gene transfer. Bioconj. Chem. 2003, 14, 222–231. [Google Scholar] [CrossRef]
- Schaffert, D.; Kiss, M.; Rödl, W.; Shir, A.; Levitzki, A.; Ogris, M.; Wagner, E. Poly(I:C)-mediated tumor growth suppression in EGF-receptor overexpressing tumors using EGF-polyethylene glycol-linear polyethylenimine as carrier. Pharm. Res. 2011, 28, 731–741. [Google Scholar] [CrossRef]
- Dohmen, C.; Edinger, D.; Fröhlich, T.; Schreiner, L.; Lächelt, U.; Troiber, C.; Rädler, J.; Hadwiger, P.; Vornlocher, H.P.; Wagner, E. Nanosized multifunctional polyplexes for receptor-mediated siRNA delivery. ACS Nano 2012, 6, 5198–5208. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H. The enhanced permeability and retention (EPR) effect in tumor vasculature: The key role of tumor-selective macromolecular drug targeting. Adv. Enzym. Regul. 2001, 41, 189–207. [Google Scholar] [CrossRef]
- Sindhwani, S.; Syed, A.M.; Ngai, J.; Kingston, B.R.; Maiorino, L.; Rothschild, J.; MacMillan, P.; Zhang, Y.; Rajesh, N.U.; Hoang, T. The entry of nanoparticles into solid tumours. Nat. Mater. 2020, 19, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Kos, P.; Lächelt, U.; He, D.; Nie, Y.; Gu, Z.; Wagner, E. Dual-targeted polyplexes based on sequence-defined peptide-PEG-oligoamino amides. J. Pharm. Sci. 2015, 104, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Lächelt, U.; Wittmann, V.; Müller, K.; Edinger, D.; Kos, P.; Höhn, M.; Wagner, E. Synthetic polyglutamylation of dual-functional MTX ligands for enhanced combined cytotoxicity of poly(I:C) nanoplexes. Mol. Pharm. 2014, 11, 2631–2639. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Reinhard, S.; Li, C.; Qian, M.; Jiang, H.; Du, Y.; Lächelt, U.; Lu, W.; Wagner, E.; Huang, R. Antitumoral Cascade-Targeting Ligand for IL-6 Receptor-Mediated Gene Delivery to Glioma. Mol. Ther. 2017, 25, 1556–1566. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.J.; Kessel, E.; Edinger, D.; He, D.; Klein, P.M.; Voith von Voithenberg, L.; Lamb, D.C.; Lächelt, U.; Lehto, T.; Wagner, E. Dual antitumoral potency of EG5 siRNA nanoplexes armed with cytotoxic bifunctional glutamyl-methotrexate targeting ligand. Biomaterials 2016, 77, 98–110. [Google Scholar] [CrossRef]
- Dohmen, C.; Fröhlich, T.; Lächelt, U.; Röhl, I.; Vornlocher, H.P.; Hadwiger, P.; Wagner, E. Defined Folate-PEG-siRNA Conjugates for Receptor-specific Gene Silencing. Mol. Ther. Nucleic Acids 2012, 1, e7. [Google Scholar] [CrossRef]
- Plank, C.; Oberhauser, B.; Mechtler, K.; Koch, C.; Wagner, E. The influence of endosome-disruptive peptides on gene transfer using synthetic virus-like gene transfer systems. J. Biol. Chem. 1994, 269, 12918–12924. [Google Scholar]
- Mechtler, K.; Wagner, E. Gene transfer mediated by influenza virus peptides: The role of peptide sequences. New J. Chem. 1997, 21, 105–111. [Google Scholar]
- Lee, D.J.; He, D.; Kessel, E.; Padari, K.; Kempter, S.; Lächelt, U.; Rädler, J.O.; Pooga, M.; Wagner, E. Tumoral gene silencing by receptor-targeted combinatorial siRNA polyplexes. J. Control. Release 2016, 244, 280–291. [Google Scholar] [CrossRef]
- Lee, D.-J.; Kessel, E.; Lehto, T.; Liu, X.; Yoshinaga, N.; Padari, K.; Chen, Y.-C.; Kempter, S.; Uchida, S.; Rädler, J.O. Systemic delivery of folate-PEG siRNA lipopolyplexes with enhanced intracellular stability for in vivo gene silencing in leukemia. Bioconj. Chem. 2017, 28, 2393–2409. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Müller, K.; Kessel, E.; Reinhard, S.; He, D.; Klein, P.M.; Höhn, M.; Rödl, W.; Kempter, S.; Wagner, E. Targeted siRNA Delivery Using a Lipo-Oligoaminoamide Nanocore with an Influenza Peptide and Transferrin Shell. Adv. Healthc. Mater. 2016, 5, 1493–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinborn, B.; Truebenbach, I.; Morys, S.; Lächelt, U.; Wagner, E.; Zhang, W. Epidermal growth factor receptor targeted methotrexate and small interfering RNA co-delivery. J. Gene Med. 2018, 20, e3041. [Google Scholar] [CrossRef]
- Müller, K.; Klein, P.M.; Heissig, P.; Roidl, A.; Wagner, E. EGF receptor targeted lipo-oligocation polyplexes for antitumoral siRNA and miRNA delivery. Nanotechnology 2016, 27, 464001. [Google Scholar] [CrossRef] [PubMed]
- Müller, K.; Kessel, E.; Klein, P.M.; Höhn, M.; Wagner, E. Post-PEGylation of siRNA Lipo-oligoamino Amide Polyplexes Using Tetra-glutamylated Folic Acid as Ligand for Receptor-Targeted Delivery. Mol. Pharm. 2016, 13, 2332–2345. [Google Scholar] [CrossRef] [PubMed]
- Truebenbach, I.; Zhang, W.; Wang, Y.; Kern, S.; Höhn, M.; Reinhard, S.; Gorges, J.; Kazmaier, U.; Wagner, E. Co-delivery of pretubulysin and siEG5 to EGFR overexpressing carcinoma cells. Int. J. Pharm. 2019, 569, 118570. [Google Scholar] [CrossRef]
- Klein, P.M.; Kern, S.; Lee, D.J.; Schmaus, J.; Höhn, M.; Gorges, J.; Kazmaier, U.; Wagner, E. Folate receptor-directed orthogonal click-functionalization of siRNA lipopolyplexes for tumor cell killing in vivo. Biomaterials 2018, 178, 630–642. [Google Scholar] [CrossRef]
- Klein, P.M.; Klinker, K.; Zhang, W.; Kern, S.; Kessel, E.; Wagner, E.; Barz, M. Efficient Shielding of Polyplexes Using Heterotelechelic Polysarcosines. Polymers 2018, 10, 689. [Google Scholar] [CrossRef] [Green Version]
- Ogris, M.; Brunner, S.; Schüller, S.; Kircheis, R.; Wagner, E. PEGylated DNA/transferrin–PEI complexes: Reduced interaction with blood components, extended circulation in blood and potential for systemic gene delivery. Gene Ther. 1999, 6, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Beckert, L.; Kostka, L.; Kessel, E.; Levacic, A.K.; Kostkova, H.; Etrych, T.; Lächelt, U.; Wagner, E. Acid-labile pHPMA modification of four-arm oligoaminoamide pDNA polyplexes balances shielding and gene transfer activity in vitro and in vivo. Eur. J. Pharm. Biopharm. 2016, 105, 85–96. [Google Scholar] [CrossRef]
- Rostovtsev, V.V.; Green, L.G.; Fokin, V.V.; Sharpless, K.B. A stepwise huisgen cycloaddition process: Copper(I)-catalyzed regioselective “ligation” of azides and terminal alkynes. Angew. Chem. 2002, 41, 2596–2599. [Google Scholar] [CrossRef]
- Chang, P.V.; Prescher, J.A.; Sletten, E.M.; Baskin, J.M.; Miller, I.A.; Agard, N.J.; Lo, A.; Bertozzi, C.R. Copper-free click chemistry in living animals. Proc. Natl. Acad. Sci. USA 2010, 107, 1821–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskin, J.M.; Prescher, J.A.; Laughlin, S.T.; Agard, N.J.; Chang, P.V.; Miller, I.A.; Lo, A.; Codelli, J.A.; Bertozzi, C.R. Copper-free click chemistry for dynamic in vivo imaging. Proc. Natl. Acad. Sci. USA 2007, 104, 16793–16797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behr, J.-P. The proton sponge: A trick to enter cells the viruses did not exploit. Chim. Int. J. Chem. 1997, 51, 34–36. [Google Scholar]
- Gilleron, J.; Querbes, W.; Zeigerer, A.; Borodovsky, A.; Marsico, G.; Schubert, U.; Manygoats, K.; Seifert, S.; Andree, C.; Stöter, M. Image-based analysis of lipid nanoparticle–mediated siRNA delivery, intracellular trafficking and endosomal escape. Nat. Biotechnol. 2013, 31, 638–646. [Google Scholar] [CrossRef]
- Wittrup, A.; Ai, A.; Liu, X.; Hamar, P.; Trifonova, R.; Charisse, K.; Manoharan, M.; Kirchhausen, T.; Lieberman, J. Visualizing lipid-formulated siRNA release from endosomes and target gene knockdown. Nat. Biotechnol. 2015, 33, 870–876. [Google Scholar] [CrossRef] [Green Version]
- Brunner, S.; Sauer, T.; Carotta, S.E.A.; Cotten, M.; Saltik, M.; Wagner, E. Cell cycle dependence of gene transfer by lipoplex, polyplex and recombinant adenovirus. Gene Ther. 2000, 7, 401–407. [Google Scholar] [CrossRef]
- Brunner, S.; Fürtbauer, E.; Sauer, T.; Kursa, M.; Wagner, E. Overcoming the nuclear barrier: Cell cycle independent nonviral gene transfer with linear polyethylenimine or electroporation. Mol. Ther. 2002, 5, 80–86. [Google Scholar] [CrossRef]
- Grosse, S.; Thévenot, G.; Monsigny, M.; Fajac, I. Which mechanism for nuclear import of plasmid DNA complexed with polyethylenimine derivatives? J. Gene Med. 2006, 8, 845–851. [Google Scholar] [CrossRef]
- Grandinetti, G.; Reineke, T.M. Exploring the mechanism of plasmid DNA nuclear internalization with polymer-based vehicles. Mol. Pharm. 2012, 9, 2256–2267. [Google Scholar] [CrossRef] [PubMed]
- Pack, D.W.; Hoffman, A.S.; Pun, S.; Stayton, P.S. Design and development of polymers for gene delivery. Nat. Rev. Drug Discov. 2005, 4, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Ranganathan, P.; Diamond, S.L. Nuclear targeting peptide scaffolds for lipofection of nondividing mammalian cells. Nat. Biotechnol. 1999, 17, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Brandén, L.J.; Mohamed, A.J.; Smith, C.E. A peptide nucleic acid–nuclear localization signal fusion that mediates nuclear transport of DNA. Nat. Biotechnol. 1999, 17, 784–787. [Google Scholar] [CrossRef]
- Munkonge, F.M.; Dean, D.A.; Hillery, E.; Griesenbach, U.; Alton, E.W. Emerging significance of plasmid DNA nuclear import in gene therapy. Adv. Drug Deliv. Rev. 2003, 55, 749–760. [Google Scholar] [CrossRef]
- Ciolina, C.; Byk, G.; Blanche, F.; Thuillier, V.; Scherman, D.; Wils, P. Coupling of nuclear localization signals to plasmid DNA and specific interaction of the conjugates with importin α. Bioconj. Chem. 1999, 10, 49–55. [Google Scholar] [CrossRef]
- Cartier, R.; Reszka, R. Utilization of synthetic peptides containing nuclear localization signals for nonviral gene transfer systems. Gene Ther. 2002, 9, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Bremner, K.H.; Seymour, L.W.; Logan, A.; Read, M.L. Factors influencing the ability of nuclear localization sequence peptides to enhance nonviral gene delivery. Bioconj. Chem. 2004, 15, 152–161. [Google Scholar] [CrossRef]
- Mesika, A.; Kiss, V.; Brumfeld, V.; Ghosh, G.; Reich, Z. Enhanced intracellular mobility and nuclear accumulation of DNA plasmids associated with a karyophilic protein. Hum. Gene Ther. 2005, 16, 200–208. [Google Scholar] [CrossRef]
- Remaut, K.; Symens, N.; Lucas, B.; Demeester, J.; De Smedt, S. Cell division responsive peptides for optimized plasmid DNA delivery: The mitotic window of opportunity? J. Control. Release 2014, 179, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wagstaff, K.M.; Glover, D.J.; Tremethick, D.J.; Jans, D.A. Histone-mediated transduction as an efficient means for gene delivery. Mol. Ther. 2007, 15, 721–731. [Google Scholar] [CrossRef]
- Fritz, J.D.; Herweijer, H.; Zhang, G.; Wolff, J.A. Gene transfer into mammalian cells using histone-condensed plasmid DNA. Hum. Gene Ther. 1996, 7, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Pigeon, L.; Gonçalves, C.; Gosset, D.; Pichon, C.; Midoux, P. An E3-14.7 K Peptide that Promotes Microtubules-Mediated Transport of Plasmid DNA Increases Polyplexes Transfection Efficiency. Small 2013, 9, 3845–3851. [Google Scholar] [CrossRef] [PubMed]
- Midoux, P.; Pigeon, L.; Gonçalves, C.; Pichon, C. Peptides mediating DNA transport on microtubules and their impact on non-viral gene transfer efficiency. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, S.-T.; Leng, Q.; Scaria, P.; Woodle, M.; Mixson, A.J. Selective modification of HK peptides enhances siRNA silencing of tumor targets in vivo. Cancer Gene Ther. 2011, 18, 707–716. [Google Scholar] [CrossRef]
- Love, K.T.; Mahon, K.P.; Levins, C.G.; Whitehead, K.A.; Querbes, W.; Dorkin, J.R.; Qin, J.; Cantley, W.; Qin, L.L.; Racie, T. Lipid-like materials for low-dose, in vivo gene silencing. Proc. Natl. Acad. Sci. USA 2010, 107, 1864–1869. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.R.; Marshall, J.; Siegel, C.S.; Jiang, C.; Yew, N.S.; Nichols, M.R.; Nietupski, J.B.; Ziegler, R.J.; Lane, M.B.; Wang, K.X. Detailed analysis of structures and formulations of cationic lipids for efficient gene transfer to the lung. Hum. Gene Ther. 1996, 7, 1701–1717. [Google Scholar] [CrossRef]
- Dong, Y.; Love, K.T.; Dorkin, J.R.; Sirirungruang, S.; Zhang, Y.; Chen, D.; Bogorad, R.L.; Yin, H.; Chen, Y.; Vegas, A.J. Lipopeptide nanoparticles for potent and selective siRNA delivery in rodents and nonhuman primates. Proc. Natl. Acad. Sci. USA 2014, 111, 3955–3960. [Google Scholar] [CrossRef] [Green Version]
- Dahlman, J.E.; Kauffman, K.J.; Xing, Y.; Shaw, T.E.; Mir, F.F.; Dlott, C.C.; Langer, R.; Anderson, D.G.; Wang, E.T. Barcoded nanoparticles for high throughput in vivo discovery of targeted therapeutics. Proc. Natl. Acad. Sci. USA 2017, 114, 2060–2065. [Google Scholar] [CrossRef] [Green Version]
- Neri, D.; Lerner, R.A. DNA-encoded chemical libraries: A selection system based on endowing organic compounds with amplifiable information. Annu. Rev. Biochem. 2018, 87, 479–502. [Google Scholar] [CrossRef]
- Lokugamage, M.P.; Sago, C.D.; Dahlman, J.E. Testing thousands of nanoparticles in vivo using DNA barcodes. Curr. Opin. Biomed. Eng. 2018, 7, 1–8. [Google Scholar] [CrossRef]
- Sago, C.D.; Lokugamage, M.P.; Lando, G.N.; Djeddar, N.; Shah, N.N.; Syed, C.; Bryksin, A.V.; Dahlman, J.E. Modifying a commonly expressed endocytic receptor retargets nanoparticles in vivo. Nano Lett. 2018, 18, 7590–7600. [Google Scholar] [CrossRef] [PubMed]
- Paunovska, K.; Sago, C.D.; Monaco, C.M.; Hudson, W.H.; Castro, M.G.; Rudoltz, T.G.; Kalathoor, S.; Vanover, D.A.; Santangelo, P.J.; Ahmed, R. A direct comparison of in vitro and in vivo nucleic acid delivery mediated by hundreds of nanoparticles reveals a weak correlation. Nano Lett. 2018, 18, 2148–2157. [Google Scholar] [CrossRef]
- Sago, C.D.; Lokugamage, M.P.; Islam, F.Z.; Krupczak, B.R.; Sato, M.; Dahlman, J.E. Nanoparticles that deliver RNA to bone marrow identified by in vivo directed evolution. J. Am. Chem. Soc. 2018, 140, 17095–17105. [Google Scholar] [CrossRef] [PubMed]
- Paunovska, K.; Gil, C.J.; Lokugamage, M.P.; Sago, C.D.; Sato, M.; Lando, G.N.; Gamboa Castro, M.; Bryksin, A.V.; Dahlman, J.E. Analyzing 2000 in vivo drug delivery data points reveals cholesterol structure impacts nanoparticle delivery. ACS Nano 2018, 12, 8341–8349. [Google Scholar] [CrossRef] [PubMed]
- Sago, C.D.; Lokugamage, M.P.; Paunovska, K.; Vanover, D.A.; Monaco, C.M.; Shah, N.N.; Castro, M.G.; Anderson, S.E.; Rudoltz, T.G.; Lando, G.N. High-throughput in vivo screen of functional mRNA delivery identifies nanoparticles for endothelial cell gene editing. Proc. Natl. Acad. Sci. USA 2018, 115, E9944–E9952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paunovska, K.; Da Silva Sanchez, A.J.; Sago, C.D.; Gan, Z.; Lokugamage, M.P.; Islam, F.Z.; Kalathoor, S.; Krupczak, B.R.; Dahlman, J.E. Nanoparticles containing oxidized cholesterol deliver mRNA to the liver microenvironment at clinically relevant doses. Adv. Mater. 2019, 31, 1807748. [Google Scholar] [CrossRef] [PubMed]
- Lokugamage, M.P.; Sago, C.D.; Gan, Z.; Krupczak, B.R.; Dahlman, J.E. Constrained nanoparticles deliver siRNA and sgRNA to T cells in vivo without targeting ligands. Adv. Mater. 2019, 31, 1902251. [Google Scholar] [CrossRef]
- Gan, Z.; Lokugamage, M.P.; Hatit, M.Z.; Loughrey, D.; Paunovska, K.; Sato, M.; Cristian, A.; Dahlman, J.E. Nanoparticles containing constrained phospholipids deliver mRNA to liver immune cells in vivo without targeting ligands. Bioeng. Transl. Med. 2020, e10161. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cargo | Properties | Target Site | Function and Mechanism |
|---|---|---|---|
| pDNA | Circular, large (5–15 kbp) double-stranded DNA | Nucleus | Encodes a cDNA expression cassette under the control of a strong promoter/enhancer unit |
| siRNA | Short noncoding double-stranded RNA with 21–25 bp sequence | Cytosol | RNAi; unwound into single-strand bound in RISC that recognizes the complementary mRNA sequence, resulting in mRNA cleavage. |
| miRNA | MicroRNA, short noncoding endogenous double-stranded RNA | Cytosol | Regulate gene expression post-transcriptionally by binding of mRNA and thus preventing translation of mRNA into protein |
| mRNA | Single stranded sequence transcribed from DNA | Cytosol | Translated into proteins in the cytoplasm |
| sgRNA | Noncoding short RNA sequence binding Cas9 protein | Nucleus | sgRNA guides the nuclease Cas9 to a selective target DNA sequence via complementary binding to make a DSB |
| ASO | Chemically stabilized short single-stranded antisense oligonucleotide | Cytosol/Nucleus | Bind to mRNA and prevent translation, or to induce exon skipping |
| PMO | Synthetic uncharged ASO, in which the ribosyl rings and phospodiesters in the backbone are replaced with methylenemorpholine rings and a phosphorodiamidate backbone, respectively. | Nucleus | Bind to pre-mRNA in the nucleus and alter gene splicing, resulting in the exclusion or inclusion of particular genetic fragments in the mature mRNA |
| Peptide | Structure | Topology | Functions |
|---|---|---|---|
| H2K |  | Linear | Ineffective in nucleic acid delivery in vitro, need to combine with cationic carriers as liposomes [45,51]. Far more effective for in vivo pDNA delivery [49]. |
| H2K4b |  | 4-branched | Effective in pDNA delivery yet infective for in vitro siRNA transfection as compared with H3K8b [47]. Far more effective for in vivo siRNA delivery [48] as compare with H3K8b. |
| H3K4b H3K(+H)4b |  | 4-branched | H3K4b exhibited better in vitro siRNA transfection than H2K4b [47]. H3K(+H)4b exhibited comparable in vitro siRNA transfection efficiency as compared with H3K8b [53]. 4-branched HK peptide exhibited far more effective gene silencing effect for in vivo siRNA administration [48]. |
| H3K8b |  | 8-branched | Highly effective as carriers for siRNA in vitro transfection as compared with 4 branched H2K4b and H3K4b peptide [47]. Lower in vivo siRNA transfection efficiency as compared with 4 branched H2K4b and H3K4b peptide [48]. |
| CPPs | Sequences | Cargos |
|---|---|---|
| GALA | WEAALAEALAEALAEHLAEALAEALEALAA | DNA [58] |
| KALA | WEAKLAKALAKALAKHLAKALAKALKACEA | DNA [58] |
| RALA | WEARLARALARALARHLARALARALRACEA | DNA [58,59], siRNA [60], mRNA [61] |
| RAWA | RAWARALARALRALARALRALAR | DNA [70] |
| PepFect3 | Stearyl-AGYLLGKINLKALAALAKKIL-NH2 | SCOs [62], pDNA [63]. |
| PepFect6 | Stearyl-AGYLLK(K(K2(tfq4)))INLKALAALAKKIL-NH2 | siRNA [64] |
| PepFect14 | Stearyl-AGYLLGKLLOOLAAAALOOLL-NH2 | SCOs [71], pDNA [65], siRNA [72] |
| NickFect1 | Stearyl-AGY(PO3)LLKTNLKALAALAKKIL-NH2 | SCOs [66], pDNA [63] |
| NickFect51 | (Stearyl-AGYLLG)-δ-OINLKALAALAKKIL-NH2 | SCOs, pDNA, siRNA [67,68] |
| NickFect55 | (Stearyl-AGYLLG)-δ-OINLKALAALAKAIL-NH2 | pDNA [69]. |
| Name | Sequences |
|---|---|
| Glu-KW | Glu-GSGSGS-KKKKKKKK-GGSGGS-WKWEWKWEWKWEWG |
| C-Sn-B | C-(GAGAGAGQ)10-K12 |
| K3C6SPD | KKKC6-WLVFFAQQGSPD |
| H4K5-HCBzlCBzlH | HHHH-KKKK-C12LL-H-CBzlCBzl-H-LLGSPD |
| PAs | PalA-VVVAAAEEE |
| P2 | TFVETGSGTSKQVAKRVAAEKLLTKFKT |
| P3 | SIRKLEYEIEELRLRIGGG |
| P4 | SIRKLEYEIEELRLRIGGGTFVETGSGTSKQVAKRVAAEKLLTKFKT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wagner, E. Non-Viral Targeted Nucleic Acid Delivery: Apply Sequences for Optimization. Pharmaceutics 2020, 12, 888. https://doi.org/10.3390/pharmaceutics12090888
Wang Y, Wagner E. Non-Viral Targeted Nucleic Acid Delivery: Apply Sequences for Optimization. Pharmaceutics. 2020; 12(9):888. https://doi.org/10.3390/pharmaceutics12090888
Chicago/Turabian StyleWang, Yanfang, and Ernst Wagner. 2020. "Non-Viral Targeted Nucleic Acid Delivery: Apply Sequences for Optimization" Pharmaceutics 12, no. 9: 888. https://doi.org/10.3390/pharmaceutics12090888