A Colon-Targeted Prodrug, 4-Phenylbutyric Acid-Glutamic Acid Conjugate, Ameliorates 2,4-Dinitrobenzenesulfonic Acid-Induced Colitis in Rats

 , ,
, ,
Abstract
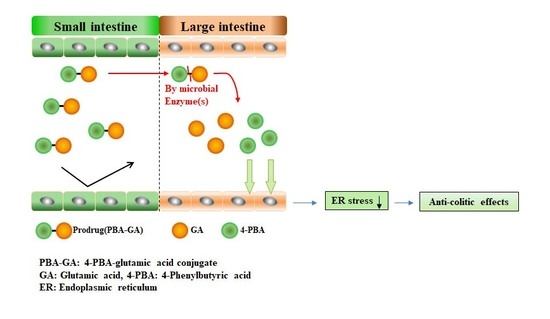
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis of N-(4-Phenylbutanoyl) Glutamic Acid (PBA-GA) and N-(4-Phenylbutanoyl) Aspartic Acid (PBA-AA)
2.3. High-Performance Liquid Chromatography (HPLC)
2.4. Apparent Distribution Coefficient and Chemical Stability
- CO: initial concentration of compound in 1-octanol
- COc: equilibrium concentration of compound in 1-octanol
- CW: equilibrium concentration of compound in isotonic phosphate buffer (pH 6.8)
2.5. Animals
2.6. Incubation of PBA-GA and PBA-AA with the Contents of the Small and Large Intestines of the Rats
2.7. Determination of Drug Concentration in Blood and Cecum
2.8. DNBS-Induced Colitis in Rats
2.9. Evaluation of the Anti-Colitic Effect
2.10. Western Blot Analysis
2.11. Enzyme-Linked Immunosorbent Assay (ELISA) for Cytokine-Induced Neutrophil Chemoattractant-3 CINC-3)
2.12. Data Analysis
3. Results
3.1. Synthesis of 4-PBA Conjugated with Acidic Amino Acids
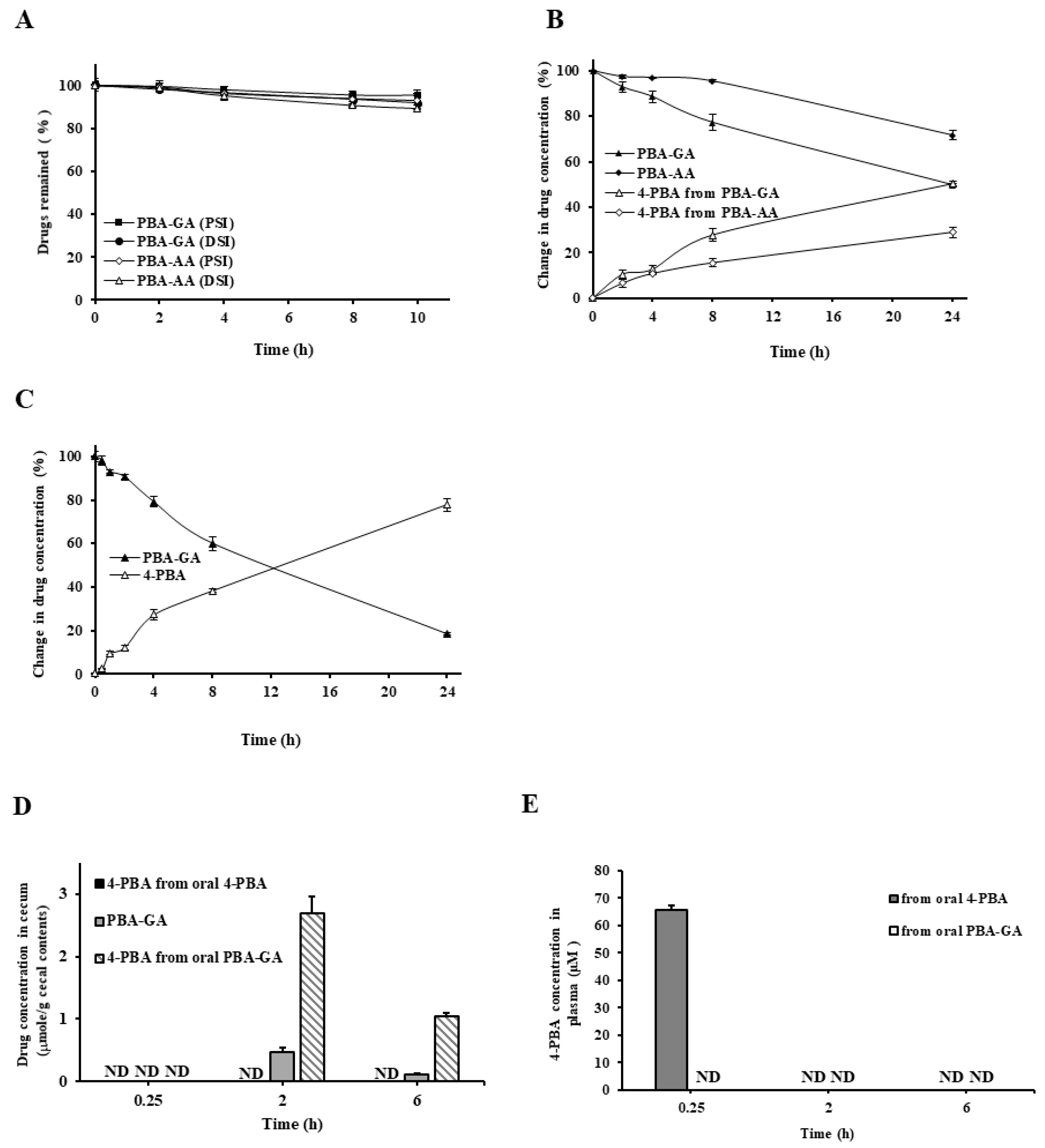
3.2. Colon Specificity of PBA-AA and PBA-GA
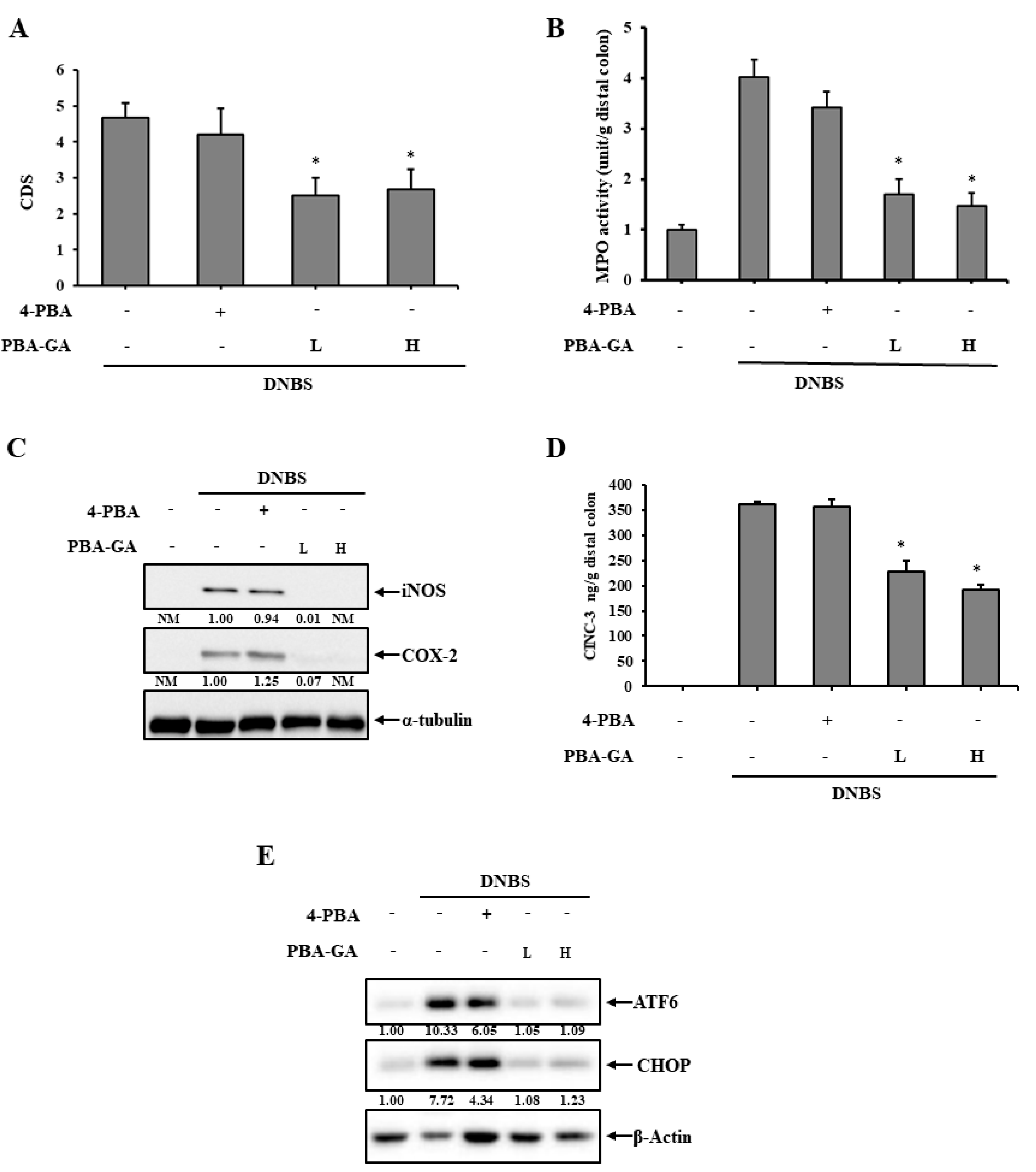
3.3. PBA-GA Mitigates DNBS-Induced Colitis in Rats
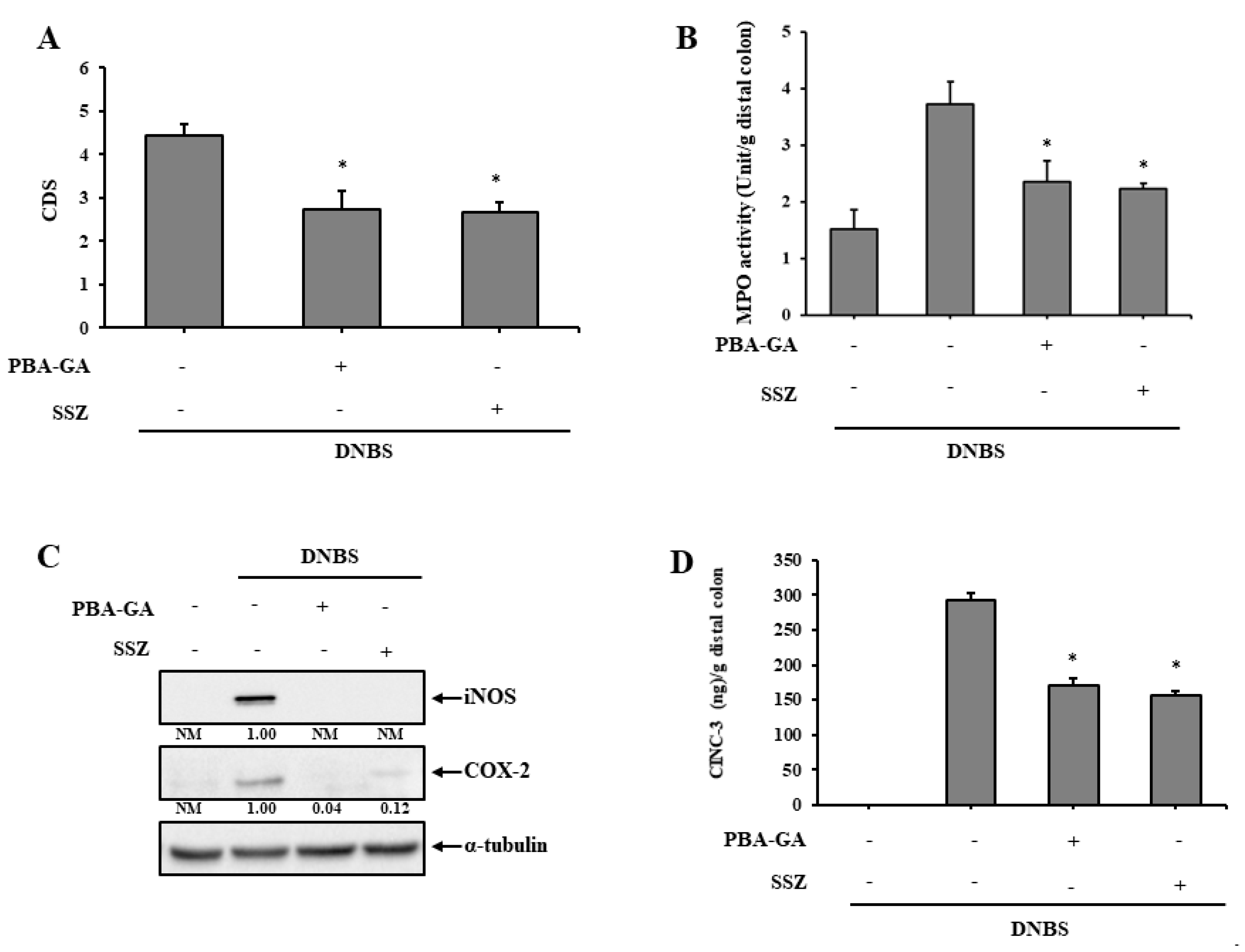
3.4. PBA-GA Is as Effective as SSZ in Mitigating DNBS-Induced Rat Colitis
4. Discussions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Abraham, C.; Cho, J.H. Inflammatory Bowel Disease. N. Engl. J. Med. 2009, 361, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Bryant, R.V.; Brain, O.; Travis, S.P. Conventional drug therapy for inflammatory bowel disease. Scand. J. Gastroenterol. 2015, 50, 90–112. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.K.; Auyeung, K.K. Inflammatory bowel disease: Etiology, pathogenesis and current therapy. Curr. Pharm. Des. 2014, 20, 1082–1096. [Google Scholar] [CrossRef] [PubMed]
- Randall, C.W.; Vizuete, J.A.; Martinez, N.; Alvarez, J.J.; Garapati, K.V.; Malakouti, M.; Taboada, C.M. From historical perspectives to modern therapy: A review of current and future biological treatments for Crohn’s disease. Ther. Adv. Gastroenterol. 2015, 8, 143–159. [Google Scholar] [CrossRef] [Green Version]
- Pithadia, A.B.; Jain, S. Treatment of inflammatory bowel disease (IBD). Pharmacol. Rep. 2011, 63, 629–642. [Google Scholar] [CrossRef]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ho, S.B. Intestinal Goblet Cells and Mucins in Health and Disease: Recent Insights and Progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Gallo, R.L.; Hooper, L.V. Epithelial antimicrobial defence of the skin and intestine. Nat. Rev. Immunol. 2012, 12, 503–516. [Google Scholar] [CrossRef] [Green Version]
- Henderson, P.; Van Limbergen, J.E.; Schwarze, J.; Wilson, D.C. Function of the intestinal epithelium and its dysregulation in inflammatory bowel disease. Inflamm. Bowel Dis. 2011, 17, 382–395. [Google Scholar] [CrossRef]
- Vaishnava, S.; Behrendt, C.L.; Ismail, A.S.; Eckmann, L.; Hooper, L.V. Paneth cells directly sense gut commensals and maintain homeostasis at the intestinal host-microbial interface. Proc. Natl. Acad. Sci. USA 2008, 105, 20858–20863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.S.; Chamaillard, M.; Ogura, Y.; Henegariu, O.; Inohara, N.; Núñez, G.; Flavell, R.A. Nod2-Dependent Regulation of Innate and Adaptive Immunity in the Intestinal Tract. Science 2005, 307, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Salim, S.Y.; Söderholm, J.D. Importance of disrupted intestinal barrier in inflammatory bowel diseases. Inflamm. Bowel Dis. 2011, 17, 362–381. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Ng, S.C. The Gut Microbiota in the Pathogenesis and Therapeutics of Inflammatory Bowel Disease. Front. Microbiol. 2018, 9, 2247. [Google Scholar] [CrossRef] [PubMed]
- Atreya, R.; Neurath, M.F. IBD pathogenesis in 2014: Molecular pathways controlling barrier function in IBD. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 67–68. [Google Scholar] [CrossRef]
- Coleman, O.I.; Haller, D. ER Stress and the UPR in Shaping Intestinal Tissue Homeostasis and Immunity. Front. Immunol. 2019, 10, 2825. [Google Scholar] [CrossRef]
- Cao, S.S. Endoplasmic Reticulum Stress and Unfolded Protein Response in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2015, 21, 636–644. [Google Scholar] [CrossRef]
- Ma, X.; Dai, Z.; Sun, K.; Zhang, Y.; Chen, J.; Yang, Y.; Tso, P.; Wu, G.; Wu, Z. Intestinal Epithelial Cell Endoplasmic Reticulum Stress and Inflammatory Bowel Disease Pathogenesis: An Update Review. Front. Immunol. 2017, 8, 1271. [Google Scholar] [CrossRef] [Green Version]
- Eri, R.D.; Adams, R.J.; Tran, T.V.; Tong, H.; Das, I.; Roche, D.K.; Oancea, I.; Png, C.W.; Jeffery, P.L.; Radford-Smith, G.L.; et al. An intestinal epithelial defect conferring ER stress results in inflammation involving both innate and adaptive immunity. Mucosal Immunol. 2011, 4, 354–364. [Google Scholar] [CrossRef]
- Cao, S.S. Epithelial ER Stress in Crohn’s Disease and Ulcerative Colitis. Inflamm. Bowel Dis. 2016, 22, 984–993. [Google Scholar] [CrossRef]
- Hetz, C.; Chevet, E.; Harding, H.P. Targeting the unfolded protein response in disease. Nat. Rev. Drug Discov. 2013, 12, 703–719. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Kaufman, R.J. The unfolded protein response. J. Cell Sci. 2003, 116, 1861–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadowaki, H.; Nishitoh, H. Signaling Pathways from the Endoplasmic Reticulum and Their Roles in Disease. Genes Basel 2013, 4, 306–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryno, L.M.; Wiseman, R.L.; Kelly, J.W. Targeting unfolded protein response signaling pathways to ameliorate protein misfolding diseases. Curr. Opin. Chem. Biol. 2013, 17, 346–352. [Google Scholar] [CrossRef] [Green Version]
- Burrows, J.A.; Willis, L.K.; Perlmutter, D.H. Chemical chaperones mediate increased secretion of mutant alpha 1-antitrypsin (alpha 1-AT) Z: A potential pharmacological strategy for prevention of liver injury and emphysema in alpha 1-AT deficiency. Proc. Natl. Acad. Sci. USA 2000, 97, 1796–1801. [Google Scholar] [CrossRef] [Green Version]
- Powers, E.T.; Morimoto, R.I.; Dillin, A.; Kelly, J.W.; Balch, W.E. Biological and Chemical Approaches to Diseases of Proteostasis Deficiency. Annu. Rev. Biochem. 2009, 78, 959–991. [Google Scholar] [CrossRef] [Green Version]
- Ozcan, U.; Yilmaz, E.; Ozcan, L.; Furuhashi, M.; Vaillancourt, É.; Smith, R.O.; Görgün, C.Z.; Hotamisligil, G.S. Chemical Chaperones Reduce ER Stress and Restore Glucose Homeostasis in a Mouse Model of Type 2 Diabetes. Science 2006, 313, 1137–1140. [Google Scholar] [CrossRef] [Green Version]
- Kolb, P.S.; Ayaub, E.A.; Zhou, W.; Yum, V.; Dickhout, J.G.; Ask, K. The therapeutic effects of 4-phenylbutyric acid in maintaining proteostasis. Int. J. Biochem. Cell Biol. 2015, 61, 45–52. [Google Scholar] [CrossRef]
- Ono, K.; Nimura, S.; Hideshima, Y.; Nabeshima, K.; Nakashima, M. Orally administered sodium 4-phenylbutyrate suppresses the development of dextran sulfate sodium-induced colitis in mice. Exp. Ther. Med. 2017, 14, 5485–5490. [Google Scholar] [CrossRef]
- Ono, K.; Nimura, S.; Nishinakagawa, T.; Hideshima, Y.; Enjyoji, M.; Nabeshima, K.; Nakashima, M. Sodium 4-phenylbutyrate suppresses the development of dextran sulfate sodium-induced colitis in mice. Exp. Ther. Med. 2014, 7, 573–578. [Google Scholar] [CrossRef]
- Cao, S.S.; Zimmermann, E.M.; Chuang, B.; Song, B.; Nwokoye, A.; Wilkinson, J.E.; Eaton, K.A.; Kaufman, R.J. The unfolded protein response and chemical chaperones reduce protein misfolding and colitis in mice. Gastroenterology 2013, 144, 989–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.; Kim, Y.M. What should be considered on design of a colon-specific prodrug? Expert Opin. Drug Deliv. 2010, 7, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sinha, V.R. Current pharmaceutical strategies for efficient site specific delivery in inflamed distal intestinal mucosa. J. Control. Release 2018, 272, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Marahatta, A.; Bhandary, B.; Lee, M.-R.; Kim, D.-S.; Lee, Y.C.; Kim, S.-R.; Kim, H.-R.; Chae, H.-J. Determination of phenylbutyric acid and its metabolite phenylacetic acid in different tissues of mouse by liquid chromatography with tandem mass spectrometry and its application in drug tissue distribution. J. Chromatogr. B 2012, 903, 118–125. [Google Scholar] [CrossRef]
- Wang, C.L.; Hsueh, P.R.; Sun, M.J.; Leu, Y.L.; Chang, F.S.; Yang, S.W.; Lian, J.F.; Wang, H.P. PBA-omega-Lys as Sustained Phenylbutyrate-Releasing Prodrug. J. Food Drug Anal. 2010, 18, 371–379. [Google Scholar]
- Morris, G.P.; Beck, P.L.; Herridge, M.S.; Depew, W.T.; Szewczuk, M.R.; Wallace, J.L. Hapten-induced model of chronic inflammation and ulceration in the rat colon. Gastroenterology 1989, 96, 795–803. [Google Scholar] [CrossRef]
- Hong, S.; Yum, S.; Yoo, H.-J.; Kang, S.; Yoon, J.-H.; Min, D.; Kim, Y.M.; Jung, Y. Colon-Targeted Cell-Permeable NFκB Inhibitory Peptide Is Orally Active against Experimental Colitis. Mol. Pharm. 2012, 9, 1310–1319. [Google Scholar] [CrossRef]
- Yano, H.; Hirayama, F.; Kamada, M.; Arima, H.; Uekama, K. Colon-specific delivery of prednisolone-appended α-cyclodextrin conjugate: Alleviation of systemic side effect after oral administration. J. Control. Release 2002, 79, 103–112. [Google Scholar] [CrossRef]
- Kim, H.; Kong, H.; Choi, B.; Yang, Y.; Kim, Y.; Lim, M.J.; Neckers, L.; Jung, Y. Metabolic and Pharmacological Properties of Rutin, a Dietary Quercetin Glycoside, for Treatment of Inflammatory Bowel Disease. Pharm. Res. 2005, 22, 1499–1509. [Google Scholar] [CrossRef]
- Soustek, M.S.; Balsa, E.; Barrow, J.J.; Jedrychowski, M.; Vogel, R.; Smeitink, J.; Gygi, S.P.; Puigserver, P. Inhibition of the ER stress IRE1α inflammatory pathway protects against cell death in mitochondrial complex I mutant cells. Cell Death Dis. 2018, 9, 658. [Google Scholar] [CrossRef] [Green Version]
- Jian, L.; Lu, Y.; Lu, S.; Lu, C. Chemical chaperone 4-phenylbutyric acid protects H9c2 cardiomyocytes from ischemia/reperfusion injury by attenuating endoplasmic reticulum stress-induced apoptosis. Mol. Med. Rep. 2016, 13, 4386–4392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, L.H.; Olson, J.; Tong, W.P.; Young, C.W.; Spriggs, D.R.; Malkin, M.G. Phase I dose escalation clinical trial of phenylbutyrate sodium administered twice daily to patients with advanced solid tumors. Investig. New Drugs 2007, 25, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Batshaw, M.L.; MacArthur, R.B.; Tuchman, M. Alternative pathway therapy for urea cycle disorders: Twenty years later. J. Pediatr. 2001, 138, S46–S55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.K.A.; Howes, D.T.; Piris, J.; Truelove, S.C. Optimum dose of sulphasalazine for maintenance treatment in ulcerative colitis. Gut 1980, 21, 232–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, S.A.; Vockley, J.; Vinks, A.A.; Dong, M.; Diaz, G.A.; McCandless, S.E.; Smith, W.E.; Harding, C.O.; Zori, R.; Ficicioglu, C.; et al. Pharmacokinetics of glycerol phenylbutyrate in pediatric patients 2 months to 2 years of age with urea cycle disorders. Mol. Genet. Metab. 2018, 125, 251–257. [Google Scholar] [CrossRef]
- Kim, W.; Nam, J.; Lee, S.; Jeong, S.; Jung, Y. 5-Aminosalicylic Acid Azo-Linked to Procainamide Acts as an Anticolitic Mutual Prodrug via Additive Inhibition of Nuclear Factor kappaB. Mol. Pharm. 2016, 13, 2126–2135. [Google Scholar] [CrossRef]
- Nenci, A.; Becker, C.; Wullaert, A.; Gareus, R.; Van Loo, G.; Danese, S.; Huth, M.; Nikolaev, A.; Neufert, C.; Madison, B.; et al. Epithelial NEMO links innate immunity to chronic intestinal inflammation. Nature 2007, 446, 557–561. [Google Scholar] [CrossRef]
- Wullaert, A.; Bonnet, M.C.; Pasparakis, M. NF-κB in the regulation of epithelial homeostasis and inflammation. Cell Res. 2011, 21, 146–158. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Lee, S.; Lee, H.; Ju, S.; Park, S.; Kwon, D.; Yoo, J.-W.; Yoon, I.-S.; Min, D.S.; Jung, Y.-S.; et al. A Colon-Targeted Prodrug, 4-Phenylbutyric Acid-Glutamic Acid Conjugate, Ameliorates 2,4-Dinitrobenzenesulfonic Acid-Induced Colitis in Rats. Pharmaceutics 2020, 12, 843. https://doi.org/10.3390/pharmaceutics12090843
Kim S, Lee S, Lee H, Ju S, Park S, Kwon D, Yoo J-W, Yoon I-S, Min DS, Jung Y-S, et al. A Colon-Targeted Prodrug, 4-Phenylbutyric Acid-Glutamic Acid Conjugate, Ameliorates 2,4-Dinitrobenzenesulfonic Acid-Induced Colitis in Rats. Pharmaceutics. 2020; 12(9):843. https://doi.org/10.3390/pharmaceutics12090843
Chicago/Turabian StyleKim, Soojin, Seunghyun Lee, Hanju Lee, Sanghyun Ju, Sohee Park, Doyoung Kwon, Jin-Wook Yoo, In-Soo Yoon, Do Sik Min, Young-Suk Jung, and et al. 2020. "A Colon-Targeted Prodrug, 4-Phenylbutyric Acid-Glutamic Acid Conjugate, Ameliorates 2,4-Dinitrobenzenesulfonic Acid-Induced Colitis in Rats" Pharmaceutics 12, no. 9: 843. https://doi.org/10.3390/pharmaceutics12090843