RETRACTED: Thymoquinone-Loaded Soy-Phospholipid-Based Phytosomes Exhibit Anticancer Potential against Human Lung Cancer Cells
 ,
,  , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Design and Optimization
2.3. Preparation of TQ-phytosomes
2.4. Fourier-Transform Infrared (FTIR) Spectroscopy Study
2.5. Vesicle Size Measurement
2.6. Transmission Electron Microscopy (TEM)
2.7. In Vitro Release
2.8. Cytotoxic Effects of Optimized TQ-phytosomes on Human Lung Cancer Cells
2.8.1. Cell Line and Culture Condition
2.8.2. Determination of IC50 Using MTT Assay
2.8.3. Cell Cycle Analysis
2.8.4. Annexin V Staining
2.8.5. Caspase-3 Assay
2.8.6. Reactive Oxygen Species (ROS) Determination
2.9. Statistical Analysis
3. Results
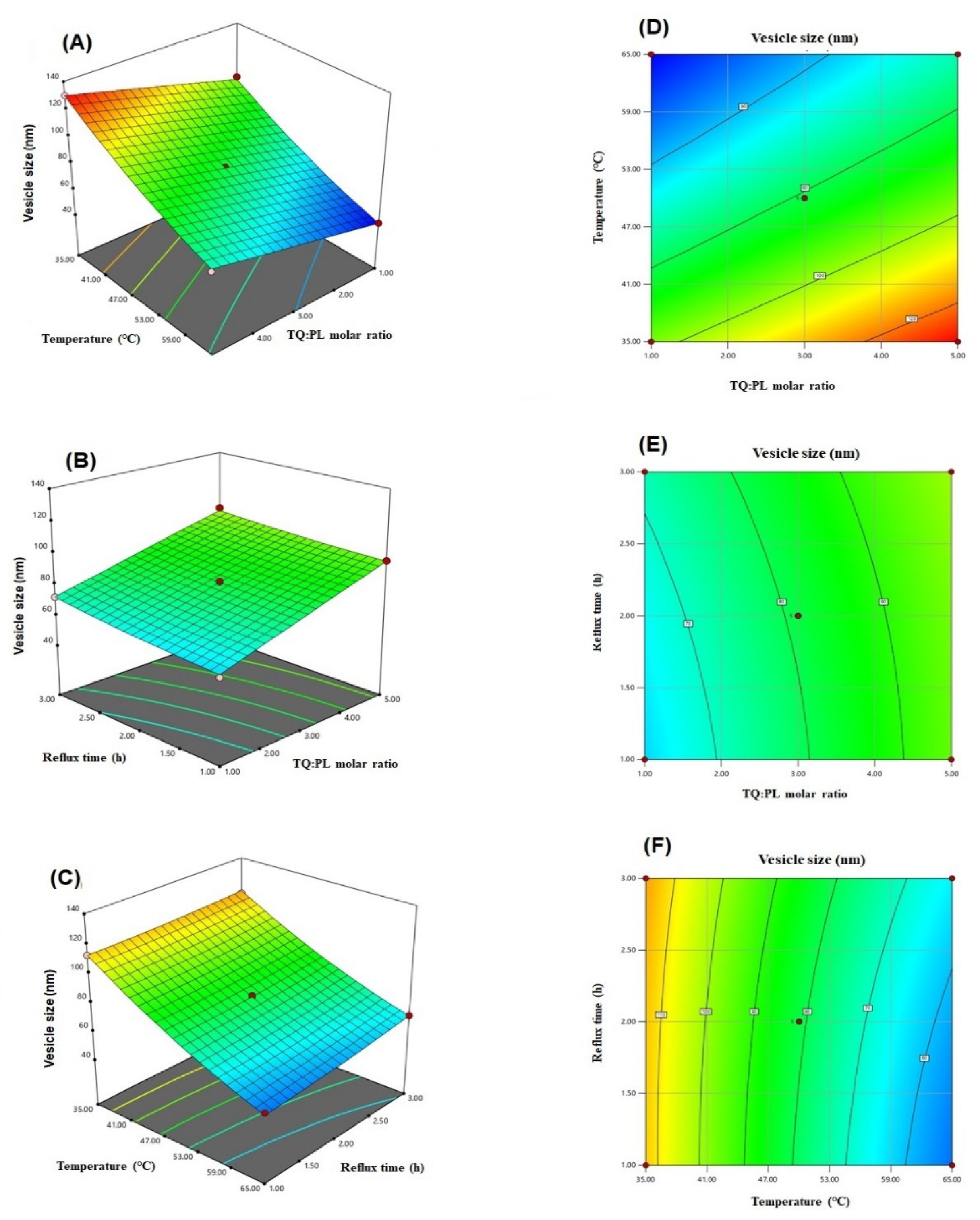
3.1. Influence of Independent Variables on Vesicle Size
3.2. Optimization of TQ-Loaded Phytosomes
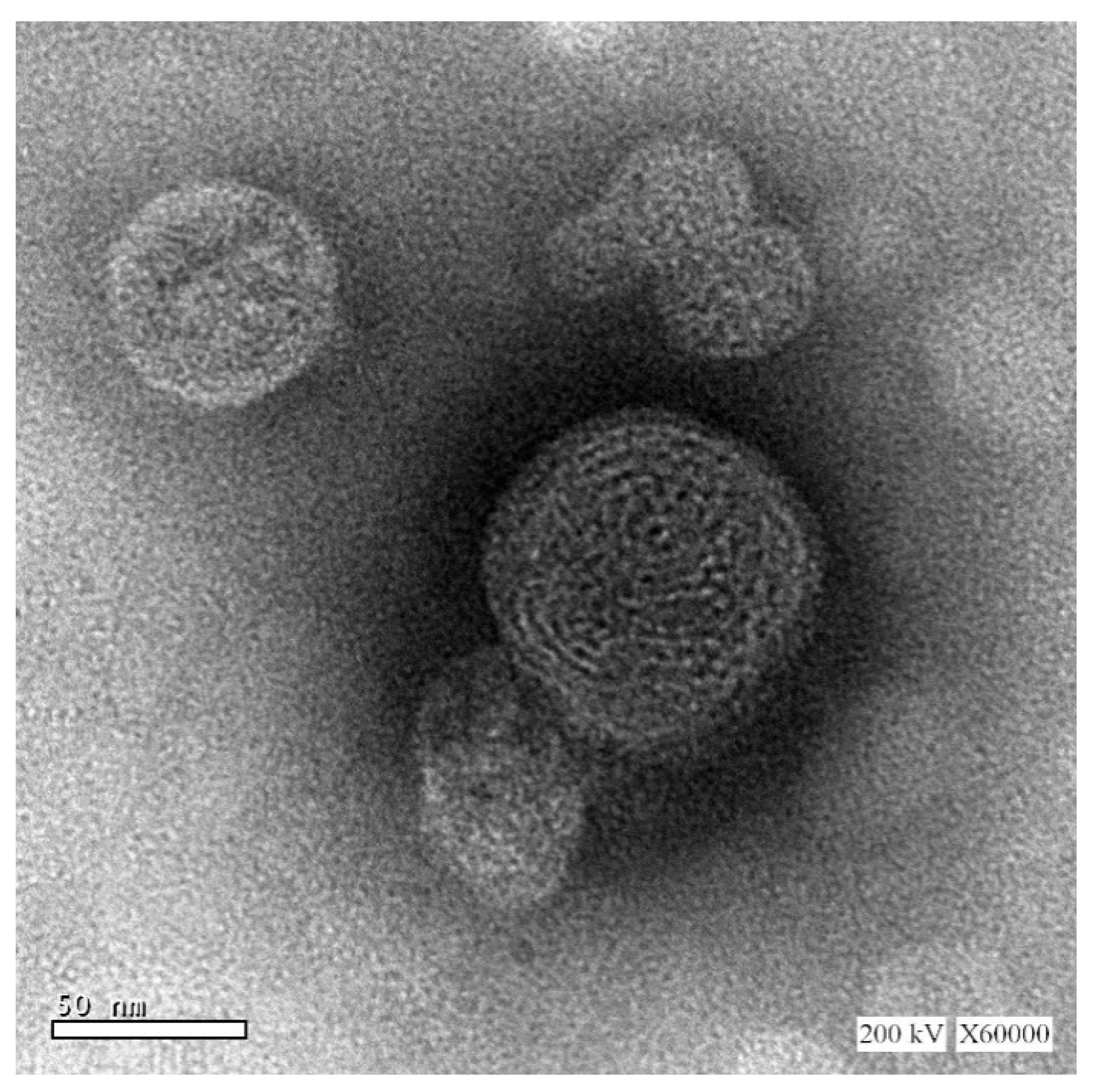
3.3. Transmission Electron Microscopy (TEM)
3.4. Fourier-Transform Infrared (FTIR) Spectroscopy
3.5. In Vitro TQ Release Study
3.6. Cytotoxic Activity
3.6.1. Determination of IC50 Values
3.6.2. Cell Cycle Analysis
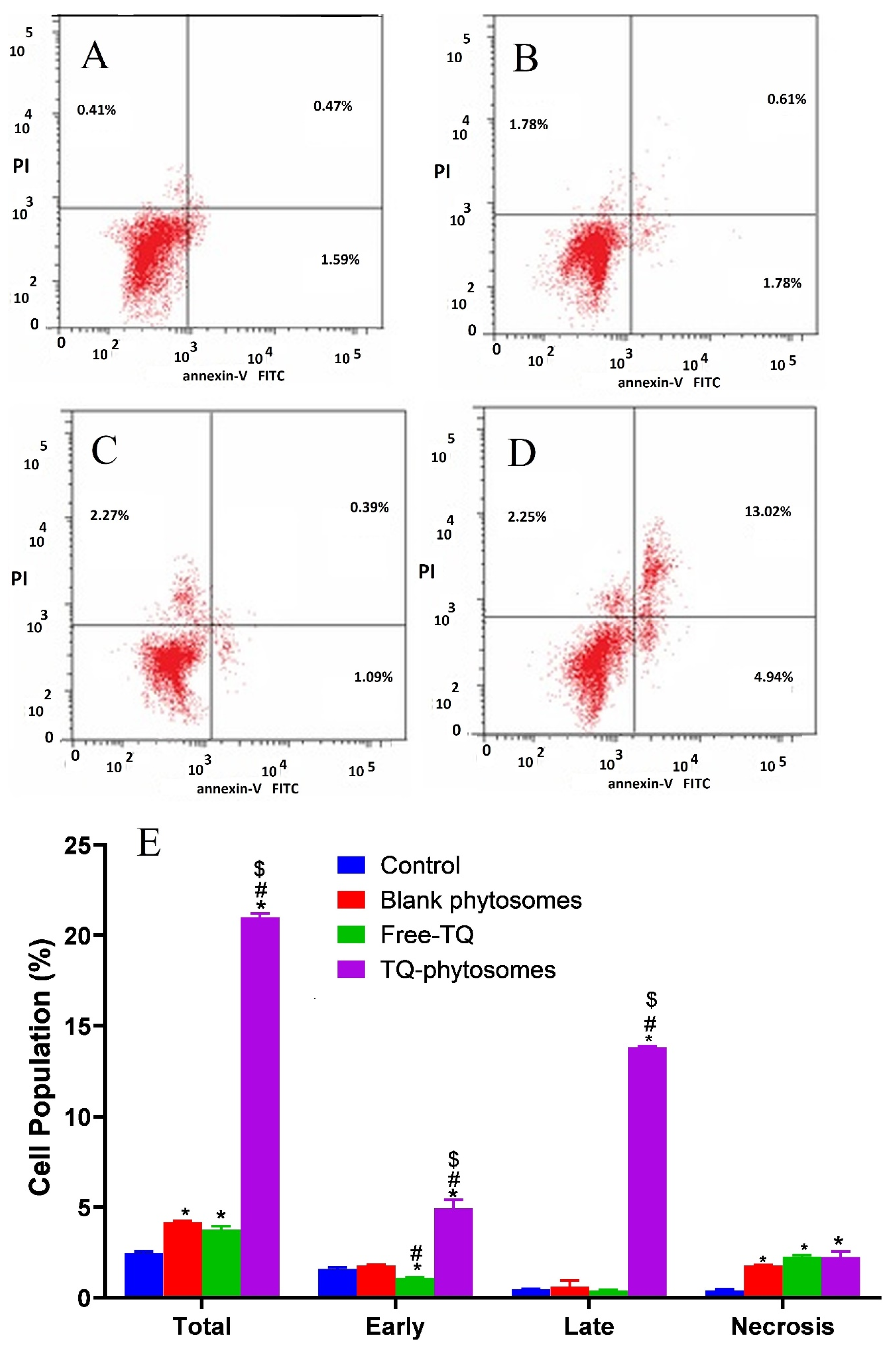
3.6.3. Apoptotic Assessment
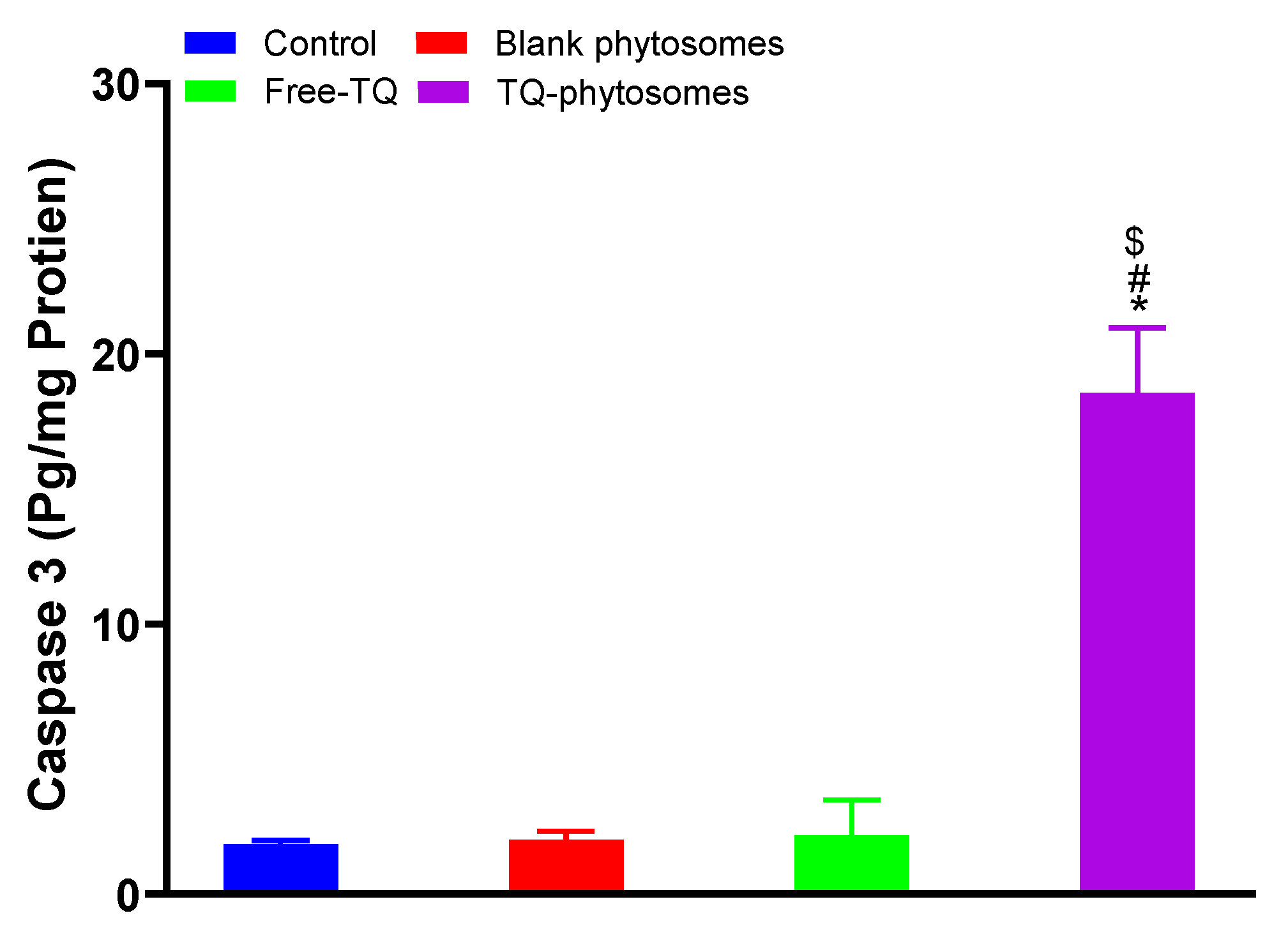
3.6.4. Caspase-3 Assay
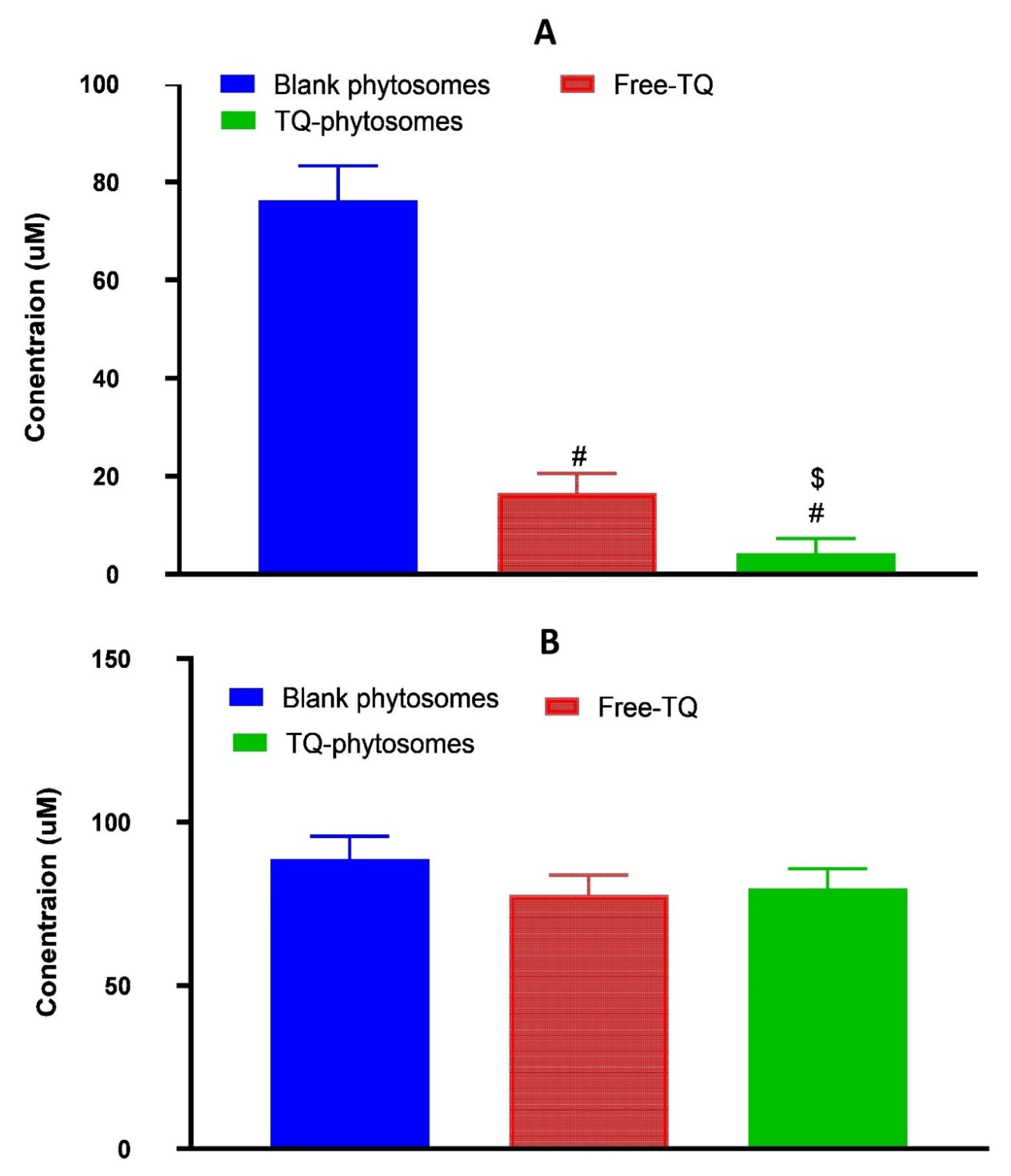
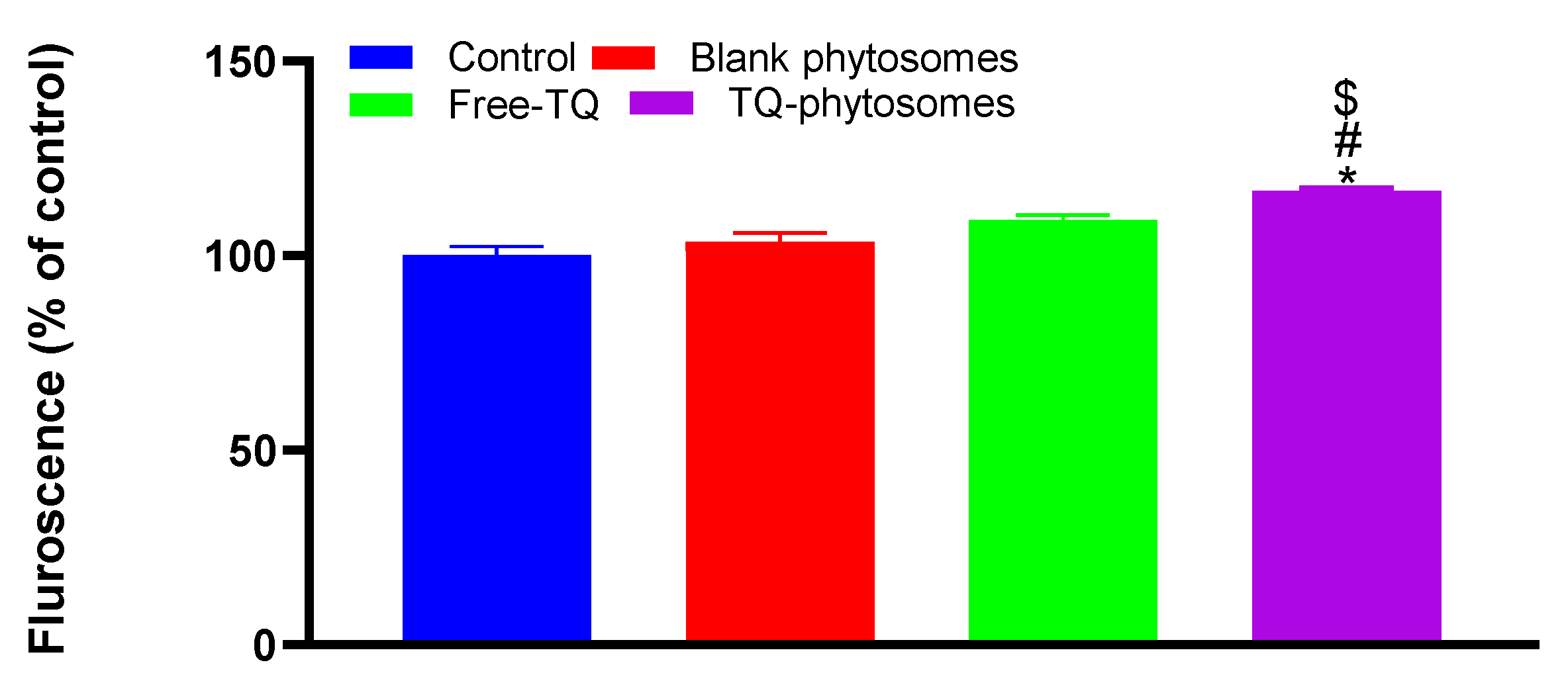
3.6.5. Determination of Generated ROS
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- McMullen, S.; Hess, L.M.; Kim, E.S.; Levy, B.; Mohamed, M.; Waterhouse, D.; Wozniak, A.; Goring, S.; Müller, K.; Muehlenbein, C.; et al. Treatment Decisions for Advanced Non-Squamous Non-Small Cell Lung Cancer: Patient and Physician Perspectives on Maintenance Therapy. Patient 2019, 12, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, J.; Yuan, X.; Hu, C. Simvastatin inhibits the proliferation of A549 lung cancer cells through oxidative stress and up-regulation of SOD2. Pharmazie 2014, 69, 610–614. [Google Scholar] [PubMed]
- Walther, U.; Emmrich, K.; Ramer, R.; Mittag, N. Lovastatin lactone elicits human lung cancer cell apoptosis via a COX-2/PPARγ-dependent pathway. Oncotarget 2016, 7, 10345–10362. [Google Scholar] [CrossRef] [PubMed]
- Alfaifi, M.Y.; Shati, A.A.; Elbehairi, S.E.I.; Fahmy, U.A.; Alhakamy, N.A.; Md, S. Anti-tumor effect of PEG-coated PLGA nanoparticles of febuxostat on A549 non-small cell lung cancer cells. 3 Biotech 2020, 10, 133. [Google Scholar] [CrossRef]
- Lin, J.J.; Ezer, N.; Sigel, K.; Mhango, G.; Wisnivesky, J.P. The Effect of Statins on Survival in Patients with Stage IV Lung Cancer. Lung Cancer 2016, 99, 137–142. [Google Scholar] [CrossRef]
- Khader, M.; Eckl, P.M. Thymoquinone: An emerging natural drug with a wide range of medical applications. Iran. J. Basic Med. Sci. 2014, 17, 950–957. [Google Scholar]
- Schneider-Stock, R.; Fakhoury, I.H.; Zaki, A.M.; El-Baba, C.O.; Gali-Muhtasib, H.U. Thymoquinone: Fifty years of success in the battle against cancer models. Drug Discov. Today 2014, 19, 18–30. [Google Scholar] [CrossRef]
- Gali-Muhtasib, H.; Roessner, A.; Schneider-Stock, R. Thymoquinone: A promising anti-cancer drug from natural sources. Int. J. Biochem. Cell Biol. 2006, 38, 1249–1253. [Google Scholar] [CrossRef]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T. Thymoquinone-induced antitumor and apoptosis in human lung adenocarcinoma cells. J. Cell. Physiol. 2019, 234, 10421–10431. [Google Scholar] [CrossRef]
- El-Najjar, N.; Chatila, M.; Moukadem, H.; Vuorela, H.; Ocker, M.; Gandesiri, M.; Schneider-Stock, R.; Gali-Muhtasib, H. Reactive oxygen species mediate thymoquinone-induced apoptosis and activate ERK and JNK signaling. Apoptosis 2010, 15, 183–195. [Google Scholar] [CrossRef]
- Kalam, M.A.; Raish, M.; Ahmed, A.; Alkharfy, K.M.; Mohsin, K.; Alshamsan, A.; Al-Jenoobi, F.I.; Al-Mohizea, A.M.; Shakeel, F. Oral bioavailability enhancement and hepatoprotective effects of thymoquinone by self-nanoemulsifying drug delivery system. Mater. Sci. Eng. C 2017, 76, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Odeh, F.; Ismail, S.I.; Abu-Dahab, R.; Mahmoud, I.S.; Al Bawab, A. Thymoquinone in liposomes: A study of loading efficiency and biological activity towards breast cancer. Drug Deliv. 2012, 19, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.; Puvvada, N.; Kumar, B.N.P.; Sarkar, S.; Konar, S.; Bharti, R.; Dey, G.; Mazumdar, A.; Pathak, A.; Fisher, P.B.; et al. Overcoming Akt Induced Therapeutic Resistance in Breast Cancer through siRNA and Thymoquinone Encapsulated Multilamellar Gold Niosomes. Mol. Pharm. 2015, 12, 4214–4225. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.K.; Yazan, L.S.; Yap, L.H.; Hafiza, W.A.G.W.N.; How, C.W.; Abdullah, R. Thymoquinone-loaded nanostructured lipid carrier exhibited cytotoxicity towards breast cancer cell lines (MDA-MB-231 and MCF-7) and cervical cancer cell lines (HeLa and SiHa). Biomed Res. Int. 2015, 2015, 263131. [Google Scholar] [CrossRef]
- Soni, P.; Kaur, J.; Tikoo, K. Dual drug-loaded paclitaxel–thymoquinone nanoparticles for effective breast cancer therapy. J. Nanoparticle Res. 2015, 17, 1–12. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Ahir, M.; Patra, P.; Mukherjee, S.; Ghosh, S.; Mazumdar, M.; Chattopadhyay, S.; Das, T.; Chattopadhyay, D.; Adhikary, A. PEGylated-thymoquinone-nanoparticle mediated retardation of breast cancer cell migration by deregulation of cytoskeletal actin polymerization through miR-34a. Biomaterials 2015, 51, 91–107. [Google Scholar] [CrossRef]
- Alhakamy, N.A.; Fahmy, U.A.; Badr-Eldin, S.M.; Ahmed, O.A.A.; Asfour, H.Z.; Aldawsari, H.M.; Algandaby, M.M.; Eid, B.G.; Abdel-Naim, A.B.; Awan, Z.A.; et al. Optimized Icariin Phytosomes Exhibit Enhanced Cytotoxicity and Apoptosis-Inducing Activities in Ovarian Cancer Cells. Pharmaceutics 2020, 12, 346. [Google Scholar] [CrossRef]
- Gnananath, K.; Sri Nataraj, K.; Ganga Rao, B. Phospholipid Complex Technique for Superior Bioavailability of Phytoconstituents. Adv. Pharm. Bull. 2017, 7, 35–42. [Google Scholar] [CrossRef]
- Bombardelli, E.; Spelta, M. Phospholipid-polyphenol complexes: A new concept in skin care ingredients. Cosmet. Toilet. 1991, 106, 69–76. [Google Scholar]
- Lu, M.; Qiu, Q.; Luo, X.; Liu, X.; Sun, J.; Wang, C.; Lin, X.; Deng, Y.; Song, Y. Phyto-phospholipid complexes (phytosomes): A novel strategy to improve the bioavailability of active constituents. Asian J. Pharm. Sci. 2019, 14, 265–274. [Google Scholar] [CrossRef]
- Azeez, N.A.; Deepa, V.S.; Sivapriya, V. Phytosomes: Emergent promising nano vesicular drug delivery system for targeted tumor therapy. Adv. Nat. Sci. Nanosci. Nanotechnol. 2018, 9, 33001. [Google Scholar] [CrossRef]
- Xu, L.; Xu, D.; Li, Z.; Gao, Y.; Chen, H. Synthesis and potent cytotoxic activity of a novel diosgenin derivative and its phytosomes against lung cancer cells. Beilstein J. Nanotechnol. 2019, 10, 1933–1942. [Google Scholar] [CrossRef] [PubMed]
- Aboul-Enein, H.Y.; Abou-Basha, L.I. Simple HPLC Method for the Determination of Thymoquinone in Black Seed Oil (Nigella Sativa Linn). J. Liq. Chromatogr. 1995, 18, 895–902. [Google Scholar] [CrossRef]
- Anwar, M.M.; Abd El-Karim, S.S.; Mahmoud, A.H.; Amr, A.E.-G.E.; Al-Omar, M.A. A Comparative Study of the Anticancer Activity and PARP-1 Inhibiting Effect of Benzofuran-Pyrazole Scaffold and Its Nano-Sized Particles in Human Breast Cancer Cells. Molecules 2019, 24, 2413. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Cun, X.; Ruan, S.; Shi, K.; Wang, Y.; Kuang, Q.; Hu, C.; Xiao, W.; He, Q.; Gao, H. Utilizing G2/M retention effect to enhance tumor accumulation of active targeting nanoparticles. Sci. Rep. 2016, 6, 27669. [Google Scholar] [CrossRef]
- Hsiao, K.Y.; Wu, Y.-J.; Liu, Z.N.; Chuang, C.W.; Huang, H.H.; Kuo, S.M. Anticancer Effects of Sinulariolide-Conjugated Hyaluronan Nanoparticles on Lung Adenocarcinoma Cells. Molecules 2016, 21, 297. [Google Scholar] [CrossRef]
- Czarnomysy, R.; Surażyński, A.; Muszynska, A.; Gornowicz, A.; Bielawska, A.; Bielawski, K. A novel series of pyrazole-platinum(II) complexes as potential anti-cancer agents that induce cell cycle arrest and apoptosis in breast cancer cells. J. Enzyme Inhib. Med. Chem. 2018, 33, 1006–1023. [Google Scholar] [CrossRef]
- Chen, X.; Zhong, Z.; Xu, Z.; Chen, L.; Wang, Y. 2′,7′-Dichlorodihydrofluorescein as a fluorescent probe for reactive oxygen species measurement: Forty years of application and controversy. Free Radic. Res. 2010, 44, 587–604. [Google Scholar] [CrossRef]
- Ramkumar, M.; Rajasankar, S.; Gobi, V.V.; Dhanalakshmi, C.; Manivasagam, T.; Justin Thenmozhi, A.; Essa, M.M.; Kalandar, A.; Chidambaram, R. Neuroprotective effect of Demethoxycurcumin, a natural derivative of Curcumin on rotenone induced neurotoxicity in SH-SY 5Y Neuroblastoma cells. BMC Complement. Altern. Med. 2017, 17, 1–11. [Google Scholar]
- Jiang, Y.; Yu, X.; Su, C.; Zhao, L.; Shi, Y. Chitosan nanoparticles induced the antitumor effect in hepatocellular carcinoma cells by regulating ROS-mediated mitochondrial damage and endoplasmic reticulum stress. Artif. Cells Nanomed. Biotechnol. 2019, 47, 747–756. [Google Scholar] [CrossRef]
- Dubey, V.; Mishra, D.; Dutta, T.; Nahar, M.; Saraf, D.K.; Jain, N.K. Dermal and transdermal delivery of an anti-psoriatic agent via ethanolic liposomes. J. Control. Release 2007, 123, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Saoji, S.D.; Raut, N.A.; Dhore, P.W.; Borkar, C.D.; Popielarczyk, M.; Dave, V.S. Preparation and Evaluation of Phospholipid-Based Complex of Standardized Centella Extract (SCE) for the Enhanced Delivery of Phytoconstituents. AAPS J. 2016, 18, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Li, Y.; Huang, Y.; Zhou, C.; Lin, J.; Wang, Y.; Cui, F.; Zhou, S.; Jia, M.; Ye, S.; et al. Phytosomes loaded with mitomycin C-soybean phosphatidylcholine complex developed for drug delivery. Mol. Pharm. 2013, 10, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Acharya, B.R.; Chatterjee, A.; Ganguli, A.; Bhattacharya, S.; Chakrabarti, G. Thymoquinone inhibits microtubule polymerization by tubulin binding and causes mitotic arrest following apoptosis in A549 cells. Biochimie 2014, 97, 78–91. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, L.; Mollica, M.; Re, A.T.; Wu, S.; Zuo, L. Nitric oxide in cancer metastasis. Cancer Lett. 2014, 353, 1–7. [Google Scholar] [CrossRef]
- Fahmy, U.A.; Badr-Eldin, S.M.; Ahmed, O.A.A.; Aldawsari, H.M.; Tima, S.; Asfour, H.Z.; Al-Rabia, M.W.; Negm, A.A.; Sultan, M.H.; Madkhali, O.A.A.; et al. Intranasal Niosomal In Situ Gel as a Promising Approach for Enhancing Flibanserin Bioavailability and Brain Delivery: In Vitro Optimization and Ex Vivo/In Vivo Evaluation. Pharmaceutics 2020, 12, 485. [Google Scholar] [CrossRef]
- Ahmed, O.A.A.; Fahmy, U.A.; Bakhaidar, R.; El-Moselhy, M.A.; Okbazghi, S.Z.; Ahmed, A.-S.F.; Hammad, A.S.A.; Alhakamy, N.A. Omega-3 Self-Nanoemulsion Role in Gastroprotection against Indomethacin-Induced Gastric Injury in Rats. Pharmaceutics 2020, 12, 140. [Google Scholar] [CrossRef]
- Alhakamy, N.A.; Badr-Eldin, S.M.; Ahmed, O.A.A.; Asfour, H.Z.; Aldawsari, H.M.; Algandaby, M.M.; Eid, B.G.; Abdel-Naim, A.B.; Awan, Z.A.; Alghaith, A.F.; et al. Piceatannol-Loaded Emulsomes Exhibit Enhanced Cytostatic and Apoptotic Activities in Colon Cancer Cells. Antioxidants 2020, 9, 419. [Google Scholar] [CrossRef]
- Alhakamy, N.A.; Ahmed, O.A.A.; Kurakula, M.; Caruso, G.; Caraci, F.; Asfour, H.Z.; Alfarsi, A.; Eid, B.G.; Mohamed, A.I.; Alruwaili, N.K.; et al. Chitosan-Based Microparticles Enhance Ellagic Acid’s Colon Targeting and Proapoptotic Activity. Pharmaceutics 2020, 12, 652. [Google Scholar] [CrossRef]
- Park, E.J.; Chauhan, A.K.; Min, K.J.; Park, D.C.; Kwon, T.K. Thymoquinone induces apoptosis through downregulation of c-FLIP and Bcl-2 in Renal carcinoma Caki cells. Oncol. Rep. 2016, 36, 2261–2267. [Google Scholar] [CrossRef]
- Gali-Muhtasib, H.U.; Abou Kheir, W.G.; Kheir, L.A.; Darwiche, N.; Crooks, P.A. Molecular pathway for thymoquinone-induced cell-cycle arrest and apoptosis in neoplastic keratinocytes. Anticancer. Drugs 2004, 15, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of reactive oxygen species in cancer progression: Molecular mechanisms and recent advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run Number | Independent Variables | Dependent Variable | ||
|---|---|---|---|---|
| A (Mole/Mole) | B (°C) | C (h) | Y (nm) | |
| R1 | 1:3 | 50.00 | 2.00 | 80.7 |
| R2 | 1:3 | 65.00 | 1.00 | 54.9 |
| R3 | 1:3 | 50.00 | 2.00 | 82.2 |
| R4 | 1:3 | 50.00 | 2.00 | 81.7 |
| R5 | 1:3 | 65.00 | 3.00 | 64.7 |
| R6 | 1:3 | 35.00 | 1.00 | 112.8 |
| R7 | 1:3 | 35.00 | 3.00 | 114.7 |
| R8 | 1:1 | 50.00 | 3.00 | 71.8 |
| R9 | 1:3 | 50.00 | 2.00 | 82.2 |
| R10 | 1:5 | 50.00 | 3.00 | 101.8 |
| R11 | 1:5 | 65.00 | 2.00 | 69.7 |
| R12 | 1:1 | 35.00 | 2.00 | 98.8 |
| R13 | 1:5 | 50.00 | 1.00 | 95.2 |
| R14 | 1:5 | 35.00 | 2.00 | 129.9 |
| R15 | 1:1 | 65.00 | 2.00 | 43.7 |
| R16 | 1:3 | 50.00 | 2.00 | 79.8 |
| R17 | 1:1 | 50.00 | 1.00 | 60.4 |
| Independent variable | Levels | |||
| Low (−1) | Medium (0) | High (1) | ||
| (0) | (−1) | (+1) | ||
| TQ to phospholipid molar ratio (A) | 1:1 | 1:3 | 1:5 | |
| Process temperature (°C, B) | 35 | 50 | 65 | |
| Reflux time (h, C) | 1 | 2 | 3 | |
| Dependent variable | Goal | |||
| Vesicle Size (Y, nm) | Minimize | |||
| Run | Mean Vesicle Size (nm) | |||
|---|---|---|---|---|
| Actual Values | Predicted Values | Residual | % Error | |
| R1 | 80.7 | 81.32 | −0.62 | 0.76% |
| R2 | 54.9 | 53.19 | 1.71 | 3.21% |
| R3 | 82.2 | 81.32 | 0.88 | 1.08% |
| R4 | 81.7 | 81.32 | 0.38 | 0.47% |
| R5 | 64.7 | 64.56 | 0.14 | 0.22% |
| R6 | 112.8 | 112.94 | −0.14 | 0.12% |
| R7 | 114.7 | 116.41 | −1.71 | 1.47% |
| R8 | 71.8 | 71.97 | −0.17 | 0.24% |
| R9 | 82.2 | 81.32 | 0.88 | 1.08% |
| R10 | 101.8 | 100.05 | 1.75 | 1.75% |
| R11 | 69.7 | 71.59 | −1.89 | 2.64% |
| R12 | 98.8 | 96.91 | 1.89 | 1.95% |
| R13 | 95.2 | 95.02 | 0.18 | 0.19% |
| R14 | 129.9 | 129.94 | −0.04 | 0.03% |
| R15 | 43.7 | 43.66 | 0.04 | 0.09% |
| R16 | 79.8 | 81.32 | −1.52 | 1.87% |
| R17 | 60.4 | 62.15 | −1.75 | 2.82% |
| Source | Sum of Squares | Degree(s) of Freedom | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 8309.905 | 9 | 923.323 | 273.803 | <0.0001 * |
| A-PL | 1857.451 | 1 | 1857.451 | 550.811 | <0.0001 * |
| B-Temp. | 6227.28 | 1 | 6227.280 | 1846.644 | <0.0001 * |
| C-Time | 110.2613 | 1 | 110.261 | 32.697 | 0.0007 * |
| AB | 6.5025 | 1 | 6.503 | 1.928 | 0.2075 |
| AC | 5.76 | 1 | 5.760 | 1.708 | 0.2325 |
| BC | 15.6025 | 1 | 15.603 | 4.627 | 0.0485 * |
| AÂ2 | 0.076737 | 1 | 0.077 | 0.023 | 0.8843 |
| BÂ2 | 79.30779 | 1 | 79.308 | 23.518 | 0.0019 * |
| CÂ2 | 5.234632 | 1 | 5.235 | 1.552 | 0.2529 |
| Residual | 23.6055 | 7 | 3.372 | ||
| Lack of Fit | 19.2175 | 3 | 6.406 | 5.839 | 0.0606 |
| Pure Error | 4.388 | 4 | 1.097 | ||
| Cor Total | 8333.511 | 16 |
| Factor | Optimized Level/Predicted Response | |
|---|---|---|
| Independent | A | 1:1.4 |
| B | 64.83 | |
| C | 1.82 | |
| Dependent | Y (nm) | 43.67 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhakamy, N.A.; Badr-Eldin, S.M.; Fahmy, U.A.; Alruwaili, N.K.; Awan, Z.A.; Caruso, G.; Alfaleh, M.A.; Alaofi, A.L.; Arif, F.O.; Ahmed, O.A.A.; et al. RETRACTED: Thymoquinone-Loaded Soy-Phospholipid-Based Phytosomes Exhibit Anticancer Potential against Human Lung Cancer Cells. Pharmaceutics 2020, 12, 761. https://doi.org/10.3390/pharmaceutics12080761
Alhakamy NA, Badr-Eldin SM, Fahmy UA, Alruwaili NK, Awan ZA, Caruso G, Alfaleh MA, Alaofi AL, Arif FO, Ahmed OAA, et al. RETRACTED: Thymoquinone-Loaded Soy-Phospholipid-Based Phytosomes Exhibit Anticancer Potential against Human Lung Cancer Cells. Pharmaceutics. 2020; 12(8):761. https://doi.org/10.3390/pharmaceutics12080761
Chicago/Turabian StyleAlhakamy, Nabil A., Shaimaa M. Badr-Eldin, Usama A. Fahmy, Nabil K. Alruwaili, Zuhier A. Awan, Giuseppe Caruso, Mohamed A. Alfaleh, Ahmed L. Alaofi, Faris O Arif, Osama A. A. Ahmed, and et al. 2020. "RETRACTED: Thymoquinone-Loaded Soy-Phospholipid-Based Phytosomes Exhibit Anticancer Potential against Human Lung Cancer Cells" Pharmaceutics 12, no. 8: 761. https://doi.org/10.3390/pharmaceutics12080761