Characterization, Stability, and In Vivo Efficacy Studies of Recombinant Human CNTF and Its Permeation into the Neural Retina in Ex Vivo Organotypic Retinal Explant Culture Models

 ,
,  ,
,  and
and 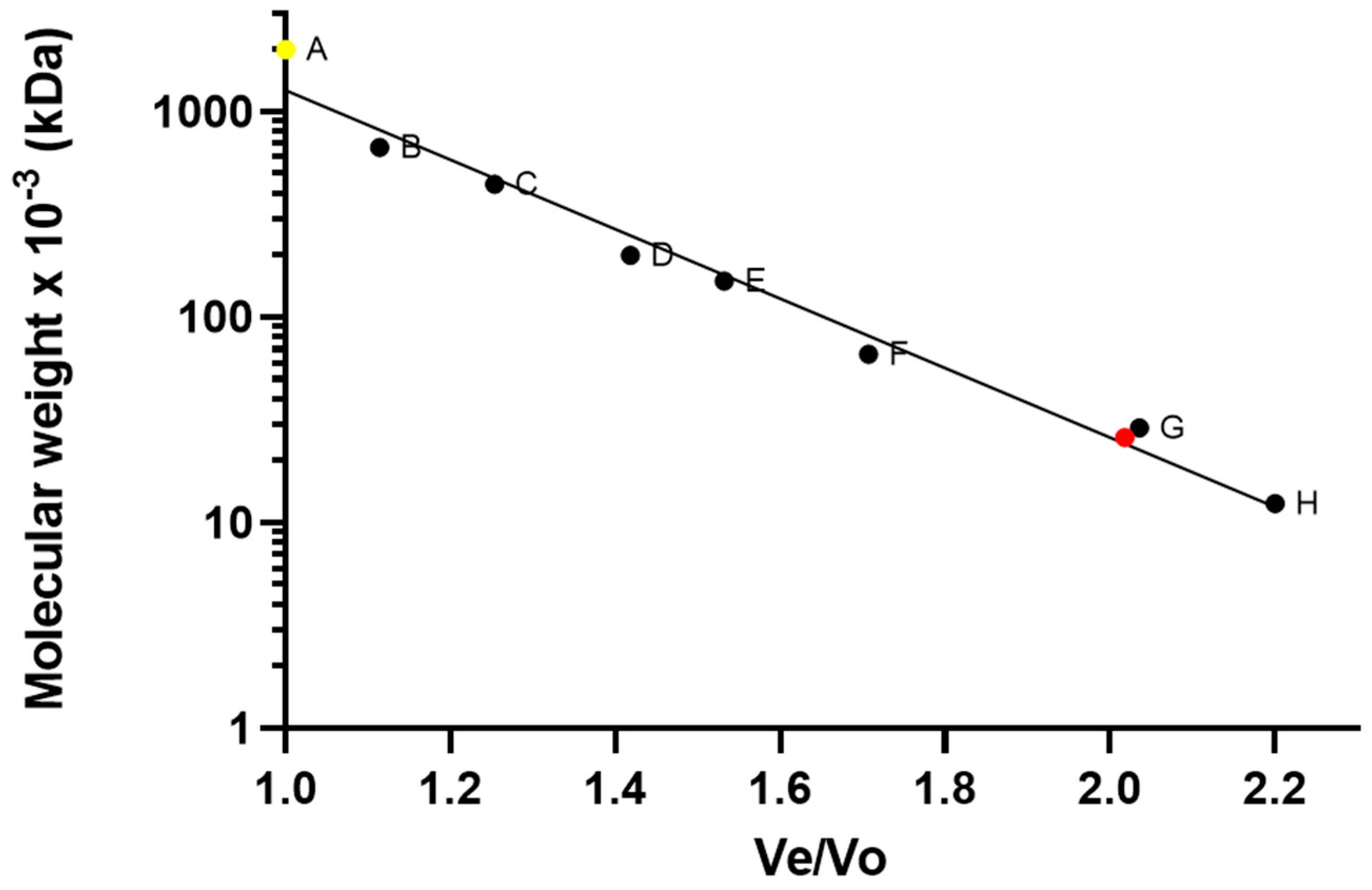{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Production
2.2. rhCNTF In Vitro Bioactivity Study
2.2.1. Cell Culture
2.2.2. Cell Proliferation Assay
2.3. rhCNTF Characterization and Stability Studies
2.3.1. Circular Dichroism
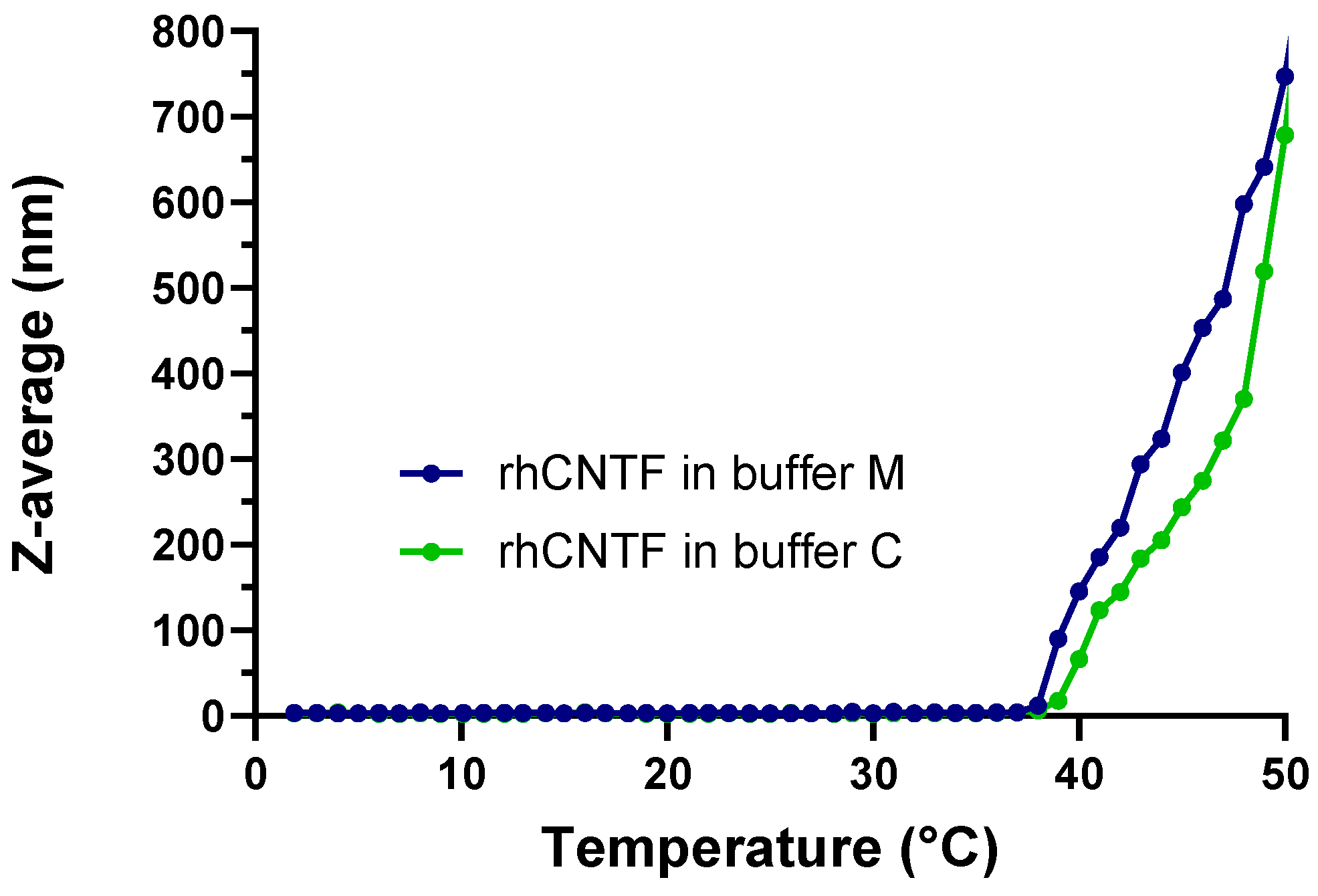
2.3.2. ThermoFluor
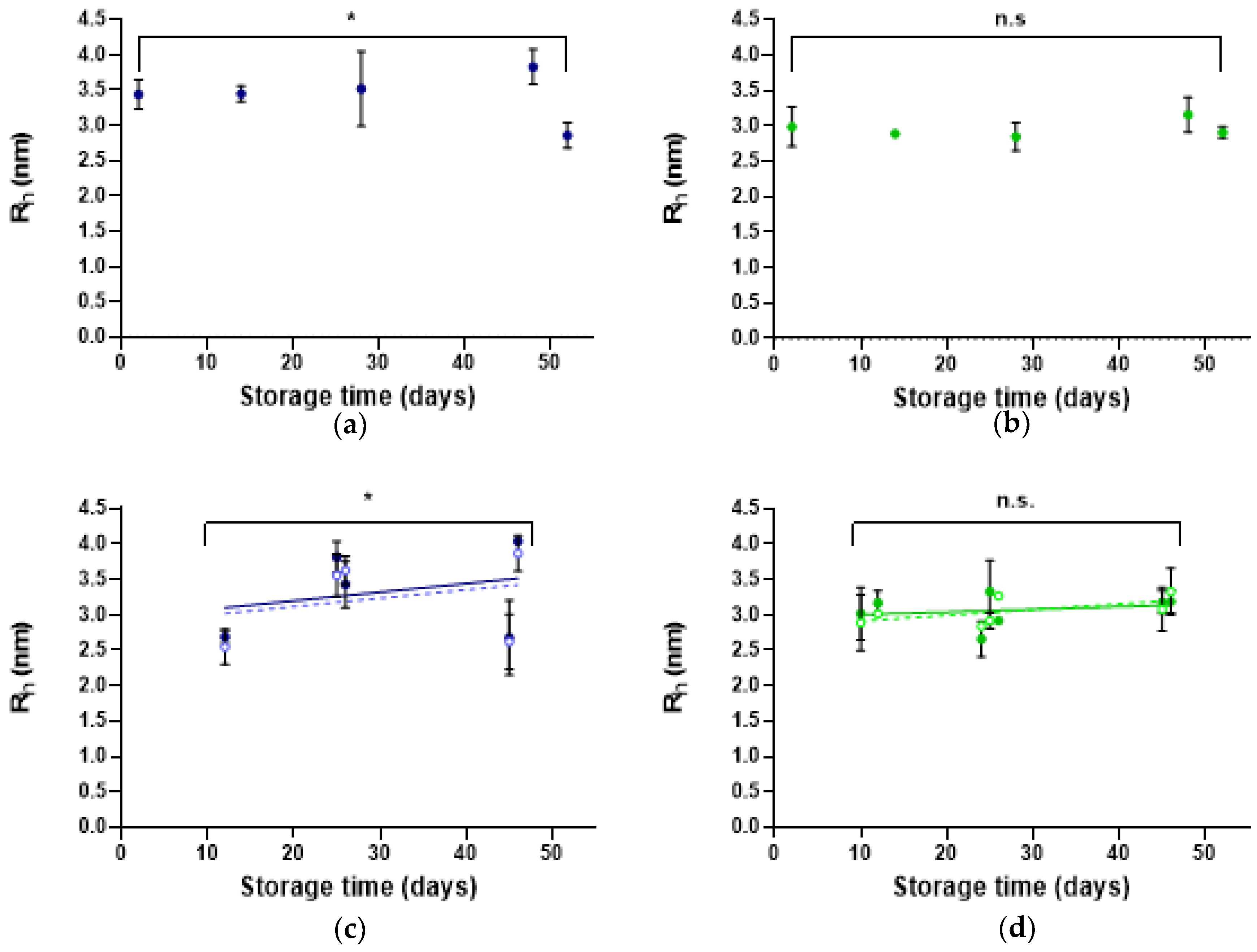
2.3.3. Dynamic Light Scattering
2.4. rhCNTF In Vivo Efficacy Study
2.4.1. Animals
2.4.2. Intravitreal Injections
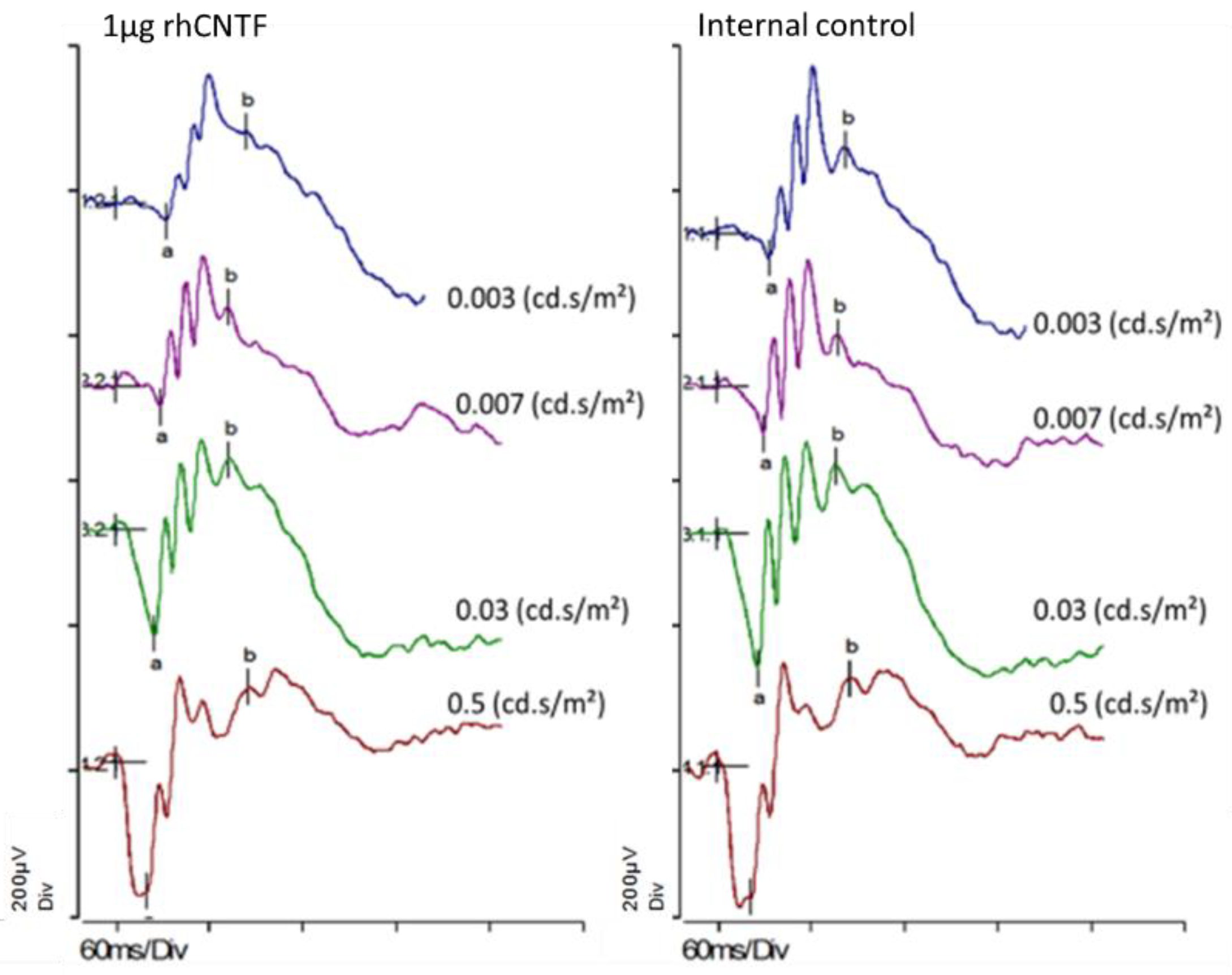
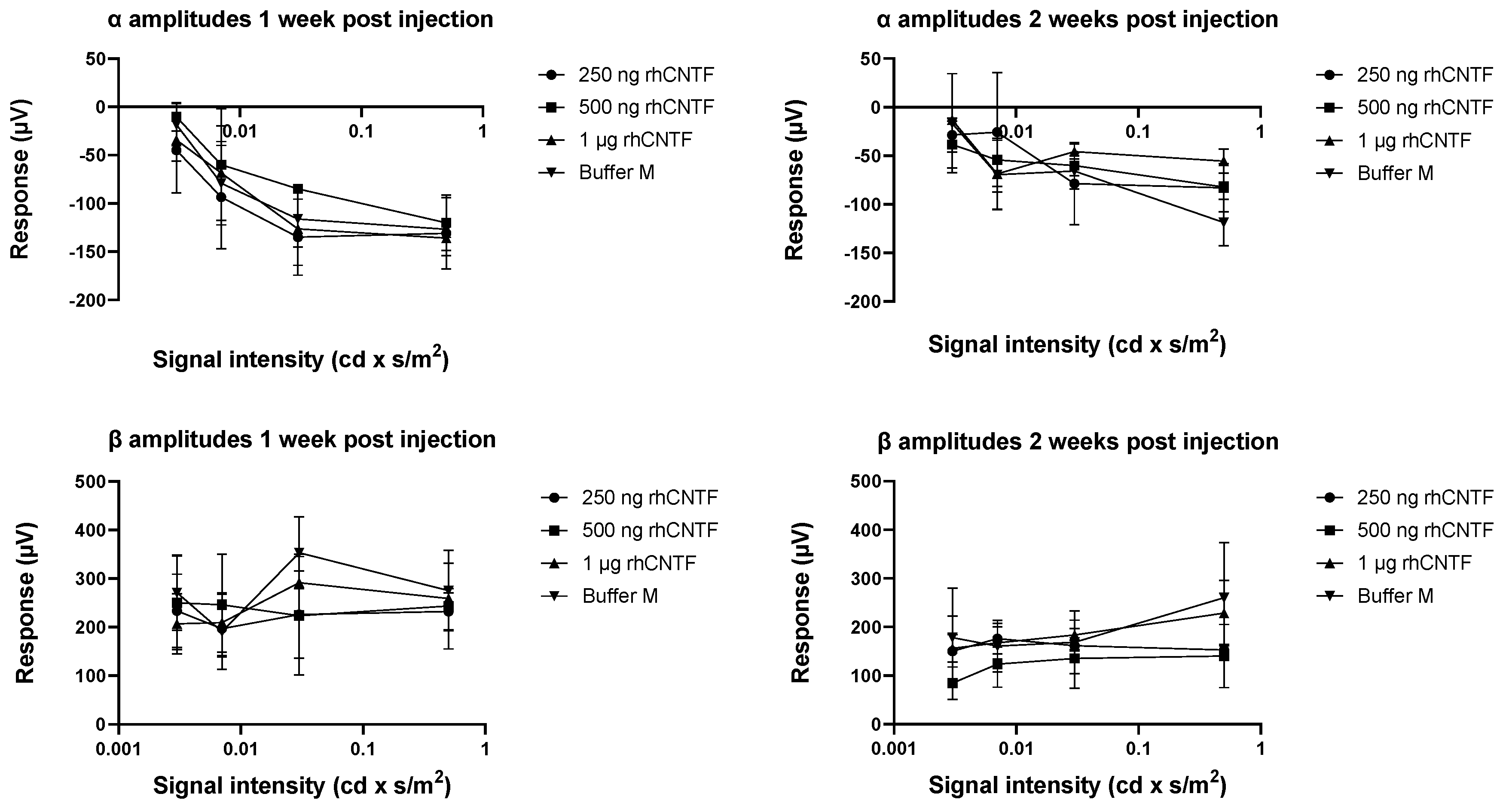
2.4.3. Electroretinogram (ERG) Recording
2.4.4. Data-Analysis
2.4.5. Histology
2.5. Retinal Penetration of rhCNTF
2.5.1. rhCNTF Fluorescent Labeling
2.5.2. Retinal Explant Culture Preparation and rhCNTF Treatment
Rat Retinal Explants
Bovine Retinal Explants
2.5.3. Tissue Culture Fixation and Sectioning
Rat Retinal Explants
Bovine Retinal Explants
2.5.4. Culture Staining and Imaging
Rat Retinal Explants
Bovine Retinal Explants
3. Results
3.1. Protein Production
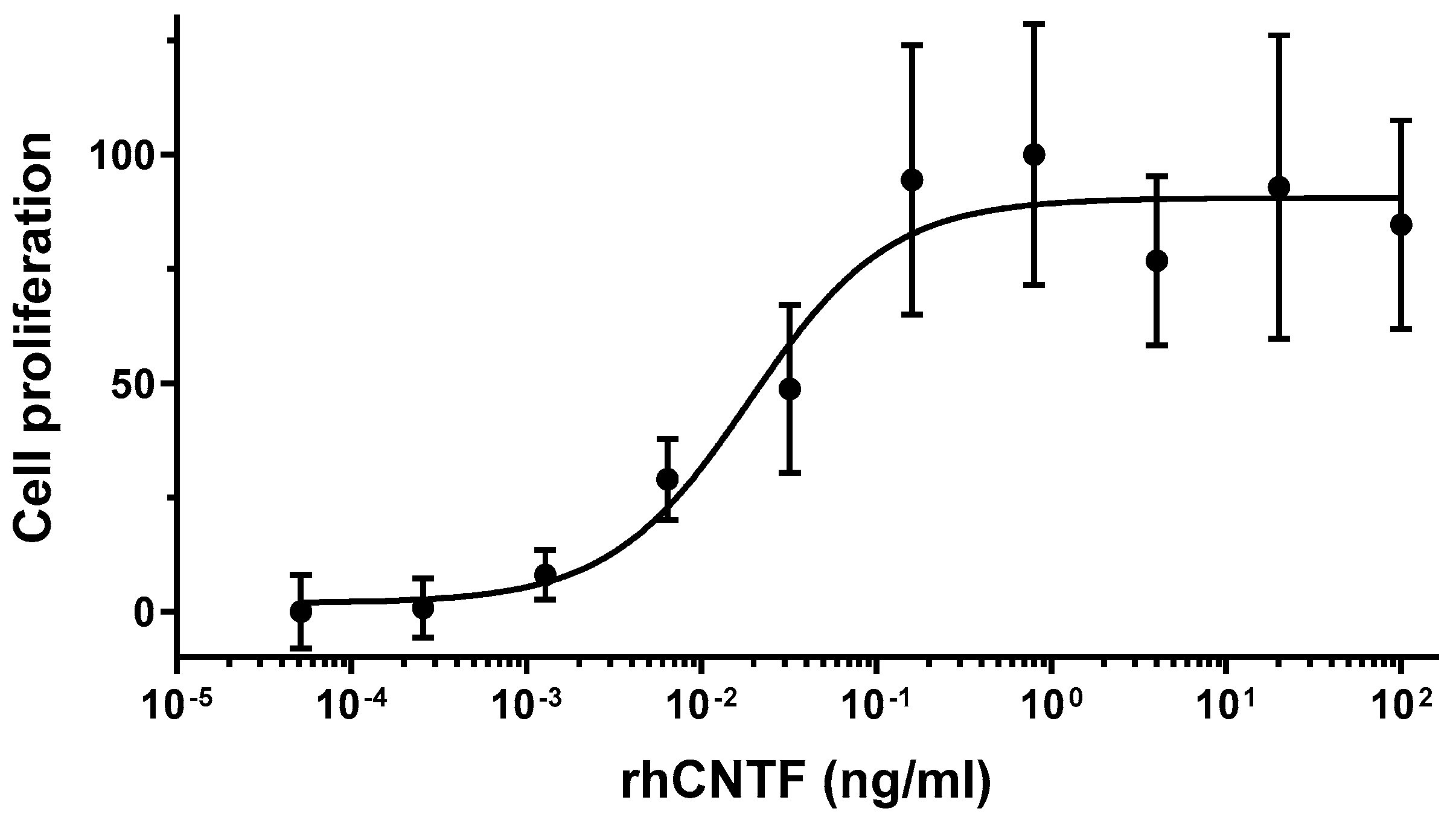
3.2. rhCNTF In Vitro Bioactivity
3.3. Characterization and Stability of rhCNTF
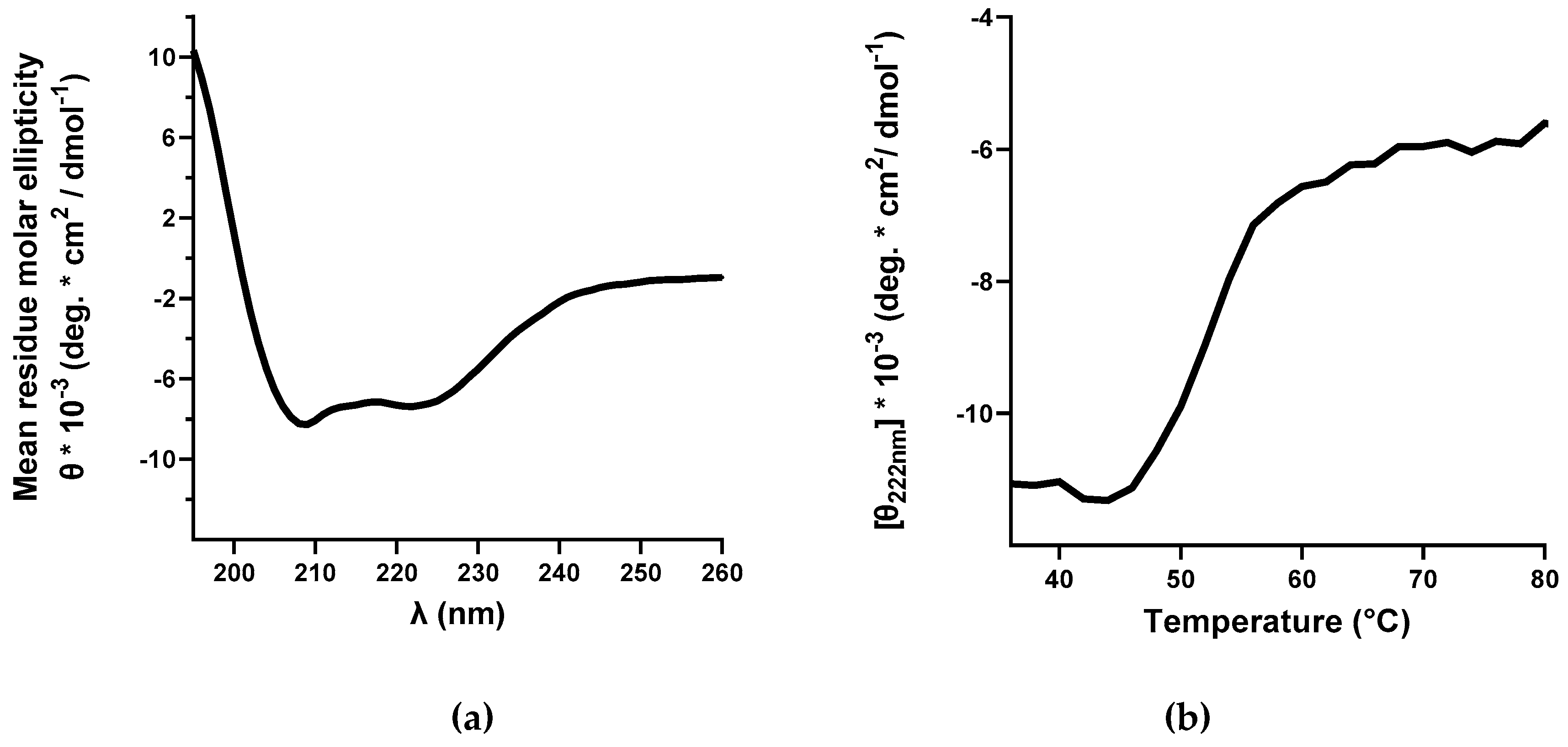
3.3.1. Circular Dichroism
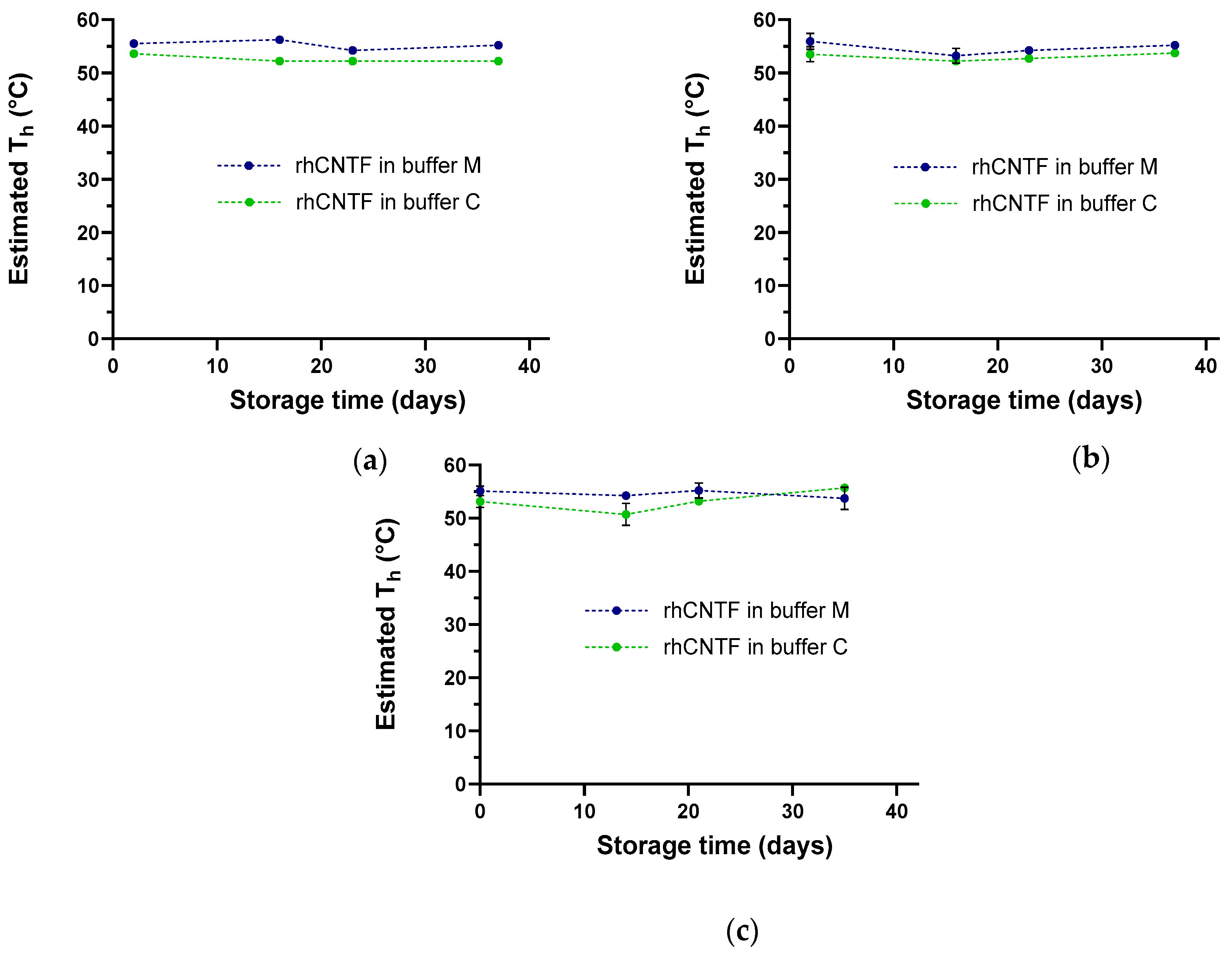
3.3.2. ThermoFluor
3.3.3. Dynamic Light Scattering
3.4. rhCNTF In Vivo Efficacy Study
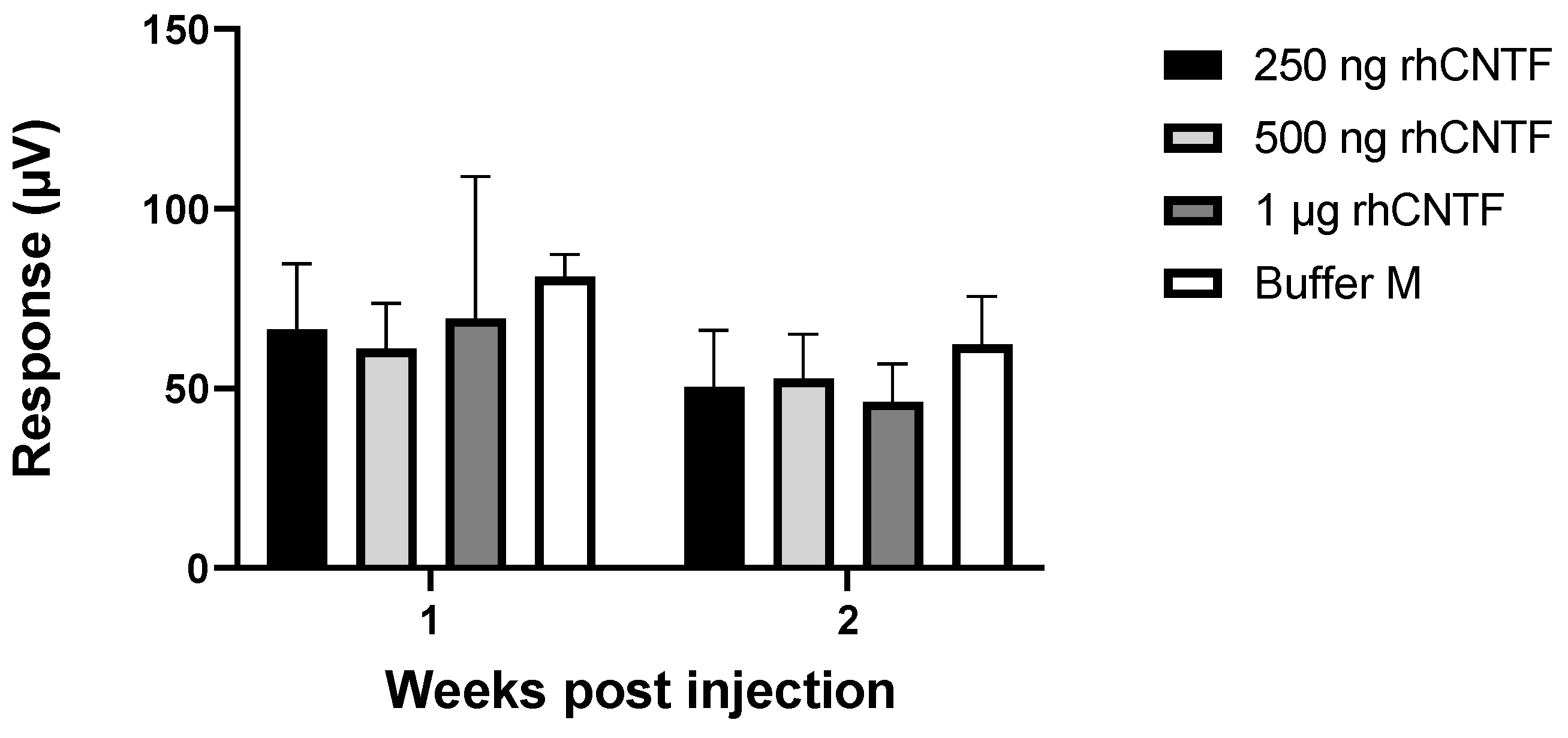
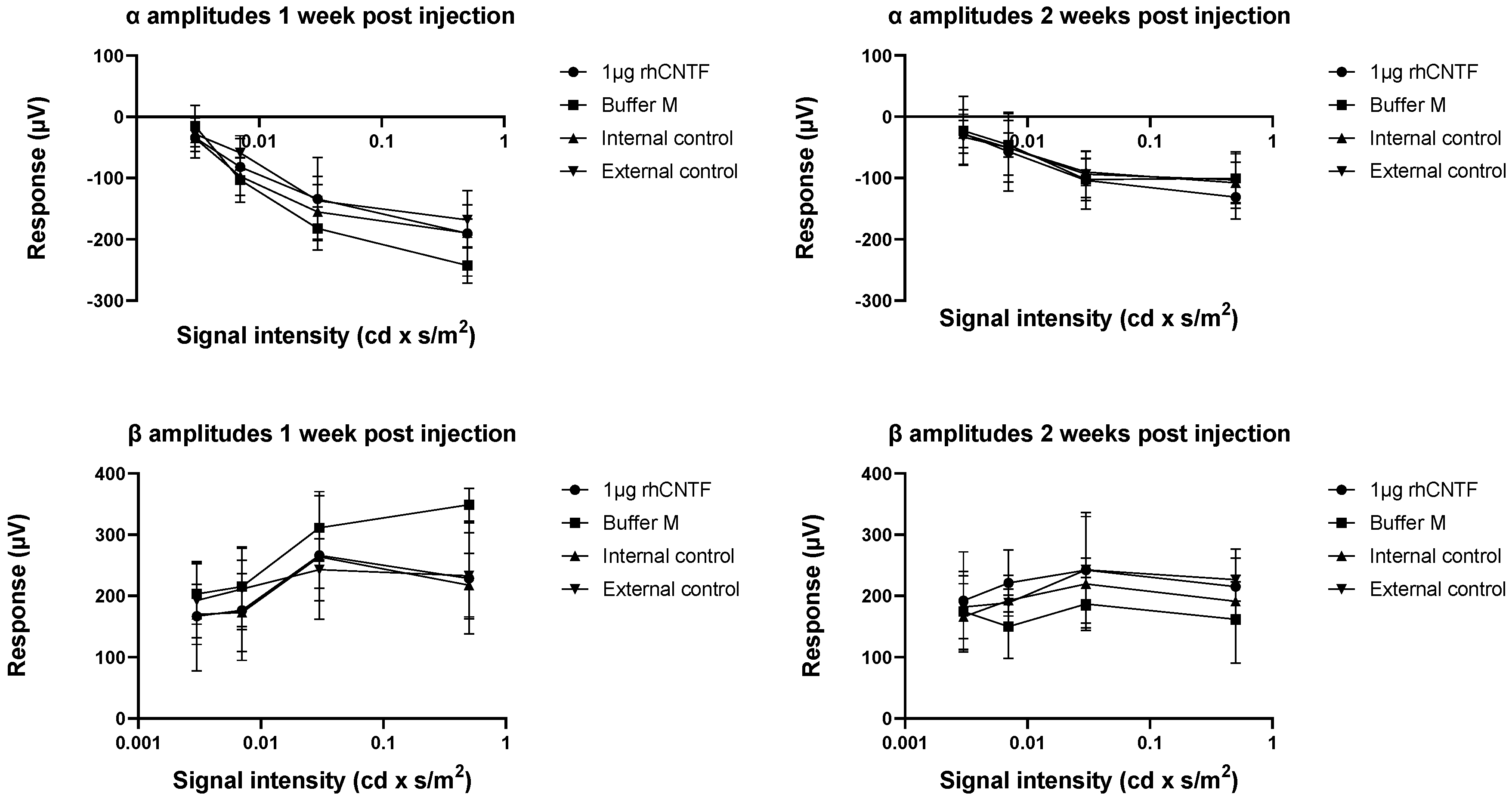
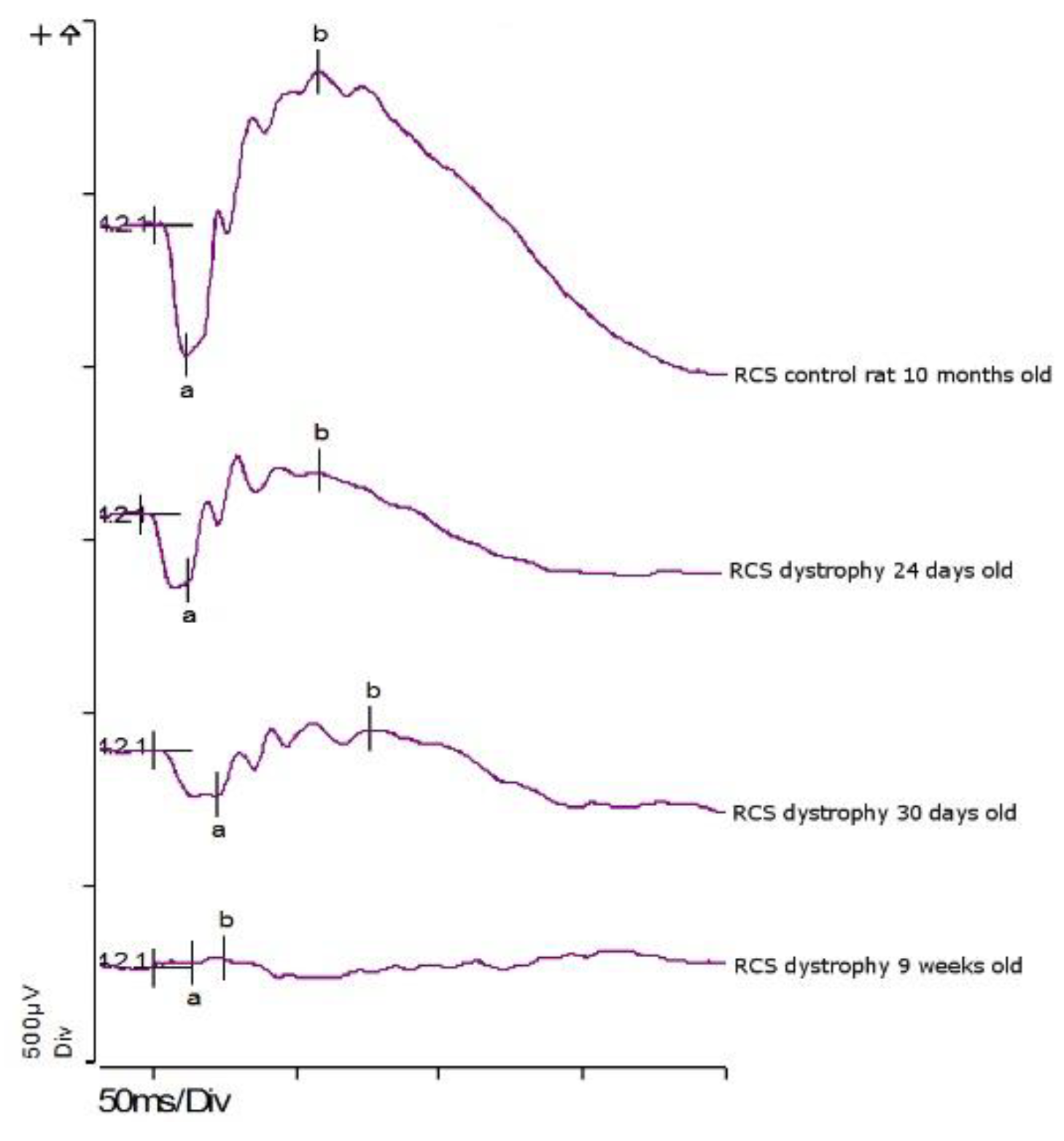
3.4.1. ERG: First Study Set
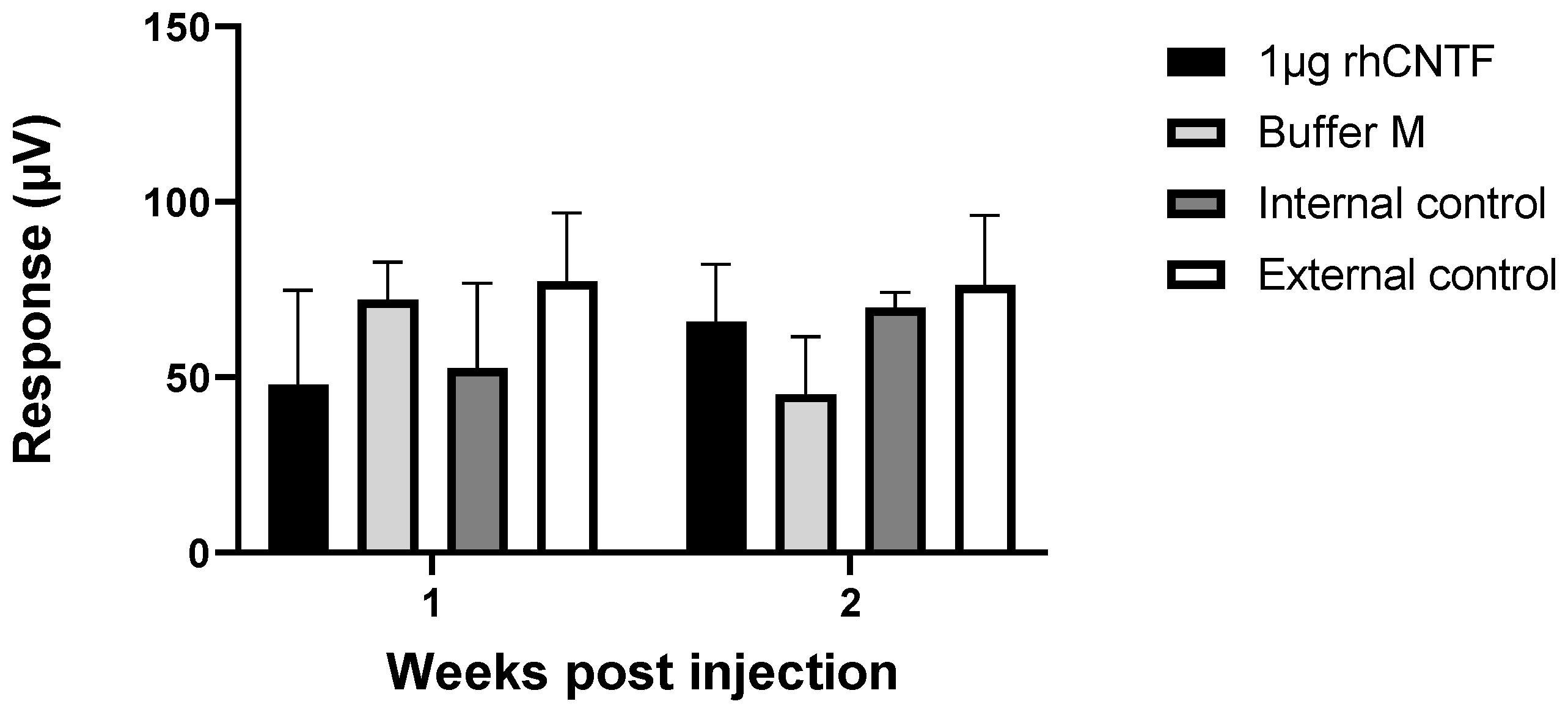
3.4.2. ERG: Second Study Set
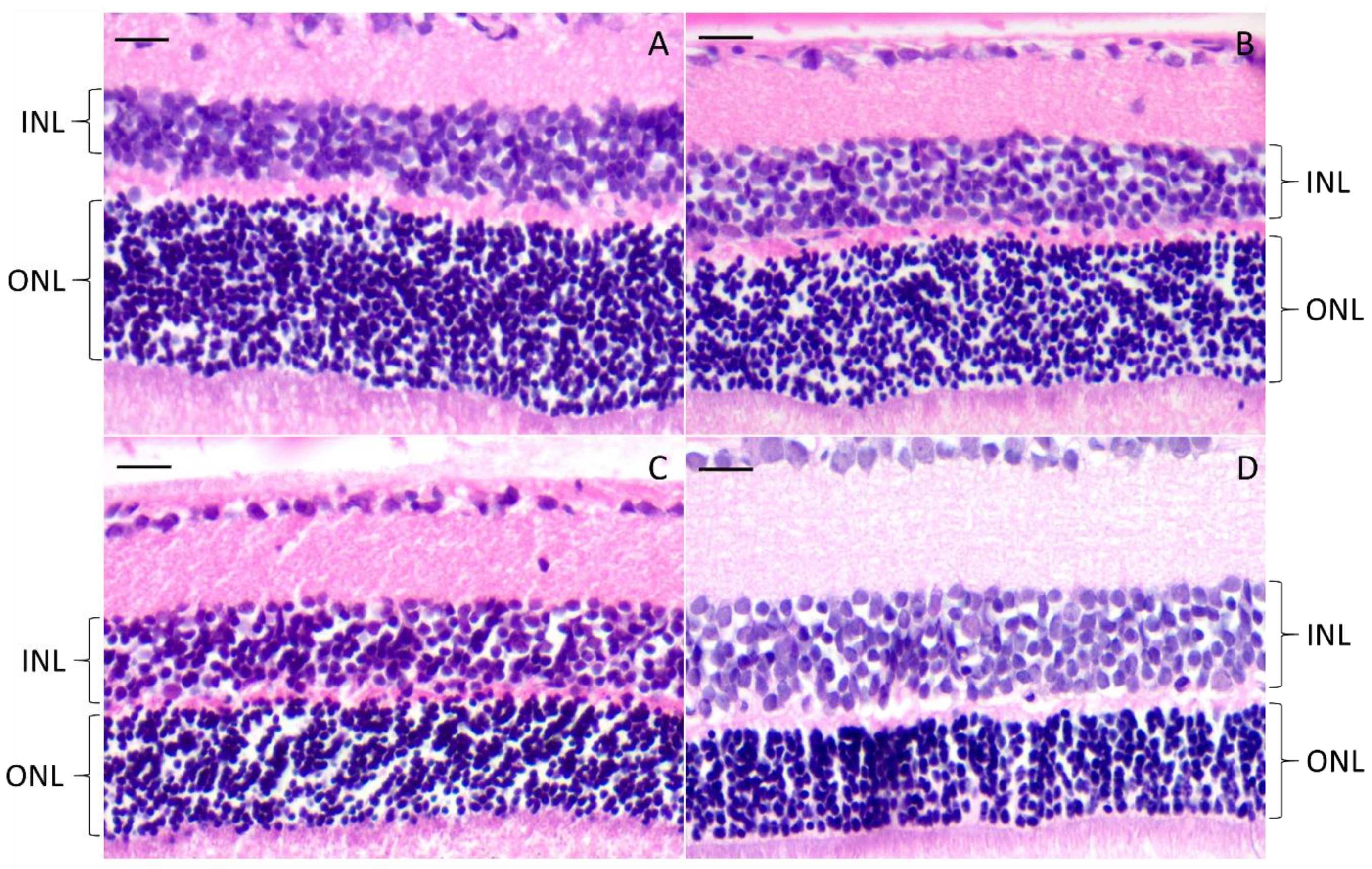
3.4.3. Histology
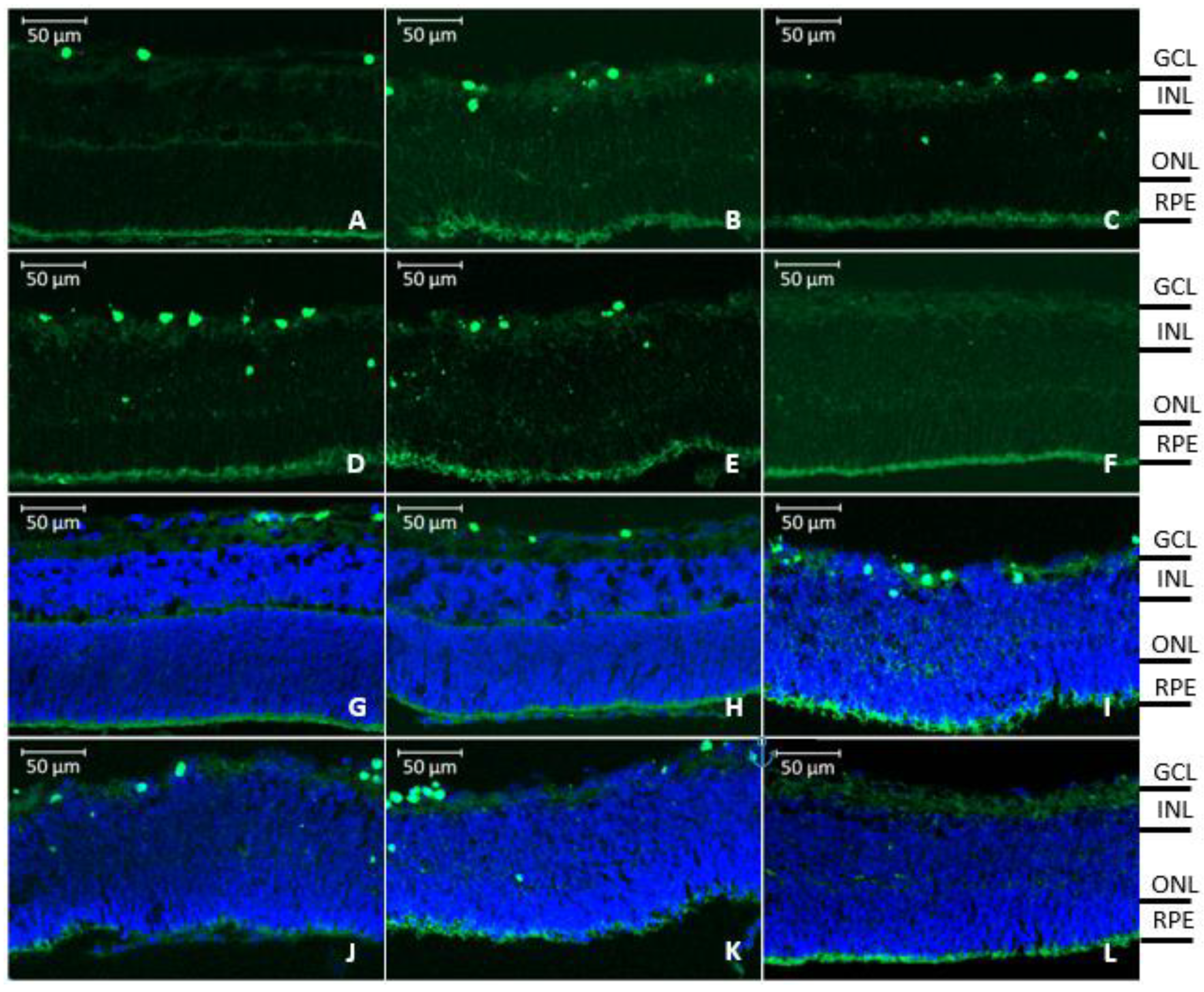
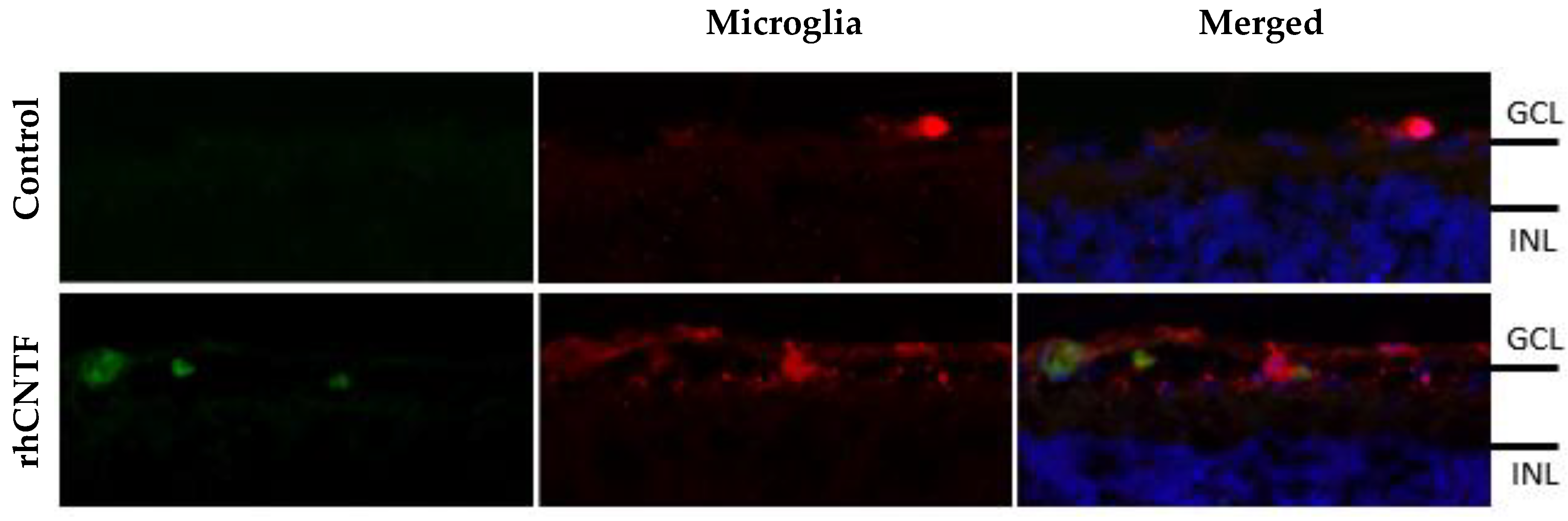
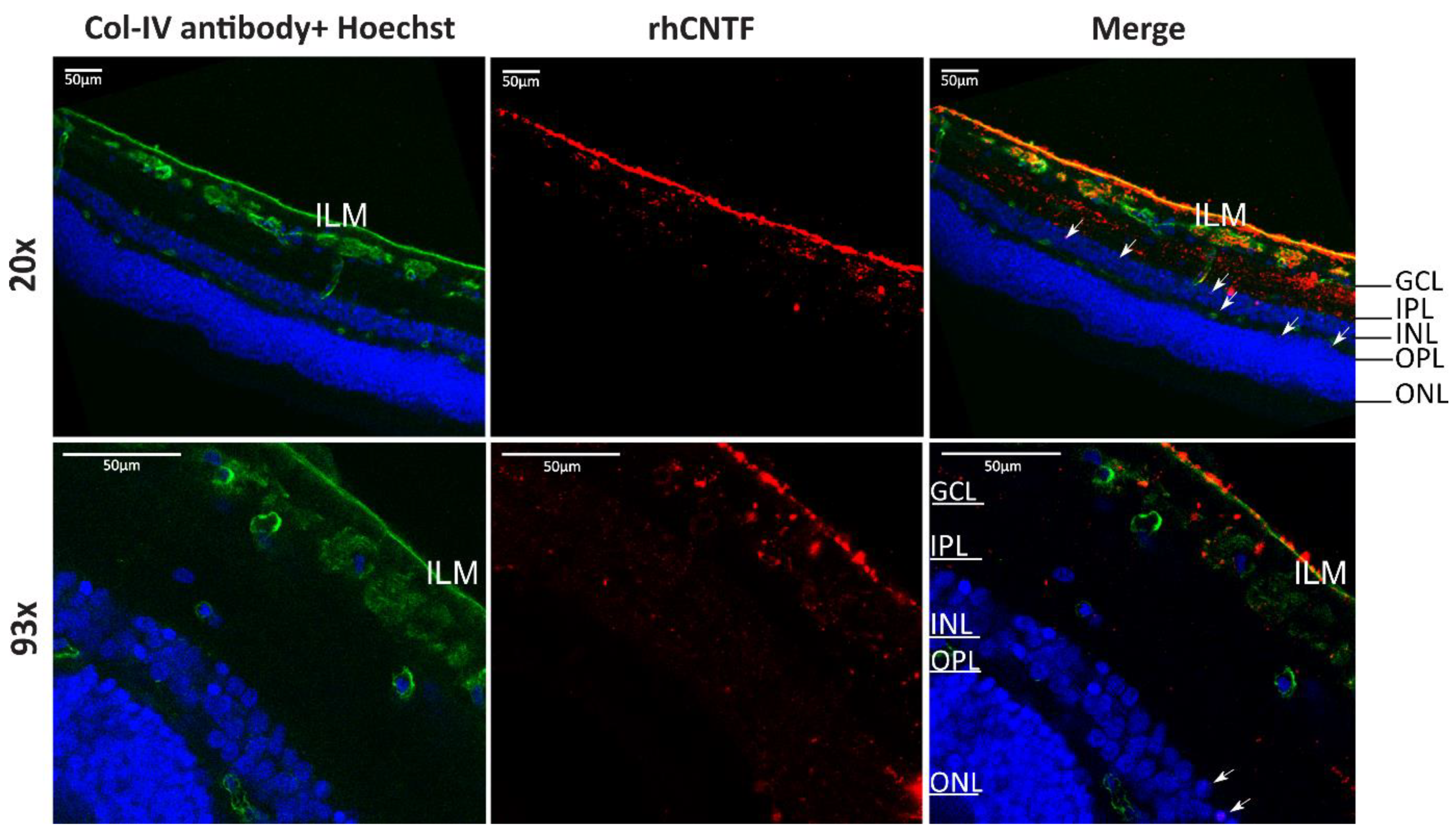
3.5. Retinal Penetration of Labeled rhCNTF
4. Discussion
4.1. Buffer Screening and Stability Studies
4.2. In Vivo Bioactivity of rhCNTF
4.3. Retinal Penetration and Effects of rhCNTF
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mandal, A.; Pal, D.; Agrahari, V.; Trinh, H.M.; Joseph, M.; Mitra, A.K. Ocular delivery of proteins and peptides: Challenges and novel formulation approaches. Adv. Drug Deliv. Rev. 2018, 126, 67–95. [Google Scholar] [CrossRef] [PubMed]
- Awwad, S.; Ahmed, A.H.M.; Sharma, G.; Heng, J.S.; Khaw, P.T.; Brocchini, S.; Lockwood, A. Principles of pharmacology in the eye. Br. J. Pharmacol. 2017, 174, 4205–4223. [Google Scholar] [CrossRef] [PubMed]
- Delplace, V.; Payne, S.; Shoichet, M. Delivery strategies for treatment of age-related ocular diseases: From a biological understanding to biomaterial solutions. J. Control. Release 2015, 219, 652–668. [Google Scholar] [CrossRef] [PubMed]
- Pardue, M.T.; Allen, R.S. Neuroprotective strategies for retinal disease. Prog. Retin. Eye Res. 2018, 65, 50–76. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, I.M.; Sauvé, Y.; Sieving, P.A. Preventing blindness in retinal disease: Ciliary neurotrophic factor intraocular implants. Can. J. Ophthal. 2007, 42, 399–402. [Google Scholar] [CrossRef]
- Kimura, A.; Namekata, K.; Guo, X.; Harada, C.; Harada, T. Neuroprotection, growth factors and BDNF-TrkB signalling in retinal degeneration. Int. J. Mol. Sci. 2016, 17, 1584. [Google Scholar] [CrossRef] [Green Version]
- Unsicker, K. Neurotrophic molecules in the treatment of neurodegenerative disease with focus on the retina: Status and perspectives. Cell Tissue Res. 2013, 353, 205–218. [Google Scholar] [CrossRef]
- Wen, R.; Tao, W.; Li, Y.; Sieving, P.A. CNTF and retina. Prog. Retin. Eye Res. 2012, 31, 136–151. [Google Scholar] [CrossRef] [Green Version]
- Sleeman, M.W.; Anderson, K.D.; Lambert, P.D.; Yancopoulos, G.D.; Wiegand, S.J. The ciliary neurotrophic factor and its receptor, CNTFRα. Pharm. Acta Helv. 2000, 74, 265–272. [Google Scholar] [CrossRef]
- Wagener, E.M.; Aurich, M.; Aparicio-Siegmund, S.; Floss, D.M.; Garbers, C.; Breusing, K.; Rabe, B.; Schwanbeck, R.; Grotzinger, J.; Rose-John, S. The amino acid exchange R28E in ciliary neurotrophic factor (CNTF) abrogates interleukin-6 receptor-dependent but retains CNTF receptor-dependent signaling via glycoprotein 130 (gp130)/leukemia inhibitory factor receptor (LIFR). J. Biol. Chem. 2014, 289, 18442–18450. [Google Scholar] [CrossRef] [Green Version]
- Schuster, B.; Kovaleva, M.; Sun, Y.; Regenhard, P.; Matthews, V.; Grotzinger, J.; Rose-John, S.; Kallen, K.J. Signaling of human ciliary neurotrophic factor (CNTF) revisited. The interleukin-6 receptor can serve as an alpha-receptor for CTNF. J. Biol. Chem. 2003, 278, 9528–9535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Wen, R.; Banzon, T.; Maminishkis, A.; Miller, S.S. CNTF mediates neurotrophic factor secretion and fluid absorption in human retinal pigment epithelium. PLoS ONE 2011, 6, e23148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Sato, K.; Gordon, W.C.; Sendtner, M.; Bazan, N.G.; Jin, M. Ciliary neurotrophic factor (CNTF) protects retinal cone and rod photoreceptors by suppressing excessive formation of the visual pigments. J. Biol. Chem. 2018, 293, 15256–15268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiduschka, P.; Renninger, D.; Fischer, D.; Müller, A.; Hofmeister, S.; Schraermeyer, U. Lens injury has a protective effect on photoreceptors in the RCS rat. ISRN Ophthalmol. 2013, 2013, 814814. [Google Scholar] [CrossRef] [PubMed]
- Peterson, W.M.; Wang, Q.; Tzekova, R.; Wiegand, S.J. Ciliary neurotrophic factor and stress stimuli activate the Jak-STAT pathway in retinal neurons and glia. J. Neurosci. 2000, 20, 4081–4090. [Google Scholar] [CrossRef] [Green Version]
- Beltran, W.; Rohrer, H.; Aguirre, G.D. Immunolocalization of ciliary neurotrophic factor receptor α (CNTFRα) in mammalian photoreceptor cells. Mol. Vis. 2005, 11, 232–244. [Google Scholar]
- Bucher, F.; Walz, J.M.; Bühler, A.; Aguilar, E.; Lange, C.; Diaz-Aguilar, S.; Martin, G.; Schlunck, G.; Agostini, H.; Friedlander, M. CNTF attenuates vasoproliferative changes through upregulation of SOCS3 in a mouse-model of oxygen-induced retinopathy. Investig. Ophthalmol. Vis. Sci. 2016, 57, 4017–4026. [Google Scholar] [CrossRef] [Green Version]
- Rhee, K.D.; Nusinowitz, S.; Chao, K.; Yu, F.; Bok, D.; Yang, X.J. CNTF-mediated protection of photoreceptors requires initial activation of the cytokine receptor gp130 in Müller glial cells. Proc. Natl. Acad. Sci. USA 2013, 110, E4520–E4529. [Google Scholar] [CrossRef] [Green Version]
- Sieving, P.A.; Caruso, R.C.; Tao, W.; Coleman, H.R.; Thompson, D.J.S.; Fullmer, K.R.; Bush, R.A. Ciliary neurotrophic factor (CNTF) for human retinal degeneration: Phase I trial of CNTF delivered by encapsulated cell intraocular implants. Proc. Natl. Acad. Sci. USA 2006, 103, 3896–3901. [Google Scholar] [CrossRef] [Green Version]
- Krady, J.K.; Lin, H.; Liberto, C.M.; Basu, A.; Kremlev, S.G.; Levison, S.W. Ciliary neurotrophic factor and interleukin-6 differentially activate microglia. J. Neurosci. Res. 2008, 86, 1538–1547. [Google Scholar] [CrossRef]
- Baek, J.; Jeong, J.; Kim, K.; Won, S.; Chung, Y.; Nam, J.; Cho, E.; Ahn, T.; Bok, E.; Shin, W. Inhibition of microglia-derived oxidative stress by ciliary neurotrophic factor protects dopamine neurons in vivo from MPP neurotoxicity. Int. J. Mol. Sci. 2018, 19, 3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Jain, M.R.; Li, H.; Levison, S.W. Ciliary neurotrophic factor (CNTF) plus soluble CNTF receptor α increases cyclooxygenase-2 expression, PGE 2 release and interferon-γ-induced CD40 in murine microglia. J. Neuroinflammation 2009, 6, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cen, L.; Luo, J.; Zhang, C.; Fan, Y.; Song, Y.; So, K.; van Rooijen, N.; Pang, C.P.; Lam, D.S.; Cui, Q. Chemotactic effect of ciliary neurotrophic factor on macrophages in retinal ganglion cell survival and axonal regeneration. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4257–4266. [Google Scholar] [CrossRef] [PubMed]
- Van Adel, B.; Arnold, J.; Phipps, J.; Doering, L.; Ball, A. Ciliary neurotrophic factor protects retinal ganglion cells from axotomy-induced apoptosis via modulation of retinal glia in vivo. J. Neurobiol. 2005, 63, 215–234. [Google Scholar] [CrossRef]
- Wen, R.; Song, Y.; Kjellstrom, S.; Tanikawa, A.; Liu, Y.; Li, Y.; Zhao, L.; Bush, R.A.; Laties, A.M.; Sieving, P.A. Regulation of rod phototransduction machinery by ciliary neurotrophic factor. J. Neurosci. 2006, 26, 13523–13530. [Google Scholar] [CrossRef] [Green Version]
- Harada, T.; Harada, C.; Kohsaka, S.; Wada, E.; Yoshida, K.; Ohno, S.; Mamada, H.; Tanaka, K.; Parada, L.F.; Wada, K. Microglia-Müller glia cell interactions control neurotrophic factor production during light-induced retinal degeneration. J. Neurosci. 2002, 22, 9228–9236. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.; Hauk, T.G.; Leibinger, M.; Marienfeld, R.; Fischer, D. Exogenous CNTF stimulates axon regeneration of retinal ganglion cells partially via endogenous CNTF. Mol. Cell Neurosci. 2009, 41, 233–246. [Google Scholar] [CrossRef]
- Wahlin, K.J.; Campochiaro, P.A.; Zack, D.J.; Adler, R. Neurotrophic factors cause activation of intracellular signaling pathways in Müller cells and other cells of the inner retina, but not photoreceptors. Investig. Ophthalmol. Vis. Sci. 2000, 41, 927–936. [Google Scholar]
- Wen, R.; Song, Y.; Cheng, T.; Matthes, M.T.; Yasumura, D.; LaVail, M.M.; Steinberg, R.H. Injury-induced upregulation of bFGF and CNTF mRNAs in the rat retina. J. Neurosci. 1995, 15, 7377–7385. [Google Scholar] [CrossRef]
- Nir, I.; Liu, C.; Wen, R. Light treatment enhances photoreceptor survival in dystrophic retinas of Royal College of Surgeons rats. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2383–2390. [Google Scholar]
- Schlichtenbrede, F.; MacNeil, A.; Bainbridge, J.; Tschernutter, M.; Thrasher, A.; Smith, A.; Ali, R. Intraocular gene delivery of ciliary neurotrophic factor results in significant loss of retinal function in normal mice and in the Prph2 Rd2/Rd2 model of retinal degeneration. Gene Ther. 2003, 10, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Bok, D.; Yasumura, D.; Matthes, M.T.; Ruiz, A.; Duncan, J.L.; Chappelow, A.V.; Zolutukhin, S.; Hauswirth, W.; LaVail, M.M. Effects of adeno-associated virus-vectored ciliary neurotrophic factor on retinal structure and function in mice with a P216L Rds/peripherin mutation. Exp. Eye Res. 2002, 74, 719–735. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Lin, P.; Liu, J.; Khor, C.; Lee, Y. Intraocular gene transfer of ciliary neurotrophic factor rescues photoreceptor degeneration in RCS rats. J. Biomed. Sci. 2004, 11, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Bush, R.A.; Lei, B.; Tao, W.; Raz, D.; Chan, C.; Cox, T.A.; Santos-Muffley, M.; Sieving, P.A. Encapsulated cell-based intraocular delivery of ciliary neurotrophic factor in normal rabbit: Dose-dependent effects on ERG and retinal histology. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2420–2430. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Tao, W.; Luo, L.; Huang, D.; Kauper, K.; Stabila, P.; LaVail, M.M.; Laties, A.M.; Wen, R. CNTF induces regeneration of cone outer segments in a rat model of retinal degeneration. PLoS ONE 2010, 5, e9495. [Google Scholar] [CrossRef] [Green Version]
- Cayouette, M.; Behn, D.; Sendtner, M.; Lachapelle, P.; Gravel, C. Intraocular gene transfer of ciliary neurotrophic factor prevents death and increases responsiveness of rod photoreceptors in the retinal degeneration slow mouse. J. Neurosci. 1998, 18, 9282–9293. [Google Scholar] [CrossRef] [Green Version]
- LaVail, M.M.; Yasumura, D.; Matthes, M.T.; Lau-Villacorta, C.; Unoki, K.; Sung, C.; Steinberg, R.H. Protection of mouse photoreceptors by survival factors in retinal degenerations. Investig. Ophthalmol. Vis. Sci. 1998, 39, 592–602. [Google Scholar]
- Beltran, W.A.; Wen, R.; Acland, G.M.; Aguirre, G.D. Intravitreal injection of ciliary neurotrophic factor (CNTF) causes peripheral remodeling and does not prevent photoreceptor loss in canine RPGR mutant Retina. Exp. Eye Res. 2007, 84, 753–771. [Google Scholar] [CrossRef] [Green Version]
- LaVail, M.M.; Unoki, K.; Yasumura, D.; Matthes, M.T.; Yancopoulos, G.D.; Steinberg, R.H. Multiple growth factors, cytokines, and neurotrophins rescue photoreceptors from the damaging effects of constant light. Proc. Natl. Acad. Sci. USA 1992, 89, 11249–11253. [Google Scholar] [CrossRef] [Green Version]
- Wen, R.; Tao, W.; Luo, L.; Huang, D.; Kauper, K.; Stabila, P.; LaVail, M.M.; Laties, A.M.; Li, Y. Regeneration of cone outer segments induced by CNTF. Adv. Exp. Med. Biol. 2012, 723, 93–99. [Google Scholar]
- Fandl, J.P.; Stahl, N.E.; Wiegand, S.J. Modified Ciliary Neurotrophic Factor (CNTF). U.S. Patent 7119066B2, 10 October 2006. [Google Scholar]
- Itkonen, J.M.; Urtti, A.; Bird, L.E.; Sarkhel, S. Codon optimization and factorial screening for enhanced soluble expression of human ciliary neurotrophic factor in Escherichia coli. BMC Biotechnol. 2014, 14, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berrow, N.S.; Alderton, D.; Owens, R.J. The precise engineering of expression vectors using high-throughput in-Fusion PCR cloning. Methods Mol. Biol. 2009, 498, 75–90. [Google Scholar]
- Studier, F.W. Protein production by auto-induction in high-density shaking cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef] [PubMed]
- Panula-Perälä, J.; Šiurkus, J.; Vasala, A.; Wilmanowski, R.; Casteleijn, M.G.; Neubauer, P. Enzyme controlled glucose auto-delivery for high cell density cultivations in microplates and shake flasks. Microb. Cell Fact. 2008, 7, 31. [Google Scholar] [CrossRef] [Green Version]
- Porstmann, T.; Ternynck, T.; Avrameas, S. Quantitation of 5-bromo-2-deoxyuridine incorporation into DNA: An enzyme immunoassay for the assessment of the lymphoid cell proliferative response. J. Immunol. Methods 1985, 82, 169–179. [Google Scholar] [CrossRef]
- Micsonai, A.; Wien, F.; Kernya, L.; Lee, Y.H.; Goto, Y.; Refregiers, M.; Kardos, J. Accurate secondary structure prediction and fold recognition for circular dichroism spectroscopy. Proc. Natl. Acad. Sci. USA 2015, 112, E3095–E3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sviben, D.; Bertoša, B.; Hloušek-Kasun, A.; Forcic, D.; Halassy, B.; Brgles, M. Investigation of the thermal shift assay and its power to predict protein and virus stabilizing conditions. J. Pharm. Biomed. Anal. 2018, 161, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Kopec, J.; Schneider, G. Comparison of fluorescence and light scattering based methods to assess formation and stability of protein–protein complexes. J. Struct. Biol. 2011, 175, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Boivin, S.; Kozak, S.; Meijers, R. Optimization of protein purification and characterization using Thermofluor screens. Protein Expr. Purif. 2013, 91, 192–206. [Google Scholar] [CrossRef]
- Caffe, A.; Ahuja, P.; Holmqvist, B.; Azadi, S.; Forsell, J.; Holmqvist, I.; Söderpalm, A.; Van Veen, T. Mouse retina explants after long-term culture in serum free medium. J. Chem. Neuroanat. 2002, 22, 263–273. [Google Scholar] [CrossRef]
- Arango-Gonzalez, B.; Szabó, A.; Pinzon-Duarte, G.; Lukáts, Á.; Guenther, E.; Kohler, K. In vivo and in vitro development of S-and M-cones in rat retina. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5320–5327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peynshaert, K.; Devoldere, J.; Forster, V.; Picaud, S.; Vanhove, C.; De Smedt, S.C.; Remaut, K. Toward smart design of retinal drug carriers: A novel bovine retinal explant model to study the barrier role of the vitreoretinal interface. Drug Deliv. 2017, 24, 1384–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, C.; Liu, L.; Zhang, Y.; Wang, Q.; Wang, J.; Liu, Y.; Su, Z. Purification and characterization of a long-acting ciliary neurotrophic factor via genetically fused with an albumin-binding domain. Protein Expr. Purif. 2017, 139, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, Y.; Zhang, C.; Guo, F.; Feng, C.; Li, X.; Shi, H.; Su, Z. High hydrostatic pressure enables almost 100% refolding of recombinant human ciliary neurotrophic factor from inclusion bodies at high concentration. Protein Expr. Purif. 2017, 133, 152–159. [Google Scholar] [CrossRef]
- Negro, A.; Grassato, L.; Polverino De Laureto, P.; Skaper, S.D. Genetic construction, properties and application of a green fluorescent protein-tagged ciliary neurotrophic factor. Protein Eng. 1997, 10, 1077–1083. [Google Scholar] [CrossRef] [Green Version]
- Delplace, V.; Ortin-Martinez, A.; Tsai, E.L.S.; Amin, A.N.; Wallace, V.; Shoichet, M.S. Controlled release strategy designed for intravitreal protein delivery to the retina. J. Control. Release 2019, 293, 10–20. [Google Scholar] [CrossRef]
- Stetefeld, J.; McKenna, S.A.; Patel, T.R. Dynamic light scattering: A practical guide and applications in biomedical sciences. Biophys. Rev. 2016, 8, 409–427. [Google Scholar] [CrossRef]
- Kitamura, T.; Tange, T.; Terasawa, T.; Chiba, S.; Kuwaki, T.; Miyagawa, K.; Piao, Y.; Miyazono, K.; Urabe, A.; Takaku, F. Establishment and characterization of a unique human cell line that proliferates dependently on GM-CSF, IL-3, or erythropoietin. J. Cell. Physiol. 1989, 140, 323–334. [Google Scholar] [CrossRef]
- Auguste, P.; Robledo, O.; Olivier, C.; Froger, J.; Praloran, V.; Pouplard-Barthelaix, A.; Gascan, H. Alanine substitution for Thr268 and Asp269 of soluble ciliary neurotrophic factor (CNTF) receptor alpha component defines a specific antagonist for the CNTF response. J. Biol. Chem. 1996, 271, 26049–26056. [Google Scholar] [CrossRef] [Green Version]
- Bhatnagar, B.S.; Bogner, R.H.; Pikal, M.J. Protein stability during freezing: Separation of stresses and mechanisms of protein stabilization. Pharm. Dev. Technol. 2007, 12, 505–523. [Google Scholar] [CrossRef] [PubMed]
- Zbacnik, T.J.; Holcomb, R.E.; Katayama, D.S.; Murphy, B.M.; Payne, R.W.; Coccaro, R.C.; Evans, G.J.; Matsuura, J.E.; Henry, C.S.; Manning, M.C. Role of buffers in protein formulations. J. Pharm. Sci. 2017, 106, 713–733. [Google Scholar] [CrossRef] [PubMed]
- Szkudlarek, B.A. Selective Crystallization of Phosphate Buffer Components and pH Changes during Freezing: Implications to Protein Stability. Ph.D. Thesis, University of Michigan, Ann Arbor, MI, USA, 1997. [Google Scholar]
- Seabrook, S.A.; Newman, J. High-throughput thermal scanning for protein stability: Making a good technique more robust. ACS Comb. Sci. 2013, 15, 387–392. [Google Scholar] [CrossRef]
- Jiskoot, W.; Randolph, T.W.; Volkin, D.B.; Middaugh, C.R.; Schöneich, C.; Winter, G.; Friess, W.; Crommelin, D.J.; Carpenter, J.F. Protein instability and immunogenicity: Roadblocks to clinical application of injectable protein delivery systems for sustained release. J. Pharm. Sci. 2012, 101, 946–954. [Google Scholar] [CrossRef]
- Wakshull, E.; Quarmby, V.; Mahler, H.; Rivers, H.; Jere, D.; Ramos, M.; Szczesny, P.; Bechtold-Peters, K.; Masli, S.; Gupta, S. Advancements in understanding immunogenicity of biotherapeutics in the intraocular space. AAPS J. 2017, 19, 1656–1668. [Google Scholar] [CrossRef] [PubMed]
- Lorber, B.; Fischer, F.; Bailly, M.; Roy, H.; Kern, D. Protein analysis by dynamic light scattering: Methods and techniques for students. Biochem. Mol. Biol. Educ. 2012, 40, 372–382. [Google Scholar] [CrossRef]
- Parmar, A.S.; Muschol, M. Hydration and hydrodynamic interactions of lysozyme: Effects of chaotropic versus kosmotropic ions. Biophys. J. 2009, 97, 590–598. [Google Scholar] [CrossRef] [Green Version]
- Amin, S.; Barnett, G.V.; Pathak, J.A.; Roberts, C.J.; Sarangapani, P.S. Protein aggregation, particle formation, characterization & rheology. Curr. Opin. Colloid Interface Sci. 2014, 19, 438–449. [Google Scholar]
- Filipe, V.; Hawe, A.; Carpenter, J.F.; Jiskoot, W. Analytical approaches to assess the degradation of therapeutic proteins. Trends Anal. Chem. 2013, 49, 118–125. [Google Scholar] [CrossRef]
- Patel, S.; Müller, G.; Stracke, J.O.; Altenburger, U.; Mahler, H.; Jere, D. Evaluation of protein drug stability with vitreous humor in a novel ex-vivo intraocular model. Eur. J. Pharm. Biopharm. 2015, 95, 407–417. [Google Scholar] [CrossRef]
- Patel, S.; Stracke, J.O.; Altenburger, U.; Mahler, H.; Metzger, P.; Shende, P.; Jere, D. Prediction of intraocular antibody drug stability using ex-vivo ocular model. Eur. J. Pharm. Biopharm. 2017, 112, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Rösch, S.; Aretzweiler, C.; Müller, F.; Walter, P. Evaluation of retinal function and morphology of the pink-eyed Royal College of Surgeons (RCS) rat: A comparative study of in vivo and in vitro methods. Curr. Eye Res. 2017, 42, 273–281. [Google Scholar] [CrossRef] [PubMed]
- McGill, T.J.; Prusky, G.T.; Douglas, R.M.; Yasumura, D.; Matthes, M.T.; Nune, G.; Donohue-Rolfe, K.; Yang, H.; Niculescu, D.; Hauswirth, W.W. Intraocular CNTF reduces vision in normal rats in a dose-dependent manner. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5756–5766. [Google Scholar] [CrossRef] [PubMed]
- Do Rhee, K.; Ruiz, A.; Duncan, J.L.; Hauswirth, W.W.; LaVail, M.M.; Bok, D.; Yang, X. Molecular and cellular alterations induced by sustained expression of ciliary neurotrophic factor in a mouse model of retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2007, 48, 1389–1400. [Google Scholar] [CrossRef] [PubMed]
- Chaitin, M.; Hall, M.O. Defective ingestion of rod outer segments by cultured dystrophic rat pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 1983, 24, 812–820. [Google Scholar] [PubMed]
- D’Cruz, P.M.; Yasumura, D.; Weir, J.; Matthes, M.T.; Abderrahim, H.; LaVail, M.M.; Vollrath, D. Mutation of the receptor tyrosine kinase gene Mertk in the retinal dystrophic RCS rat. Hum. Mol. Genet. 2000, 9, 645–651. [Google Scholar] [CrossRef] [Green Version]
- Dittrich, F.; Theonen, H.; Sendtner, M. Ciliary neurotrophic factor: Pharmacokinetics and acute-phase response in rat. Ann. Neurol. 1994, 35, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Sendtner, M.; Dittrich, F.; Hughes, R.A.; Thoenen, H. Actions of CNTF and neurotrophins on degenerating motoneurons: Preclinical studies and clinical implications. J. Neurol. Sci. 1994, 124, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Pasquin, S.; Sharma, M.; Gauchat, J. Ciliary neurotrophic factor (CNTF): New facets of an old molecule for treating neurodegenerative and metabolic syndrome pathologies. Cytokine Growth Factor Rev. 2015, 26, 507–515. [Google Scholar] [CrossRef]
- del Amo, E.M.; Vellonen, K.; Kidron, H.; Urtti, A. Intravitreal clearance and volume of distribution of compounds in rabbits: In silico prediction and pharmacokinetic simulations for drug development. Eur. J. Pharm. Biopharm. 2015, 95, 215–226. [Google Scholar] [CrossRef]
- del Amo, E.M.; Rimpelä, A.; Heikkinen, E.; Kari, O.K.; Ramsay, E.; Lajunen, T.; Schmitt, M.; Pelkonen, L.; Bhattacharya, M.; Richardson, D. Pharmacokinetic aspects of retinal drug delivery. Prog. Retin. Eye Res. 2017, 57, 134–185. [Google Scholar] [CrossRef] [PubMed]
- Shatz, W.; Hass, P.E.; Mathieu, M.; Kim, H.S.; Leach, K.; Zhou, M.; Crawford, Y.; Shen, A.; Wang, K.; Chang, D.P. Contribution of antibody hydrodynamic size to vitreal clearance revealed through rabbit studies using a species-matched Fab. Mol. Pharm. 2016, 13, 2996–3003. [Google Scholar] [CrossRef] [PubMed]
- Crowell, S.R.; Wang, K.; Famili, A.; Shatz, W.; Loyet, K.M.; Chang, V.; Liu, Y.; Prabhu, S.; Kamath, A.V.; Kelley, R.F. Influence of charge, hydrophobicity, and size on vitreous pharmacokinetics of large molecules. Transl. Visc. Sci. Technol. 2019, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekdawi, N.S.; Pulido, J.S.; Itty, S.; Marler, R.J.; Herman, D.C.; Hardwig, P.; Mohney, B.G.; Valyi-Nagy, T.; Shukla, D. Intravitreal alemtuzumab penetrates full-thickness retina in rabbit eyes. Retina 2009, 29, 1532–1534. [Google Scholar] [CrossRef] [PubMed]
- Melo, G.B.; Moraes Filho, M.N.; Rodrigues, E.B.; Regatieri, C.V.; Dreyfuss, J.L.; Penha, F.M.; Pinheiro, M.M.; Coimbra, R.C.; Haapalainen, E.F.; Farah, M.E. Toxicity and retinal penetration of infliximab in primates. Retina 2012, 32, 606–612. [Google Scholar] [CrossRef]
- Heiduschka, P.; Fietz, H.; Hofmeister, S.; Schultheiss, S.; Mack, A.F.; Peters, S.; Ziemssen, F.; Niggemann, B.; Julien, S.; Bartz-Schmidt, K.U. Penetration of bevacizumab through the retina after intravitreal injection in the monkey. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2814–2823. [Google Scholar] [CrossRef] [Green Version]
- Jankowiak, W.; Kruszewski, K.; Flachsbarth, K.; Skevas, C.; Richard, G.; Rüther, K.; Braulke, T.; Bartsch, U. Sustained neural stem cell-based intraocular delivery of CNTF attenuates photoreceptor loss in the Nclf mouse model of neuronal ceroid lipofuscinosis. PLoS ONE 2015, 10, e0127204. [Google Scholar] [CrossRef] [Green Version]
- Leaver, S.G.; Cui, Q.; Plant, G.W.; Arulpragasam, A.; Hisheh, S.; Verhaagen, J.; Harvey, A.R. AAV-mediated expression of CNTF promotes long-term survival and regeneration of adult rat retinal ganglion cells. Gene Ther. 2006, 13, 1328. [Google Scholar] [CrossRef] [Green Version]
- Pease, M.E.; Zack, D.J.; Berlinicke, C.; Bloom, K.; Cone, F.; Wang, Y.; Klein, R.L.; Hauswirth, W.W.; Quigley, H.A. Effect of CNTF on retinal ganglion cell survival in experimental glaucoma. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- LeVaillant, C.J.; Sharma, A.; Muhling, J.; Wheeler, L.P.; Cozens, G.S.; Hellström, M.; Rodger, J.; Harvey, A.R. Significant changes in endogenous retinal gene expression assessed 1 year after a single intraocular injection of AAV-CNTF or AAV-BDNF. Mol. Ther. Methods Clin. Dev. 2016, 3, 16078. [Google Scholar] [CrossRef]
- Lipinski, D.M.; Barnard, A.R.; Singh, M.S.; Martin, C.; Lee, E.J.; Davies, W.I.; MacLaren, R.E. CNTF gene therapy confers lifelong neuroprotection in a mouse model of human retinitis pigmentosa. Mol. Ther. 2015, 23, 1308–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, F.; Dejneka, N.S.; Cohen, D.R.; Krasnoperova, N.V.; Lem, J.; Maguire, A.M.; Dudus, L.; Fisher, K.J.; Bennett, J. AAV-mediated delivery of ciliary neurotrophic factor prolongs photoreceptor survival in the rhodopsin knockout mouse. Mol. Ther. 2001, 3, 241–248. [Google Scholar] [CrossRef]
- Kauper, K.; McGovern, C.; Sherman, S.; Heatherton, P.; Rapoza, R.; Stabila, P.; Dean, B.; Lee, A.; Borges, S.; Bouchard, B. Two-year intraocular delivery of ciliary neurotrophic factor by encapsulated cell technology implants in patients with chronic retinal degenerative diseases. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7484–7491. [Google Scholar] [CrossRef]
- Zhang, K.; Hopkins, J.J.; Heier, J.S.; Birch, D.G.; Halperin, L.S.; Albini, T.A.; Brown, D.M.; Jaffe, G.J.; Tao, W.; Williams, G.A. Ciliary neurotrophic factor delivered by encapsulated cell intraocular implants for treatment of geographic atrophy in age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2011, 108, 6241–6245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zein, W.M.; Jeffrey, B.G.; Wiley, H.E.; Turriff, A.E.; Tumminia, S.J.; Tao, W.; Bush, R.A.; Marangoni, D.; Wen, R.; Wei, L.L. CNGB3-achromatopsia clinical trial with CNTF: Diminished rod pathway responses with no evidence of improvement in cone function. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6301–6308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemi, M.; Alizadeh, E.; Saei Arezoumand, K.; Fallahi Motlagh, B.; Zarghami, N. Ciliary neurotrophic factor (CNTF) delivery to retina: An overview of current research advancements. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1694–1707. [Google Scholar] [CrossRef]
- Talcott, K.E.; Ratnam, K.; Sundquist, S.M.; Lucero, A.S.; Lujan, B.J.; Tao, W.; Porco, T.C.; Roorda, A.; Duncan, J.L. Longitudinal study of cone photoreceptors during retinal degeneration and in response to ciliary neurotrophic factor treatment. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2219–2226. [Google Scholar] [CrossRef]
- Chew, E.Y.; Clemons, T.E.; Jaffe, G.J.; Johnson, C.A.; Farsiu, S.; Lad, E.M.; Guymer, R.; Rosenfeld, P.; Hubschman, J.; Constable, I.; et al. Effect of ciliary neurotrophic factor on retinal neurodegeneration in patients with macular telangiectasia type 2: A randomized clinical trial. Ophthalmology 2019, 126, 540–549. [Google Scholar] [CrossRef]
- El Sanharawi, M.; Kowalczuk, L.; Touchard, E.; Omri, S.; De Kozak, Y.; Behar-Cohen, F. Protein delivery for retinal diseases: From basic considerations to clinical applications. Prog. Retin. Eye Res. 2010, 29, 443–465. [Google Scholar] [CrossRef]
- Lau, C.M.L.; Yu, Y.; Jahanmir, G.; Chau, Y. Controlled release technology for anti-angiogenesis treatment of posterior eye diseases: Current status and challenges. Adv. Drug Deliv. Rev. 2018, 126, 145–161. [Google Scholar] [CrossRef]
- Peynshaert, K.; Devoldere, J.; Minnaert, A.; De Smedt, S.C.; Remaut, K. Morphology and composition of the inner limiting membrane: Species-specific variations and relevance toward drug delivery research. Curr. Eye Res. 2019, 44, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Valdés, J.; Trachsel-Moncho, L.; Sahaboglu, A.; Trifunović, D.; Miranda, M.; Ueffing, M.; Paquet-Durand, F.; Schmachtenberg, O. Organotypic retinal explant cultures as in vitro alternative for diabetic retinopathy studies. ALTEX 2016, 33, 459–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peynshaert, K.; Devoldere, J.; De Smedt, S.C.; Remaut, K. In vitro and ex vivo models to study drug delivery barriers in the posterior segment of the eye. Adv. Drug Deliv. Rev. 2018, 126, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Alarautalahti, V.; Ragauskas, S.; Hakkarainen, J.J.; Uusitalo-Järvinen, H.; Uusitalo, H.; Hyttinen, J.; Kalesnykas, G.; Nymark, S. Viability of mouse retinal explant cultures assessed by preservation of functionality and morphology. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1914–1927. [Google Scholar] [CrossRef] [Green Version]
- Shahar, J.; Avery, R.L.; Heilweil, G.; Barak, A.; Zemel, E.; Lewis, G.P.; Johnson, P.T.; Fisher, S.K.; Perlman, I.; Loewenstein, A. Electrophysiologic and retinal penetration studies following intravitreal injection of bevacizumab (Avastin). Retina 2006, 26, 262–269. [Google Scholar] [CrossRef]
- Hutton-Smith, L.A.; Gaffney, E.A.; Byrne, H.M.; Maini, P.K.; Gadkar, K.; Mazer, N.A. Ocular pharmacokinetics of therapeutic antibodies given by intravitreal injection: Estimation of retinal permeabilities using a 3-Compartment semi-mechanistic model. Mol. Pharm. 2017, 14, 2690–2696. [Google Scholar] [CrossRef]
- Hutton-Smith, L.A.; Gaffney, E.A.; Byrne, H.M.; Caruso, A.; Maini, P.K.; Mazer, N.A. Theoretical insights into the retinal dynamics of vascular endothelial growth factor in patients treated with ranibizumab, based on an ocular pharmacokinetic/pharmacodynamic model. Mol. Pharm. 2018, 15, 2770–2784. [Google Scholar] [CrossRef]
















© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Itkonen, J.; Annala, A.; Tavakoli, S.; Arango-Gonzalez, B.; Ueffing, M.; Toropainen, E.; Ruponen, M.; Casteleijn, M.G.; Urtti, A. Characterization, Stability, and In Vivo Efficacy Studies of Recombinant Human CNTF and Its Permeation into the Neural Retina in Ex Vivo Organotypic Retinal Explant Culture Models. Pharmaceutics 2020, 12, 611. https://doi.org/10.3390/pharmaceutics12070611
Itkonen J, Annala A, Tavakoli S, Arango-Gonzalez B, Ueffing M, Toropainen E, Ruponen M, Casteleijn MG, Urtti A. Characterization, Stability, and In Vivo Efficacy Studies of Recombinant Human CNTF and Its Permeation into the Neural Retina in Ex Vivo Organotypic Retinal Explant Culture Models. Pharmaceutics. 2020; 12(7):611. https://doi.org/10.3390/pharmaceutics12070611
Chicago/Turabian StyleItkonen, Jaakko, Ada Annala, Shirin Tavakoli, Blanca Arango-Gonzalez, Marius Ueffing, Elisa Toropainen, Marika Ruponen, Marco G. Casteleijn, and Arto Urtti. 2020. "Characterization, Stability, and In Vivo Efficacy Studies of Recombinant Human CNTF and Its Permeation into the Neural Retina in Ex Vivo Organotypic Retinal Explant Culture Models" Pharmaceutics 12, no. 7: 611. https://doi.org/10.3390/pharmaceutics12070611