Nano-Polyplexes Mediated Transfection of Runx2-shRNA Mitigates the Osteodifferentiation of Human Valvular Interstitial Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Valvular Interstitial Cells (VIC): Isolation and Culture
2.3. Preparation of Fullerene (C60)-PEI/Short Hairpin (sh)RNA Plasmid Nano-Polyplexes
2.4. Characterization of Nano-Polyplexes
2.4.1. Size and Zeta Potential
2.4.2. Agarose Gel Retardation Assay
2.4.3. Evaluation of Nano-Polyplexes Cytotoxicity
2.4.4. Uptake of Nano-Polyplexes by VIC
2.4.5. Transfection Assay
2.5. Assessment of the Expression of Osteogenic Proteins in VIC Exposed to Medium Containing High Glucose Concentrations in the Absence or Presence of Osteogenic Factors
2.5.1. Western Blot Assays
2.6. Treatment of VIC with Nano-Polyplexes Carrying shRNA Sequences Specific for Runx2
2.6.1. Transfection of VIC with C60-PEI/shRNA-Runx2 Nano-Polyplexes
2.6.2. Quantitative Real-Time Polymerase Chain Reaction
2.6.3. Western Blot Assay
2.6.4. Determination of Alkaline Phosphatase (ALP) Activity in VIC after Transfection with C60-PEI/shRNA-Runx2 Nano-Polyplexes
Quantification of Alkaline Phosphatase Activity
Alkaline Phosphatase Staining
2.7. Statistical Analysis
3. Results
3.1. Characterization of Nano-Polyplexes
3.1.1. Size and Zeta Potential
3.1.2. Nano-Polyplexes Effectively Packs shRNA Plasmid
3.1.3. Cytotoxicity of C60-PEI/shRNA Plasmid Nano-Polyplexes
3.1.4. C60-PEI/Cy3-Labelled Plasmid Nano-Polyplexes Are Efficiently Taken up by VIC
3.1.5. The Transfection Efficiency of Nano-Polyplexes in VIC
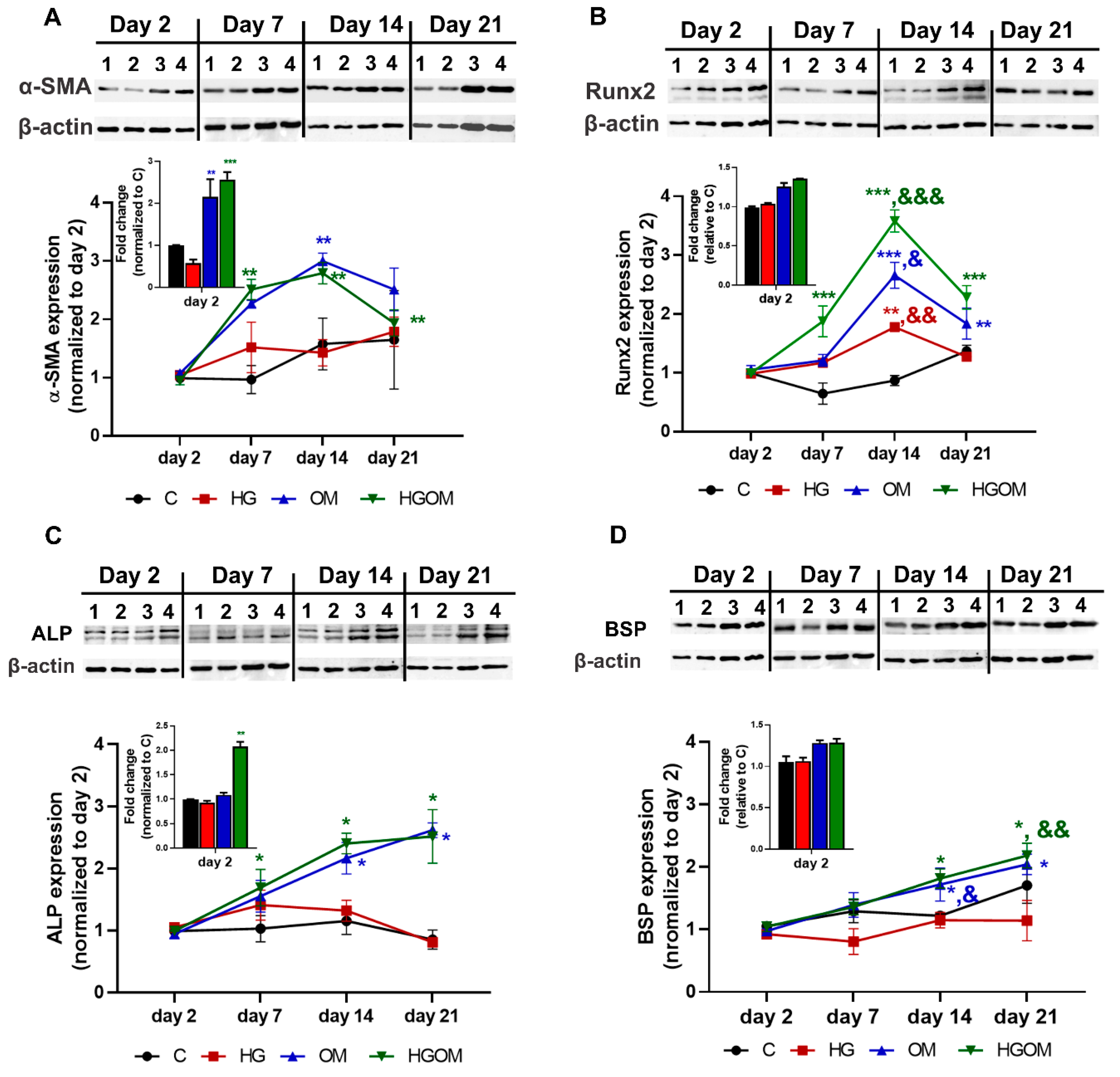
3.2. Time Course Expression of Osteogenesis-Related Markers in VIC Exposed to High Glucose Concentrations in the Absence or Presence of Osteogenic Factors
3.3. In VIC, C60-PEI/shRNA-Runx2 Nano-Polyplexes Down-Regulate Runx2 Gene and Protein Expression
3.4. In VIC Exposed to HGOM, Down-Regulation of Runx2 Expression Diminishes the Expression of Osteoblast Differentiation Markers
3.5. Reduction of Runx2 Expression Mitigates the Alkaline Phosphatase Activity in VIC Exposed to HGOM
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lindman, B.R.; Clavel, M.A.; Mathieu, P.; Iung, B.; Lancellotti, P.; Otto, C.M.; Pibarot, P. Calcific aortic stenosis. Nat. Rev. Dis. Primers 2016, 2, 16006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yutzey, K.E.; Demer, L.L.; Body, S.C.; Huggins, G.S.; Towler, D.A.; Giachelli, C.M.; Hofmann-Bowman, M.A.; Mortlock, D.P.; Rogers, M.B.; Sadeghi, M.M.; et al. Calcific aortic valve disease: A consensus summary from the Alliance of Investigators on Calcific Aortic Valve Disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2387–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawade, T.A.; Newby, D.E.; Dweck, M.R. Calcification in Aortic Stenosis: The Skeleton Key. J. Am. Coll. Cardiol. 2015, 66, 561–577. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.I.; Sakuma, I.; Sohn, I.S.; Jo, S.H.; Koh, K.K. Inflammatory and metabolic mechanisms underlying the calcific aortic valve disease. Atherosclerosis 2018, 277, 60–65. [Google Scholar] [CrossRef]
- Virtanen, M.P.O.; Airaksinen, J.; Niemela, M.; Laakso, T.; Husso, A.; Jalava, M.P.; Tauriainen, T.; Maaranen, P.; Kinnunen, E.M.; Dahlbacka, S.; et al. Comparison of Survival of Transfemoral Transcatheter Aortic Valve Implantation Versus Surgical Aortic Valve Replacement for Aortic Stenosis in Low-Risk Patients Without Coronary Artery Disease. Am. J. Cardiol. 2020, 125, 589–596. [Google Scholar] [CrossRef]
- Akin, I.; Nienaber, C.A. Is there evidence for statins in the treatment of aortic valve stenosis? World J. Cardiol. 2017, 9, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Rutkovskiy, A.; Malashicheva, A.; Sullivan, G.; Bogdanova, M.; Kostareva, A.; Stenslokken, K.O.; Fiane, A.; Vaage, J. Valve Interstitial Cells: The Key to Understanding the Pathophysiology of Heart Valve Calcification. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef]
- Filip, D.A.; Radu, A.; Simionescu, M. Interstitial cells of the heart valves possess characteristics similar to smooth muscle cells. Circ. Res. 1986, 59, 310–320. [Google Scholar] [CrossRef] [Green Version]
- Filip, D.A.; Nistor, A.; Bulla, A.; Radu, A.; Lupu, F.; Simionescu, M. Cellular events in the development of valvular atherosclerotic lesions induced by experimental hypercholesterolemia. Atherosclerosis 1987, 67, 199–214. [Google Scholar] [CrossRef]
- Osman, L.; Chester, A.H.; Sarathchandra, P.; Latif, N.; Meng, W.; Taylor, P.M.; Yacoub, M.H. A novel role of the sympatho-adrenergic system in regulating valve calcification. Circulation 2007, 116, I282–I287. [Google Scholar] [CrossRef] [Green Version]
- Rajamannan, N.M.; Subramaniam, M.; Rickard, D.; Stock, S.R.; Donovan, J.; Springett, M.; Orszulak, T.; Fullerton, D.A.; Tajik, A.J.; Bonow, R.O.; et al. Human aortic valve calcification is associated with an osteoblast phenotype. Circulation 2003, 107, 2181–2184. [Google Scholar] [CrossRef] [PubMed]
- Osman, L.; Yacoub, M.H.; Latif, N.; Amrani, M.; Chester, A.H. Role of human valve interstitial cells in valve calcification and their response to atorvastatin. Circulation 2006, 114, I547–I552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducy, P. Cbfa1: A molecular switch in osteoblast biology. Dev. Dyn. 2000, 219, 461–471. [Google Scholar] [CrossRef]
- Bruderer, M.; Richards, R.G.; Alini, M.; Stoddart, M.J. Role and regulation of RUNX2 in osteogenesis. Eur. Cell. Mater. 2014, 28, 269–286. [Google Scholar] [CrossRef] [PubMed]
- Wirrig, E.E.; Hinton, R.B.; Yutzey, K.E. Differential expression of cartilage and bone-related proteins in pediatric and adult diseased aortic valves. J. Mol. Cell. Cardiol. 2011, 50, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Cheek, J.D.; Wirrig, E.E.; Alfieri, C.M.; James, J.F.; Yutzey, K.E. Differential activation of valvulogenic, chondrogenic, and osteogenic pathways in mouse models of myxomatous and calcific aortic valve disease. J. Mol. Cell. Cardiol. 2012, 52, 689–700. [Google Scholar] [CrossRef] [Green Version]
- Wirrig, E.E.; Yutzey, K.E. Developmental pathways in CAVD. In Calcific Aortic Valve Disease; IntechOpen Limited: London, UK, 2013. [Google Scholar]
- Nagy, E.; Eriksson, P.; Yousry, M.; Caidahl, K.; Ingelsson, E.; Hansson, G.K.; Franco-Cereceda, A.; Back, M. Valvular osteoclasts in calcification and aortic valve stenosis severity. Int. J. Cardiol. 2013, 168, 2264–2271. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Lin, Y.; Sun, Z. Deficiency in the anti-aging gene Klotho promotes aortic valve fibrosis through AMPK alpha-mediated activation of RUNX2. Aging Cell 2016, 15, 853–860. [Google Scholar] [CrossRef]
- Mishra, S.; Vaughn, A.D.; Devore, D.I.; Roth, C.M. Delivery of siRNA silencing Runx2 using a multifunctional polymer-lipid nanoparticle inhibits osteogenesis in a cell culture model of heterotopic ossification. Integr. Biol. 2012, 4, 1498–1507. [Google Scholar] [CrossRef] [Green Version]
- Le Quang, K.; Bouchareb, R.; Lachance, D.; Laplante, M.A.; El Husseini, D.; Boulanger, M.C.; Fournier, D.; Fang, X.P.; Avramoglu, R.K.; Pibarot, P.; et al. Early development of calcific aortic valve disease and left ventricular hypertrophy in a mouse model of combined dyslipidemia and type 2 diabetes mellitus. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2283–2291. [Google Scholar] [CrossRef]
- Banovic, M.; Athithan, L.; McCann, G.P. Aortic stenosis and diabetes mellitus: An ominous combination. Diab. Vasc. Dis. Res. 2019, 16, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Barth, M.; Selig, J.I.; Klose, S.; Schomakers, A.; Kiene, L.S.; Raschke, S.; Boeken, U.; Akhyari, P.; Fischer, J.W.; Lichtenberg, A. Degenerative aortic valve disease and diabetes: Implications for a link between proteoglycans and diabetic disorders in the aortic valve. Diab. Vasc. Dis. Res. 2019, 16, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Tucureanu, M.M.; Filippi, A.; Alexandru, N.; Ana Constantinescu, C.; Ciortan, L.; Macarie, R.; Vadana, M.; Voicu, G.; Frunza, S.; Nistor, D.; et al. Diabetes-induced early molecular and functional changes in aortic heart valves in a murine model of atherosclerosis. Diab. Vasc. Dis. Res. 2019, 16, 562–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, C.B.; Guthrie, E.H.; Huang, M.T.; Taxman, D.J. Short hairpin RNA (shRNA): Design, delivery, and assessment of gene knockdown. Methods Mol. Biol. 2010, 629, 141–158. [Google Scholar] [CrossRef] [Green Version]
- Vadana, M.; Cecoltan, S.; Ciortan, L.; Macarie, R.D.; Tucureanu, M.M.; Mihaila, A.C.; Droc, I.; Butoi, E.; Manduteanu, I. Molecular mechanisms involved in high glucose-induced valve calcification in a 3D valve model with human valvular cells. J. Cell. Mol. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- World Medical Association. World Medical Association Declaration of Helsinki: Recommendations Guiding Physicians in Biomedical Research Involving Human Subjects. JAMA 1997, 277, 925–926. [Google Scholar] [CrossRef]
- Uritu, C.M.; Varganici, C.D.; Ursu, L.; Coroaba, A.; Nicolescu, A.; Dascalu, A.I.; Peptanariu, D.; Stan, D.; Constantinescu, C.A.; Simion, V.; et al. Hybrid fullerene conjugates as vectors for DNA cell-delivery. J. Mater. Chem. B 2015, 3, 2433–2446. [Google Scholar] [CrossRef]
- Constantinescu, C.A.; Fuior, E.V.; Rebleanu, D.; Deleanu, M.; Simion, V.; Voicu, G.; Escriou, V.; Manduteanu, I.; Simionescu, M.; Calin, M. Targeted Transfection Using PEGylated Cationic Liposomes Directed Towards P-Selectin Increases siRNA Delivery into Activated Endothelial Cells. Pharmaceutics 2019, 11, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Titorencu, I.; Jinga, V.; Constantinescu, E.; Gafencu, A.; Ciohodaru, C.; Manolescu, I.; Zaharia, C.; Simionescu, M. Proliferation, differentiation and characterization of osteoblasts from human BM mesenchymal cells. Cytotherapy 2007, 9, 682–696. [Google Scholar] [CrossRef]
- Towler, D.A. Molecular and cellular aspects of calcific aortic valve disease. Circ. Res. 2013, 113, 198–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnett, J.C.; Rossi, J.J. RNA-based therapeutics: Current progress and future prospects. Chem. Biol. 2012, 19, 60–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnett, J.C.; Rossi, J.J.; Tiemann, K. Current progress of siRNA/shRNA therapeutics in clinical trials. Biotechnol. J. 2011, 6, 1130–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Tang, C.; Yin, C. Dual-targeting and pH/redox-responsive multi-layered nanocomplexes for smart co-delivery of doxorubicin and siRNA. Biomaterials 2015, 60, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Marquez, A.R.M.; Madu, C.O.; Lu, Y. An overview of various carriers for siRNA delivery. Oncomedicine 2018, 3, 48–58. [Google Scholar] [CrossRef]
- Yang, X.; Fullerton, D.A.; Su, X.; Ao, L.; Cleveland, J.C., Jr.; Meng, X. Pro-osteogenic phenotype of human aortic valve interstitial cells is associated with higher levels of Toll-like receptors 2 and 4 and enhanced expression of bone morphogenetic protein 2. J. Am. Coll. Cardiol. 2009, 53, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.W.; Zhang, B.Y.; Wang, S.W.; Gong, D.J.; Han, L.; Xu, Z.Y.; Liu, X.H. Twist-related protein 1 negatively regulated osteoblastic transdifferentiation of human aortic valve interstitial cells by directly inhibiting runt-related transcription factor 2. J. Thorac. Cardiovasc. Surg. 2014, 148. [Google Scholar] [CrossRef] [Green Version]
- Carrion, K.; Dyo, J.; Patel, V.; Sasik, R.; Mohamed, S.A.; Hardiman, G.; Nigam, V. The long non-coding HOTAIR is modulated by cyclic stretch and WNT/beta-CATENIN in human aortic valve cells and is a novel repressor of calcification genes. PLoS ONE 2014, 9, e96577. [Google Scholar] [CrossRef] [Green Version]
- Nigam, V.; Srivastava, D. Notch1 represses osteogenic pathways in aortic valve cells. J. Mol. Cell. Cardiol. 2009, 47, 828–834. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Zhao, Z.; Cai, Z.; Dong, N.; Liu, Y. Oxidized low-density lipoprotein promotes osteoblastic differentiation of valvular interstitial cells through RAGE/MAPK. Cardiology 2015, 130, 55–61. [Google Scholar] [CrossRef]
- New, S.E.; Aikawa, E. Molecular imaging insights into early inflammatory stages of arterial and aortic valve calcification. Circ. Res. 2011, 108, 1381–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Meng, X.; Su, X.; Mauchley, D.C.; Ao, L.; Cleveland, J.C., Jr.; Fullerton, D.A. Bone morphogenic protein 2 induces Runx2 and osteopontin expression in human aortic valve interstitial cells: Role of Smad1 and extracellular signal-regulated kinase 1/2. J. Thorac. Cardiovasc. Surg. 2009, 138, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natorska, J.; Wypasek, E.; Grudzien, G.; Sobczyk, D.; Marek, G.; Filip, G.; Sadowski, J.; Undas, A. Does diabetes accelerate the progression of aortic stenosis through enhanced inflammatory response within aortic valves? Inflammation 2012, 35, 834–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamalesh, M.; Ng, C.; El Masry, H.; Eckert, G.; Sawada, S. Does diabetes accelerate progression of calcific aortic stenosis? Eur. J. Echocardiogr. 2009, 10, 723–725. [Google Scholar] [CrossRef] [Green Version]
- Shemesh, J.; Tenenbaum, A.; Fisman, E.Z.; Koren-Morag, N.; Grossman, E. Coronary calcium in patients with and without diabetes: First manifestation of acute or chronic coronary events is characterized by different calcification patterns. Cardiovasc. Diabetol. 2013, 12, 161. [Google Scholar] [CrossRef] [Green Version]
- Hjortnaes, J.; Goettsch, C.; Hutcheson, J.D.; Camci-Unal, G.; Lax, L.; Scherer, K.; Body, S.; Schoen, F.J.; Kluin, J.; Khademhosseini, A.; et al. Simulation of early calcific aortic valve disease in a 3D platform: A role for myofibroblast differentiation. J. Mol. Cell. Cardiol. 2016, 94, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, R.T.; Xiao, G.; Jiang, D.; Gopalakrishnan, R.; Yang, S.; Reith, E. Multiple signaling pathways converge on the Cbfa1/Runx2 transcription factor to regulate osteoblast differentiation. Connect. Tissue Res. 2003, 44 Suppl 1, 109–116. [Google Scholar] [CrossRef]
- Boroumand, S.; Asadpour, S.; Akbarzadeh, A.; Faridi-Majidi, R.; Ghanbari, H. Heart valve tissue engineering: An overview of heart valve decellularization processes. Regen. Med. 2018, 13, 41–54. [Google Scholar] [CrossRef]
- Bogdanova, M.; Kostina, A.; Zihlavnikova Enayati, K.; Zabirnyk, A.; Malashicheva, A.; Stenslokken, K.O.; Sullivan, G.J.; Kaljusto, M.L.; Kvitting, J.P.; Kostareva, A.; et al. Inflammation and Mechanical Stress Stimulate Osteogenic Differentiation of Human Aortic Valve Interstitial Cells. Front. Physiol. 2018, 9, 1635. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voicu, G.; Rebleanu, D.; Constantinescu, C.A.; Fuior, E.V.; Ciortan, L.; Droc, I.; Uritu, C.M.; Pinteala, M.; Manduteanu, I.; Simionescu, M.; et al. Nano-Polyplexes Mediated Transfection of Runx2-shRNA Mitigates the Osteodifferentiation of Human Valvular Interstitial Cells. Pharmaceutics 2020, 12, 507. https://doi.org/10.3390/pharmaceutics12060507
Voicu G, Rebleanu D, Constantinescu CA, Fuior EV, Ciortan L, Droc I, Uritu CM, Pinteala M, Manduteanu I, Simionescu M, et al. Nano-Polyplexes Mediated Transfection of Runx2-shRNA Mitigates the Osteodifferentiation of Human Valvular Interstitial Cells. Pharmaceutics. 2020; 12(6):507. https://doi.org/10.3390/pharmaceutics12060507
Chicago/Turabian StyleVoicu, Geanina, Daniela Rebleanu, Cristina Ana Constantinescu, Elena Valeria Fuior, Letitia Ciortan, Ionel Droc, Cristina Mariana Uritu, Mariana Pinteala, Ileana Manduteanu, Maya Simionescu, and et al. 2020. "Nano-Polyplexes Mediated Transfection of Runx2-shRNA Mitigates the Osteodifferentiation of Human Valvular Interstitial Cells" Pharmaceutics 12, no. 6: 507. https://doi.org/10.3390/pharmaceutics12060507