Repurposing Old Drugs into New Epigenetic Inhibitors: Promising Candidates for Cancer Treatment?

 , , ,
, , ,  and
and 
Abstract
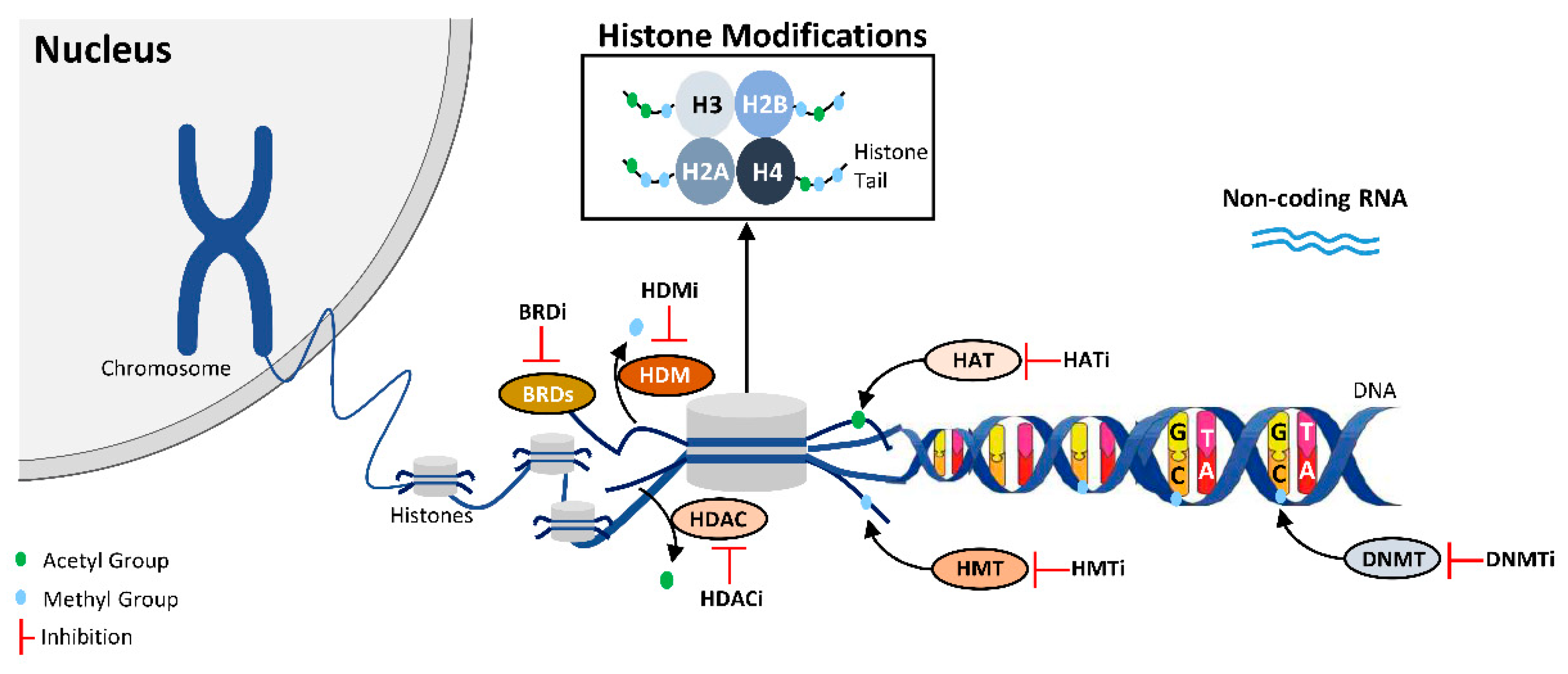
:1. Introduction
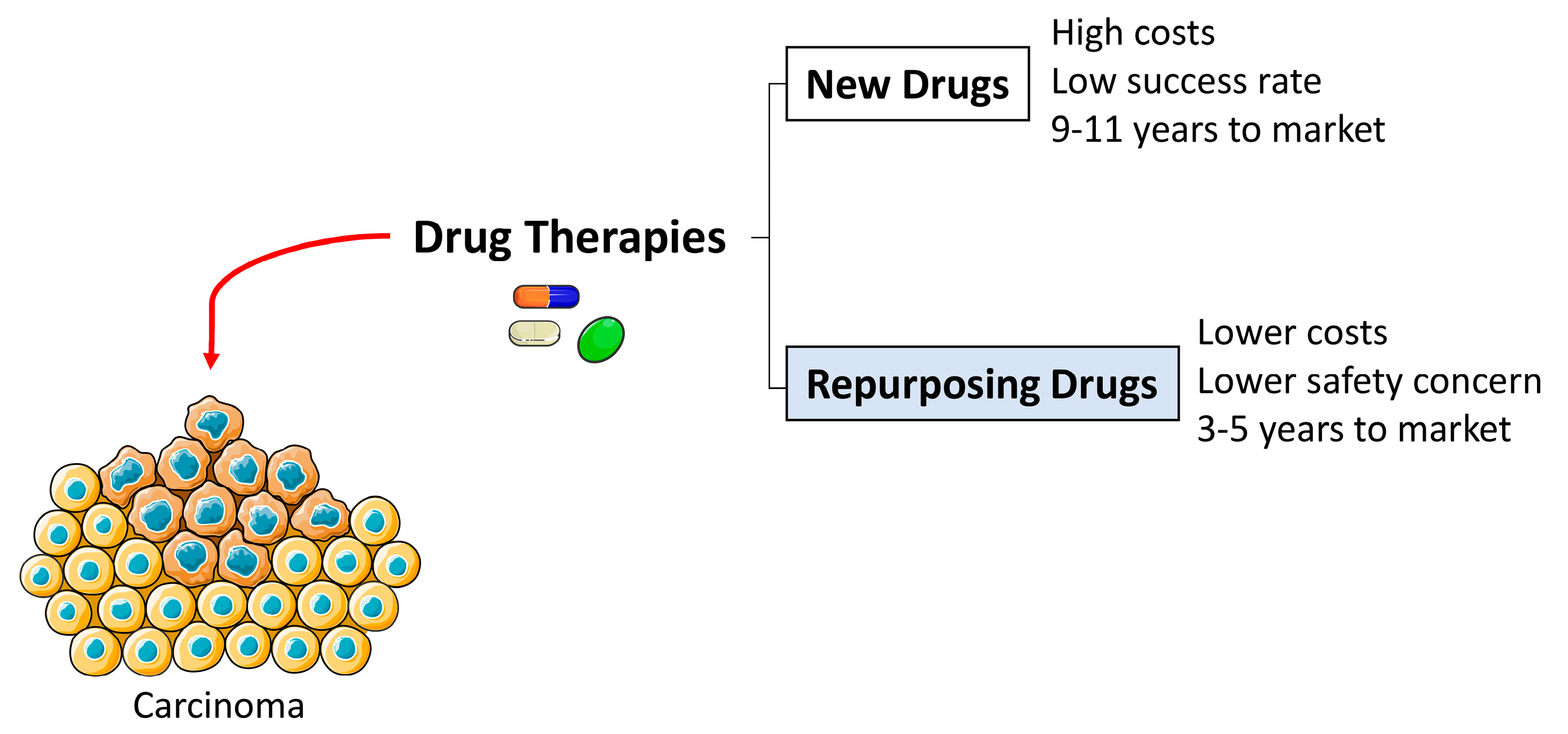
2. Drug Repurposing
3. DNMT Inhibitors
4. Inhibitors of Histone Modulators
4.1. HDAC Inhibitors
4.2. HAT, HMT, HDM and BET Inhibitors
5. DNMT and HDAC Dual Inhibitors
6. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Jerónimo, C.; Bastian, P.J.; Bjartell, A.; Carbone, G.M.; Catto, J.W.; Clark, S.J.; Henrique, R.; Nelson, W.G.; Shariat, S.F. Epigenetics in Prostate Cancer: Biologic and Clinical Relevance. Eur. Urol. 2011, 60, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Jurkowska, R.Z.; Jurkowski, T.; Jeltsch, A. Structure and Function of Mammalian DNA Methyltransferases. ChemBioChem 2010, 12, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Millar, D.S.; Ow, K.K.; Paul, C.L.; Russell, P.J.; Molloy, P.L.; Clark, S.J. Detailed methylation analysis of the glutathione S-transferase pi (GSTP1) gene in prostate cancer. Oncogene 1999, 18, 1313–1324. [Google Scholar] [CrossRef]
- Schayek, H.; Bentov, I.; Sun, S.; Plymate, S.R.; Werner, H. Progression to metastatic stage in a cellular model of prostate cancer is associated with methylation of the androgen receptor gene and transcriptional suppression of the insulin-like growth factor-I receptor gene. Exp. Cell Res. 2010, 316, 1479–1488. [Google Scholar] [CrossRef] [Green Version]
- Kouzarides, T. Chromatin Modifications and Their Function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Weichert, W.; Röske, A.; Gekeler, V.; Beckers, T.; Stephan, C.; Jung, K.; Fritzsche, F.R.; Niesporek, S.; Denkert, C.; Dietel, M.; et al. Histone deacetylases 1, 2 and 3 are highly expressed in prostate cancer and HDAC2 expression is associated with shorter PSA relapse time after radical prostatectomy. Br. J. Cancer 2008, 98, 604–610. [Google Scholar] [CrossRef] [Green Version]
- Novotny-Diermayr, V.; Sangthongpitag, K.; Hu, C.Y.; Wu, X.; Sausgruber, N.; Yeo, P.; Greicius, G.; Pettersson, S.; Liang, A.L.; Loh, Y.K.; et al. SB939, a Novel Potent and Orally Active Histone Deacetylase Inhibitor with High Tumor Exposure and Efficacy in Mouse Models of Colorectal Cancer. Mol. Cancer Ther. 2010, 9, 642–652. [Google Scholar] [CrossRef] [Green Version]
- Varambally, S.; Dhanasekaran, S.M.; Zhou, M.; Barrette, T.R.; Kumar-Sinha, C.; Sanda, M.G.; Ghosh, D.; Pienta, K.J.; Sewalt, R.G.A.B.; Otte, A.P.; et al. The polycomb group protein EZH2 is involved in progression of prostate cancer. Nature 2002, 419, 624–629. [Google Scholar] [CrossRef]
- Metzger, E.; Wissmann, M.; Yin, N.; Müller, J.M.; Schneider, R.; Peters, A.H.F.M.; Günther, T.; Buettner, R.; Schüle, R. LSD1 demethylates repressive histone marks to promote androgen-receptor-dependent transcription. Nature 2005, 437, 436–439. [Google Scholar] [CrossRef]
- Ashburn, T.T.; Thor, K.B. Drug repositioning: Identifying and developing new uses for existing drugs. Nat. Rev. Drug Discov. 2004, 3, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Peyvandipour, A.; Saberian, N.; Shafi, A.; Donato, M.; Draghici, S. A novel computational approach for drug repurposing using systems biology. Bioinformatics 2018, 34, 2817–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, J.S.; Liu, J.O. Recent Advances in Drug Repositioning for the Discovery of New Anticancer Drugs. Int. J. Boil. Sci. 2014, 10, 654–663. [Google Scholar] [CrossRef] [Green Version]
- Naveja, J.J.; Dueñas-González, A.; Medina-Franco, J.L. Chapter 12—Drug Repurposing for Epigenetic Targets Guided by Computational Methods. In Epi-Informatics; Medina-Franco, J.L., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 327–357. [Google Scholar]
- Li, Y.Y.; Jones, S.J.M. Drug repositioning for personalized medicine. Genome Med. 2012, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantziarka, P.; Bouche, G.; Meheus, L.; Sukhatme, V.; Sukhatme, V.P.; Vikas, P. The Repurposing Drugs in Oncology (ReDO) Project. Ecancermedicalscience 2014, 8, 442. [Google Scholar]
- Nygren, P.; Larsson, R. Drug repositioning from bench to bedside: Tumour remission by the antihelmintic drug mebendazole in refractory metastatic colon cancer. Acta Oncol. 2013, 53, 427–428. [Google Scholar] [CrossRef]
- Christman, J.K. 5-Azacytidine and 5-aza-2′-deoxycytidine as inhibitors of DNA methylation: Mechanistic studies and their implications for cancer therapy. Oncogene 2002, 21, 5483–5495. [Google Scholar] [CrossRef] [Green Version]
- Fenaux, P.; Mufti, G.J.; Hellström-Lindberg, E.; Santini, V.; Finelli, C.; Giagounidis, A.; Schoch, R.; Gattermann, N.; Sanz, G.; List, A.F.; et al. Efficacy of azacitidine compared with that of conventional care regimens in the treatment of higher-risk myelodysplastic syndromes: A randomised, open-label, phase III study. Lancet Oncol. 2009, 10, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Kantarjian, H.; Issa, J.-P.; Rosenfeld, C.S.; Bennett, J.M.; Albitar, M.; DiPersio, J.; Klimek, V.; Slack, J.; De Castro, C.; Ravandi, F.; et al. Decitabine improves patient outcomes in myelodysplastic syndromes. Cancer 2006, 106, 1794–1803. [Google Scholar] [CrossRef]
- Lee, W.J.; Zhu, B.T. Inhibition of DNA methylation by caffeic acid and chlorogenic acid, two common catechol-containing coffee polyphenols. Carcinogenesis 2005, 27, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Oodi, A.; Norouzi, H.; Amirizadeh, N.; Nikougoftar, M.; Vafaie, Z. Harmine, a Novel DNA Methyltransferase 1 Inhibitor in the Leukemia Cell Line. Indian J. Hematol. Blood Transfus. 2017, 33, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Lu, Q.; Zhang, Z.; Rao, T.; Attwood, J.; Yung, R.; Richardson, B. Hydralazine may induce autoimmunity by inhibiting extracellular signal-regulated kinase pathway signaling. Arthritis Rheum. 2003, 48, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Segura-Pacheco, B.; Trejo-Becerril, C.; Perez-Cardenas, E.; Taja, L.; Mariscal, I.; Chavez, A.; Acuña, C.; Salazar, A.M.; Lizano, M.; Dueñas-Gonzalez, A. Reactivation of tumor suppressor genes by the cardiovascular drugs hydralazine and procainamide and their potential use in cancer therapy. Clin. Cancer Res. 2003, 9, 1596–1603. [Google Scholar] [PubMed]
- Segura-Pacheco, B.; Perez-Cardenas, E.; Taja-Chayeb, L.; Chávez-Blanco, A.D.; Vázquez, A.L.R.; Bribiesca, L.B.; Duenas-Gonzalez, A. Global DNA hypermethylation-associated cancer chemotherapy resistance and its reversion with the demethylating agent hydralazine. J. Transl. Med. 2006, 4, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Zhang, C. Hydralazine inhibits human cervical cancer cell growth in vitro in association with APC demethylation and re-expression. Cancer Chemother. Pharmacol. 2008, 63, 605–613. [Google Scholar] [CrossRef]
- Graça, M.I.P.D.S.; Sousa, E.J.; Costa-Pinheiro, P.; Vieira, A.F.Q.; Torres-Ferreira, J.; Martins, M.G.; Henrique, R.; Jerónimo, C. Anti-neoplastic properties of hydralazine in prostate cancer. Oncotarget 2014, 5, 5950–5964. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Stewart, A.; Maity, B.; Hagen, J.; Fagan, R.L.; Yang, J.; Quelle, D.E.; Brenner, C.; Fisher, R.A. RGS6 suppresses Ras-induced cellular transformation by facilitating Tip60-mediated Dnmt1 degradation and promoting apoptosis. Oncogene 2013, 33, 3604–3611. [Google Scholar] [CrossRef] [Green Version]
- Fagan, R.L.; Cryderman, D.E.; Kopelovich, L.; Wallrath, L.; Brenner, C. Laccaic Acid A Is a Direct, DNA-competitive Inhibitor of DNA Methyltransferase 1*. J. Boil. Chem. 2013, 288, 23858–23867. [Google Scholar] [CrossRef] [Green Version]
- Jagadeesh, S.; Sinha, S.; Pal, B.C.; Bhattacharya, S.; Banerjee, P.P. Mahanine reverses an epigenetically silenced tumor suppressor gene RASSF1A in human prostate cancer cells. Biochem. Biophys. Res. Commun. 2007, 362, 212–217. [Google Scholar] [CrossRef]
- Agarwal, S.; Amin, K.S.; Jagadeesh, S.; Baishya, G.; Rao, P.G.; Barua, N.C.; Bhattacharya, S.; Banerjee, P.P. Mahanine restores RASSF1A expression by down-regulating DNMT1 and DNMT3B in prostate cancer cells. Mol. Cancer 2013, 12, 99. [Google Scholar] [CrossRef] [Green Version]
- Lin, R.-K.; Hsu, C.-H.; Wang, Y.-C. Mithramycin A inhibits DNA methyltransferase and metastasis potential of lung cancer cells. Anti-Cancer Drugs 2007, 18, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Kuck, D.; Caulfield, T.; Lyko, F.; Medina-Franco, J.L. Nanaomycin A Selectively Inhibits DNMT3B and Reactivates Silenced Tumor Suppressor Genes in Human Cancer Cells. Mol. Cancer Ther. 2010, 9, 3015–3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, C.J.; Zheng, W.; Lodh, A.; Yevtodiyenko, A.; Liefwalker, D.; Li, H.; Felsher, D.W.; Van Riggelen, J. DNMT3B overexpression contributes to aberrant DNA methylation and MYC-driven tumor maintenance in T-ALL and Burkitt’s lymphoma. Oncotarget 2017, 8, 76898–76920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez-Lucio, O.; Tran, J.; Medina-Franco, J.L.; Meurice, N.; Muller, M. Toward Drug Repurposing in Epigenetics: Olsalazine as a Hypomethylating Compound Active in a Cellular Context. ChemMedChem 2014, 9, 560–565. [Google Scholar] [CrossRef]
- Lin, X.; Asgari, K.; Putzi, M.J.; Gage, W.R.; Yu, X.; Cornblatt, B.S.; Kumar, A.; Piantadosi, S.; Deweese, T.L.; De Marzo, A.M.; et al. Reversal of GSTP1 CpG island hypermethylation and reactivation of pi-class glutathione S-transferase (GSTP1) expression in human prostate cancer cells by treatment with procainamide. Cancer Res. 2001, 61, 8611–8616. [Google Scholar]
- Jablons, D.; Gao, Z.; Xu, Z.; Hung, M.-S.; Lin, Y.-C.; Wang, T.; Gong, M.; Zhi, X.; You, L. Procaine and procainamide inhibit the Wnt canonical pathway by promoter demethylation of WIF-1 in lung cancer cells. Oncol. Rep. 2009, 22, 1479–1484. [Google Scholar] [CrossRef]
- Lee, B.H.; Yegnasubramanian, S.; Lin, X.; Nelson, W. Procainamide Is a Specific Inhibitor of DNA Methyltransferase 1. J. Boil. Chem. 2005, 280, 40749–40756. [Google Scholar] [CrossRef] [Green Version]
- Villar-Garea, A.; Fraga, M.F.; Espada, J.; Esteller, M. Procaine is a DNA-demethylating agent with growth-inhibitory effects in human cancer cells. Cancer Res. 2003, 63, 4984–4989. [Google Scholar]
- Tada, M.; Imazeki, F.; Fukai, K.; Sakamoto, A.; Arai, M.; Mikata, R.; Tokuhisa, T.; Yokosuka, O. Procaine inhibits the proliferation and DNA methylation in human hepatoma cells. Hepatol. Int. 2007, 1, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-C.; Wang, Y.; Li, D.-D.; Zhang, Y.; Li, C.-F.; Zhao, T.-C. Procaine is a specific DNA methylation inhibitor with anti-tumor effect for human gastric cancer. J. Cell. Biochem. 2017, 119, 2440–2449. [Google Scholar] [CrossRef]
- Wong, K.K.; Lawrie, C.H.; Green, T.M. Oncogenic Roles and Inhibitors of DNMT1, DNMT3A, and DNMT3B in Acute Myeloid Leukaemia. Biomark. Insights 2019, 14, 1177271919846454. [Google Scholar] [CrossRef] [PubMed]
- Arce, C.; Segura-Pacheco, B.; Perez-Cardenas, E.; Taja-Chayeb, L.; Candelaria, M.; Duenas-Gonzalez, A. Hydralazine target: From blood vessels to the epigenome. J. Transl. Med. 2006, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeles, E.; HugoVazquez-Valadez, V.; Vazquez-Valadez, O.; Velazquez-Sanchez, A.; Ramirez, A.; Martínez, L.; Díaz-Barriga, S.; Romero-Rojas, A.; Cabrera, G.; Lopez-Castañares, R.; et al. Computational Studies of 1-Hydrazinophthalazine (Hydralazine) as Antineoplasic Agent. Docking Studies on Methyltransferase. Lett. Drug Des. Discov. 2005, 2, 282–286. [Google Scholar] [CrossRef]
- Hong, J.; Ishihara, K.; Yamaki, K.; Hiraizumi, K.; Ohno, T.; Ahn, J.W.; Zee, O.; Ohuchi, K. Apicidin, a histone deacetylase inhibitor, induces differentiation of HL-60 cells. Cancer Lett. 2003, 189, 197–206. [Google Scholar] [CrossRef]
- Wu, L.-P.; Wang, X.; Li, L.; Zhao, Y.; Lu, S.; Yu, Y.; Zhou, W.; Liu, X.; Yang, J.; Zheng, Z.; et al. Histone Deacetylase Inhibitor Depsipeptide Activates Silenced Genes through Decreasing both CpG and H3K9 Methylation on the Promoter. Mol. Cell. Boil. 2008, 28, 3219–3235. [Google Scholar] [CrossRef] [Green Version]
- Durczak, M.; Jagodzinski, P.P. Apicidin upregulates PHD2 prolyl hydroxylase gene expression in cervical cancer cells. Anti-Cancer Drugs 2010, 21, 619–624. [Google Scholar] [CrossRef]
- Im, J.Y.; Park, H.; Kang, K.W.; Choi, W.S.; Kim, H.S. Modulation of cell cycles and apoptosis by apicidin in estrogen receptor (ER)-positive and-negative human breast cancer cells. Chem. Interact. 2008, 172, 235–244. [Google Scholar] [CrossRef]
- Reddy, E.S.P.; Fortson, W.S.; Kayarthodi, S.; Fujimura, Y.; Xu, H.; Matthews, R.; Grizzle, W.E.; Rao, V.N.; Bhat, G.K. Histone deacetylase inhibitors, valproic acid and trichostatin-A induce apoptosis and affect acetylation status of p53 in ERG-positive prostate cancer cells. Int. J. Oncol. 2011, 39, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Ahn, M.Y.; Kang, D.O.; Na, Y.J.; Yoon, S.; Choi, W.S.; Kang, K.W.; Chung, H.Y.; Jung, J.H.; Min, S.; Kim, H.S. Histone deacetylase inhibitor, apicidin, inhibits human ovarian cancer cell migration via class II histone deacetylase 4 silencing. Cancer Lett. 2012, 325, 189–199. [Google Scholar] [CrossRef]
- Ahn, M. HDAC inhibitor apicidin suppresses murine oral squamous cell carcinoma cell growth in vitro and in vivo via inhibiting HDAC8 expression. Oncol. Lett. 2018, 16, 6552–6560. [Google Scholar] [CrossRef] [Green Version]
- Kumari, K.; Keshari, S.; Sengupta, D.; Sabat, S.C.; Mishra, S.K. Transcriptome analysis of genes associated with breast cancer cell motility in response to Artemisinin treatment. BMC Cancer 2017, 17, 858. [Google Scholar] [CrossRef] [Green Version]
- Pandey, M.; Kaur, P.; Shukla, S.; Abbas, A.; Fu, P.; Gupta, S. Plant flavone apigenin inhibits HDAC and remodels chromatin to induce growth arrest and apoptosis in human prostate cancer cells: In vitro and in vivo study. Mol. Carcinog. 2011, 51, 952–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Q.; Chen, X.; Sun, L.; Zhao, C.; Sui, G.; Cai, L. Carbamazepine promotes Her-2 protein degradation in breast cancer cells by modulating HDAC6 activity and acetylation of Hsp90. Mol. Cell. Biochem. 2010, 348, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Beutler, A.; Li, S.; Nicol, R.; Walsh, M.J. Carbamazepine is an inhibitor of histone deacetylases. Life Sci. 2005, 76, 3107–3115. [Google Scholar] [CrossRef]
- Akbarzadeh, L.; Zanjani, T.M.; Sabetkasaei, M. Comparison of Anticancer Effects of Carbamazepine and Valproic Acid. Iran. Red Crescent Med. J. 2016, 18, 37230. [Google Scholar] [CrossRef] [Green Version]
- Byun, M.R.; Lee, D.H.; Jang, Y.P.; Lee, H.S.; Choi, J.W.; Lee, S. Repurposing natural products as novel HDAC inhibitors by comparative analysis of gene expression profiles. Phytomedicine 2019, 59, 152900. [Google Scholar] [CrossRef]
- Joung, K.E.; Kim, D.-K.; Sheen, Y. Antiproliferative effect of trichostatin A and HC-toxin in T47D human breast cancer cells. Arch. Pharmacal Res. 2004, 27, 640–645. [Google Scholar] [CrossRef]
- Deubzer, H.E.; Ehemann, V.; Westermann, F.; Heinrich, R.; Mechtersheimer, G.; Kulozik, A.E.; Schwab, M.; Witt, O. Histone deacetylase inhibitor Helminthosporium carbonum (HC)-toxin suppresses the malignant phenotype of neuroblastoma cells. Int. J. Cancer 2007, 122, 1891–1900. [Google Scholar] [CrossRef]
- Pina, I.C.; Gautschi, J.T.; Wang, G.-Y.-S.; Sanders, M.L.; Schmitz, F.J.; France, D.; Cornell-Kennon, S.; Sambucetti, L.C.; Remiszewski, S.W.; Perez, L.B.; et al. Psammaplins from the SpongePseudoceratinapurpurea:Inhibition of Both Histone Deacetylase and DNA Methyltransferase. J. Org. Chem. 2003, 68, 3866–3873. [Google Scholar] [CrossRef]
- Baud, M.; Leiser, T.; Haus, P.; Samlal, S.; Wong, A.C.; Wood, R.J.; Petrucci, V.; Gunaratnam, M.; Hughes, S.M.; Buluwela, L.; et al. Defining the Mechanism of Action and Enzymatic Selectivity of Psammaplin A against Its Epigenetic Targets. J. Med. Chem. 2012, 55, 1731–1750. [Google Scholar] [CrossRef]
- Kim, T.H.; Kim, H.S.; Kang, Y.J.; Yoon, S.; Lee, J.; Choi, W.S.; Jung, J.H.; Kim, H.S. Psammaplin A induces Sirtuin 1-dependent autophagic cell death in doxorubicin-resistant MCF-7/adr human breast cancer cells and xenografts. Biochim. Biophys. Acta 2015, 1850, 401–410. [Google Scholar] [PubMed]
- Ahn, M.Y.; Jung, J.H.; Na, Y.J.; Kim, H.S. A natural histone deacetylase inhibitor, Psammaplin A, induces cell cycle arrest and apoptosis in human endometrial cancer cells. Gynecol. Oncol. 2008, 108, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Shin, J.; Kwon, H.J. Psammaplin A is a natural prodrug that inhibits class I histone deacetylase. Exp. Mol. Med. 2007, 39, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Lee, Y.S.; Lee, Y.C. Sodium butyrate-induced DAPK-mediated apoptosis in human gastric cancer cells. Oncol. Rep. 2011, 27, 1111–1115. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Sun, Y.; Liu, J.; Wu, X.; Chen, L.; Ma, L.; Wu, P. Histone deacetylase inhibitor sodium butyrate suppresses DNA double strand break repair induced by etoposide more effectively in MCF-7 cells than in HEK293 cells. BMC Biochem. 2015, 16, 2. [Google Scholar] [CrossRef] [Green Version]
- Cang, S.; Xu, X.; Ma, Y.; Liu, D.; Chiao, J.W. Hypoacetylation, hypomethylation, and dephosphorylation of H2B histones and excessive histone deacetylase activity in DU-145 prostate cancer cells. J. Hematol. Oncol. 2016, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Vigushin, D.M.; Ali, S.; Pace, P.E.; Mirsaidi, N.; Ito, K.; Adcock, I.; Coombes, R.C. Trichostatin A is a histone deacetylase inhibitor with potent antitumor activity against breast cancer in vivo. Clin. Cancer Res. 2001, 7, 971–976. [Google Scholar]
- Chambers, A.; Banerjee, S.; Chaplin, T.; Dunne, J.; Debernardi, S.; Joel, S.; Young, B. Histone acetylation-mediated regulation of genes in leukaemic cells. Eur. J. Cancer 2003, 39, 1165–1175. [Google Scholar] [CrossRef]
- Ma, J.; Guo, X.; Zhang, S.; Liu, H.; Lu, J.; Dong, Z.; Liu, K.; Ming, L. Trichostatin A, a histone deacetylase inhibitor, suppresses proliferation and promotes apoptosis of esophageal squamous cell lines. Mol. Med. Rep. 2015, 11, 4525–4531. [Google Scholar] [CrossRef]
- Motawi, T.; Darwish, H.A.; Diab, I.; Helmy, M.W.; Noureldin, M.H. Combinatorial strategy of epigenetic and hormonal therapies: A novel promising approach for treating advanced prostate cancer. Life Sci. 2018, 198, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, X.; Liu, H.; Jin, H.; Ji, Y. Trichostatin A inhibits proliferation of PC3 prostate cancer cells by disrupting the EGFR pathway. Oncol. Lett. 2019, 18, 687–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Liu, X.; Zhu, S.; Hu, X.; Niu, H.; Zhang, X.; Zhu, D.; Nesa, E.U.; Tian, K.; Yuan, H. Hyper-acetylation contributes to the sensitivity of chemo-resistant prostate cancer cells to histone deacetylase inhibitor Trichostatin A. J. Cell. Mol. Med. 2018, 22, 1909–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiffon, C. Histone Deacetylase Inhibition Restores Expression of Hypoxia-Inducible Protein NDRG1 in Pancreatic Cancer. Pancreas 2018, 47, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Kavoosi, F.; Sanaei, M. Effect of 5-aza-2′-deoxycytidine in comparison to valproic acid and trichostatin a on histone deacetylase 1, dna methyltransferase 1, and cip/kip family (p21, p27, and p57) genes expression, cell growth inhibition, and apoptosis induction in colon cancer sw480 cell line. Adv. Biomed. Res. 2019, 8, 52. [Google Scholar] [CrossRef]
- Hernández-Oliveras, A.; Izquierdo-Torres, E.; Meneses-Morales, I.; Rodríguez, G.; Zarain-Herzberg, A.; Santiago-García, J. Histone deacetylase inhibitors promote ATP2A3 gene expression in hepatocellular carcinoma cells: p300 as a transcriptional regulator. Int. J. Biochem. Cell Boil. 2019, 113, 8–16. [Google Scholar] [CrossRef]
- Ahn, M.; Chung, H.Y.; Choi, W.S.; Lee, B.-M.; Yoon, S.; Kim, H.S. Anti-tumor effect of apicidin on Ishikawa human endometrial cancer cells both in vitro and in vivo by blocking histone deacetylase 3 and 4. Int. J. Oncol. 2010, 36, 125–131. [Google Scholar]
- You, J.S.; Kang, J.; Lee, E.K.; Lee, J.C.; Lee, S.H.; Jeon, Y.J.; Koh, D.H.; Ahn, S.H.; Seo, D.-W.; Lee, H.Y.; et al. Histone deacetylase inhibitor apicidin downregulates DNA methyltransferase 1 expression and induces repressive histone modifications via recruitment of corepressor complex to promoter region in human cervix cancer cells. Oncogene 2007, 27, 1376–1386. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanyam, K.; Swaminathan, V.; Ranganathan, A.; Kundu, T.K. Small Molecule Modulators of Histone Acetyltransferase p300. J. Boil. Chem. 2003, 278, 19134–19140. [Google Scholar] [CrossRef] [Green Version]
- Sung, B.; Pandey, M.K.; Ahn, K.S.; Yi, T.; Chaturvedi, M.M.; Liu, M.; Aggarwal, B.B. Anacardic acid (6-nonadecyl salicylic acid), an inhibitor of histone acetyltransferase, suppresses expression of nuclear factor-kappaB-regulated gene products involved in cell survival, proliferation, invasion, and inflammation through inhibition of the inhibitory subunit of nuclear factor-kappaBalpha kinase, leading to potentiation of apoptosis. Blood 2008, 111, 4880–4891. [Google Scholar]
- Han, H.; Yang, X.; Pandiyan, K.; Liang, G. Synergistic Re-Activation of Epigenetically Silenced Genes by Combinatorial Inhibition of DNMTs and LSD1 in Cancer Cells. PLoS ONE 2013, 8, e75136. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanyam, K.; Altaf, M.; Varier, R.A.; Swaminathan, V.; Ravindran, A.; Sadhale, P.P.; Kundu, T.K. Polyisoprenylated Benzophenone, Garcinol, a Natural Histone Acetyltransferase Inhibitor, Represses Chromatin Transcription and Alters Global Gene Expression. J. Boil. Chem. 2004, 279, 33716–33726. [Google Scholar] [CrossRef] [Green Version]
- Collins, H.; Abdelghany, M.K.; Messmer, M.; Yue, B.; Deeves, S.E.; Kindle, K.B.; Mantelingu, K.; Aslam, A.; Winkler, S.; Kundu, T.K.; et al. Differential effects of garcinol and curcumin on histone and p53 modifications in tumour cells. BMC Cancer 2013, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Sethi, G.; Chatterjee, S.; Rajendran, P.; Li, F.; Shanmugam, M.K.; Wong, K.-F.; Kumar, A.P.; Senapati, P.; Behera, A.K.; Hui, K.M.; et al. Inhibition of STAT3 dimerization and acetylation by garcinol suppresses the growth of human hepatocellular carcinoma in vitro and in vivo. Mol. Cancer 2014, 13, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wu, M.; Zheng, D.; Zhang, H.; Lv, Y.; Zhang, L.; Tan, H.S.; Zhou, H.; Lao, Y.Z.; Xu, H.X. Garcinol inhibits esophageal cancer metastasis by suppressing the p300 and TGF-beta1 signaling pathways. Acta Pharmacol. Sin. 2019, 41, 82–92. [Google Scholar] [PubMed]
- Sakane, C.; Okitsu, T.; Wada, A.; Sagami, H.; Shidoji, Y. Inhibition of lysine-specific demethylase 1 by the acyclic diterpenoid geranylgeranoic acid and its derivatives. Biochem. Biophys. Res. Commun. 2014, 444, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Xing, J.; Wang, C.; Zhang, H.; Yue, L.; Wan, X.; Chen, W.; Ding, H.; Xie, Y.; Tao, H.; et al. Discovery of novel BET inhibitors by drug repurposing of nitroxoline and its analogues. Org. Biomol. Chem. 2017, 15, 9352–9361. [Google Scholar] [CrossRef]
- Wang, M.; Liu, X.; Guo, J.; Weng, X.; Jiang, G.; Wang, Z.; He, L. Inhibition of LSD1 by Pargyline inhibited process of EMT and delayed progression of prostate cancer in vivo. Biochem. Biophys. Res. Commun. 2015, 467, 310–315. [Google Scholar] [CrossRef]
- Ravindra, K.C.; Selvi, B.R.; Arif, M.; Reddy, B.A.A.; Thanuja, G.R.; Agrawal, S.; Pradhan, S.K.; Nagashayana, N.; Dasgupta, D.; Kundu, T.K. Inhibition of Lysine Acetyltransferase KAT3B/p300 Activity by a Naturally Occurring Hydroxynaphthoquinone, Plumbagin*. J. Boil. Chem. 2009, 284, 24453–24464. [Google Scholar] [CrossRef] [Green Version]
- Casaos, J.; Huq, S.; Lott, T.; Felder, R.; Choi, J.; Gorelick, N.; Peters, M.; Xia, Y.; Maxwell, R.; Zhao, T.; et al. Ribavirin as a potential therapeutic for atypical teratoid/rhabdoid tumors. Oncotarget 2018, 9, 8054–8067. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Xu, X.; Chen, J. Clinically relevant concentration of anti-viral drug ribavirin selectively targets pediatric osteosarcoma and increases chemosensitivity. Biochem. Biophys. Res. Commun. 2018, 506, 604–610. [Google Scholar] [CrossRef]
- De La Cruz-Hernández, E.; Medina-Franco, J.L.; Trujillo, J.; Chávez-Blanco, A.D.; Dominguez-Gomez, G.; Perez-Cardenas, E.; Gonzalez-Fierro, A.; Taja, L.; Dueñas-Gonzalez, A. Ribavirin as a tri-targeted antitumor repositioned drug. Oncol. Rep. 2015, 33, 2384–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.M.; Manton, C.; Bhat, K.P.; Tsai, W.-W.; Aldape, K.; Barton, S.; Chandra, J. Inhibition of LSD1 sensitizes glioblastoma cells to histone deacetylase inhibitors. Neuro-Oncology 2011, 13, 894–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennani-Baiti, I.M.; Machado, I.; Llombart-Bosch, A.; Kovar, H. Lysine-specific demethylase 1 (LSD1/KDM1A/AOF2/BHC110) is expressed and is an epigenetic drug target in chondrosarcoma, Ewing’s sarcoma, osteosarcoma, and rhabdomyosarcoma. Hum. Pathol. 2012, 43, 1300–1307. [Google Scholar] [PubMed]
- Lee, M.G.; Wynder, C.; Schmidt, D.M.; McCafferty, D.G.; Shiekhattar, R. Histone H3 Lysine 4 Demethylation Is a Target of Nonselective Antidepressive Medications. Chem. Boil. 2006, 13, 563–567. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Jiang, X.; Chen, S.; Price, B.D. Inhibition of histone acetyltransferase activity by anacardic acid sensitizes tumor cells to ionizing radiation. FEBS Lett. 2006, 580, 4353–4356. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Chen, S.; Gao, C.; Dai, Q.; Zhang, C.; Sun, Q.; Lin, J.-S.; Guo, C.; Chen, Y.Z.; Jiang, Y. Development of a versatile DNMT and HDAC inhibitor C02S modulating multiple cancer hallmarks for breast cancer therapy. Bioorganic Chem. 2019, 87, 200–208. [Google Scholar] [CrossRef]
- Huang, Z.-H.; Zheng, H.-F.; Wang, W.-L.; Wang, Y.; Zhong, L.-F.; Wu, J.-L.; Li, Q.-X. Berberine targets epidermal growth factor receptor signaling to suppress prostate cancer proliferation in vitro. Mol. Med. Rep. 2014, 11, 2125–2128. [Google Scholar] [CrossRef] [Green Version]
- Qing, Y.; Hu, H.; Liu, Y.; Feng, T.; Meng, W.; Jiang, L.; Sun, Y.; Yao, Y. Berberine induces apoptosis in human multiple myeloma cell line U266 through hypomethylation of p53 promoter. Cell Boil. Int. 2014, 38, 563–570. [Google Scholar] [CrossRef]
- Kalaiarasi, A.; Anusha, C.; Sankar, R.; Rajasekaran, S.; Marshal, J.J.; Muthusamy, K.; Ravikumar, V. Plant Isoquinoline Alkaloid Berberine Exhibits Chromatin Remodeling by Modulation of Histone Deacetylase To Induce Growth Arrest and Apoptosis in the A549 Cell Line. J. Agric. Food Chem. 2016, 64, 9542–9550. [Google Scholar] [CrossRef]
- Gopal, Y.V.; Arora, T.S.; Van Dyke, M.W. Parthenolide Specifically Depletes Histone Deacetylase 1 Protein and Induces Cell Death through Ataxia Telangiectasia Mutated. Chem. Boil. 2007, 14, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Dawood, M.; Ooko, E.; Efferth, T. Collateral Sensitivity of Parthenolide via NF-κB and HIF-α Inhibition and Epigenetic Changes in Drug-Resistant Cancer Cell Lines. Front. Pharmacol. 2019, 10, 542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Liu, S.; Xie, Z.; Pavlovicz, R.E.; Wu, J.; Chen, P.; Aimiuwu, J.; Pang, J.; Bhasin, D.; Neviani, P.; et al. Modulation of DNA methylation by a sesquiterpene lactone parthenolide. J. Pharmacol. Exp. Ther. 2009, 329, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Hartman, M.L.; Talar, B.; Sztiller-Sikorska, M.; Nejc, D.; Czyz, M. Parthenolide induces MITF-M downregulation and senescence in patient-derived MITF-M(high) melanoma cell populations. Oncotarget 2016, 7, 9026–9040. [Google Scholar]
- Fudhaili, A.; Yoon, N.A.; Kang, S.; Ryu, J.; Jeong, J.Y.; Lee, N.H.; Kang, S.S. Resveratrol epigenetically regulates the expression of zinc finger protein 36 in nonsmall cell lung cancer cell lines. Oncol. Rep. 2018, 41, 1377–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, B.; Ghosh, K.; Kanade, S.R. Resveratrol modulates epigenetic regulators of promoter histone methylation and acetylation that restores BRCA1, p53, p21CIP1 in human breast cancer cell lines. BioFactors 2019, 45, 818–829. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Torres, E.; Hernández-Oliveras, A.; Meneses-Morales, I.; Rodríguez, G.; Fuentes-García, G.; Zarain-Herzberg, A. Resveratrol up-regulates ATP2A3 gene expression in breast cancer cell lines through epigenetic mechanisms. Int. J. Biochem. Cell Boil. 2019, 113, 37–47. [Google Scholar] [CrossRef]
- Liu, X.; Li, H.; Wu, M.-L.; Wu, J.; Sun, Y.; Zhang, K.-L.; Liu, J. Resveratrol Reverses Retinoic Acid Resistance of Anaplastic Thyroid Cancer Cells via Demethylating CRABP2 Gene. Front. Endocrinol. 2019, 10, 734. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Y.; Xue, Y.; Hu, H.; Ye, J.; Li, X.; Lu, Z.-G.; Meng, F.; Liang, S. Berberine acts as a putative epigenetic modulator by affecting the histone code. Toxicol. Vitr. 2016, 36, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Koprowska, K.; Czyz, M. Molecular mechanisms of parthenolide’s action: Old drug with a new face. Postepy Hig. Med. Dosw. (Online) 2010, 64, 100–114. [Google Scholar]
- Venturelli, S.; Berger, A.; Böcker, A.; Busch, C.; Weiland, T.; Noor, S.; Leischner, C.; Schleicher, S.; Mayer, M.; Weiss, T.S.; et al. Resveratrol as a pan-HDAC inhibitor alters the acetylation status of histone [corrected] proteins in human-derived hepatoblastoma cells. PLoS ONE 2013, 8, e73097. [Google Scholar]
- Härmä, V.; Virtanen, J.; Mäkelä, R.; Happonen, A.; Mpindi, J.-P.; Knuuttila, M.; Kohonen, P.; Lötjönen, J.; Kallioniemi, O.-P.; Nees, M. A Comprehensive Panel of Three-Dimensional Models for Studies of Prostate Cancer Growth, Invasion and Drug Responses. PLoS ONE 2010, 5, e10431. [Google Scholar] [CrossRef]
- Mittler, F.; Obeïd, P.; Rulina, A.V.; Haguet, V.; Gidrol, X.; Maxim, B. High-Content Monitoring of Drug Effects in a 3D Spheroid Model. Front. Oncol. 2017, 7, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, D.; Buchanan, G. Stromal Androgen Receptor in Prostate Cancer Development and Progression. Cancers 2017, 9, 10. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Drug | Approved for | Epigenetic Target | Cancer Model |
|---|---|---|---|
| Chlorogenic Acid | Natural Compound (not approved) | DNMT1 | Breast Cancer [21] |
| Harmine | Natural Compound (not approved) | DNMT1 | Acute Myeloid Leukemia [22] |
| Hydralazine | Hypertension | DNMT1 | T-Cell Leukemia [23], Breast Cancer [24,25], Bladder Cancer [24], Cervical Cancer [26], Prostate Cancer [27] |
| Laccaic Acid A | Natural Compound (not approved) | DNMT1 | Breast Cancer [28,29] |
| Mahanine | Natural Compound (not approved) | DNMT1, DNMT3B | Prostate Cancer [30,31] |
| Mithramycin A | Hypercalcemia, especially due to malignancies | DNMT1 | Lung Cancer [32] |
| Nanaomycin A | Quinone antibiotic (not approved) | DNMT3B | Lung Cancer, Colon Cancer [33], T-Cell Acute Lymphoblastic Leukemia, Burkitt Lymphoma [34] |
| Olsalazine | Inflammatory bowel disease and ulcerative colitis | DNMT | Cervical Cancer [35] |
| Procainamide | Cardiac arrythmias | DNMT1 | Prostate Cancer [36], Breast Cancer, Bladder Cancer [37], Colon Cancer [38], Nonsmall Cell Lung Cancer [37] |
| Procaine | Infiltration anesthesia, peripheral nerve and spinal block | DNMT1, DNMT3A | Breast Cancer [39], Hepatocellular Carcinoma [40], Nonsmall Cell Lung Cancer [37], Gastric Cancer [41] |
| Drug | Approved for | Epigenetic Target | Cancer Model |
|---|---|---|---|
| Apicidin | Antiprotozoal (not approved) | HDAC3, HDAC4, HDAC8 | Acute Promyelocytic Leukemia [45], Lung Cancer, Colon Cancer, Pancreatic Cancer [46], Cervical Cancer [47] Breast Cancer [48], Endometrial Cancer [49], Ovarian Cancer [50], Oral Squamous Cell Carcinoma [51] |
| Artemisin | Malaria | HDAC1, HDAC2, HDAC6 | Breast Cancer [52] |
| Aspigenin | Natural Compound (not approved) | HDAC class I | Prostate Cancer [53] |
| Carbamazepine | Control of psychomotor or focal seizures | HDAC3, HDAC6, HDAC7 | Breast Cancer [54], Liver Cancer [55], Colon Cancer [56] |
| Ginseng | Natural Compound (not approved) | HDAC | Nonsmall Cell Lung Cancer [57] |
| HC Toxin | Natural Compound (not approved) | HDAC | Breast Cancer [58], Neuroblastoma [59] |
| Psammaplin A | Natural Compound (not approved) | HDAC1, HDAC6, SIRT1 | Lung Cancer [60], Breast Cancer [61,62], Endometrial Cancer [63], Cervical Cancer [64] |
| Sodium Butyrate | Anti-inflammatory | HDAC1 | Gastric Cancer [65], Breast Cancer [66], Prostate Cancer [67] |
| TSA | Antifungal antibiotic | HDAC class I, II and SIRT6 | Breast Cancer [68], Leukemia [69], Esophageal Squamous Carcinoma [70], Prostate Cancer [49,71,72,73], Pancreatic Cancer [74], Colon Cancer [75], Hepatocellular Carcinoma [76] |
| Drug | Approved for | Epigenetic Target | Cancer Model |
|---|---|---|---|
| Anarcadic Acid | Anti-inflammatory and radio-sensitization activities | Ep300 and Tip60 | Cervical Cancer [79], Myeloid Leukemia, T-Cell Lymphoma, Lung Cancer, Prostate Cancer [80] |
| Clorgyline | MAO inhibitor | LSD1 | Bladder Cancer, Colon Cancer, Leukemia [81] |
| Garcinol | Antioxidant (not approved) | Ep300 and KAT2B | Cervical Cancer [82], Breast Cancer [83], Hepatocellular Carcinoma [84], Esophageal Carcinoma [85] |
| Geranylgeranoic Acid | Natural Compound (not approved) | LSD1 | Neuroblastoma [86] |
| Nitroxoline | Urinary antibacterial agent | BRD4 | Mixed-Lineage Leukemia [87] |
| Pargyline | Irreversible selective MAO-B and antihypertensive | LSD1 | Prostate Cancer [88] |
| Plumbagin | Natural Compound (not approved) | KAT3B/p300 | Liver Carcinoma [89] |
| Ribavirin | RSV infections and Hepatitis C | EZH2 | Solid Tumors [90,91,92] |
| Tranylcypromine | Depression, Dysthymic disorder, atypical depression, panic and phobic disorders | LSD1 | Glioblastoma Multiforme [93], Sarcomas [94], Embryonal Carcinoma [95] |
| Drug | Approved for | Epigenetic Target | Cancer Model |
|---|---|---|---|
| Berberine | Parasitic and fungal infections | HDAC class I, II, IV and DNMT1, DNMT3A | Prostate Cancer [98], Multiple Myeloma [99], Lung Cancer [100] |
| Parthenolide | Anti-inflammatory (not approved) | HDAC1 and DNMT | Breast Cancer [101,102], Leukemia [103], Myeloma [104], Colon Cancer [102] |
| Resveratrol | Natural Compound (not approved) | HDAC and DNMT1 | Nonsmall Cell Lung Cancer [105], Breast Cancer [106,107], Thyroid Cancer [108] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira-Silva, F.; Camilo, V.; Gaspar, V.; Mano, J.F.; Henrique, R.; Jerónimo, C. Repurposing Old Drugs into New Epigenetic Inhibitors: Promising Candidates for Cancer Treatment? Pharmaceutics 2020, 12, 410. https://doi.org/10.3390/pharmaceutics12050410
Moreira-Silva F, Camilo V, Gaspar V, Mano JF, Henrique R, Jerónimo C. Repurposing Old Drugs into New Epigenetic Inhibitors: Promising Candidates for Cancer Treatment? Pharmaceutics. 2020; 12(5):410. https://doi.org/10.3390/pharmaceutics12050410
Chicago/Turabian StyleMoreira-Silva, Filipa, Vânia Camilo, Vítor Gaspar, João F. Mano, Rui Henrique, and Carmen Jerónimo. 2020. "Repurposing Old Drugs into New Epigenetic Inhibitors: Promising Candidates for Cancer Treatment?" Pharmaceutics 12, no. 5: 410. https://doi.org/10.3390/pharmaceutics12050410