Interaction of Lipoplex with Albumin Enhances Gene Expression in Hepatitis Mice

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. pDNA
2.4. Cationic Liposomes
2.5. Lipoplex
2.6. Preparation of Serum Components
2.7. In Vivo Transfection
2.8. Measurement of Serum Transaminase Activities
2.9. Evaluation of Hepatic Accumulation of Lipoplex
2.10. Cytokine ELISA
2.11. CCl4-Induced Hepatitis Mice
2.12. Statistical Analysis
3. Results
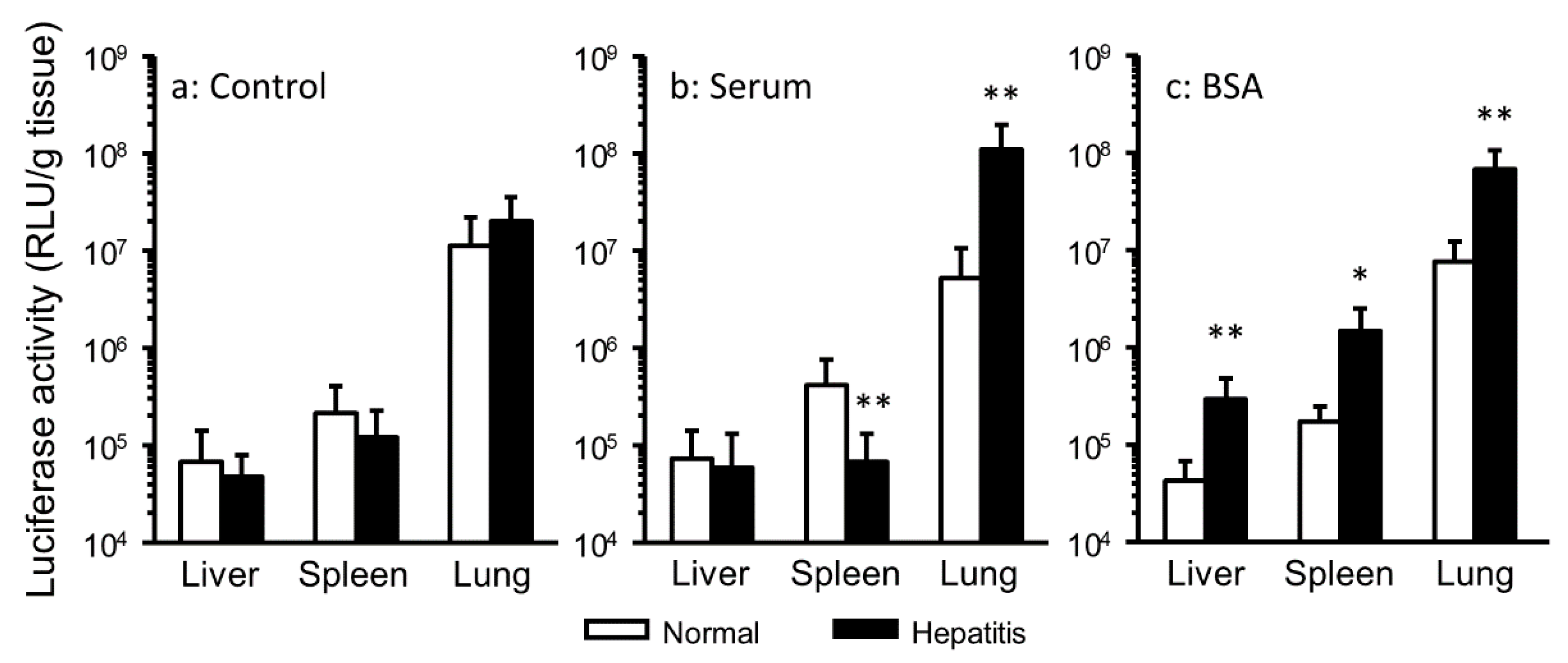
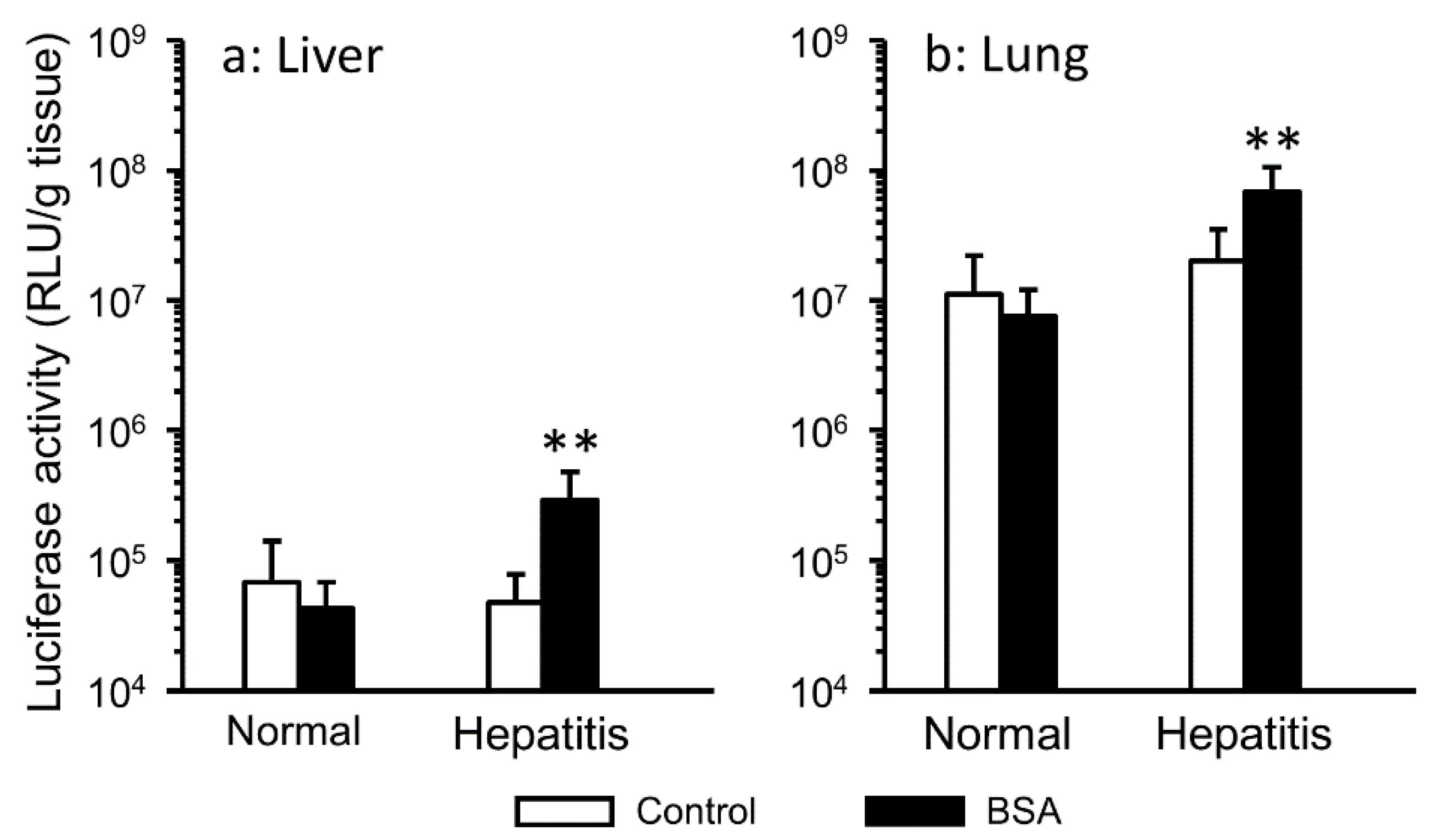
3.1. Effect of the Interaction with Serum Components on Gene Expression in Hepatitis Mice
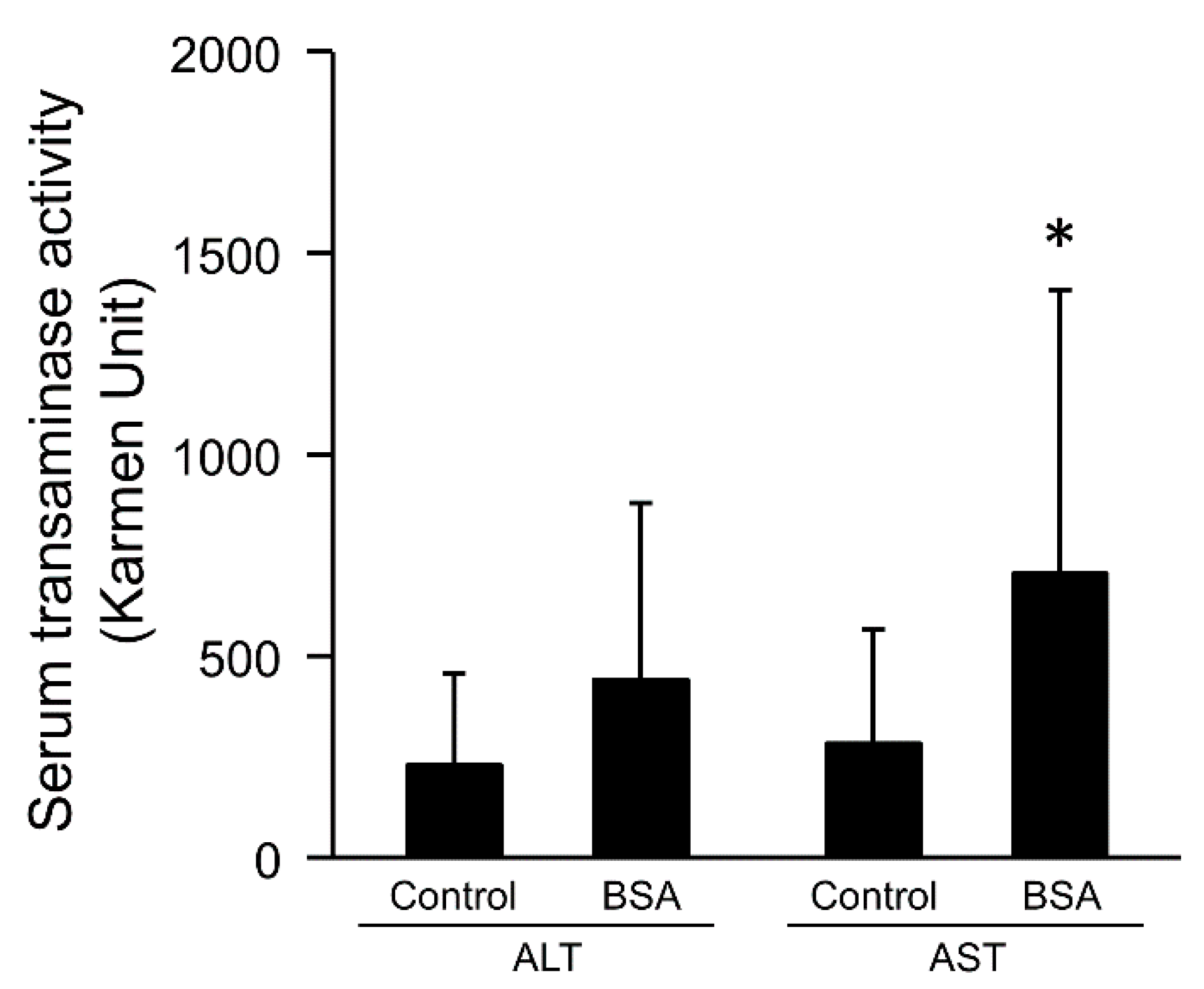
3.2. Effect of the Interaction with Serum Components on Serum Transaminase Activities
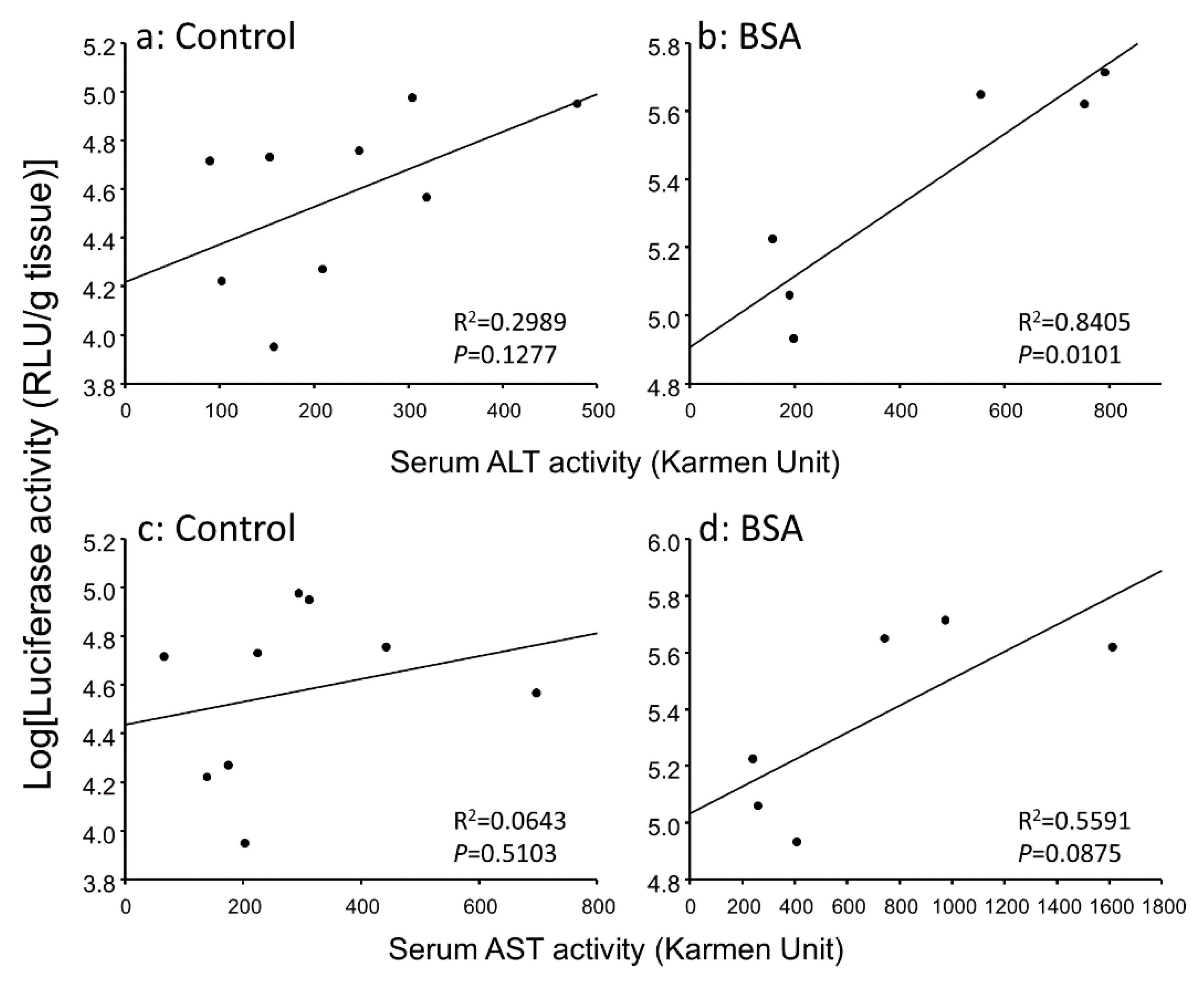
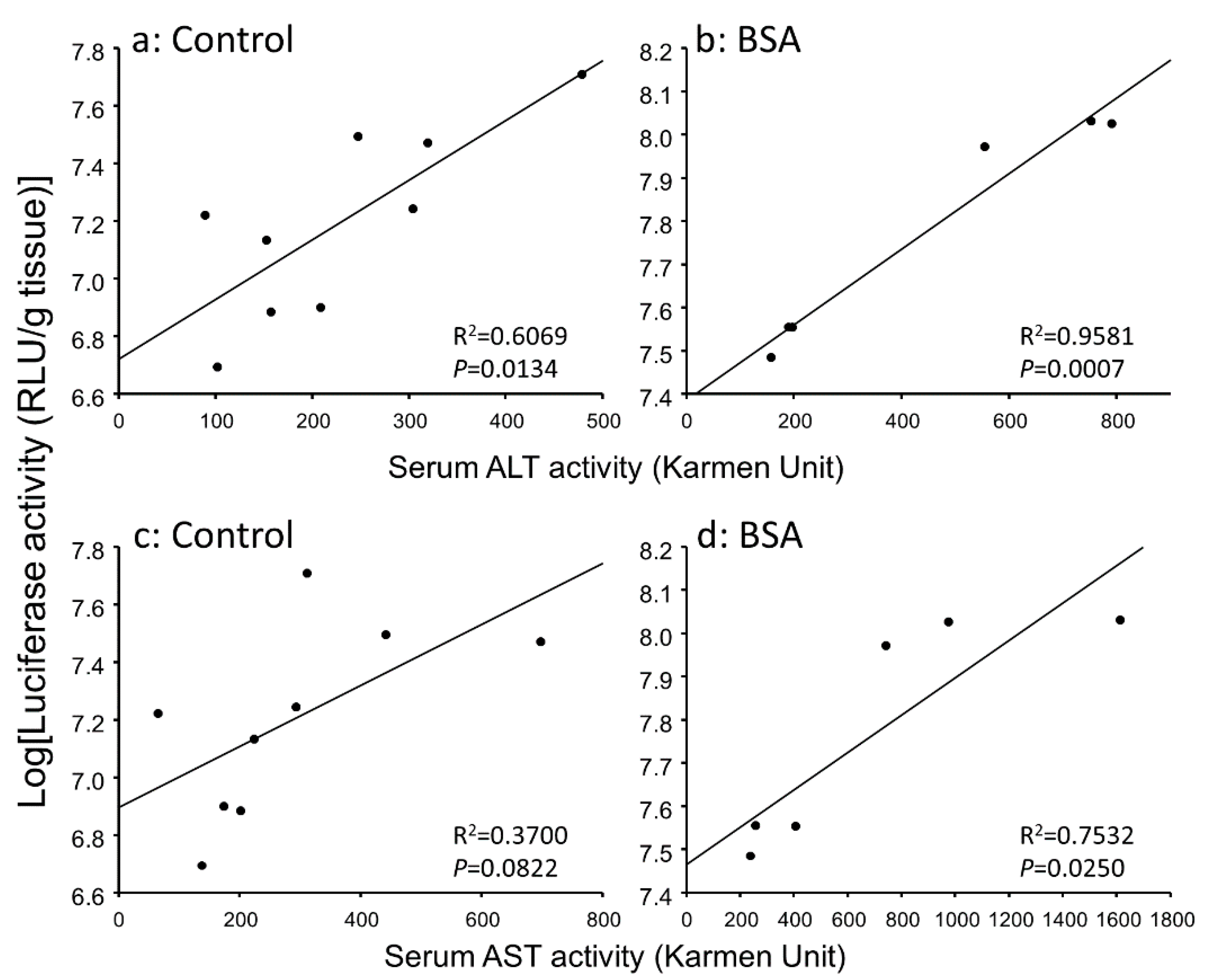
3.3. Correlation between Gene Expression Levels and Serum Transaminase Activity Levels in Hepatitis Mice
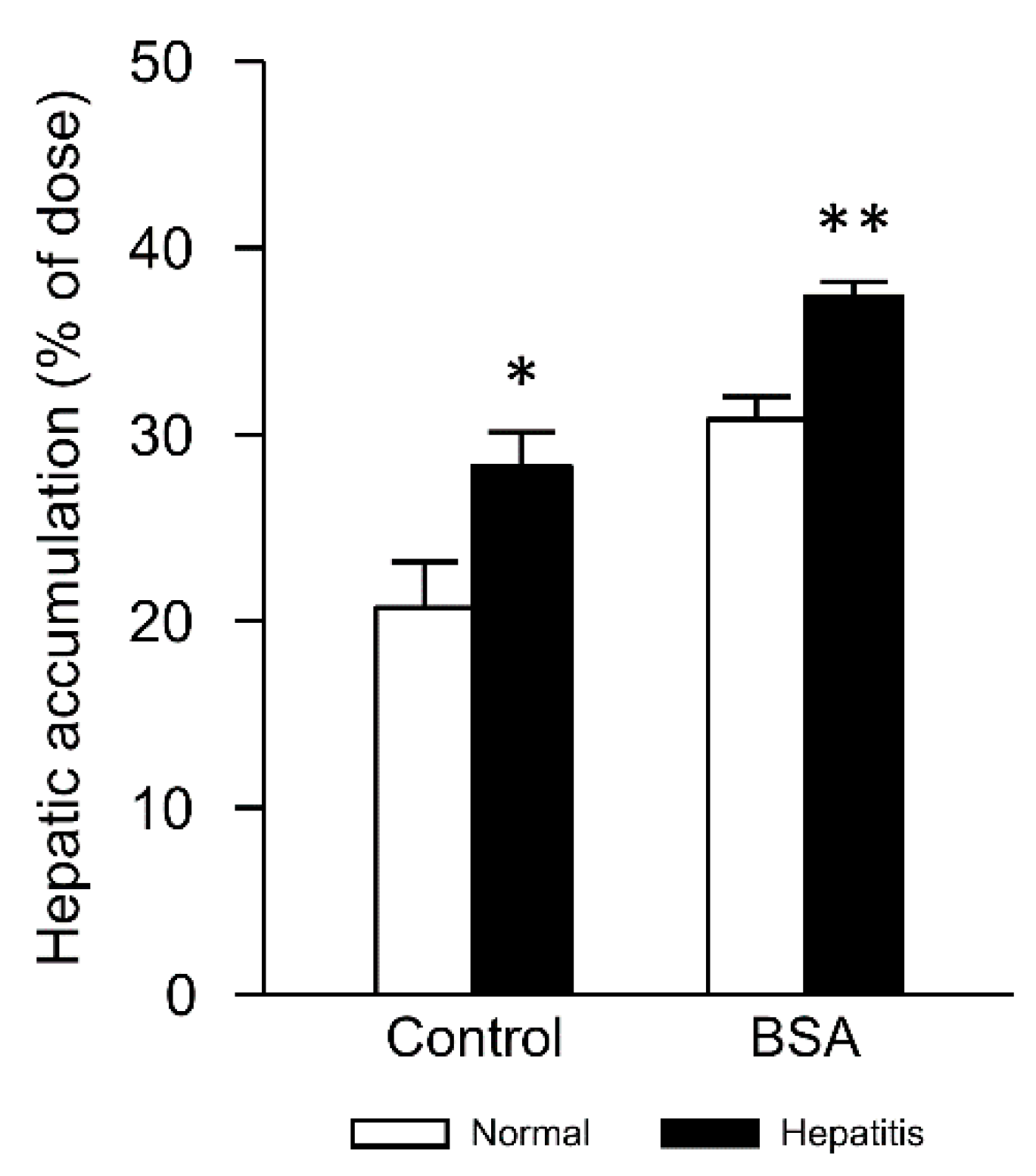
3.4. Changes in Hepatic Accumulation of Lipoplex by Interaction with Albumin in Hepatitis Mice
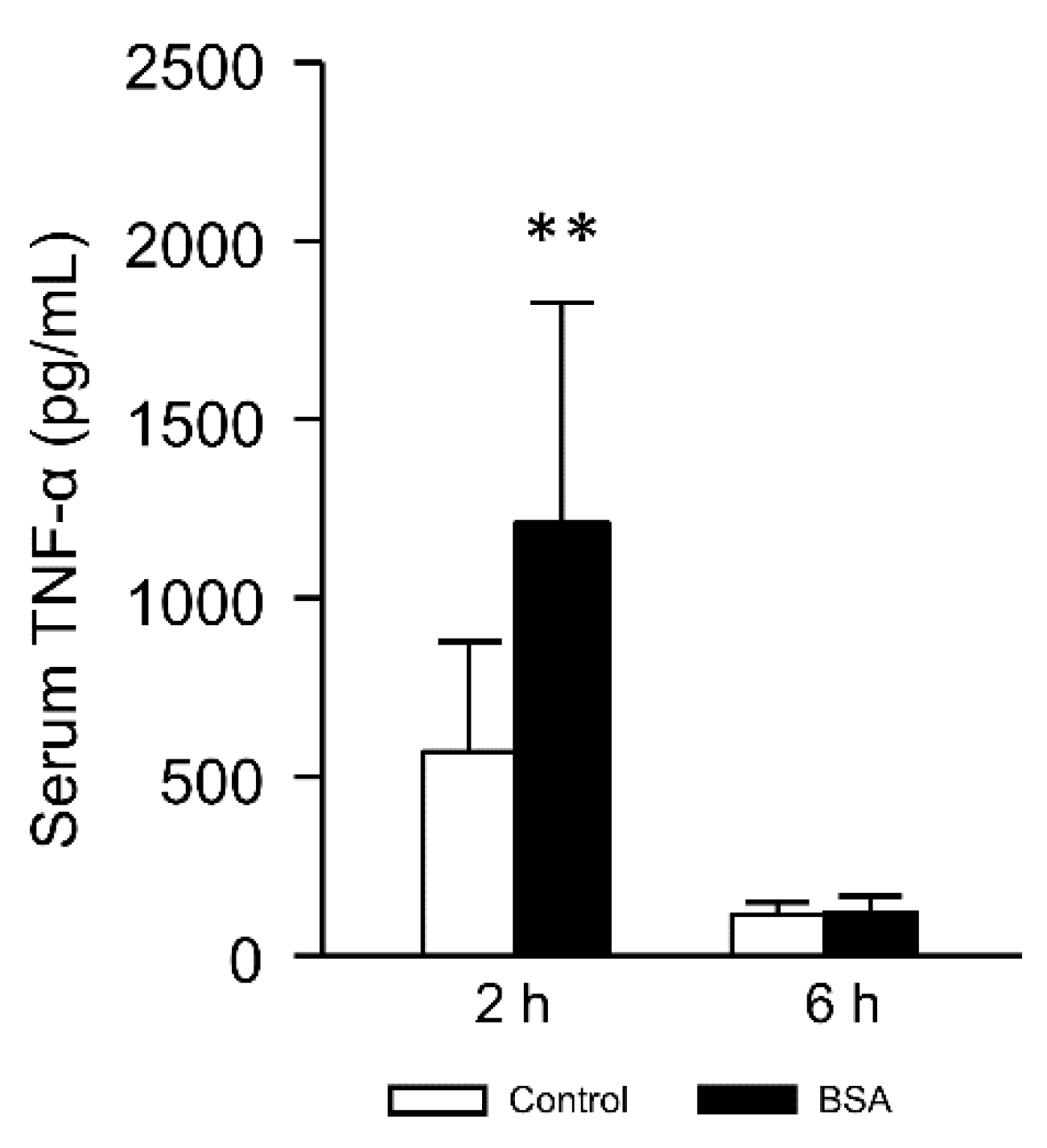
3.5. Effect of the Interaction with Albumin on Serum TNF-α Level
3.6. Effect of pH on Particle Size and Zeta Potential of Lipoplex Interacted with BSA
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shirley, J.L.; de Jong, Y.P.; Terhorst, C.; Herzog, R.W. Immune Responses to Viral Gene Therapy Vectors. Mol. Ther. 2020, 28, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.G.; Evans, A.C.; Ho, M.L.; Guenther, C.M.; Brun, M.J.; Judd, J.; Wu, E.; Suh, J. Reducing off target viral delivery in ovarian cancer gene therapy using a protease-activated AAV2 vector platform. J. Control. Release 2019, 307, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Bono, N.; Ponti, F.; Mantovani, D.; Candiani, G. Non-Viral in Vitro Gene Delivery: It is Now Time to Set the Bar! Pharmaceutics 2020, 12, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, R.E.; Zahid, M. Cell Penetrating Peptides, Novel Vectors for Gene Therapy. Pharmaceutics 2020, 12, 225. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Cheng, K. The principles and applications of avidin-based nanoparticles in drug delivery and diagnosis. J. Control. Release 2017, 245, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Taghavi, S.; Abnous, K.; Taghdisi, S.M.; Ramezani, M.; Alibolandi, M. Hybrid carbon-based materials for gene delivery in cancer therapy. J. Control. Release 2020, 318, 158–175. [Google Scholar] [CrossRef]
- Cordeiro, R.A.; Serra, A.; Coelho, J.F.J.; Faneca, H. Poly(β-amino ester)-based gene delivery systems: From discovery to therapeutic applications. J. Control. Release 2019, 310, 155–187. [Google Scholar] [CrossRef]
- Martínez-Negro, M.; Sánchez-Arribas, N.; Guerrero-Martínez, A.; Moyá, M.L.; Tros de Ilarduya, C.; Mendicuti, F.; Aicart, E.; Junquera, E. A Non-Viral Plasmid DNA Delivery System Consisting on a Lysine-Derived Cationic Lipid Mixed with a Fusogenic Lipid. Pharmaceutics 2019, 11, 632. [Google Scholar] [CrossRef] [Green Version]
- Palchetti, S.; Caputo, D.; Digiacomo, L.; Capriotti, A.L.; Coppola, R.; Pozzi, D.; Caracciolo, G. Protein Corona Fingerprints of Liposomes: New Opportunities for Targeted Drug Delivery and Early Detection in Pancreatic Cancer. Pharmaceutics 2019, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Quagliarini, E.; Di Santo, R.; Palchetti, S.; Ferri, G.; Cardarelli, F.; Pozzi, D.; Caracciolo, G. Effect of Protein Corona on The Transfection Efficiency of Lipid-Coated Graphene Oxide-Based Cell Transfection Reagents. Pharmaceutics 2020, 12, 113. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.P.; Huang, L. Overcoming the inhibitory effect of serum on lipofection by increasing the charge ratio of cationic liposome to DNA. Gene Ther. 1997, 4, 950–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, F.; Nishioka, T.; Saito, H.; Baba, T.; Okuda, A.; Matsumoto, O.; Taga, T.; Yamashita, F.; Takakura, Y.; Hashida, M. Interaction between DNA-cationic liposome complexes and erythrocytes is an important factor in systemic gene transfer via the intravenous route in mice: The role of the neutral helper lipid. Gene Ther. 2001, 8, 677–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fumoto, S.; Kawakami, S.; Shigeta, K.; Higuchi, Y.; Yamashita, F.; Hashida, M. Interaction with blood components plays a crucial role in asialoglycoprotein receptor-mediated in vivo gene transfer by galactosylated lipoplex. J. Pharmacol. Exp. Ther. 2005, 315, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, N.; Sakamoto, K.; Mizuno, S.; Sakaguchi, J.; Miyamoto, H.; Mine, T.; Sasaki, H.; Fumoto, S.; Nishida, K. Multiple components in serum contribute to hepatic transgene expression by lipoplex in mice. J. Gene Med. 2011, 13, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, N.; Fumoto, S.; Nakashima, M.; Shimokawa, K.; Miyamoto, H.; Nishida, K. The role of fibronectin in pulmonary gene transfer following intravenous administration of lipoplex in mice. Biol. Pharm. Bull. 2013, 36, 1807–1813. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, T.; Yoshida, S.; Kurosaki, T.; Teshima, M.; Nishida, K.; Nakamura, J.; Nakashima, M.; To, H.; Kitahara, T.; Sasaki, H. Cationic liposomes-mediated plasmid DNA delivery in murine hepatitis induced by carbon tetrachloride. J. Liposome Res. 2009, 19, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Smuckler, E.A.; Iseri, O.A.; Benditt, E.P. An intracellular defect in protein synthesis induced by carbon tetrachloride. J. Exp. Med. 1962, 116, 55–72. [Google Scholar] [CrossRef]
- Kawakami, S.; Fumoto, S.; Nishikawa, M.; Yamashita, F.; Hashida, M. In vivo gene delivery to the liver using novel galactosylated cationic liposomes. Pharm. Res. 2000, 17, 306–313. [Google Scholar] [CrossRef]
- Sakurai, F.; Nishioka, T.; Yamashita, F.; Takakura, Y.; Hashida, M. Effects of erythrocytes and serum proteins on lung accumulation of lipoplexes containing cholesterol or DOPE as a helper lipid in the single-pass rat lung perfusion system. Eur. J. Pharm. Biopharm. 2001, 52, 165–172. [Google Scholar] [CrossRef]
- Lausted, C.; Hu, Z.; Hood, L. Quantitative serum proteomics from surface plasmon resonance imaging. Mol. Cell. Proteom. 2008, 7, 2464–2474. [Google Scholar] [CrossRef] [Green Version]
- Fumoto, S.; Kawakami, S.; Ito, Y.; Shigeta, K.; Yamashita, F.; Hashida, M. Enhanced hepatocyte-selective in vivo gene expression by stabilized galactosylated liposome/plasmid DNA complex using sodium chloride for complex formation. Mol. Ther. 2004, 10, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Fumoto, S.; Furukawa, H.; Nakamura, J.; Nishida, K. Safety of liver surface instillation of plasmid DNA in normal and carbon tetrachloride-induced hepatitis mice. J. Pharm. Pharm. Sci. 2011, 14, 274–282. [Google Scholar] [CrossRef]
- Morimoto, K.; Kondo, M.; Kawahara, K.; Ushijima, H.; Tomino, Y.; Miyajima, M.; Kimura, J. Advances in targeting drug delivery to glomerular mesangial cells by long circulating cationic liposomes for the treatment of glomerulonephritis. Pharm. Res. 2007, 24, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Nagamura, Y.; Hirano, K. The recovering effect of betaine on carbon tetrachloride-induced liver injury. J. Nutr. Sci. Vitaminol. 1998, 44, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Un, K.; Kawakami, S.; Yoshida, M.; Higuchi, Y.; Suzuki, R.; Maruyama, K.; Yamashita, F.; Hashida, M. Efficient suppression of murine intracellular adhesion molecule-1 using ultrasound-responsive and mannose-modified lipoplexes inhibits acute hepatic inflammation. Hepatology 2012, 56, 259–269. [Google Scholar] [CrossRef]
- Janakat, S.; Al-Merie, H. Optimization of the dose and route of injection, and characterisation of the time course of carbon tetrachloride-induced hepatotoxicity in the rat. J. Pharmacol. Toxicol. Methods 2002, 48, 41–44. [Google Scholar] [CrossRef]
- Wong, F.W.; Chan, W.Y.; Lee, S.S. Resistance to carbon tetrachloride-induced hepatotoxicity in mice which lack CYP2E1 expression. Toxicol. Appl. Pharmacol. 1998, 153, 109–118. [Google Scholar] [CrossRef]
- DeCicco, L.A.; Rikans, L.E.; Tutor, C.G.; Hornbrook, K.R. Serum and liver concentrations of tumor necrosis factor α and interleukin-1β following administration of carbon tetrachloride to male rats. Toxicol. Lett. 1998, 98, 115–121. [Google Scholar] [CrossRef]
- Jain, A.; Barve, A.; Zhao, Z.; Fetse, J.P.; Liu, H.; Li, Y.; Cheng, K. Targeted Delivery of an siRNA/PNA Hybrid Nanocomplex Reverses Carbon Tetrachloride-Induced Liver Fibrosis. Adv. Ther. 2019, 2, 1900046. [Google Scholar] [CrossRef]
- Tousignant, J.D.; Gates, A.L.; Ingram, L.A.; Johnson, C.L.; Nietupski, J.B.; Cheng, S.H.; Eastman, S.J.; Scheule, R.K. Comprehensive analysis of the acute toxicities induced by systemic administration of cationic lipid:plasmid DNA complexes in mice. Hum. Gene Ther. 2000, 11, 2493–2513. [Google Scholar] [CrossRef]
- Gentry, M.J.; Snitily, M.U.; Preheim, L.C. Phagocytosis of Streptococcus pneumoniae measured in vitro and in vivo in a rat model of carbon tetrachloride-induced liver cirrhosis. J. Infect. Dis. 1995, 171, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Trewby, P.N.; Warren, R.; Contini, S.; Crosbie, W.A.; Wilkinson, S.P.; Laws, J.W.; Williams, R. Incidence and pathophysiology of pulmonary edema in fulminant hepatic failure. Gastroenterology 1978, 74, 859–865. [Google Scholar] [CrossRef]
- Andersson, S.E.; Lexmüller, K.; Johansson, A.; Ekström, G.M. Tissue and intracellular pH in normal periarticular soft tissue and during different phases of antigen induced arthritis in the rat. J. Rheumatol. 1999, 26, 2018–2024. [Google Scholar] [PubMed]
- O’Donnel, T.F., Jr.; Clowes, G.H., Jr.; Browse, N.L.; Ryan, N.T.; Blackburn, G.L. A metabolic approach to the evaluation of peripheral vascular disease. Surg. Gynecol. Obstet. 1977, 144, 51–57. [Google Scholar] [PubMed]
- Loisel, S.; Le Gall, C.; Doucet, L.; Ferec, C.; Floch, V. Contribution of plasmid DNA to hepatotoxicity after systemic administration of lipoplexes. Hum. Gene Ther. 2001, 12, 685–696. [Google Scholar] [CrossRef]
- Wunder, A.; Stehle, G.; Sinn, H.; Schrenk, H.; Hoffbiederbeck, D.; Bader, F.; Friedrich, E.; Peschke, P.; Maierborst, W.; Heene, D. Enhanced albumin uptake by rat tumors. Int. J. Oncol. 1997, 11, 497–507. [Google Scholar] [CrossRef]
- Meyer, K.A.; Kammerling, E.M.; Amtman, L.; Koller, M.; Hoffman, S.J. pH studies of malignant tissues in human beings. Cancer Res. 1948, 8, 513–518. [Google Scholar]
- Naeslund, J.; Swenson, K.E. Investigations on the pH of malignant tumors in mice and humans after the administration of glucose. Acta Obstet. Gynecol. Scand. 1953, 32, 359–367. [Google Scholar] [CrossRef]
- Chonn, A.; Cullis, P.R.; Devine, D.V. The role of surface charge in the activation of the classical and alternative pathways of complement by liposomes. J. Immunol. 1991, 146, 4234–4241. [Google Scholar]
- Devine, D.V.; Wong, K.; Serrano, K.; Chonn, A.; Cullis, P.R. Liposome-complement interactions in rat serum: Implications for liposome survival studies. Biochim. Biophys. Acta 1994, 1191, 43–51. [Google Scholar] [CrossRef]
- Noda, T.; Mimura, H.; Orita, K. Assessment of Kupffer cell function in rats with chronic liver injury caused by CCl4. Hepatogastroenterology 1990, 37, 319–323. [Google Scholar] [PubMed]
- Okazaki, M.; Furuya, E.; Kasahara, T.; Sakamoto, K. Function of reticuloendothelial system on CCl4 induced liver injury in mice. Jpn. J. Pharmacol. 1985, 39, 503–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luster, M.I.; Simeonova, P.P.; Gallucci, R.M.; Bruccoleri, A.; Blazka, M.E.; Yucesoy, B. Role of inflammation in chemical-induced hepatotoxicity. Toxicol. Lett. 2001, 120, 317–321. [Google Scholar] [CrossRef]
- Edwards, M.J.; Keller, B.J.; Kauffman, F.C.; Thurman, R.G. The involvement of Kupffer cells in carbon tetrachloride toxicity. Toxicol. Appl. Pharmacol. 1993, 119, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Czaja, M.J.; Xu, J.; Alt, E. Prevention of carbon tetrachloride-induced rat liver injury by soluble tumor necrosis factor receptor. Gastroenterology 1995, 108, 1849–1854. [Google Scholar] [CrossRef]
- Klement, J.F.; Rice, N.R.; Car, B.D.; Abbondanzo, S.J.; Powers, G.D.; Bhatt, P.H.; Chen, C.H.; Rosen, C.A.; Stewart, C.L. IκBα deficiency results in a sustained NF-κB response and severe widespread dermatitis in mice. Mol. Cell. Biol. 1996, 16, 2341–2349. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, Z.; Wang, L.; Zhang, X. NF-κB signaling pathway, inflammation and colorectal cancer. Cell. Mol. Immunol. 2009, 6, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Li, X.J.; Zeng, X.; Shen, D.Y.; Liu, C.Q.; Zhang, H.J.; Xu, C.B.; Li, X.Y. Activation of nuclear factor-κB pathway is responsible for tumor necrosis factor-α-induced up-regulation of endothelin B2 receptor expression in vascular smooth muscle cells in vitro. Toxicol. Lett. 2012, 209, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Kako, K.; Nishikawa, M.; Yoshida, H.; Takakura, Y. Effects of inflammatory response on in vivo transgene expression by plasmid DNA in mice. J. Pharm. Sci. 2008, 97, 3074–3083. [Google Scholar] [CrossRef]
- Gan, X.; Zhang, R.; Gu, J.; Ju, Z.; Wu, X.; Wang, Q.; Peng, H.; Qiu, J.; Zhou, J.; Cheng, F.; et al. Acidic Microenvironment Regulates the Severity of Hepatic Ischemia/Reperfusion Injury by Modulating the Generation and Function of Tregs via the PI3K-mTOR Pathway. Front. Immunol. 2020, 10, 2945. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipoplex | pH 7.4 | pH 7.0 | pH 6.5 | pH 6.0 | pH 5.5 |
|---|---|---|---|---|---|
| Control | 473 ± 24.5 | 491 ± 33.3 | 423 ± 20.2 | 437 ± 35.6 | 389 ± 21.9 * |
| BSA-interacted | 208 ± 5.63 | 207 ± 7.60 | 204 ± 7.50 | 200 ± 1.22 | 203 ± 5.03 |
| Lipoplex | pH 7.4 | pH 7.0 | pH 6.5 | pH 6.0 | pH 5.5 |
|---|---|---|---|---|---|
| Control | 39.7 ± 2.41 | 43.5 ± 1.25 * | 44.9 ± 0.17 ** | 47.6 ± 1.65 ** | 48.3 ± 1.04 ** |
| BSA-interacted | −11.7 ± 0.76 | −10.5 ± 0.85 | −9.50 ± 0.35 * | −8.21 ± 0.82 ** | −7.36 ± 0.66 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshikawa, N.; Fumoto, S.; Yoshikawa, K.; Hu, D.; Okami, K.; Kato, R.; Nakashima, M.; Miyamoto, H.; Nishida, K. Interaction of Lipoplex with Albumin Enhances Gene Expression in Hepatitis Mice. Pharmaceutics 2020, 12, 341. https://doi.org/10.3390/pharmaceutics12040341
Yoshikawa N, Fumoto S, Yoshikawa K, Hu D, Okami K, Kato R, Nakashima M, Miyamoto H, Nishida K. Interaction of Lipoplex with Albumin Enhances Gene Expression in Hepatitis Mice. Pharmaceutics. 2020; 12(4):341. https://doi.org/10.3390/pharmaceutics12040341
Chicago/Turabian StyleYoshikawa, Naoki, Shintaro Fumoto, Keiko Yoshikawa, Die Hu, Kazuya Okami, Riku Kato, Mikiro Nakashima, Hirotaka Miyamoto, and Koyo Nishida. 2020. "Interaction of Lipoplex with Albumin Enhances Gene Expression in Hepatitis Mice" Pharmaceutics 12, no. 4: 341. https://doi.org/10.3390/pharmaceutics12040341