Enhanced Antioxidant and Cytotoxic Potentials of Lipopolysaccharides-Injected Musca domestica Larvae

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Oxidative Stress Induction
2.3. Hemolymph Collection
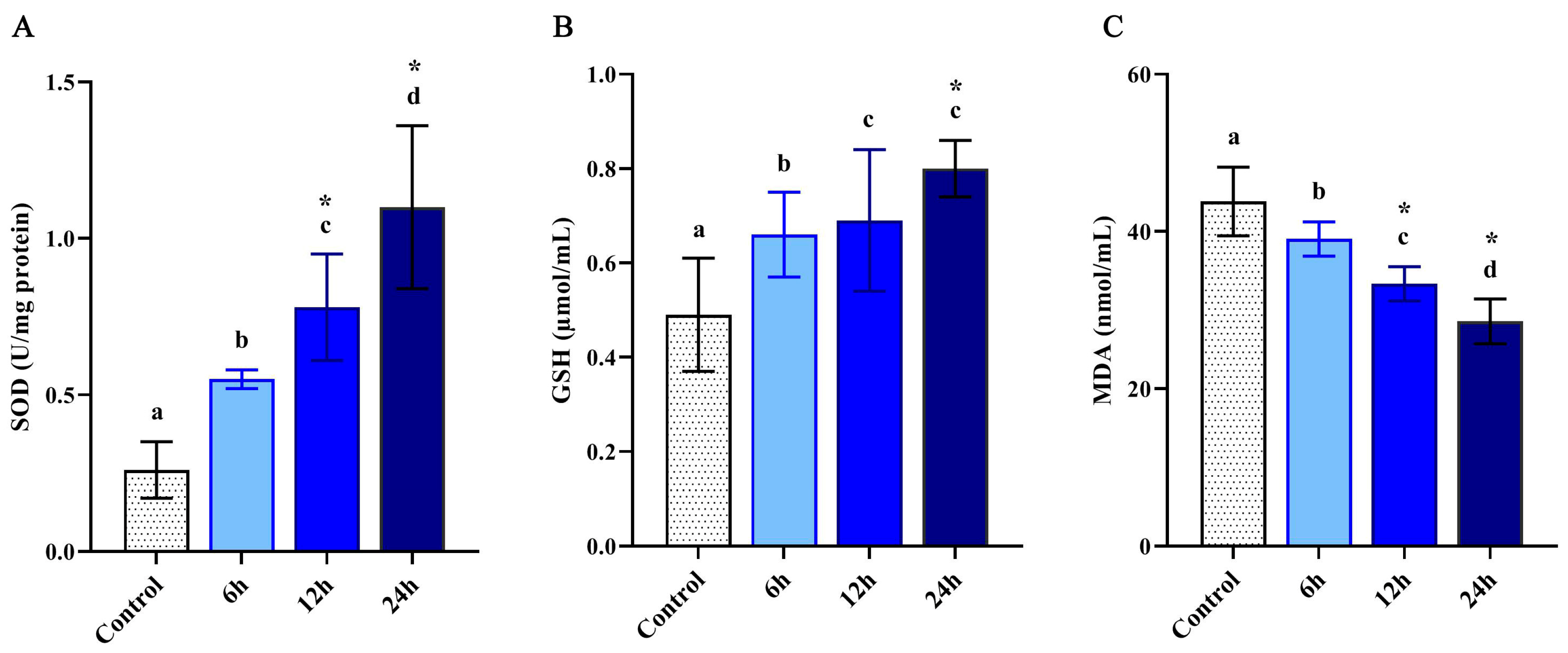
2.4. Determination of Antioxidant Enzymes Activity
2.4.1. Superoxide Dismutase (SOD)
2.4.2. Glutathione (GSH)
2.4.3. Malondialdehyde (MDA)
2.5. Protein Electrophoresis (SDS-PAGE)
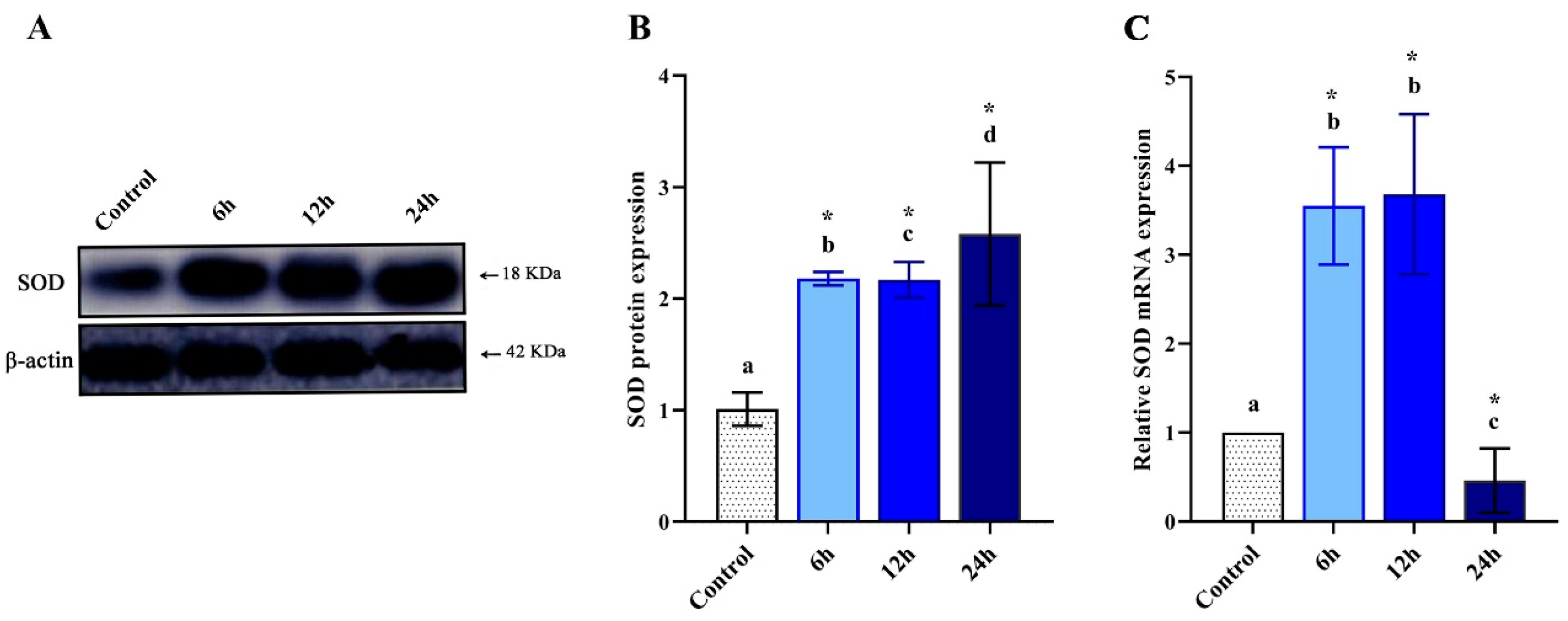
2.6. Western Blot Analysis
2.7. Quantitative Real-Time PCR (qPCR) for the MdSOD1 Gene
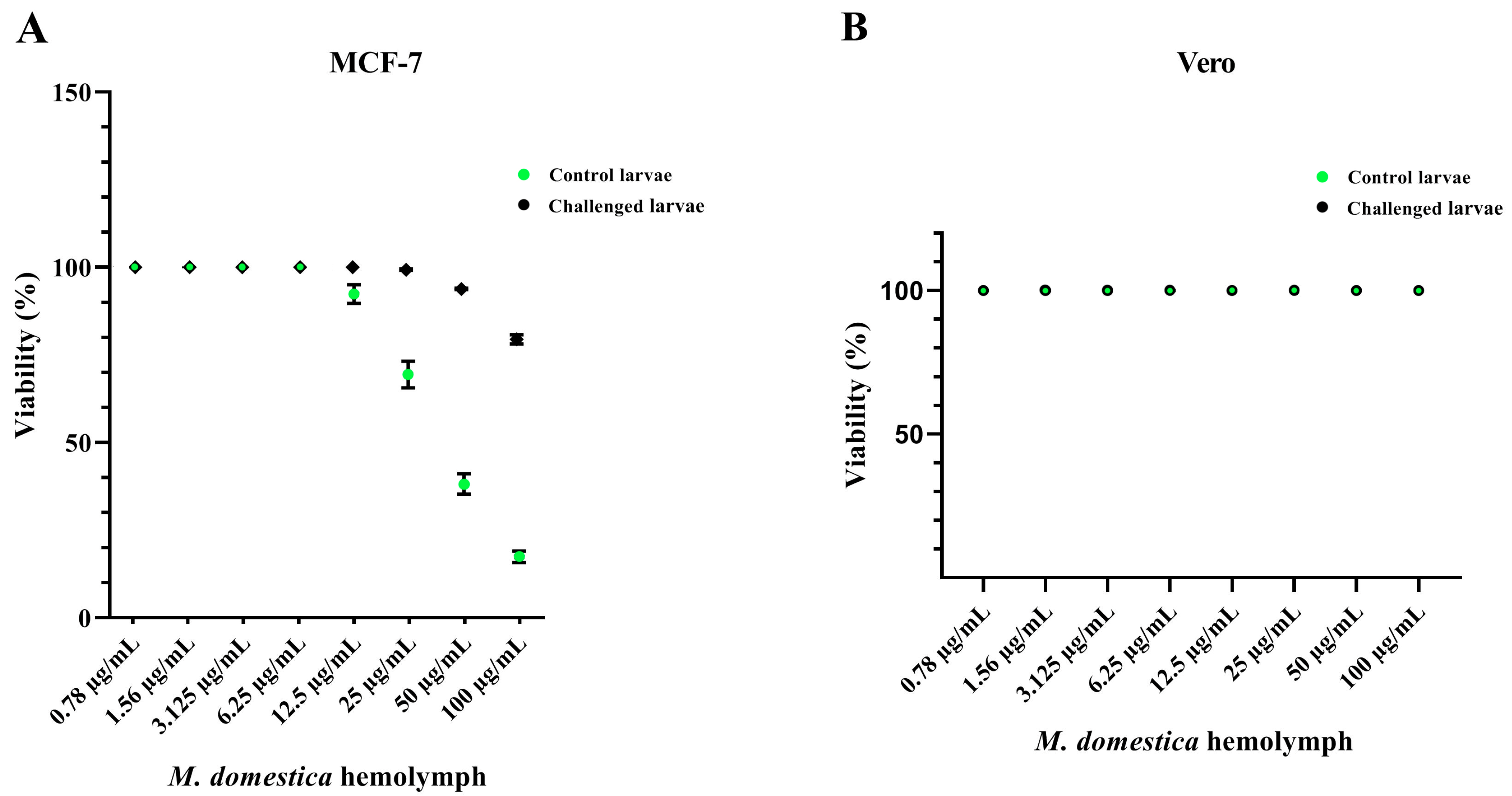
2.8. In Vitro Anticancer Activities
2.8.1. Maintenance of Cell Lines
2.8.2. Cytotoxicity Assay Using (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide Dye (MTT)
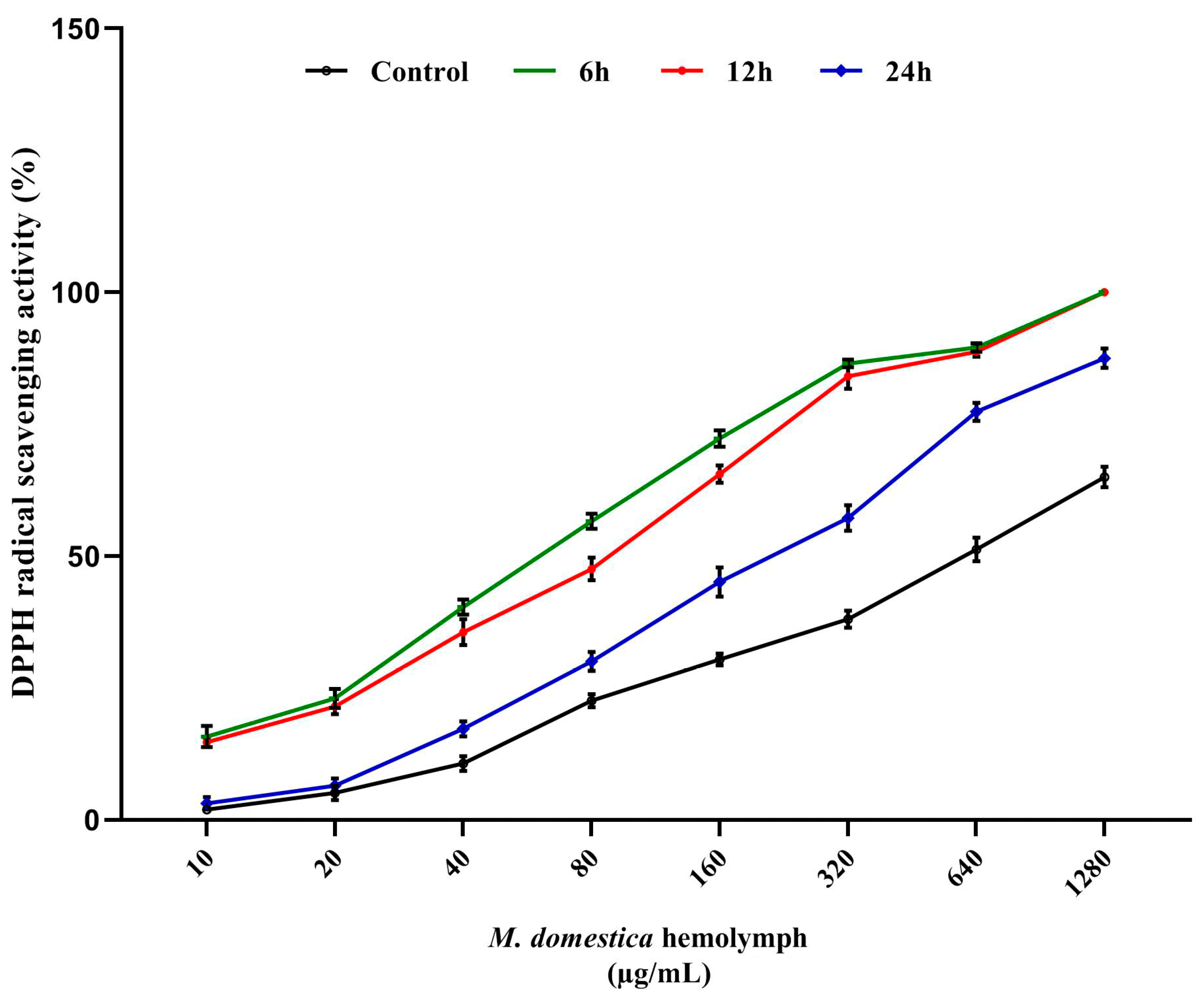
2.9. Determination of Antioxidant Activity In Vitro
2.9.1. DPPH Assay
2.9.2. Total Antioxidant Capacity (TAC)
2.10. Data Analysis
3. Results
3.1. Antioxidant Enzymes Activity
3.2. Altered Levels of MdSOD1 Protein
3.3. Altered Expression of MdSOD1 mRNA
3.4. Cytotoxicity of the M. domestica Larval Hemolymph
3.5. DPPH Radical Scavenging Assay
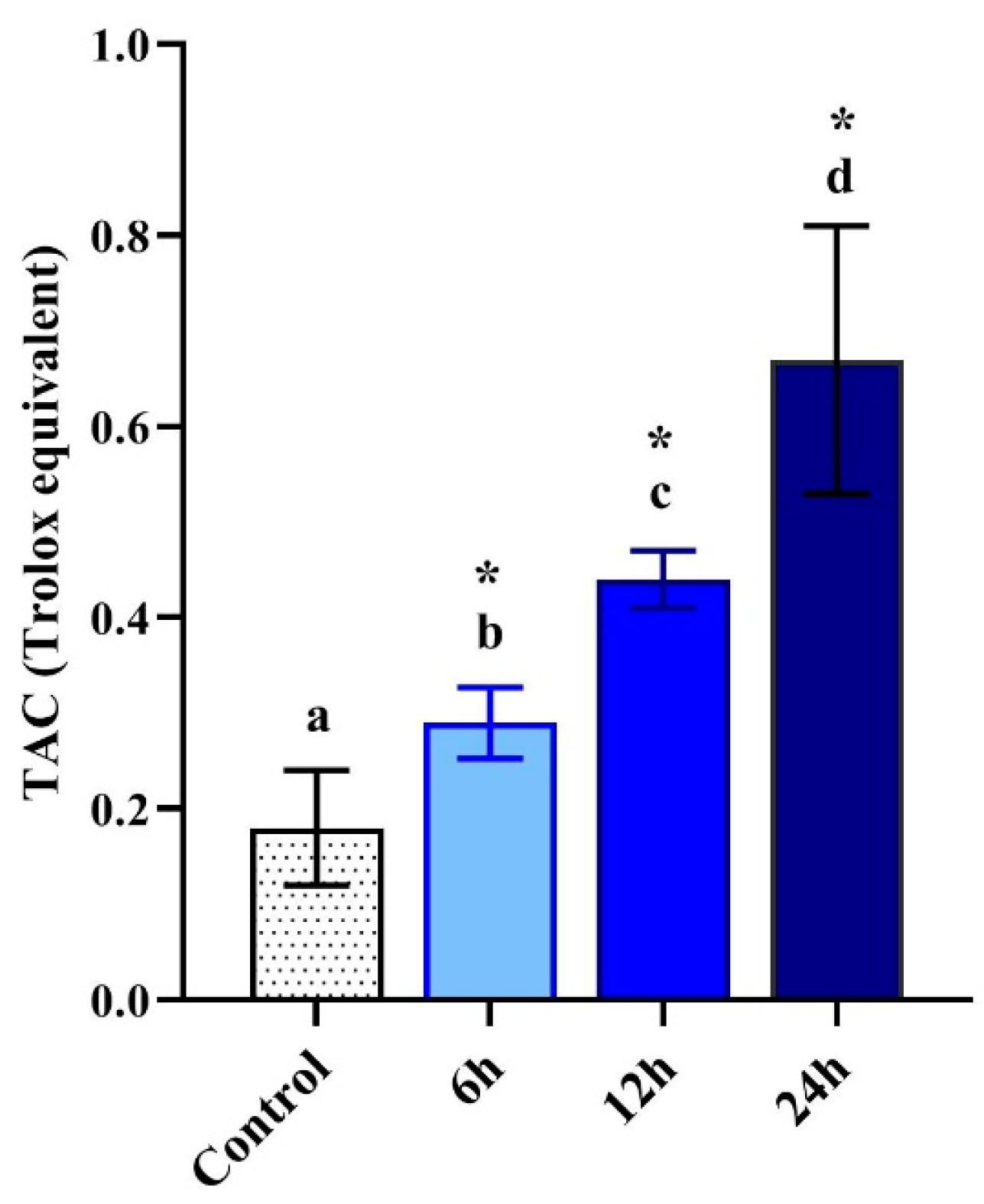
3.6. Total Antioxidant Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- El-Garawani, I.; Hassab, S.; Nabi, E.; El-Ghandour, E. The Protective Effect of (Foeniculum Vulgare) Oil on Etoposide-Induced Genotoxicity on Male Albino Rats. Eur. J. Pharm. Med. Res. 2017, 4, 180–194. [Google Scholar]
- Saeed, M.; Amen, A.; Fahmi, A.; Garawani, I.E.; Sayed, S. The Possible Protective Effect of Coriandrum Sativum Seeds Methanolic Extract on Hepato-Renal Toxicity Induced by Sodium Arsenite in Albino Rats. J. Appl. Pharm. Sci. 2014, 4. [Google Scholar] [CrossRef] [Green Version]
- El-Garawani, I.; Emam, M.; Elkhateeb, W.; El-Seedi, H.; Khalifa, S.; Oshiba, S.; Abou-Ghanima, S.; Daba, G. In Vitro Antigenotoxic, Antihelminthic and Antioxidant Potentials Based on the Extracted Metabolites from Lichen, Candelariella Vitellina. Pharmaceutics 2020, 12, 477. [Google Scholar] [CrossRef] [PubMed]
- El-Garawani, I.M.; El-Nabi, S.E.H.; Mohamed, A.H.; El-Esawy, H.M. Molecular Amelioration of Acacia Arabica Gum on Some Male Reproductive Aspects in Schistosoma Mansoni Infected Mice. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 505–512. [Google Scholar]
- Ahmed, A.; Ali, M.; El-Kholie, E.; El-Garawani, I.; Sherif, N. Anticancer Activity of Morus Nigra on Human Breast Cancer Cell Line (MCF-7): The Role of Fresh and Dry Fruit Extracts. J. Biosci. Appl. Res. 2016, 2, 352–361. [Google Scholar]
- Tohamy, A.A.; El-Garawani, I.M.; Ibrahim, S.R.; Abdel Moneim, A.E. The Apoptotic Properties of Salvia Aegyptiaca and Trigonella Foenum-Graecum Extracts on Ehrlich Ascites Carcinoma Cells: The Effectiveness of Combined Treatment. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 1842–1883. [Google Scholar]
- El-Garawani, I.; El Nabi, S.H.; Nafie, E.; Almeldin, S. Foeniculum Vulgare and Pelargonium Graveolens Essential Oil Mixture Triggers the Cell Cycle Arrest and Apoptosis in MCF-7 Cells. Anticancer Agents Med. Chem. 2019, 19, 1013–1113. [Google Scholar] [CrossRef]
- El-Garawani, I.M.; El-Nabi, S.H.; El-Shafey, S.; Elfiky, M.; Nafie, E. Coffea Arabica Bean Extracts and Vitamin C: A Novel Combination Unleashes MCF-7 Cell Death. Curr. Pharm. Biotechnol. 2020, 21, 23–36. [Google Scholar] [CrossRef]
- El-Garawani, I.M.; El-Nabi, S.H.; Dawoud, G.T.; Esmail, S.M.; Abdel Moneim, A.E. Triggering of Apoptosis and Cell Cycle Arrest by Fennel and Clove Oils in Caco-2 Cells: The Role of Combination. Toxicol. Mech. Methods 2019, 29, 710–722. [Google Scholar] [CrossRef]
- Pettit, G.R.; Meng, Y.; Herald, D.L.; Knight, J.C.; Day, J.F. Antineoplastic Agents. 553. The Texas Grasshopper Brachystola Magna. J. Nat. Prod. 2005, 68, 1256–1258. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-E.; Jo, D.-E.; Lee, A.-J.; Park, H.-K.; Youn, K.; Yun, E.-Y.; Hwang, J.-S.; Jun, M.; Kang, B.H. Hepatoprotective and Anticancer Activities of Allomyrina Dichotoma Larvae. J. Life Sci. 2015. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.M.; Ku, M.J.; Son, Y.-J.; Yun, J.-M.; Kim, S.H.; Lee, S.Y. Anti-Metastatic Effect of Cantharidin in A549 Human Lung Cancer Cells. Arch. Pharm. Res. 2013, 36, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Arumugam Sarasa, B.A. Effect of Crude Extract of Bombyx Mori Coccoons in Hyperlipidemia and Atherosclerosis. J. Ayurveda Integr. Med. 2011, 2, 72–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.L.; Wang, R.; Feng, X.; Zhao, X. Preventive Effect of Insect Tea against Reserpine-Induced Gastric Ulcers in Mice. Exp. Ther. Med. 2014, 8, 1318–1324. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S.; Singh, M.; Khan, M.A.; Arya, D.S.; Ahmad, S. Scientific Validation of Cardioprotective Attribute by Standardized Extract of Bombyx Mori against Doxorubicin-Induced Cardiotoxicity in Murine Model. Excli J. 2014, 13, 1043–1054. [Google Scholar] [CrossRef]
- Prakash, S.; Bhargava, H.R. Apis Cerana Bee Venom: It’s Anti-Diabetic and Anti-Dandruff Activity against Malassezia Furfur. World Appl. Sci. J. 2014, 32, 343–348. [Google Scholar] [CrossRef]
- Józefiak, A.; Engberg, R.M. Insect Proteins as a Potential Source of Antimicrobial Peptides in Livestock Production. A Review. J. Anim. Feed Sci. 2017, 26, 87–99. [Google Scholar] [CrossRef]
- Wu, Q.; Patočka, J.; Kuča, K. Insect Antimicrobial Peptides, a Mini Review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef]
- Tang, J.J.; Fang, P.; Xia, H.L.; Tu, Z.C.; Hou, B.Y.; Yan, Y.M.; Di, L.; Zhang, L.; Cheng, Y.X. Constituents from the Edible Chinese Black Ants (Polyrhachis Dives) Showing Protective Effect on Rat Mesangial Cells and Anti-Inflammatory Activity. Food Res. Int. 2015, 67, 163–168. [Google Scholar] [CrossRef]
- Yan, Y.M.; Li, L.J.; Qin, X.C.; Lu, Q.; Tu, Z.C.; Cheng, Y.X. Compounds from the Insect Blaps Japanensis with COX-1 and COX-2 Inhibitory Activities. Bioorganic Med. Chem. Lett. 2015, 25, 2469–2672. [Google Scholar] [CrossRef]
- Tiveron, A.P.; Rosalen, P.L.; Franchin, M.; Lacerda, R.C.C.; Bueno-Silva, B.; Benso, B.; Denny, C.; Ikegaki, M.; De Alencar, S.M. Chemical Characterization and Antioxidant, Antimicrobial, and Anti-Inflammatory Activities of South Brazilian Organic Propolis. PLoS ONE 2016, 11, e0165588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shittu, O.K.; Bashir, L.; Isaac, O.O. Effects of Methanol Extract of Musca Domestica Larvae on Antioxidants Enzymes in T. Brucei Infected Rats. Niger. J. Biochem. Mol. Biol. 2014, 29, 1–10. [Google Scholar]
- Nmorsi, O.P.G.; Ukwandu, N.C.D.; Agbozele, G.E. Detection of Some Gastrointestinal Parasites from Four Synanthropic Flies in Ekpoma, Nigeria. J. Vector Borne Dis. 2006, 43, 136–139. [Google Scholar] [PubMed]
- An, C.; Li, D.; Du, R. Analysis of Antibacterial-Relative Proteins and Peptides in Housefly Larvae. Wei Sheng Yan Jiu 2004, 33, 86–88. [Google Scholar] [PubMed]
- Guo, G.; Tao, R.; Li, Y.; Ma, H.; Xiu, J.; Fu, P.; Wu, J. Identification and Characterization of a Novel Antimicrobial Protein from the Housefly Musca Domestica. Biochem. Biophys. Res. Commun. 2017, 490, 746–752. [Google Scholar] [CrossRef]
- Ok, S.; Olayemi, I.K.; Omalu, I.C.; Adeniyi, A.K. Anti-Plasmodial properties of methanolic extract of musca domestica maggot on P. berghei—Infected mice. Int. J. Biol. Pharm. Allied Sci. 2013, 2, 1064–1070. [Google Scholar]
- Sun, H.X.; Chen, L.Q.; Zhang, J.; Chen, F.Y. Anti-Tumor and Immunomodulatory Activity of Peptide Fraction from the Larvae of Musca Domestica. J. Ethnopharmacol. 2014, 153, 831–839. [Google Scholar] [CrossRef]
- Zhao, R.J.; Zhang, Q.H.; Li, F.D. The Effection of Antimicrobial Peptides Extracted from Adult Housefly on Tumour C. Chin. J. Vector Biol. Control 2007, 18, 17. [Google Scholar]
- Hou, L.; Shi, Y.; Zhai, P.; Le, G. Antibacterial Activity and in Vitro Anti-Tumor Activity of the Extract of the Larvae of the Housefly (Musca Domestica). J. Ethnopharmacol. 2007, 42, 259–265. [Google Scholar] [CrossRef]
- Boman, H.G. Peptide Antibiotics and Their Role in Innate Immunity. Annu. Rev. Immunol. 1995, 13, 61–92. [Google Scholar] [CrossRef]
- Bulet, P.; Hetru, C.; Dimarcq, J.L.; Hoffmann, D. Antimicrobial Peptides in Insects; Structure and Function. Dev. Comp. Immunol. 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Barbehenn, R.V. Gut-Based Antioxidant Enzymes in a Polyphagous and a Graminivorous Grasshopper. J. Chem. Ecol. 2002, 28, 1329–1347. [Google Scholar] [CrossRef] [PubMed]
- Felton, G.W.; Summers, C.B. Antioxidant Systems in Insects. Arch. Insect Biochem. Physiol. 1995, 29, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, E.; Baraniak, B.; Karaś, M.; Rybczyńska, K.; Jakubczyk, A. Selected Species of Edible Insects as a Source of Nutrient Composition. Food Res. Int. 2015, 77, 460–466. [Google Scholar] [CrossRef]
- Zielińska, E.; Karaś, M.; Jakubczyk, A. Antioxidant Activity of Predigested Protein Obtained from a Range of Farmed Edible Insects. Int. J. Food Sci. Technol. 2017, 52, 306–312. [Google Scholar] [CrossRef]
- Gobbetti, M.; Stepaniak, L.; De Angelis, M.; Corsetti, A.; Di Cagno, R. Latent Bioactive Peptides in Milk Proteins: Proteolytic Activation and Significance in Dairy Processing. Crit. Rev. Food Sci. Nutr. 2002, 42, 223–239. [Google Scholar] [CrossRef]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant Activity of Peptides Obtained from Porcine Myofibrillar Proteins by Protease Treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef]
- Zhang, M.; Mu, T.H.; Wang, Y.B.; Sun, M.J. Evaluation of Free Radical-Scavenging Activities of Sweet Potato Protein and Its Hydrolysates as Affected by Single and Combination of Enzyme Systems. Int. J. Food Sci. Technol. 2012, 47, 696–702. [Google Scholar] [CrossRef]
- Karaś, M.; Jakubczyk, A.; Szymanowska, U.; Złotek, U.; Zielińska, E. Digestion and Bioavailability of Bioactive Phytochemicals. Int. J. Food Sci. Technol. 2017, 52, 291–305. [Google Scholar] [CrossRef]
- Di Mattia, C.; Battista, N.; Sacchetti, G.; Serafini, M. Antioxidant Activities in Vitro of Water and Liposoluble Extracts Obtained by Different Species of Edible Insects and Invertebrates. Front. Nutr. 2019, 6, 106. [Google Scholar] [CrossRef] [Green Version]
- Dutta, P.; Dey, T.; Manna, P.; Kalita, J. Antioxidant Potential of Vespa affinis L., a Traditional Edible Insect Species of North East India. PLoS ONE 2016, 11, e0156107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoud, S.; Moselhy, W.; El-Khashab, L.A.; Seufi, A.M. Identification and Molecular Characterisation of a Novel Manganese Superoxide Dismutase Gene from Flesh Fly Larvae, Sarcophaga Argyrostoma (Diptera: Sarcophagidae). Afr. Entomol. 2018, 26, 448–457. [Google Scholar] [CrossRef]
- Tang, T.; Huang, D.-W.; Zhou, C.-Q.; Li, X.; Xie, Q.-J.; Liu, F.-S. Molecular Cloning and Expression Patterns of Copper/Zinc Superoxide Dismutase and Manganese Superoxide Dismutase in Musca Domestica. Gene 2012, 505, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Večeřa, J.; Krishnan, N.; Alquicer, G.; Kodrík, D.; Socha, R. Adipokinetic Hormone-Induced Enhancement of Antioxidant Capacity of Pyrrhocoris Apterus Hemolymph in Response to Oxidative Stress. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wu, Y.; Wang, Z.; Chen, J.; Yang, Y.; Dong, G. Dandelion Extract Alleviated Lipopolysaccharide-Induced Oxidative Stress through the Nrf2 Pathway in Bovine Mammary Epithelial Cells. Toxins (Basel) 2020, 12, 496. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, H.H.; Nie, X.T.; Jiang, W.R.; Zhang, Y.S. Sodium Butyrate Ameliorates Lipopolysaccharide-Induced Cow Mammary Epithelial Cells from Oxidative Stress Damage and Apoptosis. J. Cell. Biochem. 2019, 120, 2370–2381. [Google Scholar] [CrossRef] [PubMed]
- Magdaleno, F.; Blajszczak, C.C.; Nieto, N. Key Events Participating in the Pathogenesis of Alcoholic Liver Disease. Biomolecules 2017, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Gomes, J.M.G.; de Assis Costa, J.; de Cássia Gonçalves Alfenas, R. Metabolic Endotoxemia and Diabetes Mellitus: A Systematic Review. Metab. Clin. Exp. 2017, 68, 133–144. [Google Scholar] [CrossRef]
- Choi, Y.S.; Lee, K.S.; Yoon, H.J.; Kim, I.; Sohn, H.D.; Jin, B.R. Bombus Ignitus Cu,Zn Superoxide Dismutase (SOD1): CDNA Cloning, Gene Structure, and up-Regulation in Response to Paraquat, Temperature Stress, or Lipopolysaccharide Stimulation. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 144, 365–371. [Google Scholar] [CrossRef]
- Parusel, R.; Steimle, A.; Lange, A.; Schäfer, A.; Maerz, J.K.; Bender, A.; Frick, J.S. An Important Question: Which LPS Do You Use? Virulence 2017, 8, 1890–1893. [Google Scholar] [CrossRef] [Green Version]
- Homa, J.; Stalmach, M.; Wilczek, G.; Kolaczkowska, E. Effective Activation of Antioxidant System by Immune-Relevant Factors Reversely Correlates with Apoptosis of Eisenia Andrei Coelomocytes. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2016, 186, 417–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, F.J.; Jin, X.B.; Xu, Y.Y.; Ma, Y.; Li, X.B.; Lu, X.M.; Liu, W.B.; Zhu, J.Y. Inflammatory Regulation Effect and Action Mechanism of Anti-Inflammatory Effective Parts of Housefly (Musca Domestica) Larvae on Atherosclerosis. Evid. Based Complement. Altern. Med. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Yi, Y.; Lv, Y.; Li, M.; Wang, J.; Qiu, L. The Lipopolysaccharide (LPS) of Photorhabdus Luminescens TT01 Can Elicit Dose- and Time-Dependent Immune Priming in Galleria Mellonella Larvae. J. Invertebr. Pathol. 2015, 127, 63–72. [Google Scholar] [CrossRef]
- Stadtman, E.R. Protein Oxidation and Aging. Free Radic. Res. 2006, 40, 1250–1258. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.S.; Kasoju, N.; Luthra, A.; Singh, A.; Sharanabasava, H.; Sahu, A.; Bora, U. Indian Medicinal Herbs as Sources of Antioxidants. Food Res. Int. 2008, 41, 1–15. [Google Scholar] [CrossRef]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; LLeonart, M.E. Oxidative Stress and Cancer: An Overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef]
- Baynes, J.W. Role of Oxidative Stress in Development of Complications in Diabetes. Diabetes 1991, 40, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Torres-Fuentes, C.; Alaiz, M.; Vioque, J. Affinity Purification and Characterisation of Chelating Peptides from Chickpea Protein Hydrolysates. Food Chem. 2011, 129, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Karaś, M.; Baraniak, B.; Rybczyńska, K.; Gmiński, J.; Gaweł-Bęben, K.; Jakubczyk, A. The Influence of Heat Treatment of Chickpea Seeds on Antioxidant and Fibroblast Growth-Stimulating Activity of Peptide Fractions Obtained from Proteins Digested under Simulated Gastrointestinal Conditions. Int. J. Food Sci. Technol. 2015, 50, 2097–2103. [Google Scholar] [CrossRef]
- Carrasco-Castilla, J.; Hernández-Álvarez, A.J.; Jiménez-Martínez, C.; Jacinto-Hernández, C.; Alaiz, M.; Girón-Calle, J.; Vioque, J.; Dávila-Ortiz, G. Antioxidant and Metal Chelating Activities of Phaseolus Vulgaris L. Var. Jamapa Protein Isolates, Phaseolin and Lectin Hydrolysates. Food Chem. 2012, 131, 1157–1164. [Google Scholar] [CrossRef]
- Tak, P.P.; Zvaifler, N.J.; Firestein, G.S.; Greene, D.R. Rheumatoid Arthritis and P53: How Oxidative Stress, Might Alter the Course of Inflammatory Diseases. Immunol. Today 2000, 21, 78–82. [Google Scholar] [CrossRef]
- Manna, P.; Bhattacharyya, S.; Das, J.; Ghosh, J.; Sil, P.C. Phytomedicinal Role of Pithecellobium Dulce against Ccl4 -Mediated Hepatic Oxidative Impairments and Necrotic Cell Death. Evid. Based Complement. Altern. Med. 2011, 2011, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O‘Brien’, P.J. Antioxidants and Cancer: Molecular Mechanism. In Free Radicals in Diagnostic Medicine; Springer: Boston, MA, USA, 1994. [Google Scholar]
- Hashem, H.O.; Youssef, N.S. Developmental Changes Induced by Methanolic Extracts of Leaves and Fruits of Melia Azedarach L. on the House Fly Musca Domestica Vicina MACQ. J. Egypt. Ger. Soc. Zool. 1991, 3, 335–352. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the Superoxide Anion Radical in the Autoxidation of Pyrogallol and a Convenient Assay for Superoxide Dismutase. Eur. J. Biochem. 1974, 17, 310. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L. Tissue Sulfhydryl Groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Mihara, M.; Uchiyama, M. Determination of Malonaldehyde Precursor in Tissues by Thiobarbituric Acid Test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 13, 519–523. [Google Scholar] [CrossRef]
- Burnette, W.N. “Western Blotting”: Electrophoretic Transfer of Proteins from Sodium Dodecyl Sulfate-Polyacrylamide Gels to Unmodified Nitrocellulose and Radiographic Detection with Antibody and Radioiodinated Protein A. Anal. Biochem. 1981, 112, 195–203. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Zhu, Z.; Ma, W.; Lei, C. The Molecular Characterization of Antioxidant Enzyme Genes in Helicoverpa Armigera Adults and Their Involvement in Response to Ultraviolet-A Stress. J. Insect Physiol. 2012, 58, 1250–1258. [Google Scholar] [CrossRef]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative Properties of Xanthone on the Auto Oxidation of Soybean in Cylcodextrin Emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Meng, J.Y.; Zhang, C.Y.; Zhu, F.; Wang, X.P.; Lei, C.L. Ultraviolet Light-Induced Oxidative Stress: Effects on Antioxidant Response of Helicoverpa Armigera Adults. J. Insect Physiol. 2009, 55, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Jovanović-Galović, A.; Blagojević, D.P.; Grubor-Lajšić, G.; Worland, R.; Spasić, M.B. Role of Antioxidant Defense during Different Stages of Preadult Life Cycle in European Corn Borer (Ostrinia Nubilalis, Hubn.): Diapause and Metamorphosis. Arch. Insect Biochem. Physiol. 2004, 55, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Pritsos, C.A.; Ahmad, S.; Elliott, A.J.; Pardini, R.S. Antioxidant Enzyme Level Response to Prooxidant Allelochemicals in Larvae of the Southern Armyworm Moth, Spodoptera Eridania. Free Radic. Res. 1990, 9, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Oberley, L.W.; Murhammer, D.W. Antioxidant Defense Systems of Two Lipidopteran Insect Cell Lines. Free Radic. Biol. Med. 2001, 30, 1254–1262. [Google Scholar] [CrossRef]
- Weirich, G.F.; Collins, A.M.; Williams, V.P. Antioxidant Enzymes in the Honey Bee, Apis Mellifera. Apidologie 2002, 33, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.G.; Mitchell, A.E.; Shibamoto, T. Determination of Antioxidant Properties of Aroma Extracts from Various Beans. J. Agric. Food Chem. 2000, 48, 4817–4820. [Google Scholar] [CrossRef]
- Dreher, D.; Junod, A.F. Role of Oxygen Free Radicals in Cancer Development. Eur. J. Cancer 1996, 32, 30–38. [Google Scholar] [CrossRef]
- Wyatt, G.R. The Biochemistry of Insect Hemolymph. Annu. Rev. Entomol. 1961, 6, 75–102. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The Host Defense of Drosophila Melanogaster. Annu. Rev. Immunol. 2007, 120, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Suh, H.J.; Kim, S.R.; Lee, K.S.; Park, S.; Kang, S.C. Antioxidant Activity of Various Solvent Extracts from Allomyrina Dichotoma (Arthropoda: Insecta) Larvae. J. Photochem. Photobiol. B Biol. 2010, 99, 67–73. [Google Scholar] [CrossRef]
- Pentreath, V.W.; Kennedy, P.G.E. Pathogenesis of Human African Trypanosomiasis. In The trypanosomiases; CABI Publishing: Wallingford, CT, USA, 2009. [Google Scholar] [CrossRef]
- Atalay, M.; Laaksonen, D.E.; Khanna, S.; Kaliste-Korhonen, E.; Hänninen, O.; Sen, C.K. Vitamin E Regulates Changes in Tissue Antioxidants Induced by Fish Oil and Acute Exercise. Med. Sci. Sports Exerc. 2000, 32, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, C.; Hassan, H.M. Induction and Inactivation of Catalase and Superoxide Dismutase of Escherichia Coli by Ozone. Arch. Biochem. Biophys. 1987, 257, 464–471. [Google Scholar] [CrossRef]
- Farr, S.B.; Kogoma, T. Oxidative Stress Responses in Escherichia Coli and Salmonella Typhimurium. Microbiol. Rev. 1991, 55, 561–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Liu, X.S.; Zhang, Q.; Zhao, H.B.; Wang, Y.F. Expression Profiles of Six Novel C-Type Lectins in Response to Bacterial and 20E Injection in the Cotton Bollworm (Helicoverpa Armigera). Dev. Comp. Immunol. 2012, 37, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Večeřa, J.; Krishnan, N.; Mithöfer, A.; Vogel, H.; Kodrík, D. Adipokinetic Hormone-Induced Antioxidant Response in Spodoptera Littoralis. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 389–395. [Google Scholar] [CrossRef]
- Wu, G.; Nie, L.; Zhang, W. Integrative Analyses of Posttranscriptional Regulation in the Yeast Saccharomyces Cerevisiae Using Transcriptomic and Proteomic Data. Curr. Microbiol. 2008, 57, 18–22. [Google Scholar] [CrossRef]
- Maier, T.; Güell, M.; Serrano, L. Correlation of MRNA and Protein in Complex Biological Samples. Febs Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, J.A.; Reichhart, J.M.; Hetru, C. Innate Immunity in Higher Insects. Curr. Opin. Immunol. 1996, 8, 8–13. [Google Scholar] [CrossRef]
- Grizanova, E.V.; Dubovskiy, I.M.; Whitten, M.M.A.; Glupov, V.V. Contributions of Cellular and Humoral Immunity of Galleria Mellonella Larvae in Defence against Oral Infection by Bacillus Thuringiensis. J. Invertebr. Pathol. 2014, 119, 40–46. [Google Scholar] [CrossRef]
- Wang, F.; Ai, H.; Lei, C. In Vitro Anti-Influenza Activity of a Protein-Enriched Fraction from Larvae of the Housefly (Musca Domestica). Pharm. Biol. 2013, 51, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Moore, A.J.; Devine, D.A.; Bibby, M.C. Preliminary Experimental Anticancer Activity of Cecropins. Pept. Res. 1994, 7, 265–269. [Google Scholar] [PubMed]
- Mader, J.S.; Hoskin, D.W. Cationic Antimicrobial Peptides as Novel Cytotoxic Agents for Cancer Treatment. Expert Opin. Investig. Drugs 2006, 15, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, D.W.; Ramamoorthy, A. Studies on Anticancer Activities of Antimicrobial Peptides. Biochim. Biophys. Acta Biomemb. 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berge, G.; Eliassen, L.T.; Camilio, K.A.; Bartnes, K.; Sveinbjørnsson, B.; Rekdal, Ø. Therapeutic Vaccination against a Murine Lymphoma by Intratumoral Injection of a Cationic Anticancer Peptide. Cancer Immunol. Immunother. 2010, 59, 1285–1294. [Google Scholar] [CrossRef]
- Tsai, J.C.; Jain, M.; Hsieh, C.M.; Lee, W.S.; Yoshizumi, M.; Patterson, C.; Perrella, M.A.; Cooke, C.; Wang, H.; Haber, E.; et al. Induction of Apoptosis by Pyrrolidinedithiocarbamate and N-Acetylcysteine in Vascular Smooth Muscle Cells. J. Biol. Chem. 1996, 271, 3667–3670. [Google Scholar] [CrossRef] [Green Version]
- Hasaballah, A.; Shehata, A.; Shehab, A. Antioxidant and Anticancer Activities of Some Maggots Methanol Extracts. Egypt. Acad. J. Biol. Sci. Aentomol. 2019, 12, 111–119. [Google Scholar] [CrossRef]
- Shehata, A.; Mehany, A.; El-Sheikh, T. Excretion/Secretion of Lucilia Sericata and Chrysomya Albiceps (Diptera: Calliphoridae) Maggots as Potential Anticancer Agent and Kinases Inhibitor. N. Y. Sci. J. 2016. [Google Scholar] [CrossRef]
- Song, C.; Yu, H.; Zhang, M.; Yang, Y.; Zhang, G. Physicochemical Properties and Antioxidant Activity of Chitosan from the Blowfly Chrysomya Megacephala Larvae. Int. J. Biol. Macromol. 2013, 60, 347–354. [Google Scholar] [CrossRef]
- Suh, H.J.; Kim, S.R.; Hwang, J.S.; Kim, M.J.; Kim, I. Antioxidant Activity of Aqueous Methanol Extracts from the Lucanid Beetle, Serrognathus Platymelus Castanicolor Motschulsky (Coleoptera: Lucanidae). J. Asia Pac. Entomol. 2011, 14, 95–98. [Google Scholar] [CrossRef]
- Suh, H.-J.; Kang, S.C. Antioxidant Activity of Aqueous Methanol Extracts of Protaetia Brevitarsis Lewis (Coleoptera: Scarabaedia) at Different Growth Stages. Nat. Prod. Res. 2012, 26, 510–517. [Google Scholar] [CrossRef]
- Mahmoud, S.H.; Moselhy, W.A.; El-Khashab, L.A.A.; Abdelbaset, B.Z.; Seufi, A.M. Variations induced in electrophoretic pattern of haemolymph proteins of flesh fly, sarcophaga argyrostoma (diptera: Sarcophagidae) larvae challenged with hydrogen peroxide. Int. J. Adv. Res. 2016. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.S.; Cook, M.L. Induction of Lysozymelike Activity in the Hemolymph and Hemocytes of an Insect, Spodoptera Eridania. J. Invertebr. Pathol. 1979, 33, 197–203. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Accession No. | Reverse | Forward |
|---|---|---|---|
| MdSOD1 | JF919738 | 5′CCTCGCCCAAAATCATCTGG′3 | 5′GCCCAATGATGATGCCTCTC′3 |
| β- actin | JN969088 | 5′CGGTGGTGGTGAACGAGTAA′3 | 5′ACACACCAAAATGTGCGACG′3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Garawani, I.; El-Seedi, H.; Khalifa, S.; El Azab, I.H.; Abouhendia, M.; Mahmoud, S. Enhanced Antioxidant and Cytotoxic Potentials of Lipopolysaccharides-Injected Musca domestica Larvae. Pharmaceutics 2020, 12, 1111. https://doi.org/10.3390/pharmaceutics12111111
El-Garawani I, El-Seedi H, Khalifa S, El Azab IH, Abouhendia M, Mahmoud S. Enhanced Antioxidant and Cytotoxic Potentials of Lipopolysaccharides-Injected Musca domestica Larvae. Pharmaceutics. 2020; 12(11):1111. https://doi.org/10.3390/pharmaceutics12111111
Chicago/Turabian StyleEl-Garawani, Islam, Hesham El-Seedi, Shaden Khalifa, Islam H. El Azab, Marwa Abouhendia, and Shaymaa Mahmoud. 2020. "Enhanced Antioxidant and Cytotoxic Potentials of Lipopolysaccharides-Injected Musca domestica Larvae" Pharmaceutics 12, no. 11: 1111. https://doi.org/10.3390/pharmaceutics12111111