Latest Advances in the Development of Eukaryotic Vaults as Targeted Drug Delivery Systems


Abstract
:
1. Introduction
2. Eukaryotic Vaults
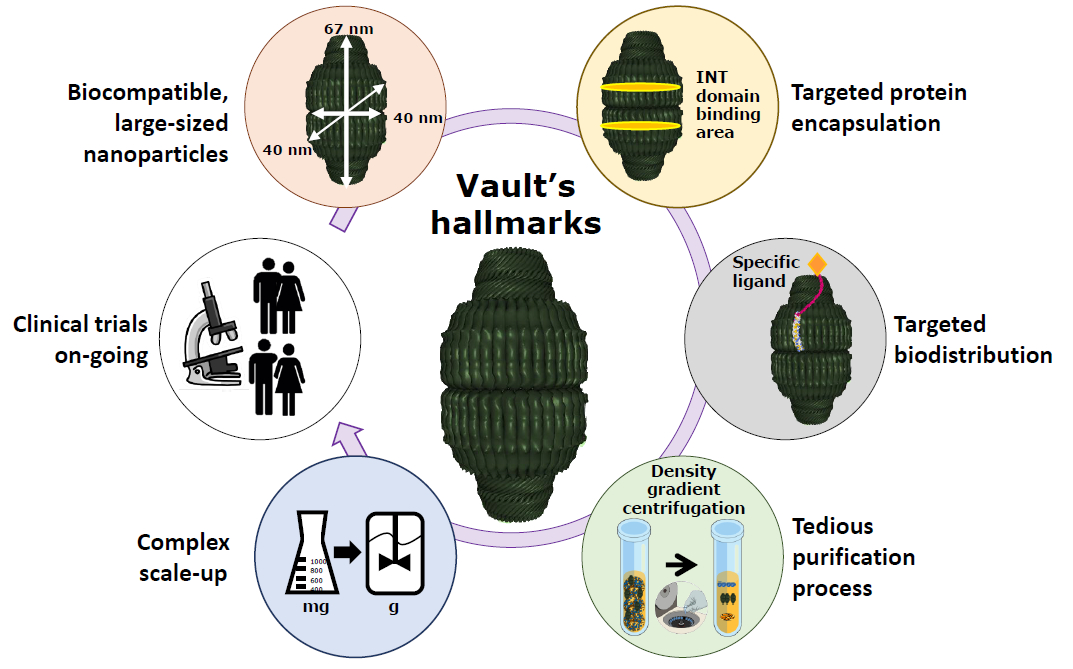
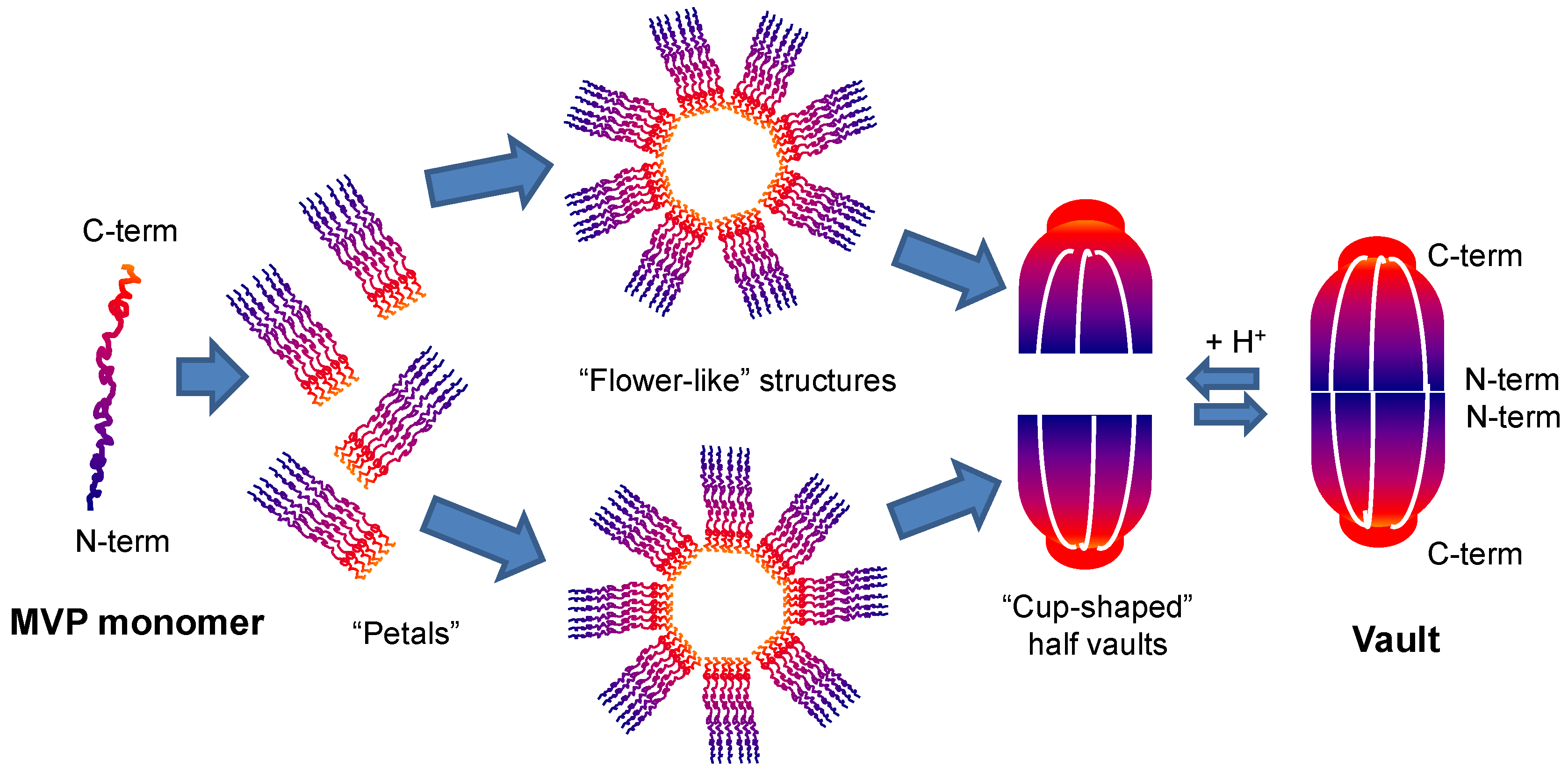
2.1. Vaults Structure
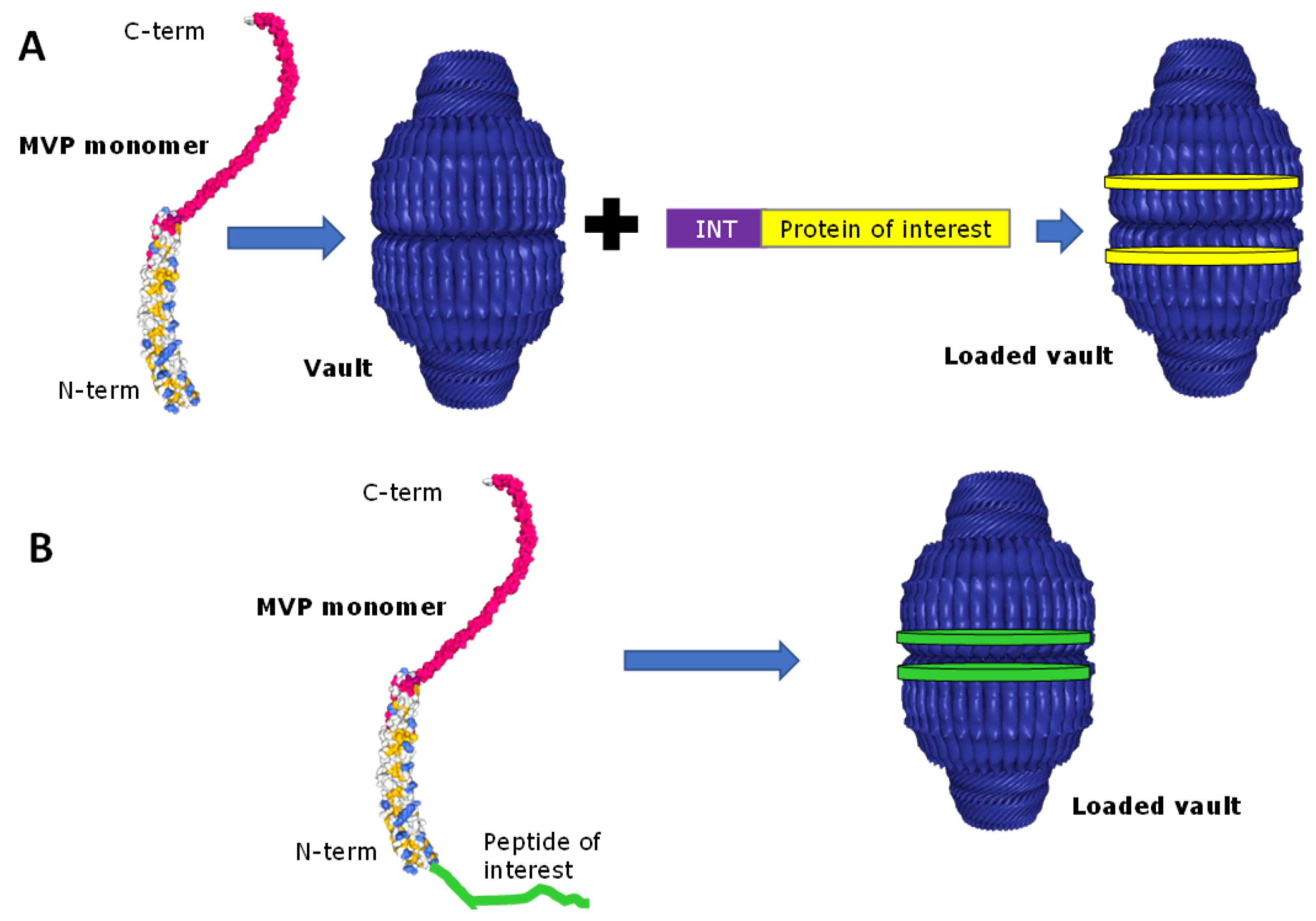
2.2. Drug Encapsulation within Vaults
2.3. Vault Targeting and Internalization
3. Recombinant Vaults Production and Purification
4. Vaults Applications in Nanomedicine and Clinical Trials
5. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Freitas, R.A. What is nanomedicine? Nanomed. Nanotechnol. Biol. Med. 2005, 1, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Burgess, R. Understanding nanomedicine: An introductory textbook; Pan Stanford: Singapore, 2012; ISBN 9789814303521. [Google Scholar]
- Safari, J.; Zarnegar, Z. Advanced drug delivery systems: Nanotechnology of health design: A review. J. Saudi Chem. Soc. 2014, 18, 85–99. [Google Scholar] [CrossRef]
- Li, W.; Feng, S.; Guo, Y. Tailoring polymeric micelles to optimize delivery to solid tumors. Nanomedicine 2012, 7, 1235–1252. [Google Scholar] [CrossRef] [PubMed]
- Pridgen, E.M.; Alexis, F.; Farokhzad, O.C. Polymeric nanoparticle drug delivery technologies for oral delivery applications. Expert Opin. Drug Deliv. 2015, 12, 1459–1473. [Google Scholar] [CrossRef] [Green Version]
- Mishra, V.; Bansal, K.; Verma, A.; Yadav, N.; Thakur, S.; Sudhakar, K.; Rosenholm, J. Solid Lipid Nanoparticles: Emerging Colloidal Nano Drug Delivery Systems. Pharmaceutics 2018, 10, 191. [Google Scholar] [CrossRef] [PubMed]
- Beloqui, A.; Solinís, M.Á.; Rodríguez-Gascón, A.; Almeida, A.J.; Préat, V. Nanostructured lipid carriers: Promising drug delivery systems for future clinics. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.W.Y.; Amador, C.; Wang, L.C.; Mody, U.V.; Bally, M.B. What Drives Innovation: The Canadian Touch on Liposomal Therapeutics. Pharmaceutics 2019, 11, 124. [Google Scholar] [CrossRef] [PubMed]
- Jiao, M.; Zhang, P.; Meng, J.; Li, Y.; Liu, C.; Luo, X.; Gao, M. Recent advancements in biocompatible inorganic nanoparticles towards biomedical applications. Biomater. Sci. 2018, 6, 726–745. [Google Scholar] [CrossRef]
- Chauhan, A. Dendrimers for Drug Delivery. Molecules 2018, 23, 938. [Google Scholar] [CrossRef]
- Liu, Y.-L.; Chen, D.; Shang, P.; Yin, D.-C. A review of magnet systems for targeted drug delivery. J. Control. Release 2019, 302, 90–104. [Google Scholar] [CrossRef]
- Hainline, K.M.; Fries, C.N.; Collier, J.H. Progress Toward the Clinical Translation of Bioinspired Peptide and Protein Assemblies. Adv. Healthc. Mater. 2018, 7, 1700930. [Google Scholar] [CrossRef] [PubMed]
- DeFrates, K.; Markiewicz, T.; Gallo, P.; Rack, A.; Weyhmiller, A.; Jarmusik, B.; Hu, X. Protein Polymer-Based Nanoparticles: Fabrication and Medical Applications. Int. J. Mol. Sci. 2018, 19, 1717. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Lee, J.; Min, J.; Kang, S. Developing Genetically Engineered Encapsulin Protein Cage Nanoparticles as a Targeted Delivery Nanoplatform. Biomacromolecules 2014, 15, 3794–3801. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, A. Nanoparticles in translational science and medicine; Academic Press: London, UK, 2011; ISBN 9780124160200. [Google Scholar]
- Elzoghby, A.O.; Samy, W.M.; Elgindy, N.A. Protein-based nanocarriers as promising drug and gene delivery systems. J. Control. Release 2012, 161, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Unzueta Elorza, U. De novo design of self-assembling protein nanoparticles towards the gene therapy of colorectal cancer. PhD Thesis, Universitat Autònoma de Barcelona, Bellaterra, Spain, 28 June 2013. [Google Scholar]
- Buehler, D.C.; Marsden, M.D.; Shen, S.; Toso, D.B.; Wu, X.; Loo, J.A.; Zhou, Z.H.; Kickhoefer, V.A.; Wender, P.A.; Zack, X.J.A.; et al. Bioengineered Vaults: Self-Assembling Protein Shell-Lipophilic Core Nanoparticles for Drug Delivery. ACS Nano 2014, 8, 7723–7732. [Google Scholar] [CrossRef] [PubMed]
- Benner, N.L.; Zang, X.; Buehler, D.C.; Kickhoefer, V.A.; Rome, M.E.; Rome, L.H.; Wender, P.A. Vault Nanoparticles: Chemical Modifications for Imaging and Enhanced Delivery. ACS Nano 2017, 11, 872–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llauró, A.; Guerra, P.; Kant, R.; Bothner, B.; Verdaguer, N.; De Pablo, P.J. Decrease in pH destabilizes individual vault nanocages by weakening the inter-protein lateral interaction. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Zheng, C.L.; Sumizawa, T.; Che, X.F.; Tsuyama, S.; Furukawa, T.; Haraguchi, M.; Gao, H.; Gotanda, T.; Jueng, H.C.; Murata, F.; et al. Characterization of MVP and VPARP assembly into vault ribonucleoprotein complexes. Biochem. Biophys. Res. Commun. 2004, 326, 100–107. [Google Scholar] [CrossRef]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the clinic. Bioeng. Transl. Med. 2016, 1, 10–29. [Google Scholar] [CrossRef]
- Chen, L.; Bai, G.; Yang, S.; Yang, R.; Zhao, G. Encapsulation of curcumin in recombinant human H-chain ferritin increases its water-solubility and stability. FRIN 2014, 62, 1147–1153. [Google Scholar] [CrossRef]
- Flenniken, M.L.; Liepold, L.O.; Crowley, B.E.; Willits, D.A.; Young, M.J.; Douglas, T. Selective attachment and release of a chemotherapeutic agent from the interior of a protein cage architecture. Chem. Commun. 2005, 2, 447–449. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Tsukihara, T. Structural studies of large nucleoprotein particles, vaults. Japan Acad. 2012, 88, 416–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, T.; Young, M. Viruses: Making Friends with Old Foes. Science. 2006, 312, 873–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, L.; Wang, Q. Adaptations of nanoscale viruses and other protein cages for medical applications. Nanomedicine Nanotechnology, Biol. Med. 2006, 2, 137–149. [Google Scholar] [CrossRef]
- Lee, E.J.; Lee, N.K.; Kim, I.-S. Bioengineered protein-based nanocage for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 157–171. [Google Scholar] [CrossRef]
- Goldsmith, L.E.; Pupols, M.; Kickhoefer, V.A.; Rome, L.H.; Monbouquette, H.G. Utilization of a Protein “Shuttle” To Load Vault Nanocapsules with Gold Probes and Proteins. ACS Nano 2009, 3, 3175–3183. [Google Scholar] [CrossRef]
- Kedersha, N.L.; Rome, L.H. Isolation and Characterization of a Novel Ribonucleoprotein Particle: Large Structures Contain a Single Species of Small RNA. J. Cell Biol. 1986, 103, 699–709. [Google Scholar] [CrossRef]
- Suprenant, K.A. Vault Ribonucleoprotein Particles: Sarcophagi, Gondolas, or Safety Deposit Boxes? Biochemistry 2002, 41, 14447–14454. [Google Scholar] [CrossRef]
- Liu, S.; Hao, Q.; Peng, N.; Yue, X.; Wang, Y.; Chen, Y.; Wu, J.; Zhu, Y. Major vault protein: A virus-induced host factor against viral replication through the induction of type-I interferon. Hepatology 2012, 56, 57–66. [Google Scholar] [CrossRef]
- Yang, J.; Srinivasan, A.; Sun, Y.; Mrazek, J.; Shu, Z.; Kickhoefer, V.A.; Rome, L.H. Vault nanoparticles engineered with the protein transduction domain, TAT48, enhances cellular uptake. Integr. Biol. 2013, 5, 151–158. [Google Scholar] [CrossRef]
- Kedersha, N.L.; Heuser, J.E.; Chugani, D.C.; Rome, L.H. Vaults. III. Vault ribonucleoprotein particles open into flower-like structures with octagonal symmetry. J. Cell Biol. 1991, 112, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.H.; Kickhoefer, V.A.; Sievers, S.A.; Rome, L.H.; Eisenberg, D. Draft Crystal Structure of the Vault Shell at 9-Å Resolution. PLoS Biol. 2007, 5, e318. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Kato, K.; Yamashita, E.; Sumizawa, T.; Zhou, Y.; Yao, M.; Iwasaki, K.; Yoshimura, M.; Tsukihara, T. The Structure of Rat Liver Vault at 3.5 Angstrom Resolution. Science. 2009, 323, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Woodward, C.L.; Mendonça, L.M.; Jensen, G.J. Direct visualization of vaults within intact cells by electron cryo-tomography. Cell. Mol. Life Sci. 2015, 72, 3401–3409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Querol-Audí, J.; Casañas, A.; Usón, I.; Luque, D.; Castón, J.R.; Fita, I.; Verdaguer, N. The mechanism of vault opening from the high resolution structure of the N-terminal repeats of MVP. EMBO J. 2009, 28, 3450–3457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poderycki, M.J.; Kickhoefer, V.A.; Kaddis, C.S.; Raval-Fernandes, S.; Johansson, E.; Zink, J.I.; Loo, J.A.; Rome, L.H. The vault exterior shell is a dynamic structure that allows incorporation of vault-associated proteins into its interior. Biochemistry 2006, 45, 12184–12193. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, N.M.; Buchman, G.W.; Rome, L.H.; Maynard, H.D. Dual pH- and temperature-responsive protein nanoparticles. Eur. Polym. J. 2015, 69, 532–539. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, L.E.; Yu, M.; Rome, L.H.; Monbouquette, H.G. Vault Nanocapsule Dissociation into Halves Triggered at Low pH. Biochemistry 2007, 46, 2865–2875. [Google Scholar] [CrossRef]
- Esfandiary, R.; Kickhoefer, V.A.; Rome, L.H.; Joshi, S.B.; Middaugh, C.R. Structural Stability of Vault Particles. J. Pharm. Sci. 2009, 98, 1376–1386. [Google Scholar] [CrossRef]
- Ding, K.; Zhang, X.; Mrazek, J.; Kickhoefer, V.A.; Lai, M.; Ng, H.L.; Yang, O.O.; Rome, L.H.; Zhou, Z.H. Solution Structures of Engineered Vault Particles. Structure 2018, 26, 619–626. [Google Scholar] [CrossRef]
- Llauró, A.; Guerra, P.; Irigoyen, N.; Rodríguez, J.F.; Verdaguer, N.; de Pablo, P.J. Mechanical stability and reversible fracture of vault particles. Biophys. J. 2014, 106, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Mikyas, Y.; Makabi, M.; Raval-Fernandes, S.; Harrington, L.; Kickhoefer, V.A.; Rome, L.H.; Stewart, P.L. Cryoelectron Microscopy Imaging of Recombinant and Tissue Derived Vaults: Localization of the MVP N Termini and VPARP. J. Mol. Biol. 2004, 344, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Kickhoefer, V.A.; Han, M.; Raval-fernandes, S.; Poderycki, M.J.; Moniz, R.J.; Vaccari, D.; Silvestry, M.; Stewart, P.L.; Kelly, K.A.; Rome, L.H. Targeting Vault Nanoparticles to Specific Cell Surface Receptor. ACS Nano 2009, 3, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Kickhoefer, V.A.; Siva, A.C.; Kedersha, N.L.; Inman, E.M.; Ruland, C.; Streuli, M.; Rome, L.H. The 193-Kd Vault Protein, Vparp, Is a Novel Poly(Adp-Ribose) Polymerase. J. Cell Biol. 1999, 146, 917–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.; Kickhoefer, V.A.; Nemerow, G.R.; Rome, L.H. Targeted Vault Nanoparticles Engineered with an Endosomolytic Peptide Deliver Biomolecules to the Cytoplasm. ACS Nano 2011, 5, 6128–6137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kickhoefer, V.A.; Garcia, Y.; Mikyas, Y.; Johansson, E.; Zhou, J.C.; Raval-Fernandes, S.; Minoofar, P.; Zink, J.I.; Dunn, B.; Stewart, P.L.; et al. Engineering of vault nanocapsules with enzymatic and fluorescent properties. Proc. Natl. Acad. Sci. U.S.A. 2005, 102, 4348–4352. [Google Scholar] [CrossRef] [Green Version]
- van Zon, A.; Mossink, M.H.; Schoester, M.; Scheffer, G.L.; Scheper, R.J.; Sonneveld, P.; Wiemer, E.A.C. Structural Domains of Vault Proteins: A Role for the Coiled Coil Domain in Vault Assembly. Biochem. Biophys. Res. Commun. 2002, 291, 535–541. [Google Scholar] [CrossRef]
- Yu, K.; Yau, Y.H.; Sinha, A.; Tan, T.; Kickhoefer, V.A.; Rome, L.H.; Lee, H.; Shochat, S.G.; Lim, S. Modulation of the Vault Protein-Protein Interaction for Tuning of Molecular Release. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Kozlov, G.; Vavelyuk, O.; Minailiuc, O.; Banville, D.; Gehring, K.; Ekiel, I. Solution Structure of a Two-repeat Fragment of Major Vault Protein. J. Mol. Biol. 2006, 356, 444–452. [Google Scholar] [CrossRef]
- Kar, U.K.; Srivastava, M.K.; Andersson, Å.; Baratelli, F.; Huang, M.; Kickhoefer, V.A.; Dubinett, S.M.; Rome, L.H.; Sharma, S. Novel ccl21-vault nanocapsule intratumoral delivery inhibits lung cancer growth. PLoS One 2011, 6, 4–11. [Google Scholar] [CrossRef]
- Champion, C.I.; Kickhoefer, V.A.; Liu, G.; Moniz, R.J.; Freed, A.S.; Bergmann, L.L.; Vaccari, D.; Raval-Fernandes, S.; Chan, A.M.; Rome, L.H.; et al. A vault nanoparticle vaccine induces protective mucosal immunity. PLoS One 2009, 4, e5409. [Google Scholar] [CrossRef] [PubMed]
- Kar, U.K.; Jiang, J.; Champion, C.I.; Salehi, S.; Srivastava, M.; Sharma, S.; Rabizadeh, S.; Niazi, K.; Kickhoefer, V.; Rome, L.H.; et al. Vault nanocapsules as adjuvants favor cell-mediated over antibody-mediated immune responses following immunization of mice. PLoS One 2012, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kickhoefer, V.A.; Ng, B.C.; Gopal, A.; Bentolila, L.A.; John, S.; Tolbert, S.H.; Rome, L.H. Vaults Are Dynamically Unconstrained Cytoplasmic Nanoparticles Capable of Half Vault Exchange. ACS Nano 2010, 4, 7229–7240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rome, L.H.; Kickhoefer, V.A. Development of the Vault Particle as a Platform Technology. ACS Nano 2013, 7, 889–902. [Google Scholar] [CrossRef] [PubMed]
- Buehler, D.C.; Toso, D.B.; Kickhoefer, V.A.; Zhou, Z.H.; Rome, L.H. Vaults engineered for hydrophobic drug delivery. Small 2011, 7, 1432–1439. [Google Scholar] [CrossRef] [PubMed]
- Stephen, A.G.; Raval-Fernandes, S.; Huynh, T.; Torres, M.; Kickhoefer, V.A.; Rome, L.H. Assembly of Vault-like Particles in Insect Cells Expressing only the Major Vault Protein. J. Biol. Chem. 2001, 276, 23217–23220. [Google Scholar] [CrossRef] [PubMed]
- Mrazek, D.A.; Hornberger, J.C.; Altar, C.A.; Degtiar, I. A Review of the Clinical, Economic, and Societal Burden of Treatment-Resistant Depression: 1996–2013. Psychiatr. Serv. 2014, 65, 977–987. [Google Scholar] [CrossRef]
- Kushnir, N.; Streatfield, S.J.; Yusibov, V. Virus-like particles as a highly efficient vaccine platform: Diversity of targets and production systems and advances in clinical development. Vaccine 2012, 31, 58–83. [Google Scholar] [CrossRef]
- Fernandes, F.; Teixeira, A.P.; Carinhas, N.; Carrondo, M.J.; Alves, P.M. Insect cells as a production platform of complex virus-like particles. Expert Rev. Vaccines 2013, 12, 225–236. [Google Scholar] [CrossRef]
- Vicente, T.; Roldão, A.; Peixoto, C.; Carrondo, M.J.T.; Alves, P.M. Large-scale production and purification of VLP-based vaccines. J. Invertebr. Pathol. 2011, 107, S42–S48. [Google Scholar] [CrossRef]
- Cregg, J.M.; Cereghino, J.L.; Shi, J.; Higgins, D.R. Recombinant Protein Expression in Pichia pastoris. Mol. Biotechnol. 2000, 16, 23–52. [Google Scholar] [CrossRef]
- Lua, L.H.L.; Connors, N.K.; Sainsbury, F.; Chuan, Y.P.; Wibowo, N.; Middelberg, A.P.J. Bioengineering virus-like particles as vaccines. Biotechnol. Bioeng. 2014, 111, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Kickhoefer, V.A.; Rome, L.H.; Foellmer, O.K.; Mahendra, S. Synthesis and assembly of human vault particles in yeast. Biotechnol. Bioeng. 2018, 115, 2941–2950. [Google Scholar] [CrossRef] [PubMed]
- Rome, L.H.; Kickhoefer, V.A.; Raval-Fernandes, S.; Stewart, P.L. Vault and vault-like carrier molecules. US Patent US7482319B2, 27 January 2009. [Google Scholar]
- Deo, V.K.; Tsuji, Y.; Yasuda, T.; Kato, T.; Sakamoto, N.; Suzuki, H.; Park, E.Y. Expression of an RSV-gag virus-like particle in insect cell lines and silkworm larvae. J. Virol. Methods 2011, 177, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.-C.; Huang, J.-H.; Lai, C.-W.; Sheng, H.-C.; Shih, S.-R.; Ho, M.-S.; Hu, Y.-C. Expression, purification and characterization of enterovirus-71 virus-like particles. World J. Gastroenterol. 2006, 12, 921. [Google Scholar] [CrossRef] [PubMed]
- Morenweiser, R. Downstream processing of viral vectors and vaccines. Gene Ther. 2005, 12, S103–S110. [Google Scholar] [CrossRef] [PubMed]
- Huhti, L.; Blazevic, V.; Nurminen, K.; Koho, T.; Hytönen, V.P.; Vesikari, T. A comparison of methods for purification and concentration of norovirus GII-4 capsid virus-like particles. Arch. Virol. 2010, 155, 1855–1858. [Google Scholar] [CrossRef] [Green Version]
- Galbiati, E.; Avvakumova, S.; La Rocca, A.; Pozzi, M.; Messali, S.; Magnaghi, P.; Colombo, M.; Prosperi, D.; Tortora, P. A fast and straightforward procedure for vault nanoparticle purification and the characterization of its endocytic uptake. Biochim. Biophys. Acta - Gen. Subj. 2018, 1862, 2254–2260. [Google Scholar] [CrossRef]
- Izquierdo, M.A.; Scheffer, G.L.; Flens, M.J.; Giaccone, G.; Broxterman, H.J.; Meijer, C.J.; van der Valk, P.; Scheper, R.J. Broad distribution of the multidrug resistance-related vault lung resistance protein in normal human tissues and tumors. Am. J. Pathol. 1996, 148, 877–887. [Google Scholar]
- Sharma, S.; Zhu, L.; Srivastava, M.K.; Harris-White, M.; Huang, M.; Lee, J.M.; Rosen, F.; Lee, G.; Wang, G.; Kickhoefer, V.; et al. CCL21 Chemokine Therapy for Lung Cancer. Int Trends Immun 2013, 1, 10–15. [Google Scholar]
- Vault Pharma An Immunology Biotech Company. Available online: https://vaultpharma.com/ (accessed on 29 April 2019).
- Endo, M.; De Graaff, M.A.; Ingram, D.R.; Lim, S.; Lev, D.C.; Briaire-De Bruijn, I.H.; Somaiah, N.; Bovée, J.V.M.G.; Lazar, A.J.; Nielsen, T.O. NY-ESO-1 (CTAG1B) expression in mesenchymal tumors. Mod. Pathol. 2015, 28, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Jiang, J.; Said-Sadier, N.; Boxx, G.; Champion, C.; Tetlow, A.; Kickhoefer, V.A.; Rome, L.H.; Ojcius, D.M.; Kelly, K.A. Activation of the NLRP3 inflammasome by vault nanoparticles expressing a chlamydial epitope. Vaccine 2015, 33, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Nagasawa, D.T.; Spasic, M.; Amolis, M.; Choy, W.; Garcia, H.M.; Prins, R.M.; Liau, L.M.; Yang, I. Endogenous Vaults and Bioengineered Vault Nanoparticles for Treatment of Glioblastomas. Neurosurg. Clin. N. Am. 2012, 23, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Abad, D.; Kickhoefer, V.A.; Rome, L.H.; Mahendra, S. Vault Nanoparticles Packaged with Enzymes as an Efficient Pollutant Biodegradation Technology. ACS Nano 2015, 9, 10931–10940. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, Y.; Kickhoefer, V.A.; Rome, L.H.; Allard, P.; Mahendra, S. A Vault-Encapsulated Enzyme Approach for Efficient Degradation and Detoxification of Bisphenol A and Its Analogues. ACS Sustain. Chem. Eng. 2019, 7, 5808–5817. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Expression System [Ref.] | Purification Method | Final Yield | Advantages | Disadvantages |
|---|---|---|---|---|
| Baculovirus infection on sf9 cells [59] | Several saccharose gradient ultra-centrifugations | ~10 mg/L | No endogenous vaults | Time-consuming, tedious downstream process |
| Baculovirus infection on sf21 cells [72] | Dialysis and size-exclusion chromatography | ~0.5–1 mg/L | No endogenous vaults Quick, easy and cheap downstream process | Low yields |
| Baculovirus oral infection on insect larvae [40] | Ion exchange (cationic) and size-exclusion chromatography | Up to several grams | No endogenous vaults High yields | Difficult scale-up Slow production rate |
| Yeast cells (Pichia pastoris) [66] | Sucrose gradient ultracentrifugation and ion exchange (anionic) chromatography | ~7–11 mg/L | No endogenous vaults Fast and cost-efficient production | Need scale-up |
| Vaults Application | Clinical Trial Stage | ||
|---|---|---|---|
| Drug Discovery | Preclinical | Phase I | |
| Oncology immunology | Pancreatic cancer Prostate cancer Head and neck cancer Renal cancer Bladder cancer Colon cancer Breast cancer Graft vs host disease Pulmonary fibrosis Solid tumors | Lung cancer Melanoma Glioblastoma | Lung cancer |
| Immunological activation for infection prevention | HPV cancer Burkholderia | Chlamydia HIV/AIDS Influenza | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz-Juan, A.; Carreño, A.; Mendoza, R.; Corchero, J.L. Latest Advances in the Development of Eukaryotic Vaults as Targeted Drug Delivery Systems. Pharmaceutics 2019, 11, 300. https://doi.org/10.3390/pharmaceutics11070300
Muñoz-Juan A, Carreño A, Mendoza R, Corchero JL. Latest Advances in the Development of Eukaryotic Vaults as Targeted Drug Delivery Systems. Pharmaceutics. 2019; 11(7):300. https://doi.org/10.3390/pharmaceutics11070300
Chicago/Turabian StyleMuñoz-Juan, Amanda, Aida Carreño, Rosa Mendoza, and José L. Corchero. 2019. "Latest Advances in the Development of Eukaryotic Vaults as Targeted Drug Delivery Systems" Pharmaceutics 11, no. 7: 300. https://doi.org/10.3390/pharmaceutics11070300