Mechanistic Studies on the Absorption-Enhancing Effects of Gemini Surfactant on the Intestinal Absorption of Poorly Absorbed Hydrophilic Drugs in Rats

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. In Vivo Enteric Absorption Studies
2.3. Preparation of Brush Border Membrane Vesicles (BBMVs)
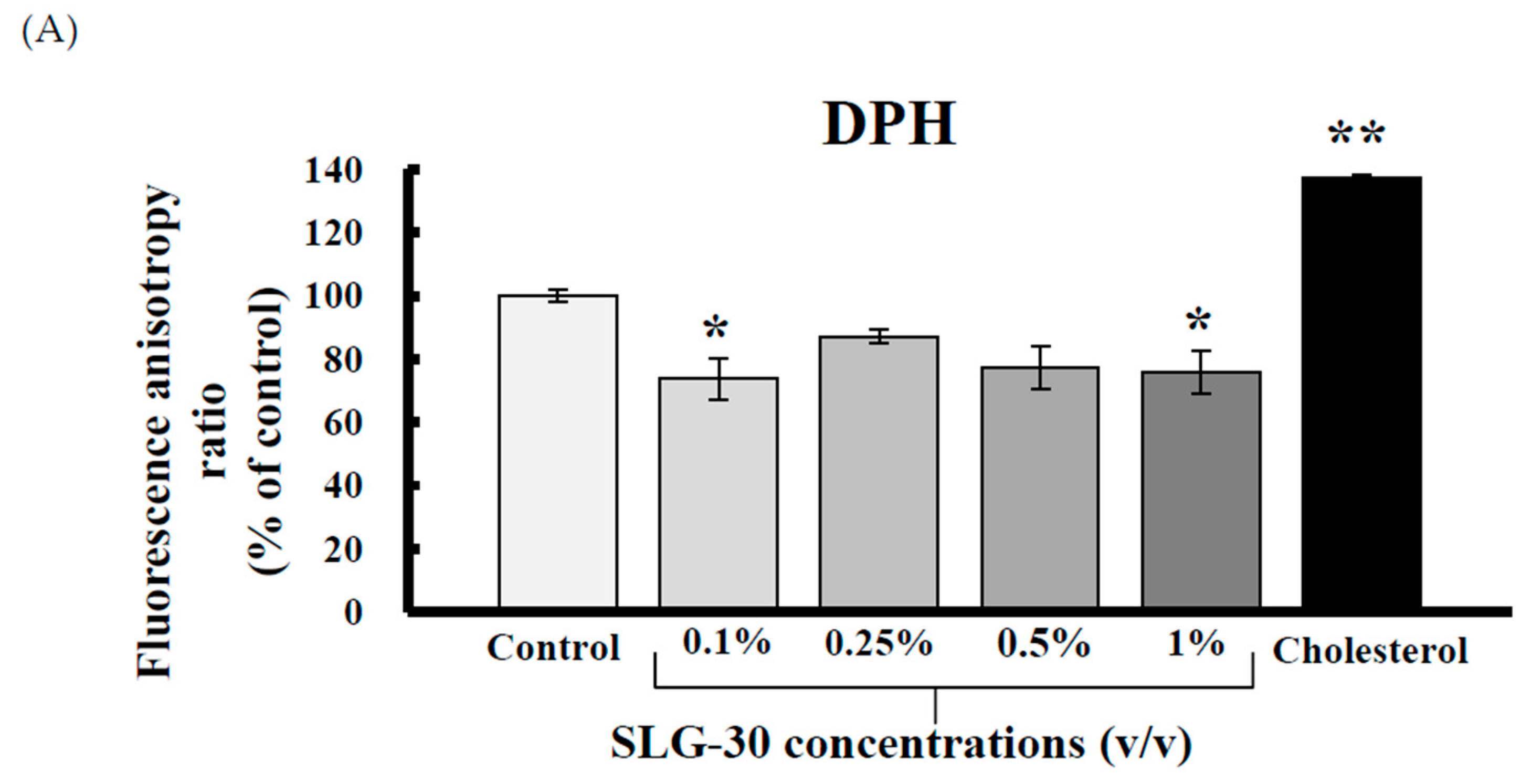
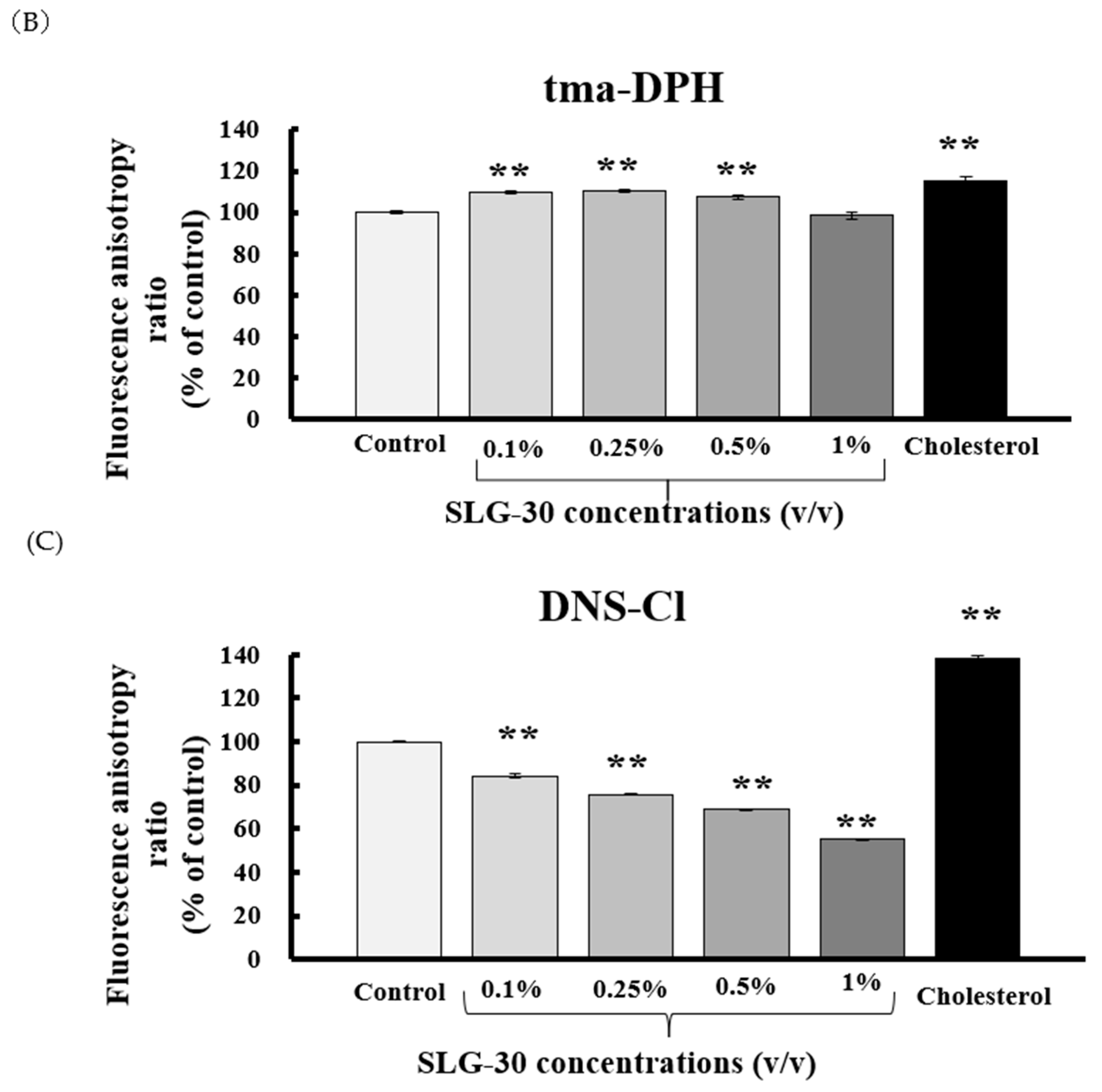
2.4. Measurement of Membrane Fluidity by Fluorescence Polarization
2.5. Measurement of Transepithelial Electrical Resistance (TEER) and the Transport of CF Using Caco-2 cell Monolayers
2.6. Assessment of Caco-2 Cell Damage
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
3.1. In Vivo Comparative Study between SLG-30 and Typical Surfactants Including Sodium Glycocholate and Sodium Laurate
3.2. Effects of SLG-30 on the Membrane Fluidity of Small Intestines in Rats
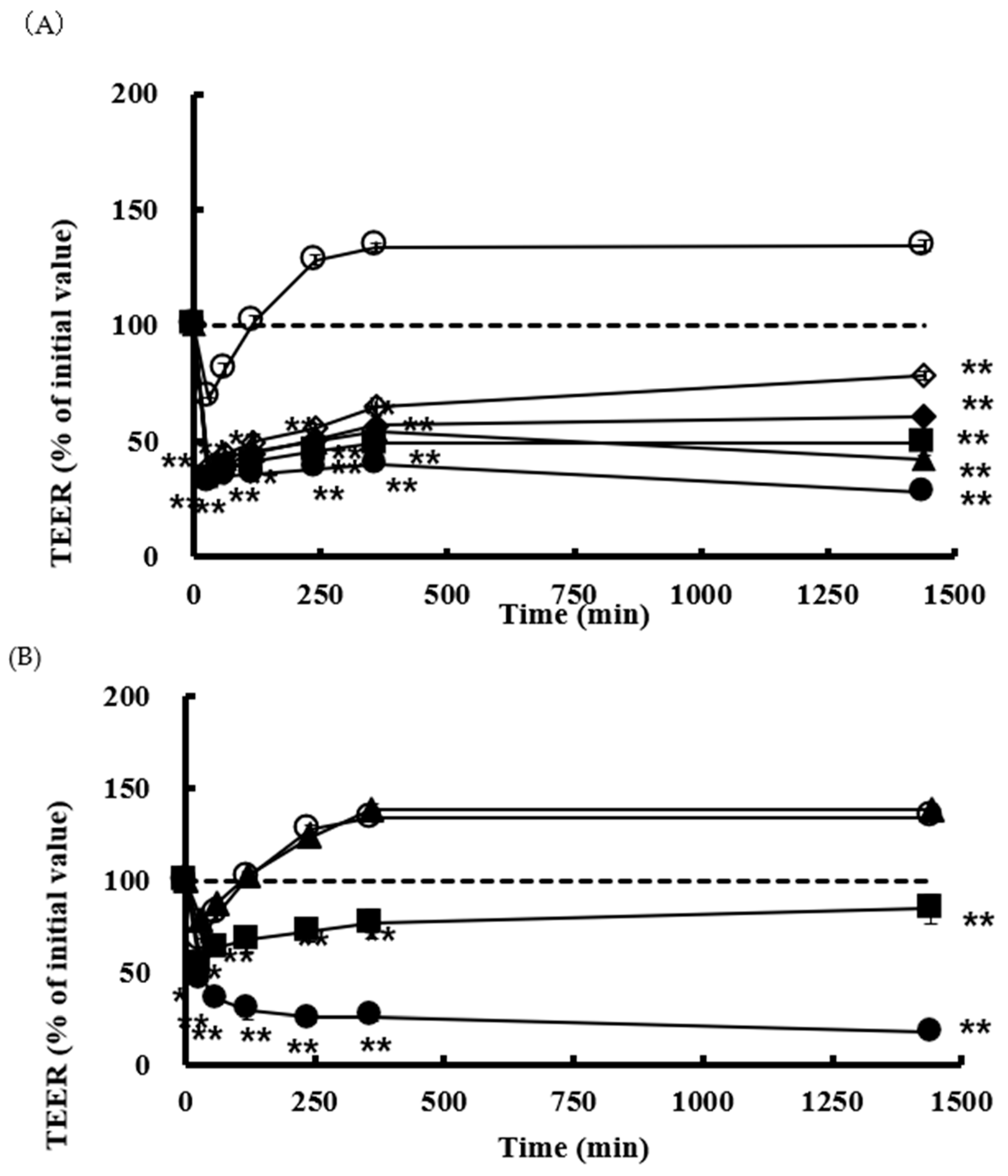
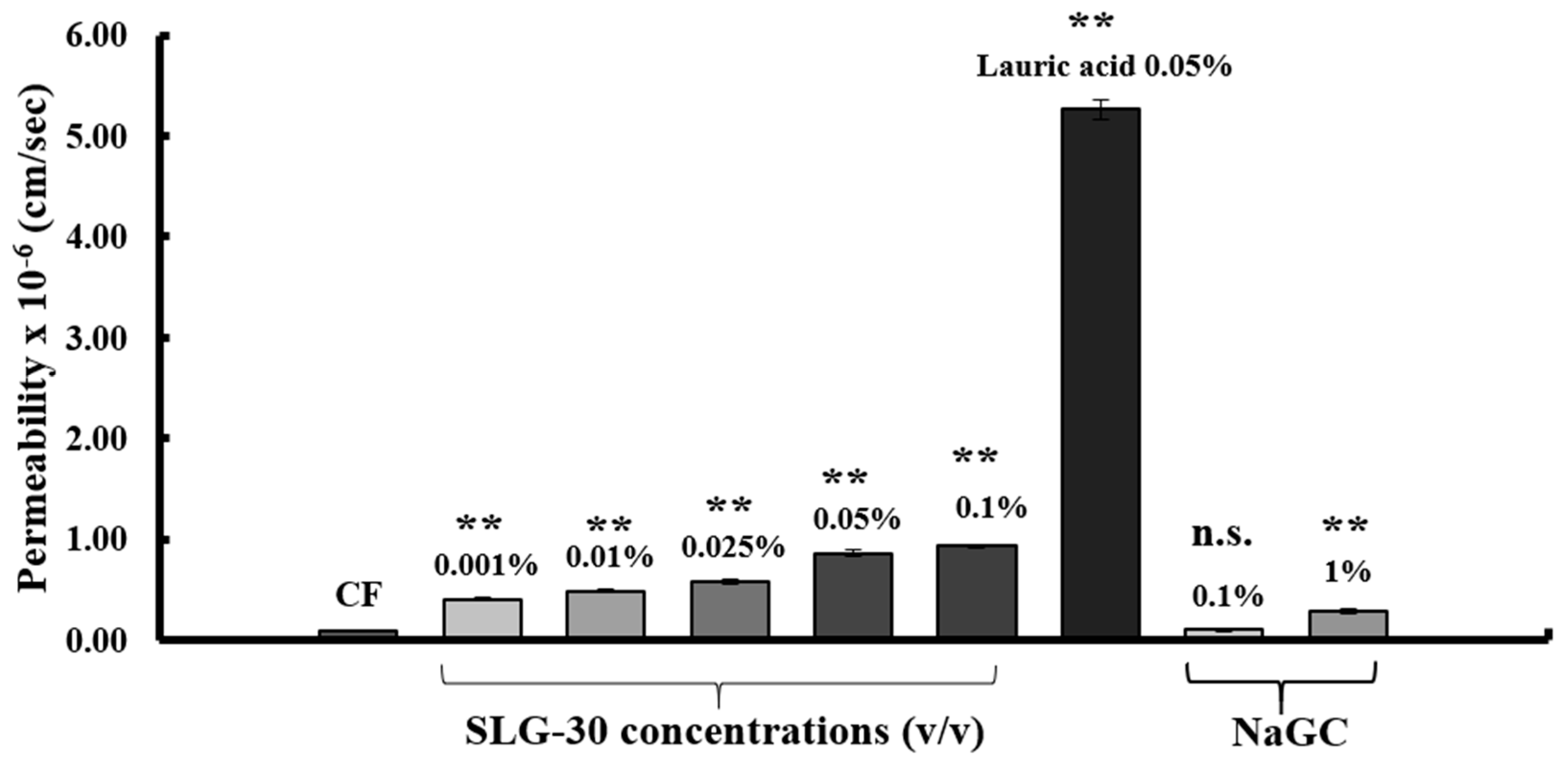
3.3. Effect of SLG-30 on TEER and Transport of CF in Caco-2 Cell Monolayers
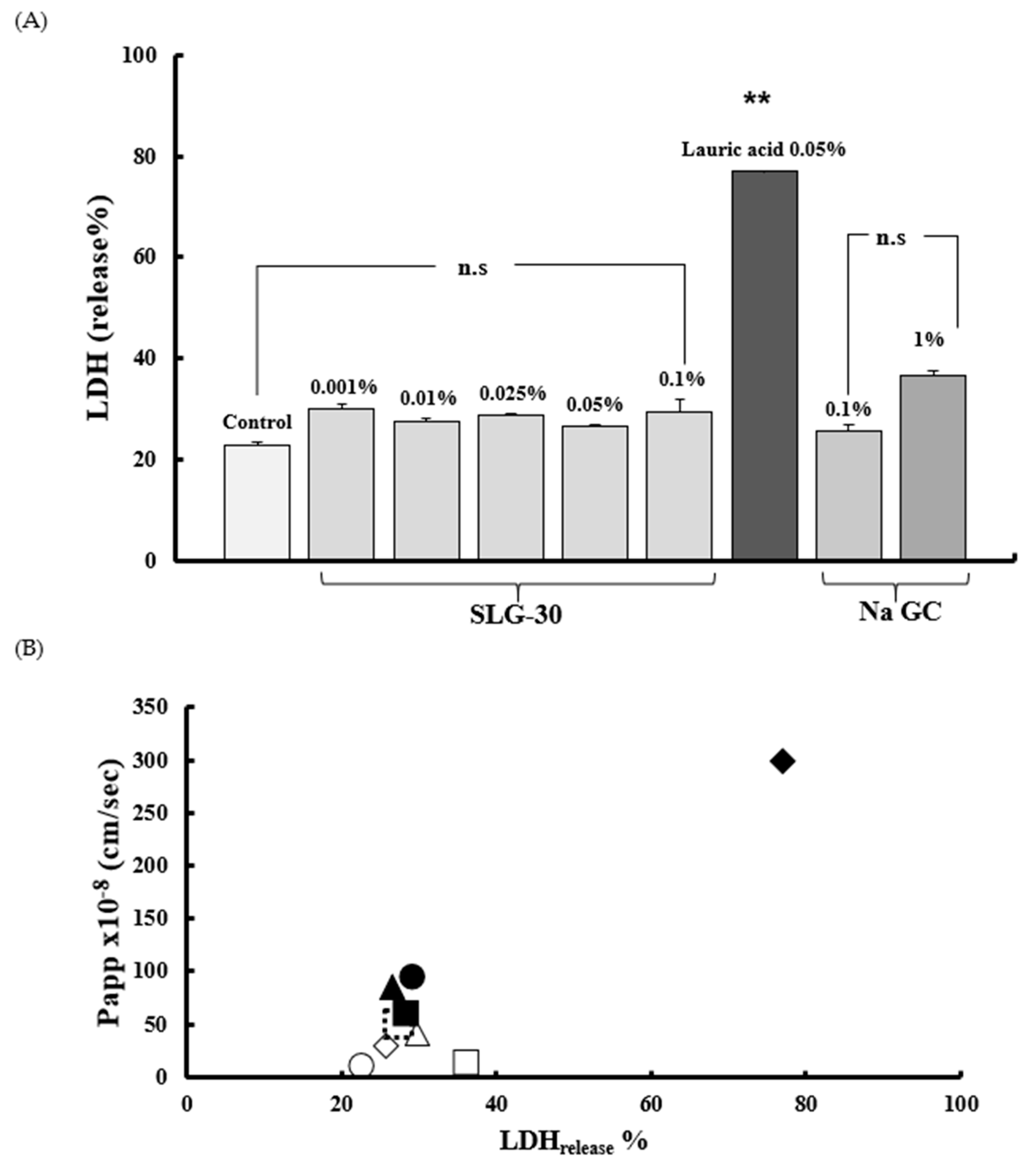
3.4. Assessment of Caco-2 Cells Injury
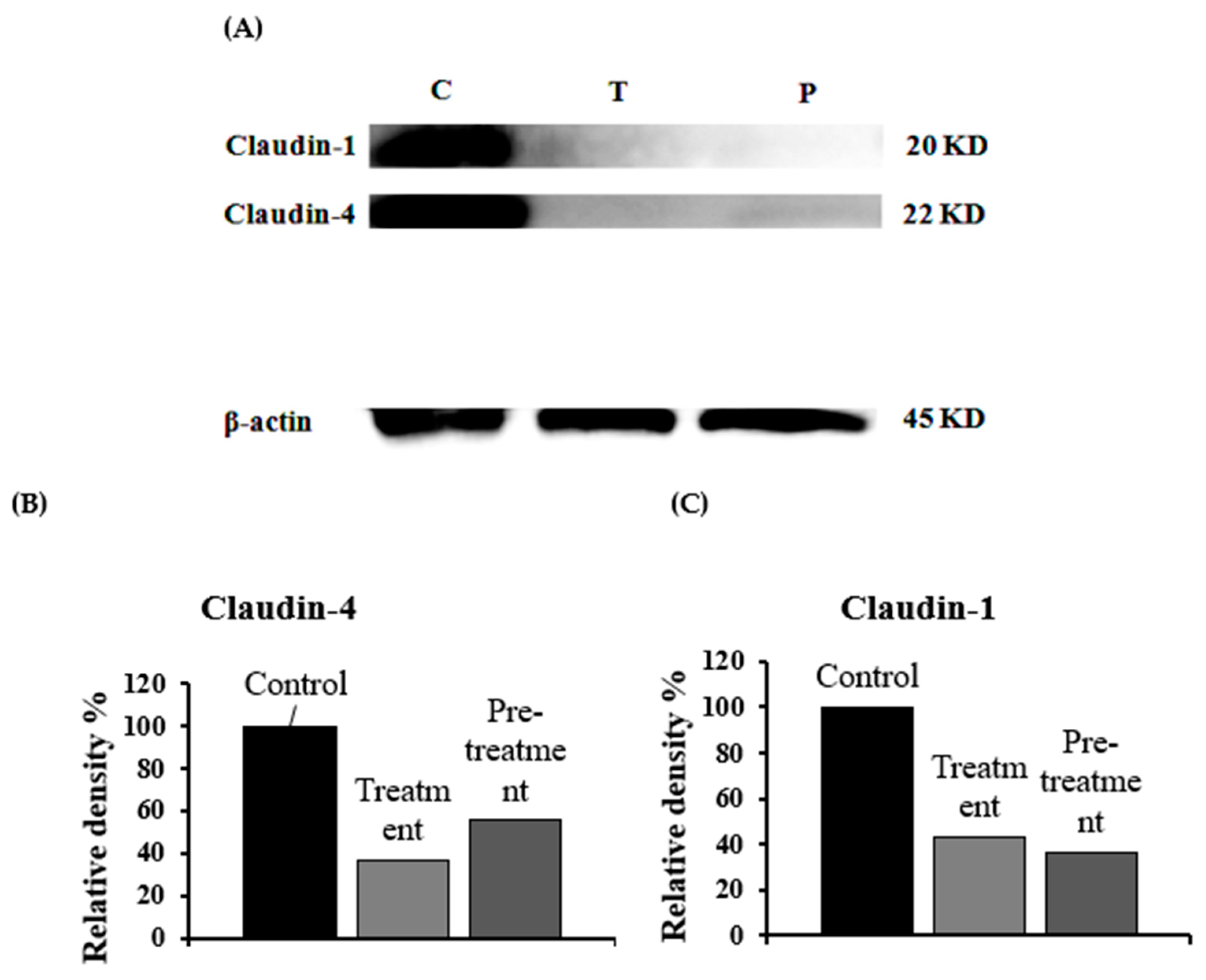
3.5. Estimation of Claudin Proteins by Western Blotting
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Beg, S.; Swain, S.; Rizwan, M.; Irfanuddin, M.; Malini, D.S. Bioavailability enhancement strategies: Basics, formulation approaches and regulatory considerations. Curr. Drug Deliv. 2011, 8, 691–702. [Google Scholar] [CrossRef]
- Adson, A.; Raub, T.J.; Burton, P.S.; Barsuhn, C.L.; Hilgers, A.R.; Audus, K.L.; Ho, N.F. Quantitative approaches to delineate paracellular diffusion in cultured epithelial cell monolayers. J. Pharm. Sci. 1994, 83, 1529–1536. [Google Scholar] [CrossRef]
- Antosova, Z.; Mackova, M.; Kral, V.; Macek, T. Therapeutic application of peptides and proteins: Parenteral forever? Trends Biotechnol. 2009, 27, 628–635. [Google Scholar] [CrossRef]
- Mitic, L.L.; Anderson, J.M. Molecular architecture of tight junctions. Annu. Rev. Physiol. 1998, 60, 121–142. [Google Scholar] [CrossRef] [PubMed]
- Fricker, G.; Drewe, J. Current concepts in intestinal peptide absorption. J. Pept. Sci. 1996, 2, 195–211. [Google Scholar] [CrossRef] [PubMed]
- Salamat-Miller, N.; Johnston, T.P. Current strategies used to enhance the paracellular transport of therapeutic polypeptides across the intestinal epithelium. Int. J. Pharm. 2005, 294, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Fix, J.A.; Engle, K.; Porter, P.A.; Leppert, P.S.; Selk, S.J.; Gardner, C.R.; Alexander, J. Acylcarnitines: Drug absorption-enhancing agents in the gastrointestinal tract. Am. J. Physiol. 1986, 251, G332–G340. [Google Scholar] [CrossRef]
- Lee, V.H.L.; Yamamoto, A. Penetration and enzymatic barriers to peptide and protein absorption. Adv. Drug Deliv. Rev. 1989, 4, 171–207. [Google Scholar] [CrossRef]
- Nellans, H.N. (B) Mechanisms of peptide and protein absorption. Adv. Drug Deliv. Rev. 1991, 7, 339–364. [Google Scholar] [CrossRef]
- Pauletti, G.M.; Okumu, F.W.; Borchardt, R.T. Effect of size and charge on the passive diffusion of peptides across Caco-2 cell monolayers via the paracellular pathway. Pharm. Res. 1997, 14, 164–168. [Google Scholar] [CrossRef]
- Aungst, B.J. Absorption enhancers: Applications and advances. AAPS J. 2012, 14, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Uchiyama, T.; Nishikawa, R.; Fujita, T.; Muranishi, S. Effectiveness and toxicity screening of various absorption enhancers in the rat small intestine: Effects of absorption enhancers on the intestinal absorption of phenol red and the release of protein and phospholipids from the intestinal membrane. J. Pharm. Pharmacol. 1996, 48, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Tatsumi, H.; Maruyama, M.; Uchiyama, T.; Okada, N.; Fujita, T. Modulation of intestinal permeability by nitric oxide donors: Implications in intestinal delivery of poorly absorbable drugs. J. Pharmacol. Exp. Ther. 2001, 296, 84–90. [Google Scholar] [PubMed]
- Gao, Y.; He, L.; Katsumi, H.; Sakane, T.; Fujita, T.; Yamamoto, A. Improvement of intestinal absorption of water-soluble macromolecules by various polyamines: Intestinal mucosal toxicity and absorption-enhancing mechanism of spermine. Int. J. Pharm. 2008, 354, 126–134. [Google Scholar] [CrossRef]
- Gao, Y.; He, L.; Katsumi, H.; Sakane, T.; Fujita, T.; Yamamoto, A. Improvement of intestinal absorption of insulin and water-soluble macromolecular compounds by chitosan oligomers in rats. Int. J. Pharm. 2008, 359, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Fujimori, T.; Kawaguchi, N.; Tsujimoto, Y.; Nishimi, M.; Dong, Z.; Katsumi, H.; Sakane, T.; Yamamoto, A. Polyamidoamine dendrimers as novel potential absorption enhancers for improving the small intestinal absorption of poorly absorbable drugs in rats. J. Control. Release 2011, 149, 21–28. [Google Scholar] [CrossRef]
- Nakaya, Y.; Takaya, M.; Hinatsu, Y.; Alama, T.; Kusamori, K.; Katsumi, H.; Sakane, T.; Yamamoto, A. Enhanced oral delivery of bisphosphonate by novel absorption enhancers: Improvement of intestinal absorption of alendronate by N-acyl amino acid and N-acyl taurates and their absorption-enhancing mechanisms. J. Pharm. Sci. 2016, 105, 3680–3690. [Google Scholar] [CrossRef]
- Alama, T.; Katayama, H.; Hirai, S.; Ono, S.; Kajiyama, A.; Kusamori, K.; Katsumi, H.; Sakane, T.; Yamamoto, A. Enhanced oral delivery of alendronate by sucrose fatty acids esters in rats and their absorption-enhancing mechanisms. Int. J. Pharm. 2016, 515, 476–489. [Google Scholar] [CrossRef]
- Dahlgren, D.; Roos, C.; Johansson, P.; Tannergren, C.; Lundqvist, A.; Langguth, P.; Sjöblom, M.; Sjögren, E.; Lennernas, H. The effects of three absorption-modifying critical excipients on the in vivo intestinal absorption of six model compounds in rats and dogs. Int. J. Pharm. 2018, 547, 158–168. [Google Scholar] [CrossRef]
- Maher, S.; Mrsny, R.J.; Braydenc, D.J. Intestinal permeation enhancers for oral peptide delivery. Adv. Drug Deliv. Rev. 2016, 106, 277–319. [Google Scholar] [CrossRef] [Green Version]
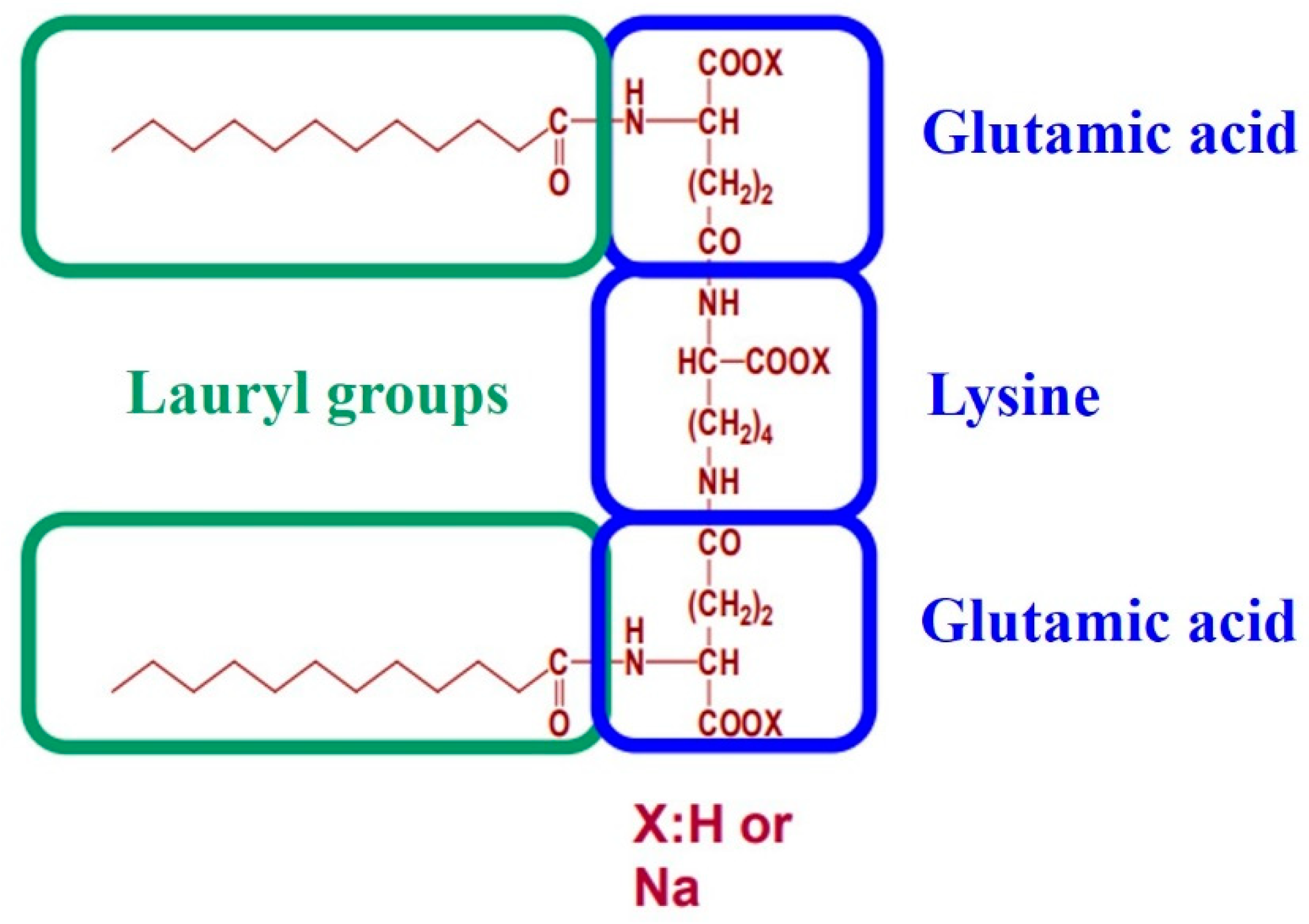
- Hait, S.K.; Moulik, S.P. Gemini surfactants: A distinct class of self- assembling molecules. Curr. Sci. Gemini Surfactants 2002, 82, 1101–1111. [Google Scholar]
- Alama, T.; Kusamori, K.; Katsumi, H.; Sakane, T.; Yamamoto, A. Absorption-enhancing effects of gemini surfactant on the intestinal absorption of poorly absorbed hydrophilic drugs including peptide and protein drugs in rats. Int. J. Pharm. 2016, 499, 58–66. [Google Scholar] [CrossRef]
- Kaneko, D.; Olsson, U.; Sakamoto, K. Self-assembly in some N-lauroyl-l-glutamate/water Systems. Langmuir 2002, 18, 4699–4703. [Google Scholar] [CrossRef]
- Yamamoto, A.; Taniguchi, T.; Rikyuu, K.; Tsuji, T.; Fujita, T.; Murakami, M.; Muranishi, S. Effects of various protease inhibitors on the intestinal absorption and degradation of insulin in rats. Pharm. Res. 1994, 11, 1496–1500. [Google Scholar] [CrossRef]
- Bhor, V.M.; Sivakami, S. Regional variations in intestinal brush border membrane fluidity and function during diabetes and the role of oxidative stress and non-enzymatic glycation. Mol. Cell. Biochem. 2003, 252, 125–132. [Google Scholar] [CrossRef]
- Ganapathy, V.; Mendicino, J.F.; Leibach, F.H. Transport of glycyl-l-proline into intestinal and renal brush border vesicles from rabbit. J. Biol. Chem. 1981, 256, 118–124. [Google Scholar]
- Prabhu, R.; Balasubramanian, K.A. A novel method of preparation of small intestinal brush border membrane vesicles by polyethylene glycol precipitation. Anal. Biochem. 2001, 289, 157–161. [Google Scholar] [CrossRef]
- Koga, K.; Kusawake, Y.; Ito, Y.; Sugioka, N.; Shibata, N.; Takada, K. Enhancing mechanism of Labrasol on intestinal membrane permeability of the hydrophilic drug gentamicin sulfate. Eur. J. Pharm. Biopharm. 2006, 64, 82–91. [Google Scholar] [CrossRef]
- Ohyashiki, T.; Sakata, N.; Matsui, K. A decrease of lipid fluidity of the porcine intestinal brush-border membranes by treatment with malondialdehyde. J. Biochem. 1992, 111, 419–423. [Google Scholar] [CrossRef]
- Hidalgo, I.J.; Raub, T.J.; Borchardt, R.T. Characterization of the human colon carcinoma cell line (Caco-2) as a model system for intestinal epithelial permeability. Gastroenterology 1989, 96, 736–749. [Google Scholar] [CrossRef]
- Choksakulnimitr, S.; Masuda, S.; Tokuda, H.; Takakura, Y.; Hashida, M. In vitro cytotoxicity of macromolecules in different cell culture systems. J. Control. Release 1995, 34, 233–241. [Google Scholar] [CrossRef]
- Quan, Y.S.; Hattori, K.; Lundborg, E.; Fujita, T.; Murakami, M.; Muranishi, S.; Yamamoto, A. Effectiveness and toxicity screening of various absorption enhancers using Caco-2 cell monolayers. Biol. Pharm. Bull. 1998, 21, 615–620. [Google Scholar] [CrossRef]
- Sugibayashi, K.; Onuki, Y.; Takayama, K. Displacement of tight junction proteins from detergent-resistant membrane domains by treatment with sodium caprate. Eur. J. Pharm. Sci. 2009, 36, 246–253. [Google Scholar] [CrossRef]
- Takizawa, Y.; Kishimoto, H.; Kitazato, T.; Tomita, M.; Hayashi, M. Changes in protein and mRNA expression levels of claudin family after mucosal lesion by intestinal ischemia/reperfusion. Int. J. Pharm. 2012, 426, 82–89. [Google Scholar] [CrossRef]
- Gordon, G.S.; Moses, A.C.; Silver, R.D.; Flier, J.S.; Carey, M.C. Nasal absorption of insulin: Enhancement by hydrophobic bile salts. Proc. Natl. Acad. Sci. USA 1985, 82, 7419–7423. [Google Scholar] [CrossRef]
- Yamamoto, A.; Hayakawa, E.; Lee, V.H. Insulin and proinsulin proteolysis in mucosal homogenates of the albino rabbit: Implications in peptide delivery from nonoral routes. Life Sci. 1990, 47, 2465–2474. [Google Scholar] [CrossRef]
- Rinken, A.; Harro, J.; Engström, L.; Oreland, L. Role of fluidity of membranes on the guanyl nucleotide-dependent binding of cholecystokinin-8S to rat brain cortical membranes. Biochem. Pharmacol. 1998, 55, 423–431. [Google Scholar] [CrossRef]
- Schedl, H.P.; Christensen, K.K.; Clark, E.D.; Buettner, G.R. Surface charge, fluidity, and calcium uptake by rat intestinal brush-border vesicles. Biochim. Biophys. Acta Biomembr. 1995, 1234, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Aungst, B.J. Intestinal permeation enhancers. J. Pharm. Sci 2000, 89, 429–442. [Google Scholar] [CrossRef]
- Chao, A.C.; Nguyen, J.V.; Broughall, M.; Griffin, A.; Fix, J.A.; Daddona, P.E. In vitro and in vivo evaluation of effects of sodium caprate on enteral peptide absorption and on mucosal morphology. Int. J. Pharm. 1999, 191, 15–24. [Google Scholar] [CrossRef]
- Morita, K.; Furuse, M.; Fujimoto, K.; Tsukita, S. Claudin multigene family encoding four-transmembrane domain protein components of tight junction strands. Proc. Natl. Acad. Sci. USA 1999, 96, 511–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonoda, N.; Furuse, M.; Sasaki, H.; Yonemura, S.; Katahira, J.; Horiguchi, Y.; Tsukita, S. Clostridium perfringens enterotoxin fragment removes specific claudins from tight junction strands: Evidence for direct involvement of claudins in tight junction barrier. J. Cell Biol. 1999, 147, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Balda, M.S.; Whitney, J.A.; Flores, C.; González, S.; Cereijido, M.; Matter, K. Functional dissociation of paracellular permeability and transepithelial electrical resistance and disruption of the apical-basolateral intramembrane diffusion barrier by expression of a mutant tight junction membrane protein. J. Cell Biol. 1996, 134, 1031–1034. [Google Scholar] [CrossRef] [PubMed]






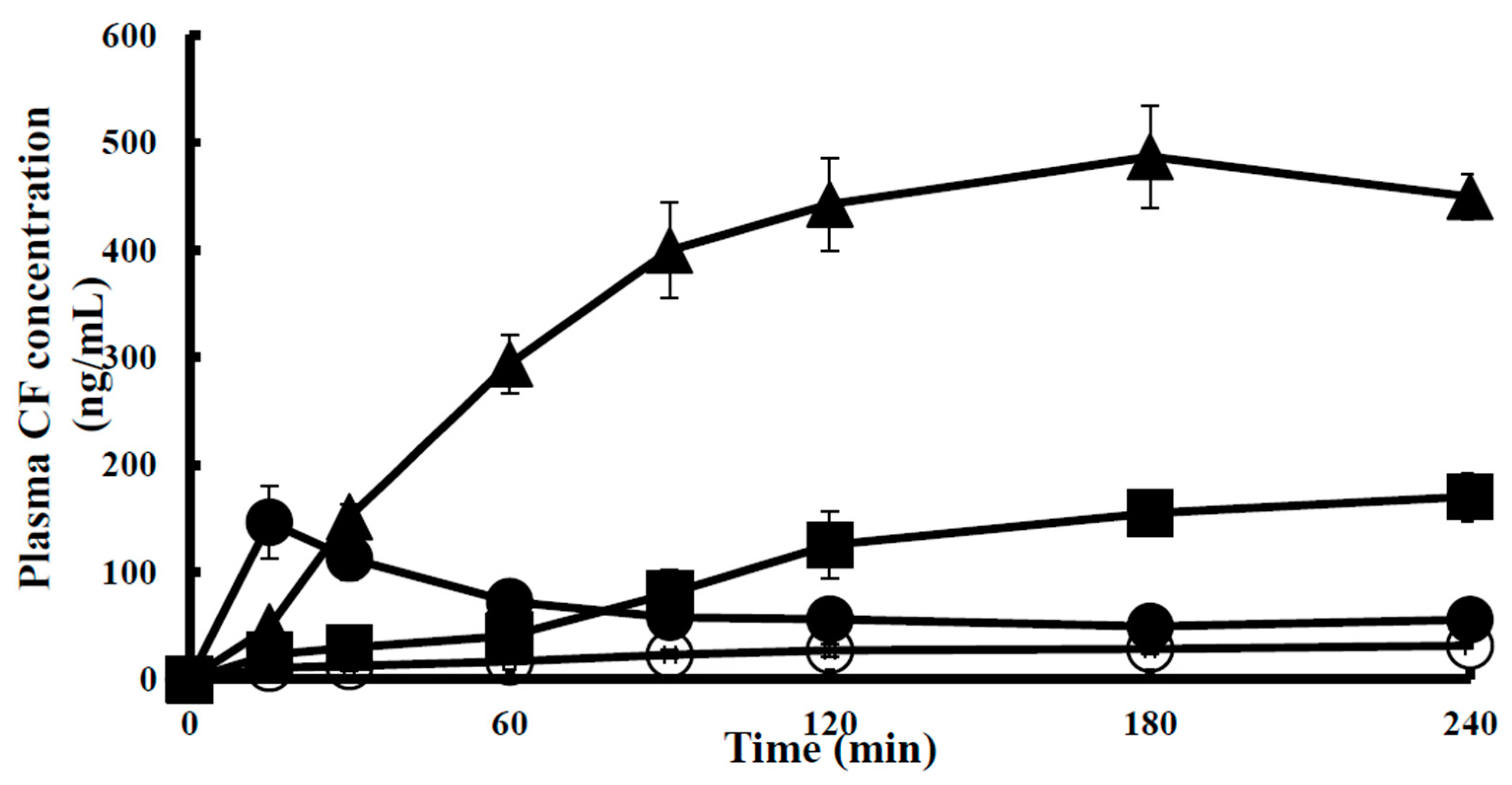
 ) 0.01% (v/v) SLG-30, (■) 0.025% (v/v) SLG-30, (▲) 0.05% (v/v) SLG-30, (●) 0.1% (v/v) SLG-30. (◆) 0.1% (w/v) sodium glycocholate, (□) 1% (w/v) sodium glycocholate, and (◆) 0.05% (w/v) sodium laurate.
) 0.01% (v/v) SLG-30, (■) 0.025% (v/v) SLG-30, (▲) 0.05% (v/v) SLG-30, (●) 0.1% (v/v) SLG-30. (◆) 0.1% (w/v) sodium glycocholate, (□) 1% (w/v) sodium glycocholate, and (◆) 0.05% (w/v) sodium laurate.
) 0.01% (v/v) SLG-30, (■) 0.025% (v/v) SLG-30, (▲) 0.05% (v/v) SLG-30, (●) 0.1% (v/v) SLG-30. (◆) 0.1% (w/v) sodium glycocholate, (□) 1% (w/v) sodium glycocholate, and (◆) 0.05% (w/v) sodium laurate.
) 0.01% (v/v) SLG-30, (■) 0.025% (v/v) SLG-30, (▲) 0.05% (v/v) SLG-30, (●) 0.1% (v/v) SLG-30. (◆) 0.1% (w/v) sodium glycocholate, (□) 1% (w/v) sodium glycocholate, and (◆) 0.05% (w/v) sodium laurate.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Absorption Enhancer | Conc. | Cmax (ng/mL) | Tmax (min) | AUC0→240 (ng·min/mL) | Enhancement Ratio |
|---|---|---|---|---|---|
| Control | - | - | 5450 ± 700 | - | |
| SLG-30 | 0.5% (v/v) | 487 ± 4 | 180 ± 0 | 87,200 ± 6000 ** | 16.0 |
| Sodium laurate | 0.5% (w/v) | 375 ± 2 | 15 ± 0 | 14,700 ± 1950 | 2.7 |
| Sodium glycocholate | 1% (w/v) | 172 ± 2 | 180 ± 0 | 24,500 ± 3700 * | 4.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alama, T.; Kusamori, K.; Morishita, M.; Katsumi, H.; Sakane, T.; Yamamoto, A. Mechanistic Studies on the Absorption-Enhancing Effects of Gemini Surfactant on the Intestinal Absorption of Poorly Absorbed Hydrophilic Drugs in Rats. Pharmaceutics 2019, 11, 170. https://doi.org/10.3390/pharmaceutics11040170
Alama T, Kusamori K, Morishita M, Katsumi H, Sakane T, Yamamoto A. Mechanistic Studies on the Absorption-Enhancing Effects of Gemini Surfactant on the Intestinal Absorption of Poorly Absorbed Hydrophilic Drugs in Rats. Pharmaceutics. 2019; 11(4):170. https://doi.org/10.3390/pharmaceutics11040170
Chicago/Turabian StyleAlama, Tammam, Kosuke Kusamori, Masaki Morishita, Hidemasa Katsumi, Toshiyasu Sakane, and Akira Yamamoto. 2019. "Mechanistic Studies on the Absorption-Enhancing Effects of Gemini Surfactant on the Intestinal Absorption of Poorly Absorbed Hydrophilic Drugs in Rats" Pharmaceutics 11, no. 4: 170. https://doi.org/10.3390/pharmaceutics11040170