An Effective Cationic Human Serum Albumin-Based Gene-Delivery Carrier Containing the Nuclear Localization Signal

Abstract
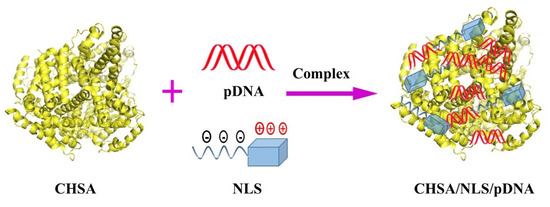
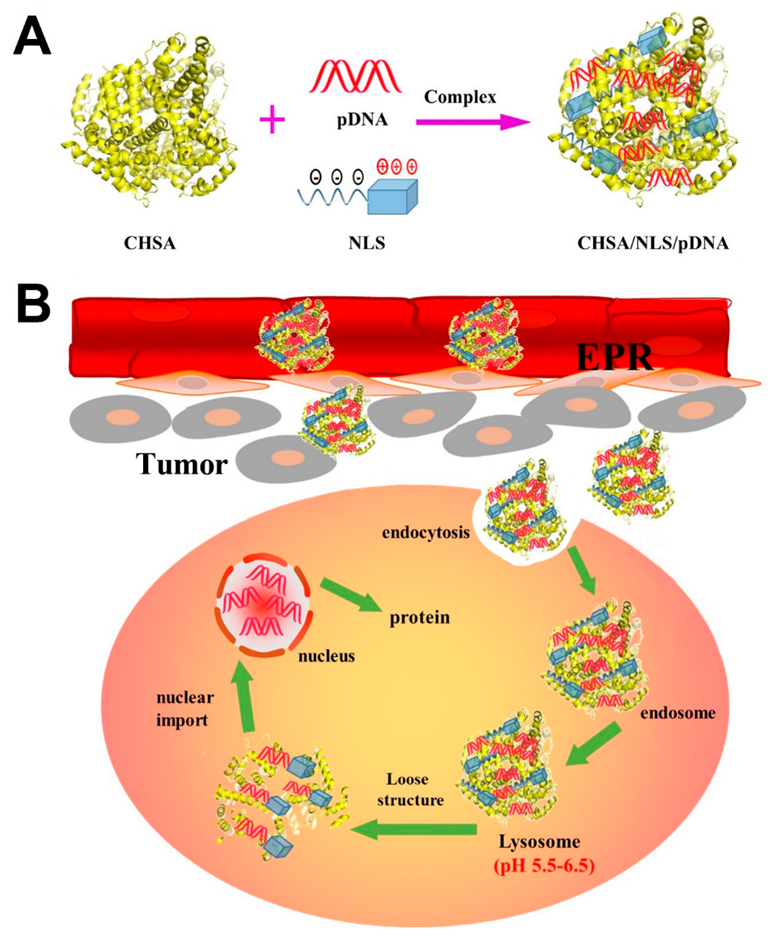
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation and Characterization of Cationic Human Serum Albumin
2.3. Preparation and Characterization of CHSA/NLS/pDNA Complexes
2.3.1. Preparation of CHSA/NLS/pDNA Complexes
2.3.2. Hoechst 33258 Intercalation Assay
2.3.3. Gel Retardation Assay
2.3.4. Particle Size and Zeta Potential Measurements and Morphological Analysis
2.3.5. Circular Dichroism Spectroscopy
2.4. MTT Assay
2.5. Immunogenicity Assay
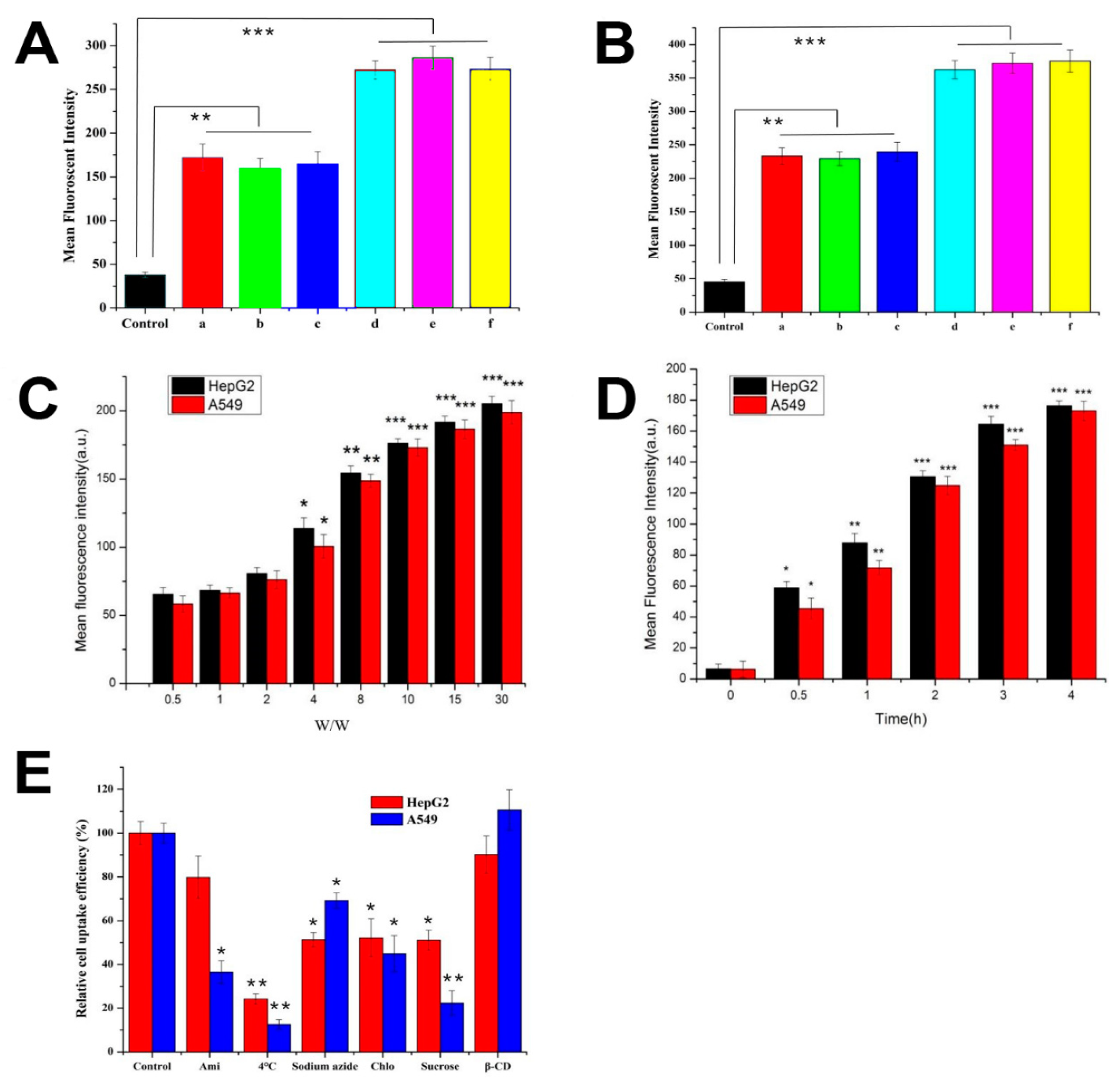
2.6. Cellular Uptake and Its Mechanisms
2.7. Gene Expression In Vitro
2.7.1. pEGFP-C1 Expression Study by LSCM
2.7.2. Luciferase Activity Assay
2.8. In Vivo Antitumor Activity
2.9. Statistics
3. Results and Discussion
3.1. Preparation and Characterization of Cationic Human Serum Albumin
3.2. Preparation and Characterization of CHSA/NLS/pDNA Complexes
3.2.1. Preparation of CHSA/NLS/pDNA Complexes
3.2.2. Hoechst 33258 Intercalation and Gel Retardation Assay
3.2.3. Particle Size and Zeta Potential Measurement
3.2.4. Morphology Characterization
3.2.5. Circular Dichroism Analysis
3.3. In Vitro Cytotoxicity
3.4. Immunogenicity Study
3.5. Cellular Uptake and Its Mechanisms
3.6. Gene Expression In Vitro
3.7. In Vivo Antitumor Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Foldvari, M.; Chen, D.W.; Nafissi, N.; Calderon, D.; Narsineni, L.; Rafiee, A. Non-viral gene therapy: Gains and challenges of non-invasive administration methods. J. Control. Release 2016, 240, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Guo, Y.; Xue, Y.; Niu, W.; Chen, M.; Ma, P.X.; Lei, B. Engineering multifunctional bioactive citric acid-based nanovectors for intrinsical targeted tumor imaging and specific siRNA gene delivery in vitro/in vivo. Biomaterials 2019, 199, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Ong, W.; Pinese, C.; Chew, S.Y. Scaffold-mediated sequential drug/gene delivery to promote nerve regeneration and remyelination following traumatic nerve injuries. Adv. Drug Deliv. Rev. 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Liu, X.; Zhu, D.; Wang, Y.; Zhang, Z.; Zhou, X.; Qin, N.; Chen, X.; Shen, Y. Nonviral cancer gene therapy: Delivery cascade and vector nanoproperty integration. Adv. Drug Deliv. Rev. 2017, 115, 115–154. [Google Scholar] [CrossRef] [PubMed]
- Kratz, F. Albumin as a drug carrier: Design of prodrugs, drug conjugates and nanoparticles. J. Control. Release 2008, 132, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Spänkuch, B.; Steinhauser, I.; Wartlick, H.; Kurunci-Csacsko, E.; Strebhardt, K.I.; Langer, K. Downregulation of Plk1 expression by receptor-mediated uptake of antisense oligonucleotide-loaded nanoparticles. Neoplasia 2008, 10, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Elzoghby, A.O.; Samy, W.M.; Elgindy, N.A. Albumin-based nanoparticles as potential controlled release drug delivery systems. J. Control. Release 2012, 157, 168–182. [Google Scholar] [CrossRef]
- Simões, S.; Slepushkin, V.; Pires, P.; Gaspar, R.; de Lima MC, P.; Düzgüneş, N. Human serum albumin enhances DNA transfection by lipoplexes and confers resistance to inhibition by serum. Biochim. Biophys. Acta 2000, 1463, 459–469. [Google Scholar] [CrossRef]
- Liu, N.; Qi, Y.H.; Cheng, C.T.; Yang, W.B.; Malhotra, A.; Zhou, Q. Potential of siRNA-albumin complex against cancer. Chem. Biol. Interact. 2018, 295, 93–96. [Google Scholar] [CrossRef]
- Piao, L.; Li, H.; Teng, L.; Yung, B.C.; Sugimoto, Y.; Brueggemeier, R.W.; Lee, R.J. Human serum albumin-coated lipid nanoparticles for delivery of siRNA to breast cancer. Nanomedicine 2013, 9, 122–129. [Google Scholar] [CrossRef]
- Loureiro, A.; Azoia, N.G.; Gomes, A.C.; Cavaco-Paulo, A. Albumin-based nanodevices as drug carriers. Curr. Pharm. Des. 2016, 22, 1371–1390. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.W.C.; Liu, Y.; Han, R.; Bai, Q.; Choi, C.H.J. Nano-cell interactions of non-cationic bionanomaterials. Acc. Chem. Res. 2019, 52, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Stephanie, R.; Hagen von, B.; Helga, R.W.; Jorg, K.; Klaus, L. Human serum albumin–polyethylenimine nanoparticles for gene delivery. J. Control. Release 2003, 92, 199–208. [Google Scholar]
- Salvatore, C.; Sante, D.G.; Elena, C.; Massimo, C. Serum albumin enhances polyethylenimine-mediated gene delivery to human respiratory epithelial cells. J. Gene Med. 2005, 7, 1555–1564. [Google Scholar]
- Jiin-Long, C.; Ssu-Wei, P.; Wei-Hsu, K.; Wei-Hsu, K.; Ching-Jan, W.; Teung-Wang, L.; Ming-Kung, Y.; Chiao-His, C. An efficient and low toxic human serum albumin conjugated polyethylenimine nano-sized complex for gene delivery. J. Nanopart. Res. 2014, 16, 2593. [Google Scholar]
- Soniat, M.; Chook, Y.M. Nuclear localization signals for four distinct karyopherin-beta nuclear import systems. Biochem. J. 2015, 468, 353–362. [Google Scholar] [CrossRef]
- Xu, Y.; Liang, W.; Qiu, Y.; Cespi, M.; Palmieri, G.F.; Mason, A.J.; Lam, J.K. Incorporation of a nuclear localization signal in pH responsive LAH4-L1 peptide enhances transfection and nuclear uptake of plasmid DNA. Mol. Pharm. 2016, 13, 3141–3152. [Google Scholar] [CrossRef]
- Sato, H.; Nakhaei, E.; Kawano, T.; Murata, M.; Kishimura, A.; Mori, T.; Katayama, Y. Ligand-mediated coating of liposomes with human serum albumin. Langmuir 2018, 34, 2324–2331. [Google Scholar] [CrossRef]
- Hoogenboezem, E.N.; Duvall, C.L. Harnessing albumin as a carrier for cancer therapies. Adv. Drug Deliv. Rev. 2018, 130, 73–89. [Google Scholar] [CrossRef]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved method for determining food protein degree of hydrolysis. J. Food Chem. Toxicol. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Martelli, L.; Ragazzi, E.; di Mario, F.; Martelli, M.; Castagliuolo, I.; Dal Maschio, M.; Palù, G.; Maschietto, M.; Scorzeto, M.; Vassanelli, S. A potential role for the vanilloid receptor TRPV1 in the therapeutic effect of curcumin in dinitrobenzene sulphonic acid-induced colitis in mice. Neurogastroenterol. Motil. 2007, 19, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Hu, Y.X.; Li, J.; Liu, Y.F.; Zhang, L.; Ai, H.X.; Liu, H.S. Probing the binding reaction of cytarabine to human serum albumin using multispectroscopic techniques with the aid of molecular docking. J. Photochem. Photobiol. B 2017, 173, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Parray, M.U.D.; Mir, M.U.H.; Dohare, N.; Maurya, N.; Khan, A.B.; Borse, M.S.; Patel, R. Effect of cationic gemini surfactant and its monomeric counterpart on the conformational stability and esterase activity of human serum albumin. J. Mol. Liq. 2018, 260, 65–77. [Google Scholar] [CrossRef]
- Tabassum, S.; Al-Asbahy, W.M.; Afzal, M.; Arjmand, F. Synthesis, characterization and interaction studies of copper based drug with Human Serum Albumin (HSA): Spectroscopic and molecular docking investigations. J. Photochem. Photobiol. B 2012, 114, 132–139. [Google Scholar] [CrossRef]
- Kandagal, P.B.; Ashoka, S.; Seetharamappa, J.; Shaikh, S.M.; Jadegoud, Y.; Ijare, O.B. Study of the interaction of an anticancer drug with human and bovine serum albumin: Spectroscopic approach. J. Pharm. Biomed. Anal. 2006, 41, 393–399. [Google Scholar] [CrossRef]
- Yin, H.; Kanasty, R.L.; Eltoukhy, A.A.; Vegas, A.J.; Dorkin, J.R.; Anderson, D.G. Non-viral vectors for gene-based therapy. Nat. Rev. Genet. 2014, 15, 541–555. [Google Scholar] [CrossRef]
- Li, W.; Xin, X.; Jing, S.; Zhang, X.; Chen, K.; Chen, D.; Hu, H. Organic metal complexes based on zoledronate–calcium: A potential pDNA delivery system. J. Mater. Chem. B 2017, 5, 1601–1610. [Google Scholar] [CrossRef]
- Guo, Z.; Kong, Z.; Wei, Y.; Li, H.; Wang, Y.; Huang, A.; Ma, L. Effects of gene carrier polyethyleneimines on the structure and binding capability of bovine serum albumin. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2017, 173, 783–791. [Google Scholar] [CrossRef]
- Remoli, M.E.; Giacomini, E.; Lutfalla, G.; Dondi, E.; Orefici, G.; Battistini, A.; Uze, G.; Pellegrini, S.; Coccia, E.M. Selective expression of Type I IFN genes in human dendritic cells infected with mycobacterium tuberculosis. J. Immunol. 2002, 169, 366–374. [Google Scholar] [CrossRef]
- Vignali, D.A.; Kuchroo, V.K. IL-12 family cytokines: Immunological playmakers. Nat. Immunol. 2012, 13, 722–728. [Google Scholar] [CrossRef]
- Xiang, S.; Tong, H.; Shi, Q.; Fernandes, J.C.; Jin, T.; Dai, K.; Zhang, X. Uptake mechanisms of non-viral gene delivery. J. Control. Release 2012, 158, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; De Maziere, A.; Iaea, D.B.; Arthur, C.P.; Klumperman, J.; Ciferri, C.; Hannoush, R.M. Visualizing the cellular route of entry of a cystine-knot peptide with Xfect transfection reagent by electron microscopy. Sci. Rep. 2019, 9, 6907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arwyn, T.J. Macropinocytosis: Searching for an endocytic identity and a role in the uptake of cell penetrating peptides. J. Cell. Mol. Med. 2007, 11, 670–684. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | HSA | CHSA-1 | CHSA-2 | CHSA-3 | CHSA-4 | |
|---|---|---|---|---|---|---|
| Character | ||||||
| pI | 5 | 7 | 8 | 9 | 10 | |
| FAG | – | 22.09 ± 3.52 | 32.71 ± 1.37 | 42.51 ± 3.11 | 57.10 ± 2.49 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, G.; Song, B.; Zhang, J.; Chen, K.; Hu, H.; Wang, M.; Chen, D. An Effective Cationic Human Serum Albumin-Based Gene-Delivery Carrier Containing the Nuclear Localization Signal. Pharmaceutics 2019, 11, 608. https://doi.org/10.3390/pharmaceutics11110608
Guan G, Song B, Zhang J, Chen K, Hu H, Wang M, Chen D. An Effective Cationic Human Serum Albumin-Based Gene-Delivery Carrier Containing the Nuclear Localization Signal. Pharmaceutics. 2019; 11(11):608. https://doi.org/10.3390/pharmaceutics11110608
Chicago/Turabian StyleGuan, Guannan, Baohui Song, Jie Zhang, Kang Chen, Haiyang Hu, Mingyue Wang, and Dawei Chen. 2019. "An Effective Cationic Human Serum Albumin-Based Gene-Delivery Carrier Containing the Nuclear Localization Signal" Pharmaceutics 11, no. 11: 608. https://doi.org/10.3390/pharmaceutics11110608