
The Influence of Organic Vanadium Complexes on an Antioxidant Profile in Adipose Tissue in Wistar Rats

 ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
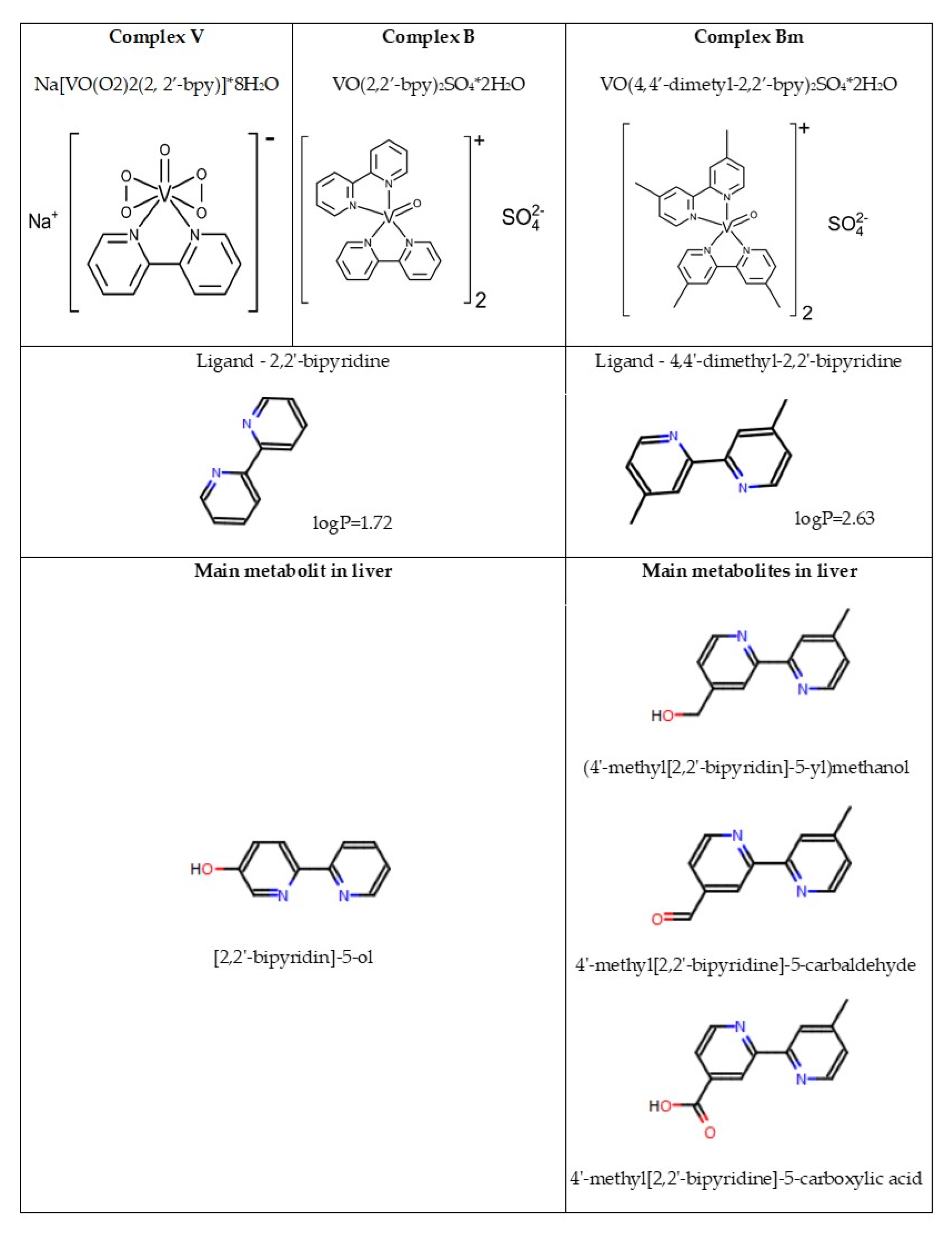
2.2. Synthesis of Vanadium Complexes
2.3. Animals
2.4. Sample Collection and Analysis
2.5. Measurement of the Vanadium Content in Adipose Tissue
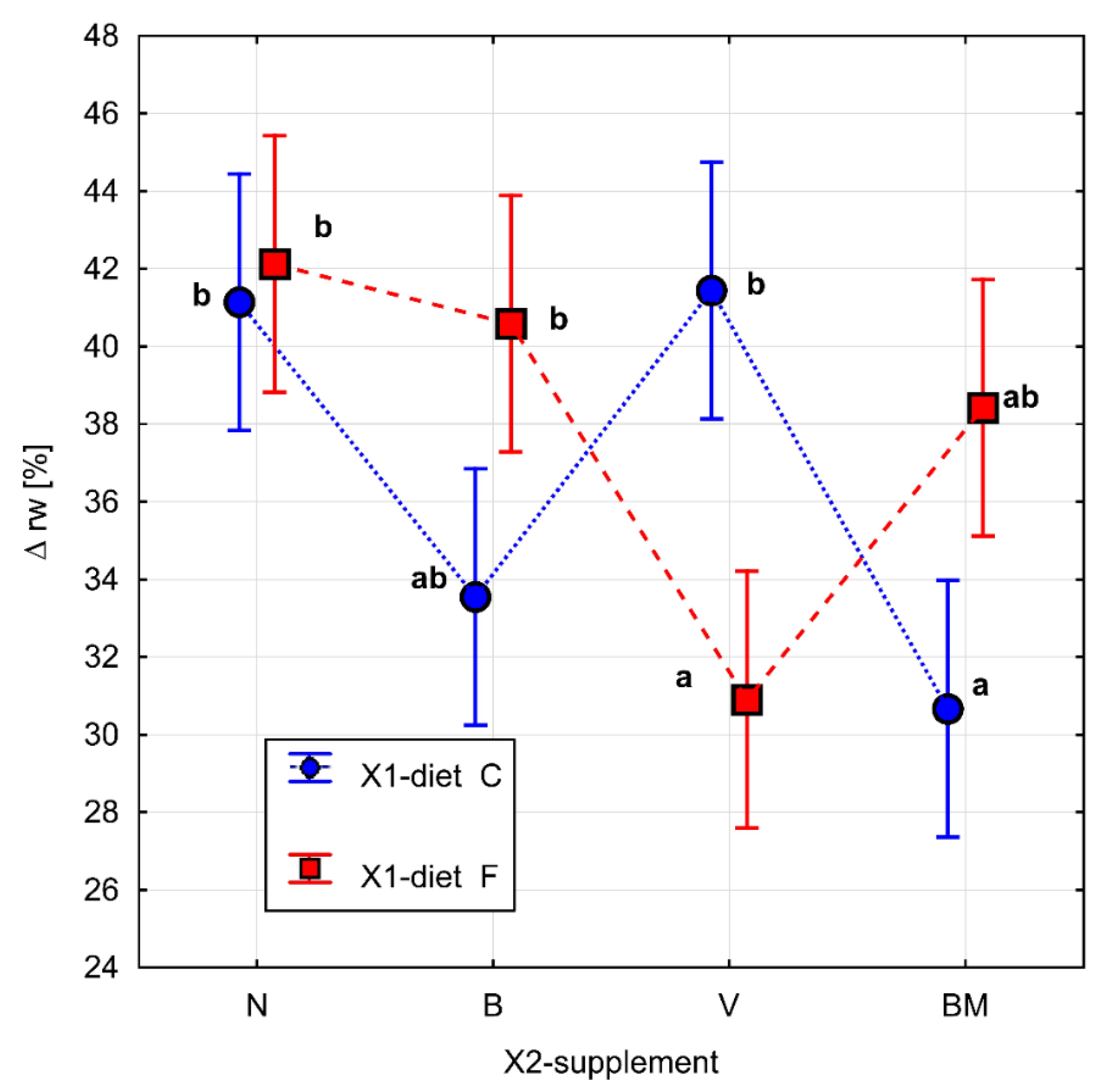
2.6. Relative Change of Rat Weight
2.7. Basal Metabolic Rate
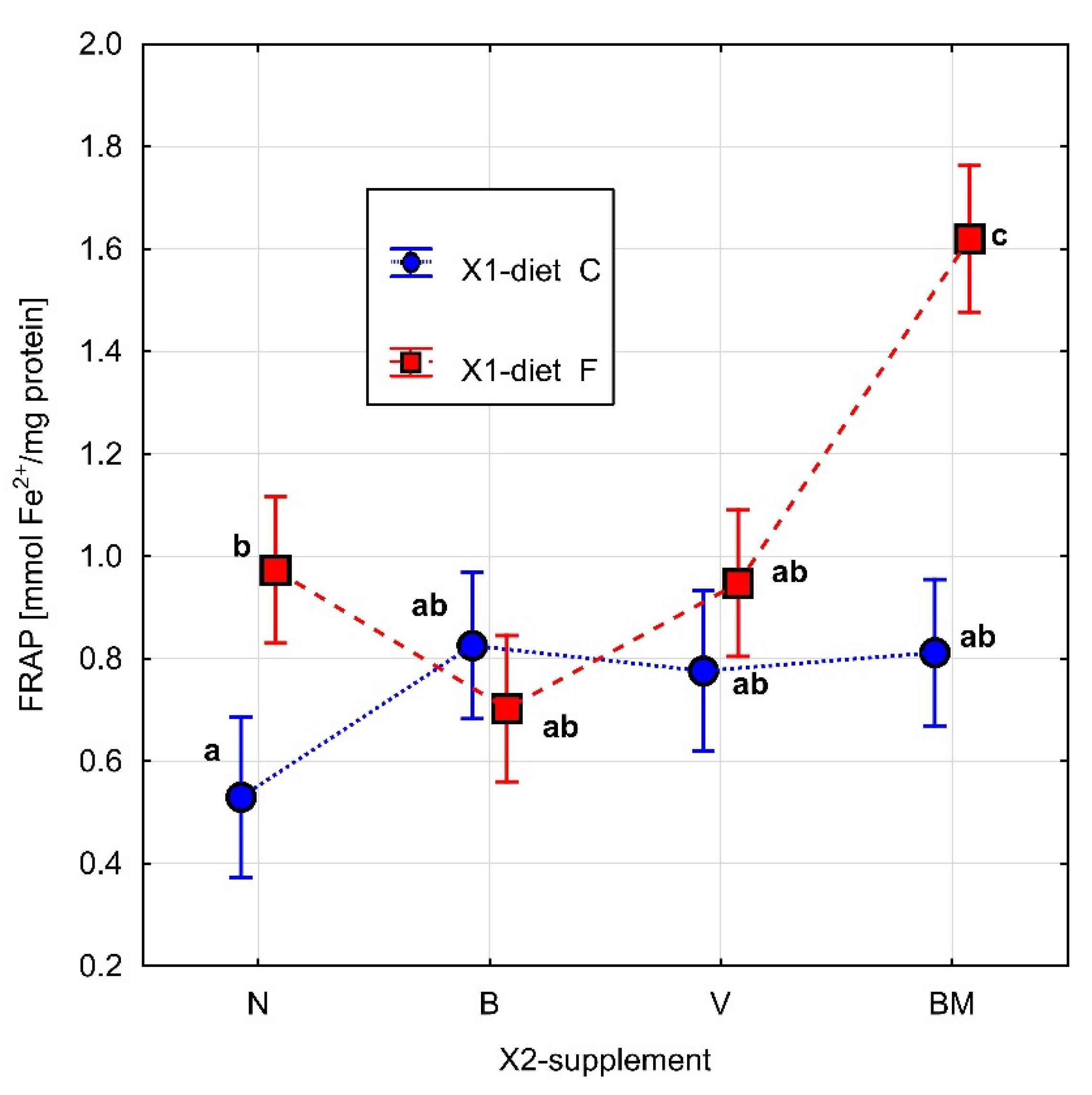
2.8. Measurement of the Total Antioxidant Capacity—Ferric Reducing Antioxidant Power (FRAP)
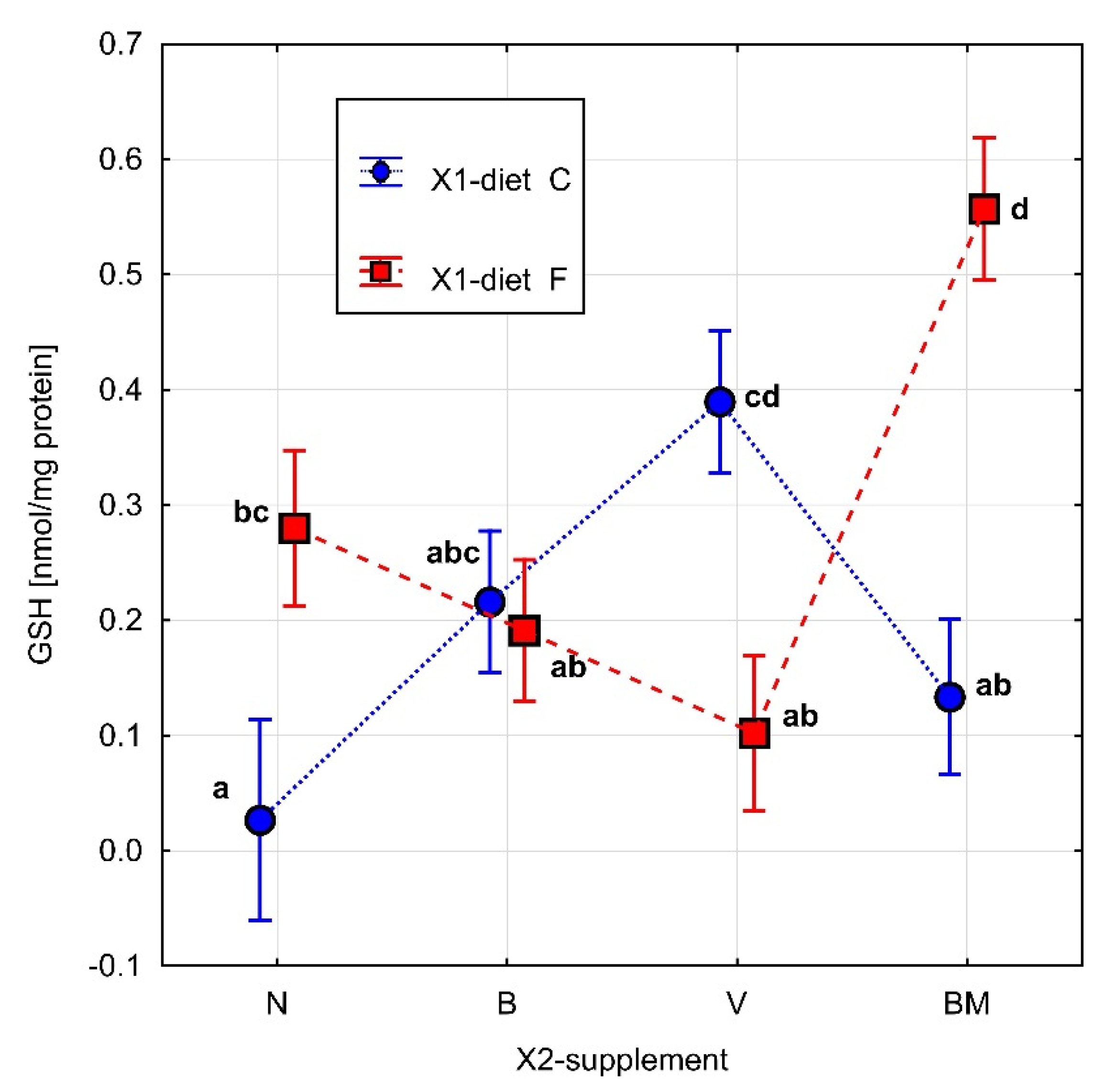
2.9. Measurement of Glutathione (GSH) Concentration
2.10. Measurement of Catalase (CAT) Activity
2.11. Measurement of Superoxide Dismutase (SOD) Activity
2.12. Statistical Analysis
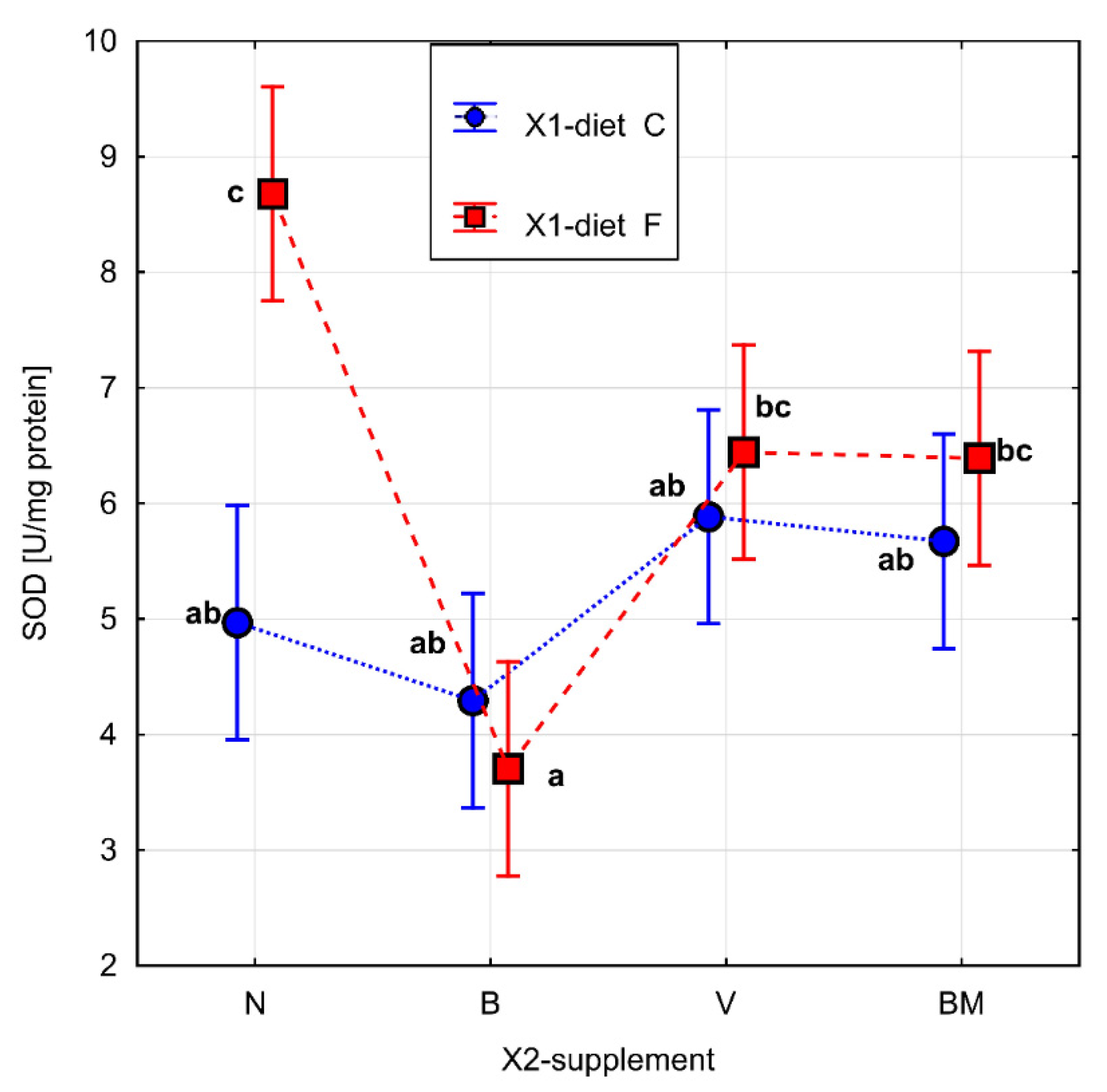
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| V complex | sodium (2,2′-bipyridine)oxidobisperoxovanadate (V) octahydrate |
| B complex | bis(2,2′-bipyridine)oxidovanadium(IV) sulfate dehydrate |
| BM complex | bis(4,4′-dimethyl-2,2′-bipyridine)oxidovanadium (IV) sulfate dehydrate |
| CN | standard diet without additives |
| CV | V complex with standard diet |
| CB | B complex with standard diet |
| CBM | BM complex with standard diet |
| FN | high-fat diet without additives |
| FV | V complex with high-fat diet |
| FB | B complex with high-fat diet |
| FBM | BM complex with high-fat diet |
| ∆rw | relative change in rat weight |
References
- Arroyo-Johnson, C.; Mincey, K.D. Obesity Epidemiology Worldwide. Gastroenterol. Clin. 2016, 45, 571–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agha, M.; Agha, R. The rising prevalence of obesity: Part A: Impact on public health. Int. J. Surg. Oncol. 2017, 2, e17. [Google Scholar] [CrossRef] [Green Version]
- Savini, I.; Catani, M.V.; Evangelista, D.; Gasperi, V.; Avigliano, L. Obesity-associated oxidative stress: Strategies finalized to improve redox state. Int. J. Mol. Sci. 2013, 14, 10497–10538. [Google Scholar] [CrossRef] [Green Version]
- Chrysohoou, C.; Panagiotakos, D.B.; Pitsavos, C.; Skoumas, I.; Papademetriou, L.; Economou, M.; Stefanadis, C. The implication of obesity on total antioxidant capacity in apparently healthy men and women: The ATTICA study. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 590–597. [Google Scholar] [CrossRef]
- Patel, C.; Ghanim, H.; Ravishankar, S.; Chang, L.S.; Viswanathan, P.; Mohanty, P.; Dandona, P. Prolonged reactive oxygen species generation and nuclear factor-κB activation after a high-fat, high-carbohydrate meal in the obese. J. Clin. Endocrinol. Metab. 2007, 92, 4476–4479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Câmara, D.A.D.; Shibli, J.A.; Müller, E.A.; De-Sá-Junior, P.L.; Porcacchia, A.S.; Blay, A.; Lizier, N.F. Adipose tissue-derived stem cells: The biologic BASIS and future directions for tissue engineering. Materials 2020, 13, 3210. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, A.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.I.; Naz, L.; Yasmeen, G. Obesity: An independent risk factor for systemic oxidative stress. Pak. J. Pharm. Sci. 2006, 19, 62–65. [Google Scholar] [PubMed]
- Duvnjak, M.; Lerotić, I.; Baršić, N.; Tomašić, V.; Jukić, L.V.; Velagić, V. Pathogenesis and management issues for non-alcoholic fatty liver disease. World J. Gastroenterol. 2007, 13, 4539–4550. [Google Scholar] [CrossRef]
- Fonseca-Alaniz, M.H.; Takada, J.; Alonso-Vale, M.I.; Lima, F.B. Adipose tissue as an endocrine organ: From theory to practice. J. Pediatr. (Rio. J). 2007, 83, S192–203. [Google Scholar] [CrossRef]
- Alcalá, M.; Sánchez-Vera, I.; Sevillano, J.; Herrero, L.; Serra, D.; Ramos, M.P.; Viana, M. Vitamin E reduces adipose tissue fibrosis, inflammation, and oxidative stress and improves metabolic profile in obesity. Obesity 2015, 23, 1598–1606. [Google Scholar] [CrossRef]
- Farhangi, M.A.; Mesgari-Abbasi, M.; Hajiluian, G.; Nameni, G.; Shahabi, P. Adipose Tissue Inflammation and Oxidative Stress: The Ameliorative Effects of Vitamin, D. Inflammation 2017, 40, 1688–1697. [Google Scholar] [CrossRef] [PubMed]
- Sohet, F.M.; Neyrinck, A.M.; Pachikian, B.D.; de Backer, F.C.; Bindels, L.B.; Niklowitz, P.; Menke, T.; Cani, P.D.; Delzenne, N.M. Coenzyme Q10 supplementation lowers hepatic oxidative stress and inflammation associated with diet-induced obesity in mice. Biochem. Pharmacol. 2009, 78, 1391–1400. [Google Scholar] [CrossRef] [Green Version]
- Pillai, S.I.; Subramanian, S.P.; Kandaswamy, M. Evaluation of antioxidant efficacy of vanadium-3-hydroxyflavone complex in streptozotocin-diabetic rats. Chem. Biol. Interact. 2013, 204, 67–74. [Google Scholar] [CrossRef]
- Tunali, S.; Yanardag, R. Protective effect of vanadyl sulfate on skin injury in streptozotocin- induced diabetic rats. Hum. Exp. Toxicol. 2013, 32, 1206–1212. [Google Scholar] [CrossRef]
- Sakurai, H.; Kojima, Y.; Yoshikawa, Y.; Kawabe, K.; Yasui, H. Antidiabetic vanadium(IV) and zinc(II) complexes. Coord. Chem. Rev. 2002, 226, 187–198. [Google Scholar] [CrossRef]
- Rangel, M.; Tamura, A.; Fukushima, C.; Sakurai, H. In vitro study of the insulin-like action of vanadyl-pyrone and -pyridinone complexes with a VO(O4) coordination mode. J. Biol. Inorg. Chem. 2001, 6, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Rehder, D.; Costa Pessoa, J.; Geraldes, C.F.G.C.; Castro, M.M.C.A.; Kabanos, T.; Kiss, T.; Meier, B.; Micera, G.; Pettersson, L.; Rangel, M.; et al. In vitro study of the insulin-mimetic behaviour of vanadium(IV, V) coordination compounds. J. Biol. Inorg. Chem. 2002, 7, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Francik, R.; Kryczyk-Kozioł, J.; Francik, S.; Gryboś, R.; Krośniak, M. Bis(4,4′-dimethyl-2,2′-bipyridine)oxidovanadium(IV) Sulfate Dehydrate: Potential Candidate for Controlling Lipid Metabolism? Biomed Res. Int. 2017, 2017, 6950516. [Google Scholar] [CrossRef]
- Przybylski, W.; Gryboś, R.; Rehder, D.; Ebel, M.; Grzywa, M.; Łasocha, W.; Lewiński, K.; Szklarzewicz, J.T. Role of the alkali metal ion and hydrogen bonds in M[VO(O2)2bpy] · nH2O (M = Li+, Na+, K+ and Rb+) and Cs[VO(O2)2bpy] · H2O2 complexes: The X-ray crystal structures and spectroscopic properties. Polyhedron 2009, 28, 1429–1436. [Google Scholar] [CrossRef]
- Krośniak, M.; Gawlik, M.; Gryboś, R. Effect of vanadium complexes and insulin administered simultaneously for oxidative stress in STZ diabetic rats. Bull. Vet. Inst. Pulawy 2009, 53, 535–540. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ellman, G.L. A colorimetric method for determining low concentrations of mercaptans. Arch. Biochem. Biophys. 1958, 74, 443–450. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Oberley, L.W.; Li, Y. A simple method for clinical assay of superoxide dismutase. Clin. Chem. 1988, 34, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Krosniak, M.; Zachwieja, Z.; Filipek, B.; Zygmunt, M.; Grybos, R. Effect of oxovanadium(IV) complexes on nondiabetic and streptozotocin-diabetic rats. Arch. Pharm. 2001, 334, 388–392. [Google Scholar] [CrossRef]
- Srivastava, A.K. Anti-diabetic and toxic effects of vanadium compounds. Mol. Cell. Biochem. 2000, 206, 177–182. [Google Scholar] [CrossRef]
- Crans, D.C. Chemistry and insulin-like properties of vanadium(IV) and vanadium(V) compounds. J. Inorg. Biochem. 2000, 80, 123–131. [Google Scholar] [CrossRef]
- Fugono, J.; Yasui, H.; Sakurai, H. Improvement of diabetic states in streptozotocin-induced type 1 diabetic rats by vanadyl sulfate in enteric-coated capsules. J. Pharm. Pharmacol. 2005, 57, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Li, M.; Ding, W. Effect of vanadate on gene expression of the insulin signaling pathway in skeletal muscle of streptozotocin-induced diabetic rats. J. Biol. Inorg. Chem. 2007, 12, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.H.; Lichter, J.; LeBel, C.; Scaife, M.C.; McNeill, J.H.; Orvig, C. Vanadium treatment of type 2 diabetes: A view to the future. J. Inorg. Biochem. 2009, 103, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Soveid, M.; Dehghani, G.A.; Omrani, G.R. Long- term efficacy and safety of vanadium in the treatment of type 1 diabetes. Arch. Iran. Med. 2013, 16, 408–411. [Google Scholar] [PubMed]
- Dong, Y.; Narla, R.K.; Sudbeck, E.; Uckun, F.M. Synthesis, X-ray structure, and anti-leukemic activity of oxovanadium(IV) complexes. J. Inorg. Biochem. 2000, 78, 321–330. [Google Scholar] [CrossRef]
- Narla, R.K.; Dong, Y.; D’Cruz, O.J.; Navara, C.; Uckun, F.M. Bis(4,7-dimethyl-1,10-phenanthroline) sulfatooxovanadium(IV) as a novel apoptosis-inducing anticancer agent. Clin. Cancer Res. 2000, 6, 1546–1556. [Google Scholar]
- Soares, S.S.; Martins, H.; Aureliano, M. Vanadium distribution following decavanadate administration. Arch. Environ. Contam. Toxicol. 2006, 50, 60–64. [Google Scholar] [CrossRef]
- Urban, J.; Antonowicz-Juchniewicz, J.; Andrzejak, R. Vanadium: Threat and hope [Wanad--zagrozenia i nadzieje.]. Med. Pr. 2001, 52, 125–133. [Google Scholar]
- Mukherjee, B.; Patra, B.; Mahapatra, S.; Banerjee, P.; Tiwari, A.; Chatterjee, M. Vanadium—An element of atypical biological significance. Toxicol. Lett. 2004, 150, 135–143. [Google Scholar] [CrossRef]
- Sanna, D.; Micera, G.; Garribba, E. New developments in the comprehension of the biotransformation and transport of insulin-enhancing vanadium compounds in the blood serum. Inorg. Chem. 2010, 49, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Sanna, D.; Buglyó, P.; Tomaz, A.I.; Pessoa, J.C.; Borović, S.; Micera, G.; Garribba, E. VIVO and CuII complexation by ligands based on pyridine nitrogen donors. Dalt. Trans. 2012, 41, 12824–12838. [Google Scholar] [CrossRef] [PubMed]
- Triantafillou, G.D.; Tolis, E.I.; Terzis, A.; Deligiannakis, Y.; Raptopoulou, C.P.; Sigalas, M.P.; Kabanos, T.A. Monomeric Oxovanadium(IV) Compounds of the General Formula cis-[V IV(=O)(X)(LNN)2]+/0 {X = OH−, Cl−, SO42− and LNN = 2,2′-Bipyridine (Bipy) or 4,4′-Disubstituted Bipy}. Inorg. Chem. 2004, 43, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Sanna, D.; Ugone, V.; Micera, G.; Buglyó, P.; Bíró, L.; Garribba, E. Speciation in human blood of Metvan, a vanadium based potential anti-tumor drug. Dalt. Trans. 2017, 46, 8950–8967. [Google Scholar] [CrossRef]
- Le, M.; Rathje, O.; Levina, A.; Lay, P.A. High cytotoxicity of vanadium(IV) complexes with 1,10-phenanthroline and related ligands is due to decomposition in cell culture medium. J. Biol. Inorg. Chem. 2017, 22, 663–672. [Google Scholar] [CrossRef]
- Levina, A.; Crans, D.C.; Lay, P.A. Speciation of metal drugs, supplements and toxins in media and bodily fluids controls in vitro activities. Coord. Chem. Rev. 2017, 352, 473–498. [Google Scholar] [CrossRef]
- Tsave, O.; Halevas, E.; Yavropoulou, M.P.; Yovos, E.; Hatzidimitriou, A.; Psycharis, V.; Ypsilantis, K.; Stathi, P.; Salifoglou, A. V(V)-Schiff base species induce adipogenesis through structure-specific influence of genetic targets. New J. Chem. 2019, 43, 17872–17890. [Google Scholar] [CrossRef]
- Liu, Y.; Jie, X.; Guo, Y.; Zhang, X.; Wang, J.; Xue, C. Green Synthesis of Oxovanadium(IV)/chitosan Nanocomposites and Its Ameliorative Effect on Hyperglycemia, Insulin Resistance, and Oxidative Stress. Biol. Trace Elem. Res. 2016, 169, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, J.; Guo, Y.; Xue, Y.; Wang, J.; Xue, C. Ameliorative effect of vanadyl(IV)-ascorbate complex on high-fat high-sucrose diet-induced hyperglycemia, insulin resistance, and oxidative stress in mice. J. Trace Elem. Med. Biol. 2015, 32, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yan, L.; Kim, S.M. Vanadium-protein complex inhibits human adipocyte differentiation through the activation of β-catenin and LKB1/AMPK signaling pathway. PLoS ONE 2020, 15, e0239547. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.P.; Grune, T.; Speckmann, B. The two faces of reactive oxygen species (ROS) in adipocyte function and dysfunction. Biol. Chem. 2016, 397, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Huang, Y.; Liu, F.; Zhang, F.; Ding, W. Vanadium(IV)-chlorodipicolinate inhibits 3T3-L1 preadipocyte adipogenesis by activating LKB1/AMPK signaling pathway. J. Inorg. Biochem. 2016, 162, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, H.; Xu, J.; Guo, Y.; Xue, Y.; Wang, J.; Xue, C. Vanadium-binding protein from vanadium-enriched sea cucumber Apostichopus japonicus inhibits adipocyte differentiation through activating WNT/β-catenin pathway. J. Funct. Foods 2015, 17, 504–513. [Google Scholar] [CrossRef]
- Dikalov, S. Cross talk between mitochondria and NADPH oxidases. Free Radic. Biol. Med. 2011, 51, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-H.; Wang, C.-C.; Huang, H.-C.; Wei, Y.-H. Mitochondrial dysfunction leads to impairment of insulin sensitivity and adiponectin secretion in adipocytes. FEBS J. 2013, 280, 1039–1050. [Google Scholar] [CrossRef]
- Wang, L.; Xu, S.; Lee, J.-E.; Baldridge, A.; Grullon, S.; Peng, W.; Ge, K. Histone H3K9 methyltransferase G9a represses PPARγ expression and adipogenesis. EMBO J. 2013, 32, 45–59. [Google Scholar] [CrossRef]
- Charradi, K.; Elkahoui, S.; Limam, F.; Aouani, E. High-fat diet induced an oxidative stress in white adipose tissue and disturbed plasma transition metals in rat: Prevention by grape seed and skin extract. J. Physiol. Sci. 2013, 63, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Flieger, J.; Flieger, W.; Baj, J.; Maciejewski, R. Antioxidants: Classification, natural sources, activity/capacity measurements, and usefulness for the synthesis of nanoparticles. Materials 2021, 14, 4135. [Google Scholar] [CrossRef] [PubMed]
- Keaney, J.F., Jr.; Larson, M.G.; Vasan, R.S.; Wilson, P.W.F.; Lipinska, I.; Corey, D.; Massaro, J.M.; Sutherland, P.; Vita, J.A.; Benjamin, E.J. Obesity and systemic oxidative stress: Clinical correlates of oxidative stress in the Framingham study. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 434–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinkov, A.A.; Sinitskii, A.I.; Popova, E.V.; Nemereshina, O.N.; Gatiatulina, E.R.; Skalnaya, M.G.; Skalny, A.V.; Nikonorov, A.A. Alteration of local adipose tissue trace element homeostasis as a possible mechanism of obesity-related insulin resistance. Med. Hypotheses 2015, 85, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Chen, D.; Zhang, F.; Willsky, G.R.; Crans, D.C.; Ding, W. Effects of vanadium (III, IV, V)-chlorodipicolinate on glycolysis and antioxidant status in the liver of STZ-induced diabetic rats. J. Inorg. Biochem. 2014, 136, 47–56. [Google Scholar] [CrossRef]
- Tinkov, A.A.; Gatiatulina, E.R.; Popova, E.V.; Polyakova, V.S.; Skalnaya, A.A.; Agletdinov, E.F.; Nikonorov, A.A.; Skalny, A. V Early High-Fat Feeding Induces Alteration of Trace Element Content in Tissues of Juvenile Male Wistar Rats. Biol. Trace Elem. Res. 2017, 175, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Tinkov, A.A.; Popova, E.V.; Polyakova, V.S.; Kwan, O.V.; Skalny, A.V.; Nikonorov, A.A. Adipose tissue chromium and vanadium disbalance in high-fat fed Wistar rats. J. Trace Elem. Med. Biol. 2015, 29, 176–181. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Group | Water Intake ± SD [mL/Day/Animal] | Feed Intake ± SD [g/Day/Animal] | BWG ± SD [g/Animal] | BMR ± SD [kcal/g Body Weight Gain] | VI ± SD [mg/kg/day] | VCAT ± SD [μg/kg of Dry Adipose Tissue] |
|---|---|---|---|---|---|---|
| CN | 27.0 ± 4.9 a | 18.4 ± 4.9 b | 105.3 ± 13.9 a | 21.0 ± 0.3 a | ND | 0.81 ± 0.09 a |
| CV | 30.3 ± 5.2 a | 17.3 ± 1.9 b | 96.8 ± 18.3 a | 22.6 ± 0.2 b | 2.25 ± 0.2 a | 3.48 ± 0.37 b |
| CB | 26.6 ± 2.5 a | 17.7 ± 3.6 b | 83.5 ± 15.4 ab | 26.1 ± 0.6 c | 1.99 ± 0.2 a | 3.67 ± 0.22 b |
| CBM | 28.0 ± 5.2 a | 17.9 ± 1.5 b | 76.8 ± 15.1 b | 27.6 ± 0.1 c | 1.80 ± 0.2 a | 4.02 ± 0.31 b |
| FN | 25.6 ± 3.6 a | 13.5 ± 0.8 a | 109.7 ± 21.5 a | 19.8 ± 1.2 a | ND | 0.78 ± 0.12 a |
| FV | 24.5 ± 3.8 a | 11.9 ± 2.2 a | 78.3 ± 14.1 b | 24.7 ± 1.0 b | 2.25 ± 0.2 a | 3.55 ± 0.29 b |
| FB | 29.2 ± 8.1 a | 12.3 ± 2.4 a | 95.6 ± 21.1 a | 23.1 ± 2.9 b | 1.99 ± 0.2 a | 3.53 ± 0.31 b |
| FBM | 29.6 ± 6.6 a | 13.3 ± 2.4 a | 94.5 ± 24.6 a | 24.6 ± 1.0 b | 1.80 ± 0.2 a | 4.19 ± 0.30 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francik, R.; Kryczyk-Kozioł, J.; Krośniak, M.; Francik, S.; Hebda, T.; Pedryc, N.; Knapczyk, A.; Berköz, M.; Ślipek, Z. The Influence of Organic Vanadium Complexes on an Antioxidant Profile in Adipose Tissue in Wistar Rats. Materials 2022, 15, 1952. https://doi.org/10.3390/ma15051952
Francik R, Kryczyk-Kozioł J, Krośniak M, Francik S, Hebda T, Pedryc N, Knapczyk A, Berköz M, Ślipek Z. The Influence of Organic Vanadium Complexes on an Antioxidant Profile in Adipose Tissue in Wistar Rats. Materials. 2022; 15(5):1952. https://doi.org/10.3390/ma15051952
Chicago/Turabian StyleFrancik, Renata, Jadwiga Kryczyk-Kozioł, Mirosław Krośniak, Sławomir Francik, Tomasz Hebda, Norbert Pedryc, Adrian Knapczyk, Mehmet Berköz, and Zbigniew Ślipek. 2022. "The Influence of Organic Vanadium Complexes on an Antioxidant Profile in Adipose Tissue in Wistar Rats" Materials 15, no. 5: 1952. https://doi.org/10.3390/ma15051952