
Rhodamine-Tagged Polymethacrylate Dyes as Alternative Tools for Analysis of Plant Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Physicochemical Characterization
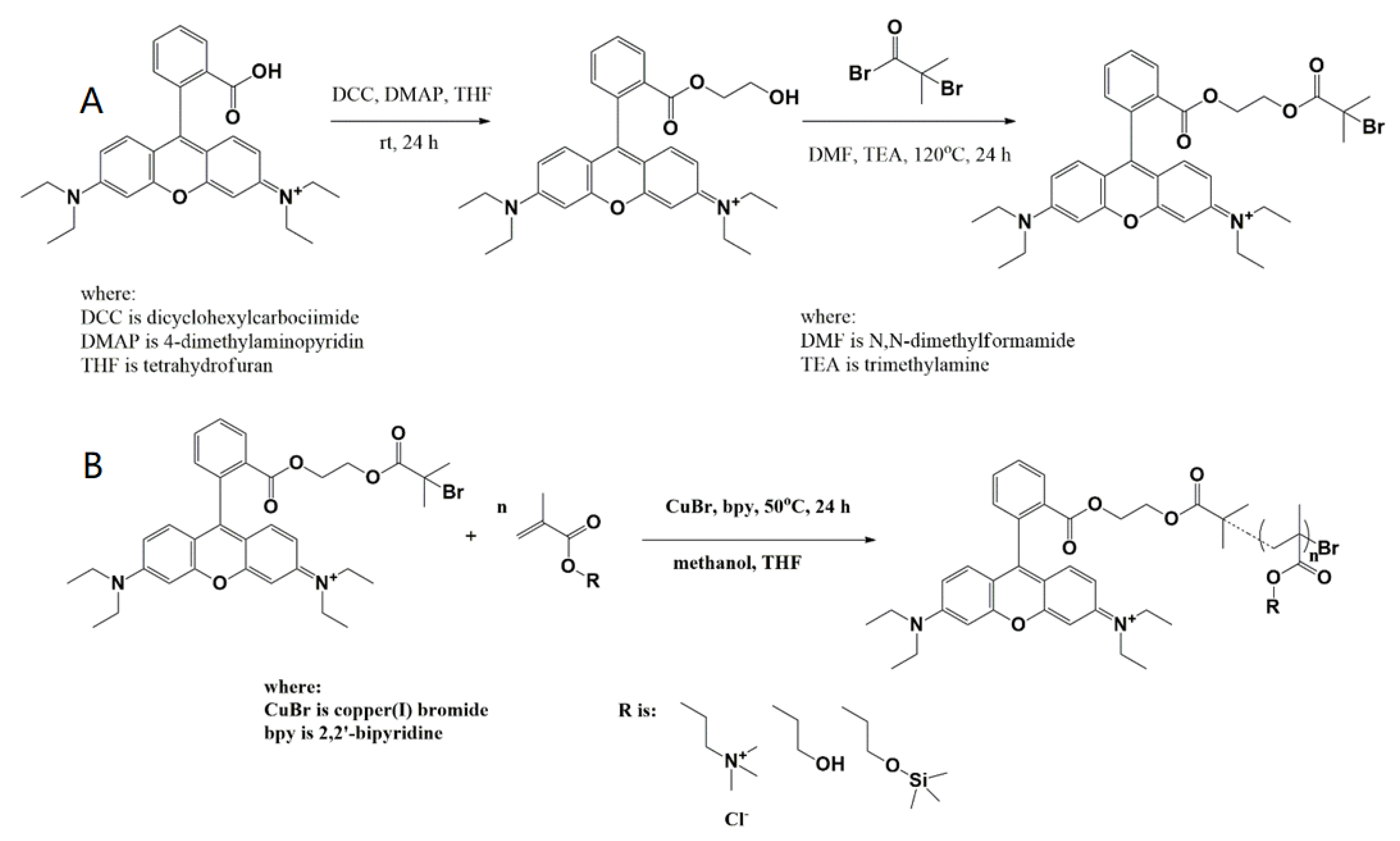
2.3. Preparation of RhB Based ATRP Initiator
2.4. General Method for RhB-Labelled Polymer Synthesis
2.5. Plant Material
2.6. Fluorochrome and Staining Procedure
2.7. Microscopical Observation
2.8. Cell Viability Detection
3. Results and Discussion
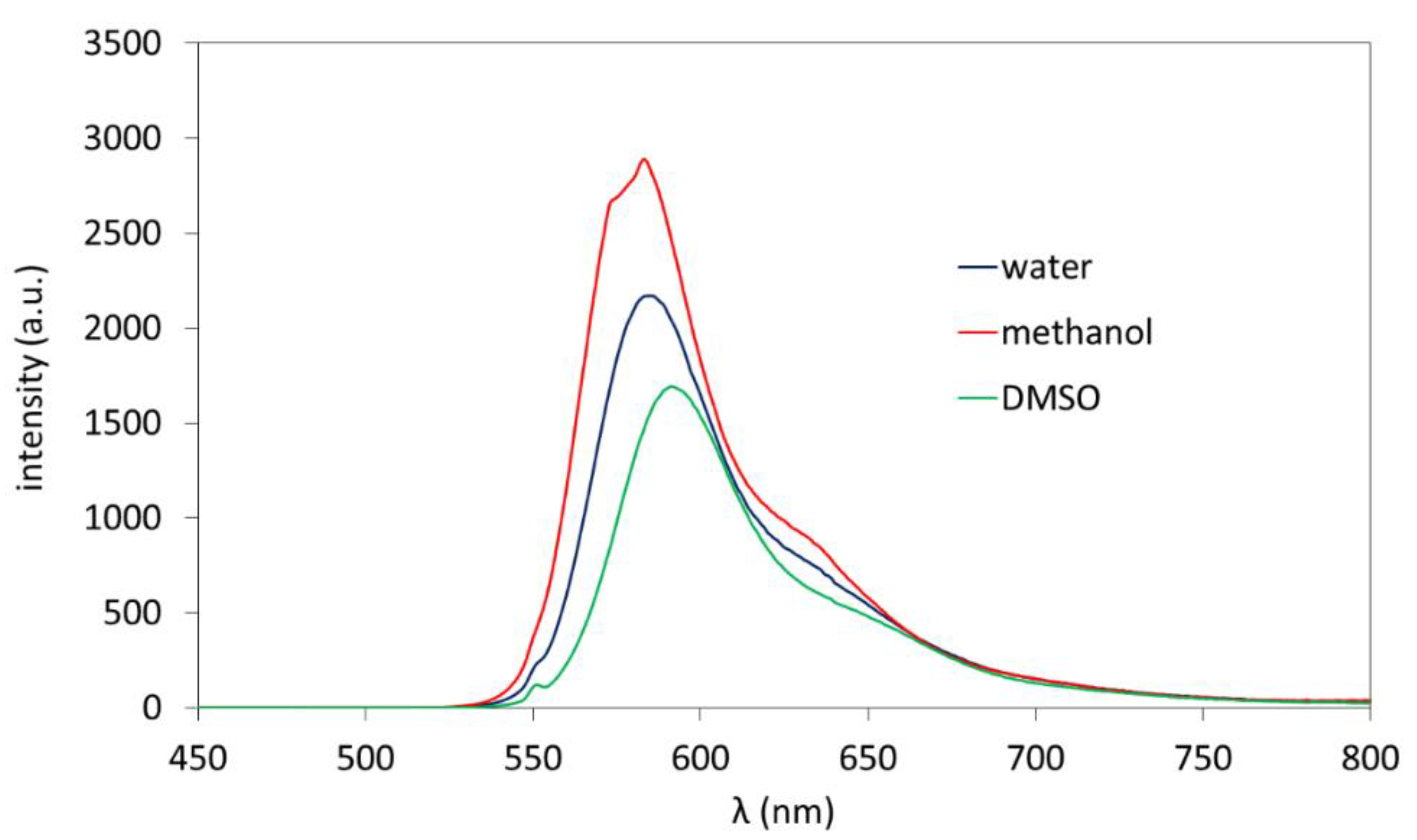
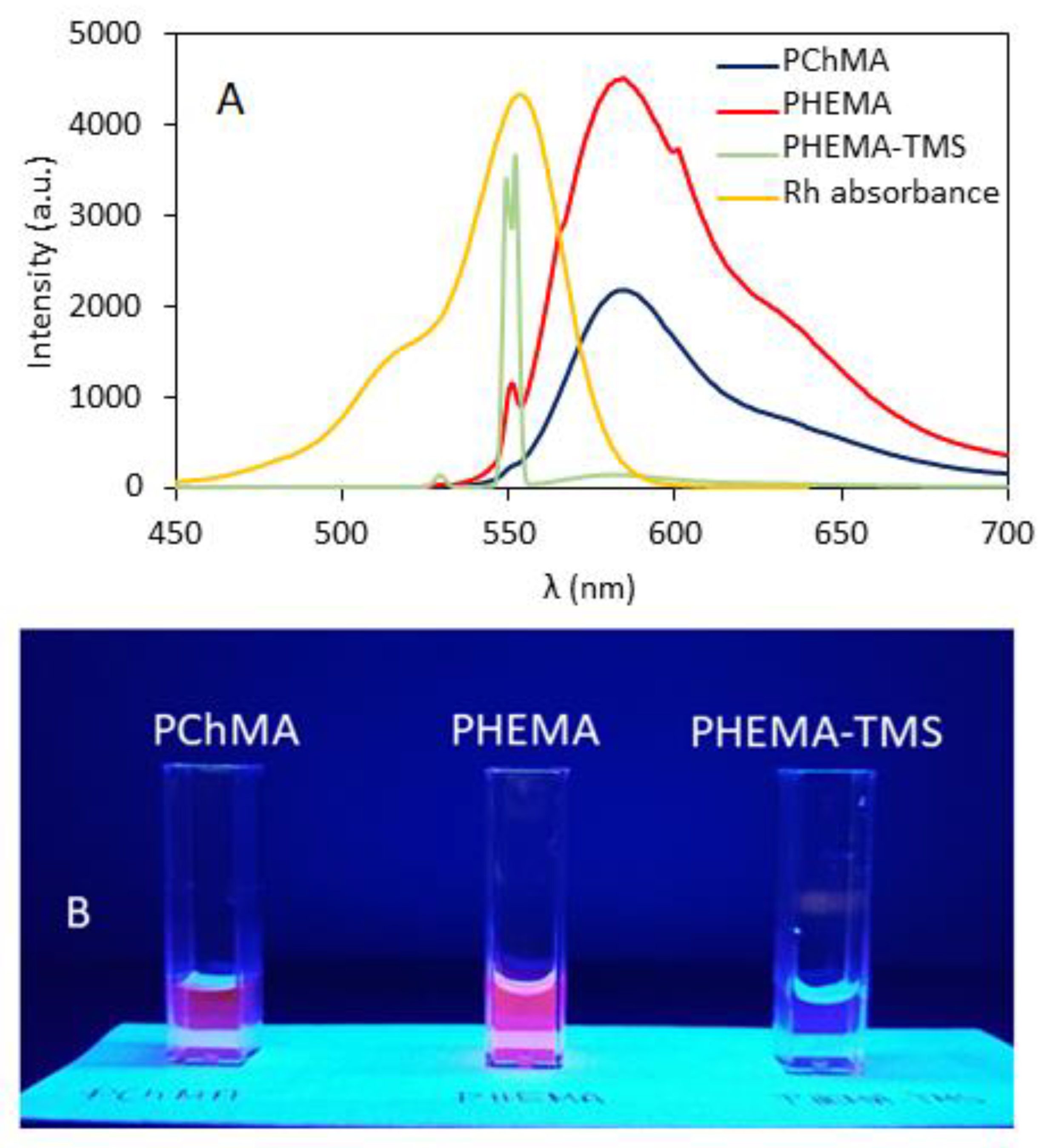
3.1. Synthesis and Polymer Characterization
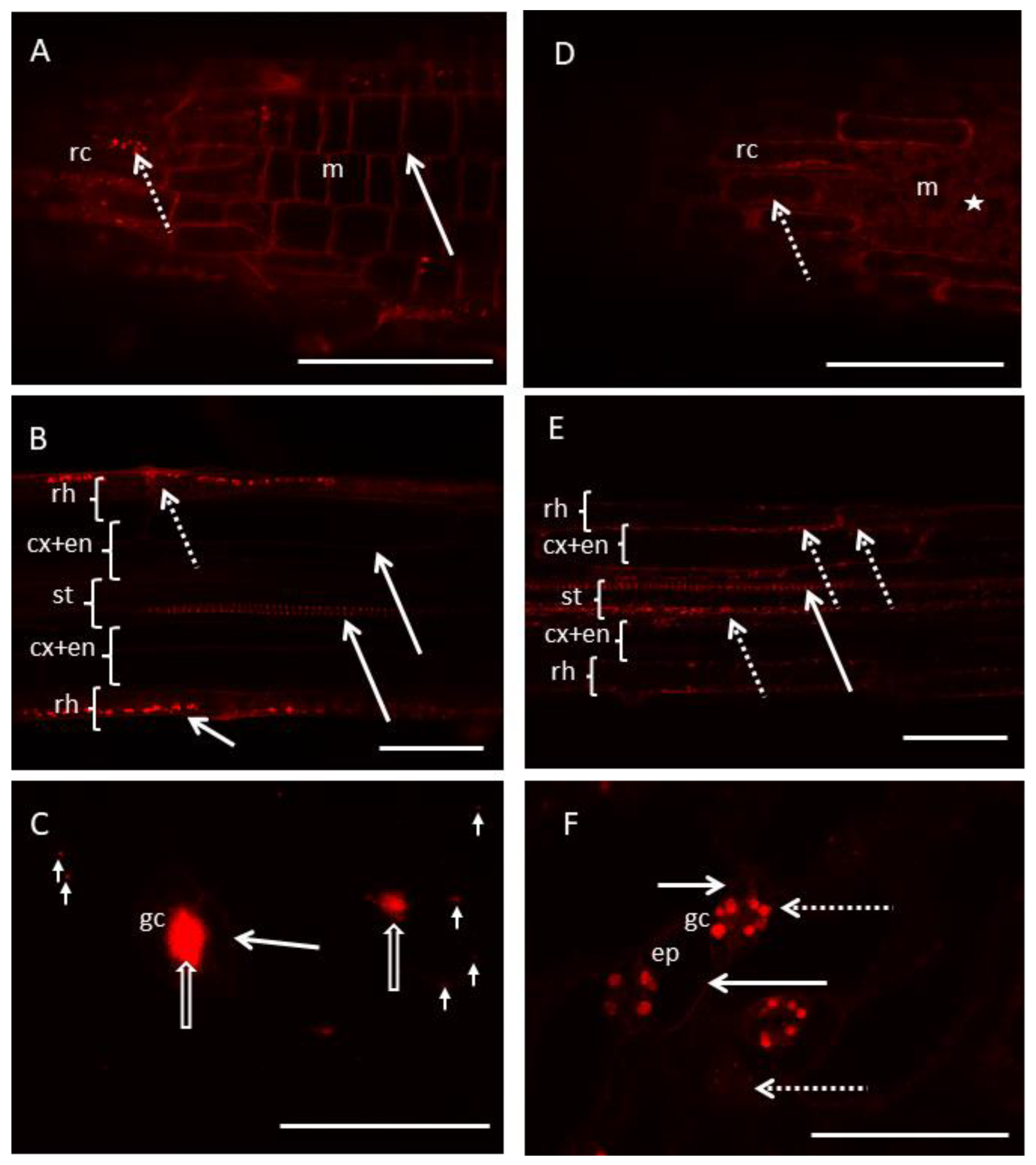
3.2. Biological Tests
3.2.1. Additional Test
3.2.2. Cell Viability Tests
4. Conclusions
- (1)
- Both RhB-polymers did not cause cell death, thus, they can be used in living plant material;
- (2)
- Different localizations of RhB-polymers depended on the charge. Positively charged RhB-PChMA was present mostly in cell walls, while neutral RhB-PHEMA was observed mostly in cytoplasm. These results suggest the differential interaction of these polymers with living cells;
- (3)
- The molecular mechanisms of such diversity are unknown and need further studies.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Breul, A.M.; Hager, M.D.; Schubert, U.S. Fluorescent monomers as building blocks for dye labeled polymers: Synthesis and application in energy conversion, biolabeling and sensors. Chem. Soc. Rev. 2013, 42, 5366–5407. [Google Scholar] [CrossRef] [PubMed]
- Wolfbeis, O.S. An overview of nanoparticles commonly used in fluorescent bioimaging. Chem. Soc. Rev. 2015, 44, 4743–4768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, J.; Ducker, R.E.; al Jaf, O.; Cartron, M.L.; Alswieleh, A.M.; Smith, C.H.; Hunter, C.N.; Armes, S.P.; Leggett, G.J. Fabrication of microstructured binary polymer brush “corrals” with integral pH sensing for studies of proton transport in model membrane systems. Chem. Sci. 2018, 9, 2238–2251. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.; Canton, I.; Warren, N.J.; Themistou, E.; Blanazs, A.; Ustbas, B.; Tian, X.; Pearson, R.; Battaglia, G.; Lewis, A.L.; et al. Nile blue-based nanosized pH sensors for simultaneous far-red and near-infrared live bioimaging. J. Am. Chem. Soc. 2013, 135, 14863–14870. [Google Scholar] [CrossRef]
- Li, G.; Zhu, X.; Cheng, Z.; Zhang, W.; Sun, B. Synthesis of poly(methyl methacrylate) labeled with fluorescein moieties via atom transfer radical polymerization. J. Macromol. Sci. Part A Pure Appl. Chem. 2008, 45, 328–334. [Google Scholar] [CrossRef]
- Mielańczyk, A.; Skonieczna, M.; Bernaczek, K.; Neugebauer, D. Fluorescein nanocarriers based on cationic star copolymers with acetal linked sugar cores. Synthesis and biochemical characterization. RSC Adv. 2014, 4, 31904–31913. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, L.; Meng, L.; Liu, Y. Synthesis of poly(N-isopropylacrylamide) by ATRP using a fluorescein-based initiator. Polym. Bull. 2007, 59, 195–206. [Google Scholar] [CrossRef]
- Neugebauer, D.; Charasim, D.; Swinarew, A.; Stolarzewicz, A.; Krompiec, M.; Janeczek, H.; Simokaitiene, J.; Gražulevičius, J.V. Polymethacrylates with anthryl and carbazolyl groups prepared by atom transfer radical polymerization. Polym. J. 2011, 43, 448–454. [Google Scholar] [CrossRef] [Green Version]
- Bielas, R.; Wróbel-Marek, J.; Kurczyńska, E.U.; Neugebauer, D. Pyranine labeled polymer nanoparticles as fluorescent markers for cell wall staining and imaging of movement within apoplast, Sensors Actuators. B Chem. 2019, 297, 126789. [Google Scholar] [CrossRef]
- Amali, A.J.; Singh, S.; Rangaraj, N.; Patra, D.; Rana, R.K. Poly(l -Lysine)–pyranine-3 coacervate mediated nanoparticle-assembly: Fabrication of dynamic pH-responsive containers. Chem. Commun. 2012, 48, 856–858. [Google Scholar] [CrossRef]
- Chou, H.-S.; Hsiao, M.-H.; Hung, W.-Y.; Yen, T.-Y.; Lin, H.-Y.; Liu, D.-M. A pH-responsive amphiphilic chitosan–pyranine core–shell nanoparticle for controlled drug delivery, imaging and intracellular pH measurement. J. Mater. Chem. B. 2014, 2, 6580–6589. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Bai, L.; Tao, F.; Deng, A.; Wang, L. A dual chemosensor for Cu2+ and Hg2+ based on a rhodamine-terminated water-soluble polymer in 100% aqueous solution. Analyst 2018, 143, 5395–5403. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Wu, S.; Zeng, F. Reusable polymer film chemosensor for ratiometric fluorescence sensing in aqueous media. Sens. Actuators B Chem. 2010, 145, 451–456. [Google Scholar] [CrossRef]
- Hirohashi, R.; Hisishi, Y. 4. Electroconductive polymers. 15. Effects of vacuum on the photoconduction of polymers. Shinku 1972, 15, 332–336. [Google Scholar] [CrossRef] [Green Version]
- Kaewtong, C.; Kampaengsri, S.; Singhana, B.; Pulpoka, B. Highly selective detection of Au3+ using rhodamine-based modified polyacrylic acid (PAA)-coated ITO. Dye. Pigment. 2017, 141, 277–285. [Google Scholar] [CrossRef]
- Min, M.; Wang, X.; Chen, Y.; Wang, L.; Huang, H.; Shi, J. Highly sensitive and selective Cu2+ sensor based on electrospun rhodamine dye doped poly(ether sulfones) nanofibers. Sensors Actuators B Chem. 2013, 188, 360–366. [Google Scholar] [CrossRef]
- Madsen, J.; Warren, N.J.; Armes, S.P.; Lewis, A.L. Synthesis of rhodamine 6G-based compounds for the ATRP synthesis of fluorescently labeled biocompatible polymers. Biomacromolecules 2011, 12, 2225–2234. [Google Scholar] [CrossRef]
- Zhu, Y.H.; Wang, J.L.; Zhang, H.B.; Khan, M.I.; Du, X.J.; Wang, J. Incorporation of a rhodamine B conjugated polymer for nanoparticle trafficking both: In vitro and in vivo. Biomater. Sci. 2019, 7, 1933–1939. [Google Scholar] [CrossRef]
- Jeong, J.H.; Park, T.G. Novel Polymer−DNA Hybrid Polymeric Micelles Composed of Hydrophobic Poly(d,l-lactic-co-glycolic Acid) and Hydrophilic Oligonucleotides. Bioconjug. Chem. 2001, 12, 917–923. [Google Scholar] [CrossRef]
- Cavalieri, F.; el Hamassi, A.; Chiessi, E.; Paradossi, G. Stable polymeric microballoons as multifunctional device for biomedical uses: Synthesis and characterization. Langmuir 2005, 21, 8758–8764. [Google Scholar] [CrossRef]
- Doll, F.; Keckeis, P.; Scheel, P.; Cölfen, H. Visualizing Cholesterol Uptake by Self-Assembling Rhodamine B-Labeled Polymer Inside Living Cells via FLIM-FRET Microscopy. Macromol. Biosci. 2019, 20, 1900081. [Google Scholar] [CrossRef] [PubMed]
- Bosma, G.; Pathmamanoharan, C.; de Hoog, E.H.A.; Kegel, W.K.; van Blaaderen, A.; Lekkerkerker, H.N.W. Preparation of monodisperse, fluorescent PMMA-latex colloids by dispersion polymerization. J. Colloid Interface Sci. 2002, 245, 292–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matyjaszewski, K.; Davis, T.P. Handbook of Radical Polymerisation; Wiley-Interscience A John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003. [Google Scholar]
- Paul, F.; Oriol, E.; Auriol, D.; Monsan, P. Acceptor reaction of a highly purified dextransucrase with maltose and oligosaccharides. Application to the synthesis of controlled-molecular-weight dextrans. Carbohydr. Res. 1986, 149, 433–441. [Google Scholar] [CrossRef]
- Milewska-Hendel, A.; Zubko, M.; Karcz, J.; Stróż, D.; Kurczyńska, E. Fate of neutral-charged gold nanoparticles in the roots of the Hordeum vulgare L. cultivar Karat. Sci. Rep. 2017, 7, 3014–3126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doblin, M.S.; Pettolino, F.; Bacic, A. Evans Review: Plant cell walls: The skeleton of the plant world. Funct. Plant Biol. 2010, 37, 357–381. [Google Scholar] [CrossRef] [Green Version]
- Liepman, A.H.; Wightman, R.; Geshi, N.; Turner, S.R.; Scheller, H.V. Arabidopsis–A powerful model system for plant cell wall research. Plant J. 2010, 61, 1107–1121. [Google Scholar] [CrossRef]
- Baron-Epel, O.; Gharyal, P.K.; Schindler, M. Pectins as mediators of wall porosity in soybean cells. Planta 1988, 175, 389–395. [Google Scholar] [CrossRef]
- Eticha, D.; Stass, A.; Horst, W.J. Cell-wall pectin and its degree of methylation in the maize root-apex: Significance for genotypic differences in aluminium resistance. Plant Cell Environ. 2005, 28, 1410–1420. [Google Scholar] [CrossRef]
- Celus, M.; Kyomugasho, C.; van Loey, A.M.; Grauwet, T.; Hendrickx, M.E. Influence of Pectin Structural Properties on Interactions with Divalent Cations and Its Associated Functionalities. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1576–1594. [Google Scholar] [CrossRef] [Green Version]
- Chrispeels, M.J.; Crawford, N.M.; Schroeder, J.I. Proteins for Transport of Water and Mineral Nutrients across the Membranes of Plant Cells. Plant Cell. 1999, 11, 661–675. [Google Scholar] [CrossRef]
- Bienert, G.P.; Bienert, M.D.; Jahn, T.P.; Boutry, M.; Chaumont, F. Solanaceae XIPs are plasma membrane aquaporins that facilitate the transport of many uncharged substrates. Plant J. 2011, 66, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Valencia, J.P.; Goodman, K.; Otegui, M.S. Endocytosis and Endosomal Trafficking in Plants. Annu. Rev. Plant Biol. 2016, 67, 309–335. [Google Scholar] [CrossRef] [PubMed]
- Horemans, N.; Asard, H.; van Gestelen, P.; Caubergs, R.J. Facilitated diffusion drives transport of oxidised ascorbate molecules into purified plasma membrane vesicles of Phaseolus vulgaris. Physiol. Plant. 1998, 104, 783–789. [Google Scholar] [CrossRef]
- Assmann, S.M.; Haubrick, L.L. Transport proteins of the plant plasma membrane. Curr. Opin. Cell Biol. 1996, 8, 458–467. [Google Scholar] [CrossRef]
- Michelet, B.; Boutry, M. The Plasma Membrane H+-ATPase (A Highly Regulated Enzyme with Multiple Physiological Functions). Plant Physiol. 1995, 108, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.; Corzo, A.; Leigh, R.A.; Sanders, D. Membrane potential-dependent calcium transport in right-side-out plasma membrane vesicles from Zea mays L. roots. Plant J. 1996, 5, 683–694. [Google Scholar] [CrossRef]
- Bielas, R.; Mielańczyk, A.; Siewniak, A.; Neugebauer, D. Trimethylammonium-Based Polymethacrylate Ionic Liquids with Tunable Hydrophilicity and Charge Distribution as Carriers of Salicylate Anions. ACS Sustain. Chem. Eng. 2016, 4, 4181–4191. [Google Scholar] [CrossRef]
- Yoon, M.; Choi, H.N.; Kwon, H.W.; Park, K.H. Solvent dependence of absorption and fluorescence spectra of piroxicam. A possible intramolecular proton transfer in the excited state. Bull. Korean Chem. Soc. 1988, 9, 171–175. [Google Scholar] [CrossRef]
- Brouwer, A.M. Standards for photoluminescence quantum yield measurements in solution (IUPAC Technical Report). Pure Appl. Chem. 2011, 83, 2213–2228. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Wróbel-Marek, J.; Kurczyńska, E.; Płachno, B.J.; Kozieradzka-Kiszkurno, M. Identification of symplasmic domains in the embryo and seed of Sedum acre L. (Crassulaceae). Planta 2017, 245, 491–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, C.J.; Mock, N.M. An improved method for monitoring cell death in cell suspension and leaf disc assays using evans blue. Plant Cell. Tissue Organ Cult. 1994, 39, 7–12. [Google Scholar] [CrossRef]
- Jones, K.; Kim, D.W.; Park, J.S.; Khang, C.H. Live-cell fluorescence imaging to investigate the dynamics of plant cell death during infection by the rice blast fungus Magnaporthe oryzae. BMC Plant Biol. 2016, 16, 69. [Google Scholar] [CrossRef] [Green Version]
- Topel, Ö.; Çakir, B.A.; Budama, L.; Hoda, N. Determination of critical micelle concentration of polybutadiene-block- poly(ethyleneoxide) diblock copolymer by fluorescence spectroscopy and dynamic light scattering. J. Mol. Liq. 2013, 177, 40–43. [Google Scholar] [CrossRef]
- Petricka, J.J.; Winter, C.M.; Benfey, P.N. Control of Arabidopsis Root Development. Annu. Rev. Plant Biol. 2012, 63, 563–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conway, L.J.; Poethig, R.S. Mutations of Arabidopsis thaliana that transform leaves into cotyledons. Proc. Natl. Acad. Sci. USA 1997, 94, 10209–10214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinke, D.W. A Homoeotic Mutant of Arabidopsis thaliana with Leafy Cotyledons. Science 1992, 258, 1647–1650. [Google Scholar] [CrossRef]
- Shtein, I.; Shelef, Y.; Marom, Z.; Zelinger, E.; Schwartz, A.; Popper, Z.A.; Bar-On, B.; Harpaz-Saad, S. Stomatal cell wall composition: Distinctive structural patterns associated with different phylogenetic groups. Ann. Bot. 2017, 119, 1021–1033. [Google Scholar] [CrossRef] [Green Version]
- Asl, L.K.; Dhondt, S.; Boudolf, V.; Beemster, G.T.S.; Beeckman, T.; Inzé, D.; Govaerts, W.; de Veylder, L. Model-based analysis of arabidopsis leaf epidermal cells reveals distinct division and expansion patterns for pavement and guard cell. Plant Physiol. 2011, 156, 2172–2183. [Google Scholar] [CrossRef] [Green Version]
- Carter, R.; Sánchez-Corrales, Y.E.; Hartley, M.; Grieneisen, V.A.; Marée, A.F.M. Pavement cells and the topology puzzle. Development 2017, 144, 4386–4397. [Google Scholar] [CrossRef]
- Ming, Y.; Sack, F.D. The too many mouths and four lips mutations affect stomatal production in Arabidopsis. Plant Cell. 1995, 7, 2227–2239. [Google Scholar] [CrossRef] [Green Version]
- Kuki, H.; Higaki, T.; Yokoyama, R.; Kuroha, T.; Shinohara, N.; Hasezawa, S.; Nishitani, K. Quantitative confocal imaging method for analyzing cellulose dynamics during cell wall regeneration in Arabidopsis mesophyll protoplasts. Plant Direct 2017, 1, e00021. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.S.; Peer, W.; Schulz, B. The Plant Plasma Membrane; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar] [CrossRef]
- Suksungworn, R.; Sanevas, N.; Wongkantrakorn, N.; Fangern, N.; Vajrodaya, S.; Duangsrisai, S. Phytotoxic effect of Haldina cordifolia on germination, seedling growth and root cell viability of weeds and crop plants, NJAS—Wageningen. J. Life Sci. 2016, 78, 175–181. [Google Scholar] [CrossRef]
- Kumpf, R.P.; Nowack, M.K. The root cap: A short story of life and death. J. Exp. Bot. 2015, 66, 5651–5662. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DP | MnNMR (g/mol) | Ð | Φ | |
|---|---|---|---|---|
| RhB-PChMA | 17 | 3500 | 1.01 | 0.15 |
| RhB-PHEMA | 28 | 3650 | 1.26 | 0.18 |
| RhB-PHEMATMS | 35 | 7100 | 1.75 | 0.06 |
| dh (nm) λ = 633 nm | dh (nm) λ = 532 nm | CMC (mg/mL) | |
|---|---|---|---|
| RhB-PChMA | 190 | 210 | 0.085 |
| RhB-PHEMA | 166 | 250 | 0.086 |
| RhB-PHEMATMS | 66 | 140 | 0.04 |
| Rhb-PChMA | RhB-PHEMA | |||
|---|---|---|---|---|
| Root | cytoplasm | cell wall | cytoplasm | cell wall |
| Root cap 1 | 95% (35 a/37 b) | 95% (35 a/37 b) | 98% (58 a/59 b) | 0% (0 a/59 b) |
| Meristematic cells 1 | 0% (0 a/37 b) | 95% (35 a/37 b) | 98% (58 a/59 b) | 0% (0 a/59 b) |
| Rhizodermis 2 | 97% (36 a/37 b) | 97% (36 a/37 b) | 93% (55 a/59 b) | 93% (55 a/59 b) * |
| Cortex and endodermis 2 | 38% (15 a/37 b) | 76% (28 a/37 b) | 93% (55 a/59 b) | 93% (55 a/59 b) * |
| Stele 3 | 38% (15 a/37 b) | 86% (32 a/37 b) | 85% (50 a/59 b) | 83% (49 a/59 b) |
| Cotyledon | cytoplasm | cell wall | cytoplasm | cell wall |
| Pavement cells | 0% (0 a/35 b) | 91% (32 a/35 b) | 86% (30 a/35 b) | 86% (30 a/35 b) |
| Guard cells | 0% (0 a/35 b) | 91% (32 a/35 b) | 86% (30 a/35 b) | 86% (30 a/35 b) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bielas, R.; Wróbel-Marek, J.; Kurczyńska, E.U.; Neugebauer, D. Rhodamine-Tagged Polymethacrylate Dyes as Alternative Tools for Analysis of Plant Cells. Materials 2022, 15, 7720. https://doi.org/10.3390/ma15217720
Bielas R, Wróbel-Marek J, Kurczyńska EU, Neugebauer D. Rhodamine-Tagged Polymethacrylate Dyes as Alternative Tools for Analysis of Plant Cells. Materials. 2022; 15(21):7720. https://doi.org/10.3390/ma15217720
Chicago/Turabian StyleBielas, Rafał, Justyna Wróbel-Marek, Ewa U. Kurczyńska, and Dorota Neugebauer. 2022. "Rhodamine-Tagged Polymethacrylate Dyes as Alternative Tools for Analysis of Plant Cells" Materials 15, no. 21: 7720. https://doi.org/10.3390/ma15217720