Synthesis of Gold Functionalised Nanoparticles with the Eranthis hyemalis Lectin and Preliminary Toxicological Studies on Caenorhabditis elegans

 , , ,
, , ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis of EHL Conjugated Gold Nanoparticles (AuNPs@EHL)
2.3. Nematode Assay
3. Results and Discussion
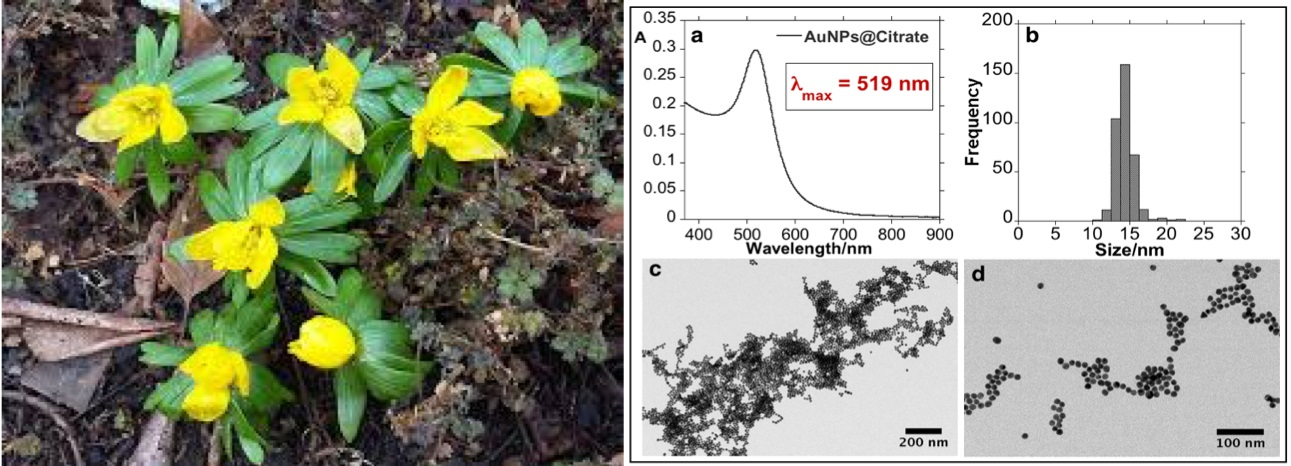
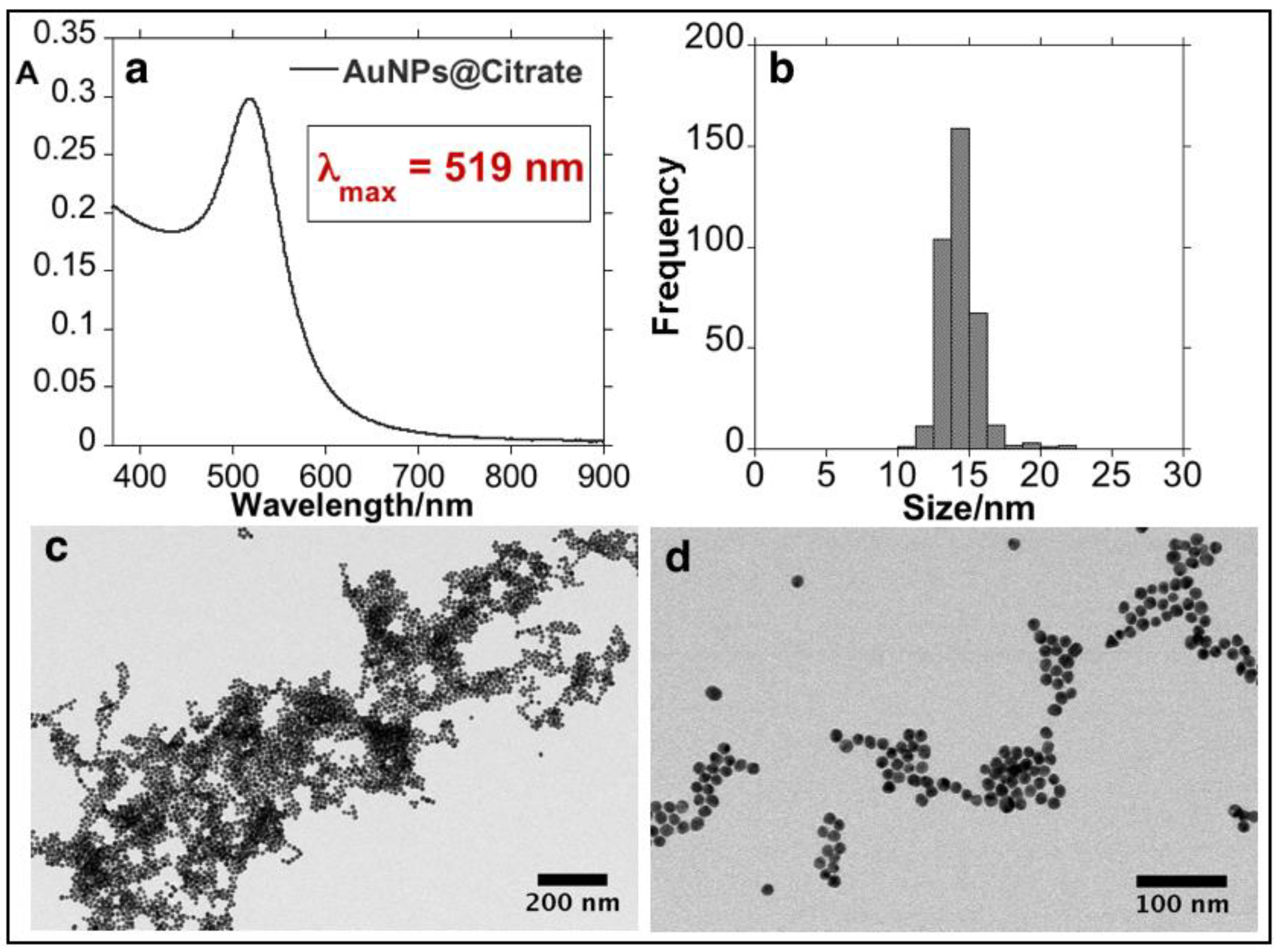
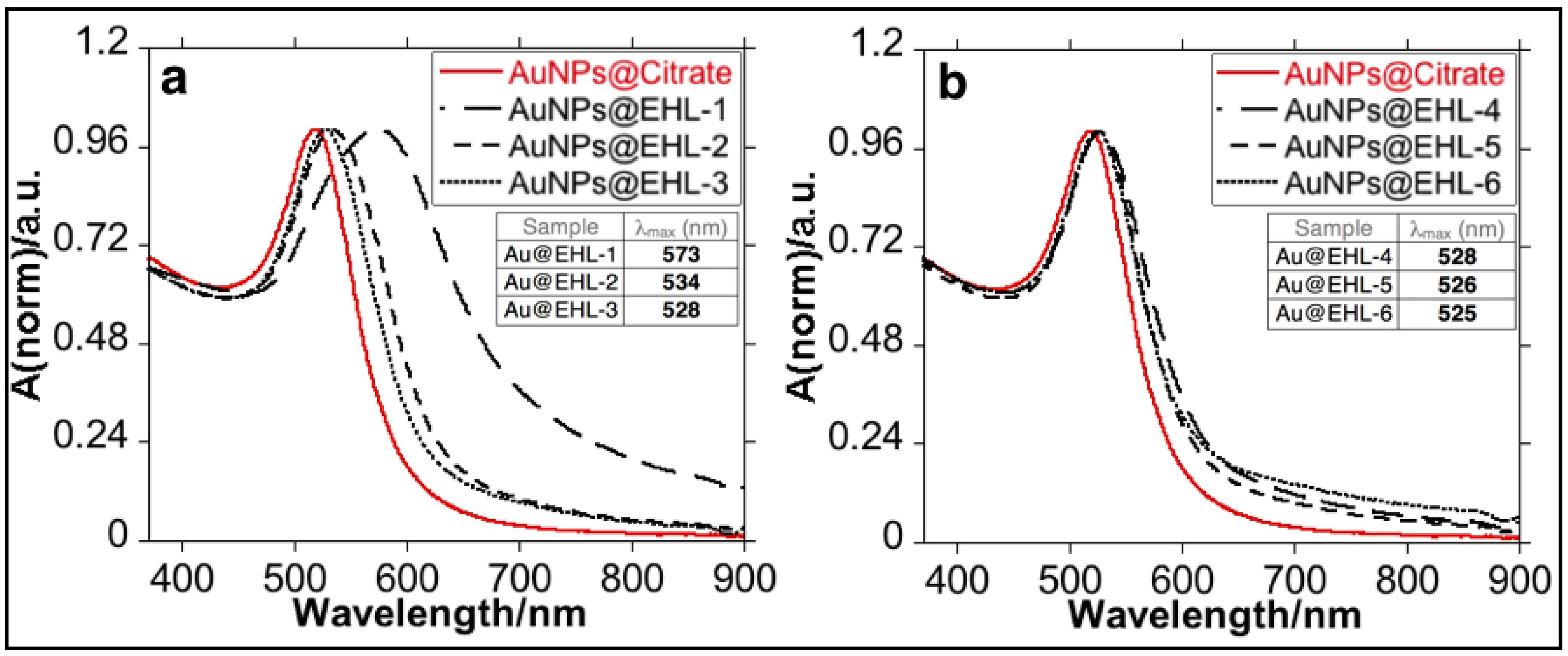
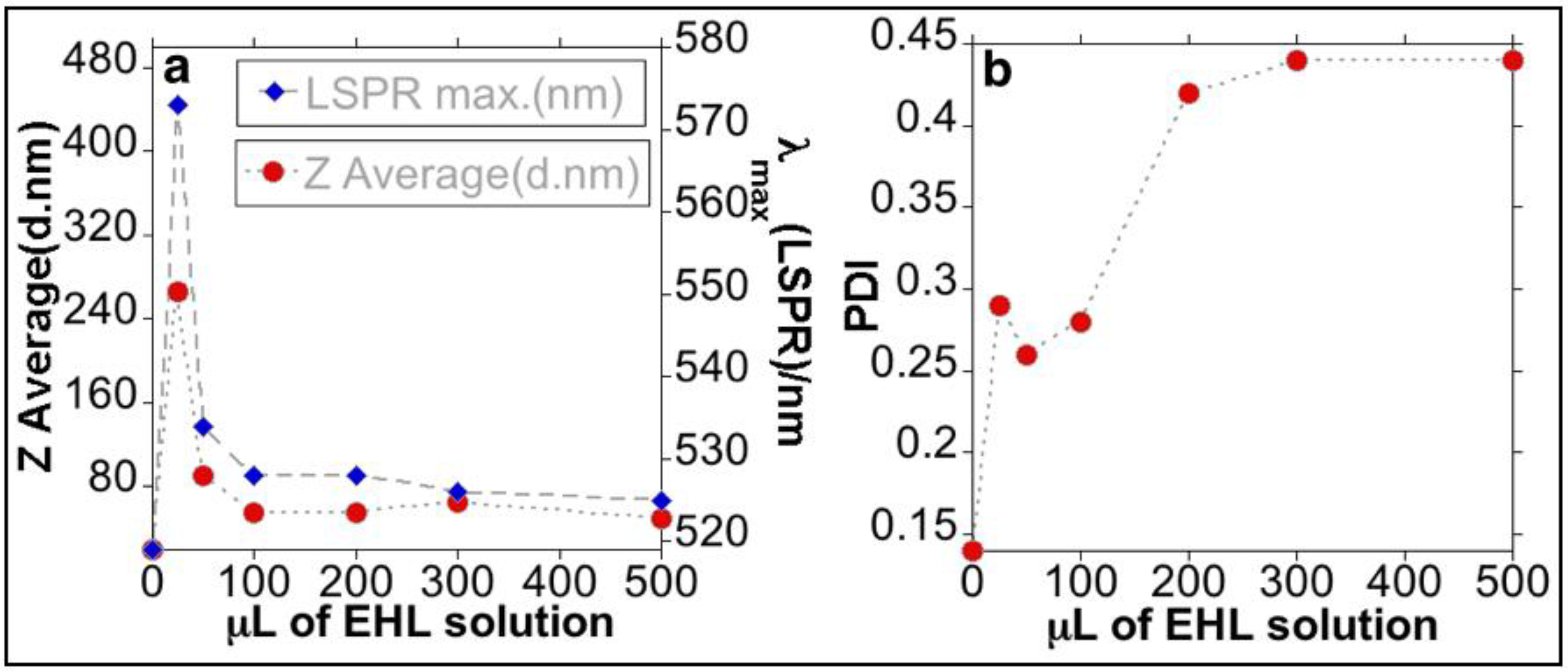
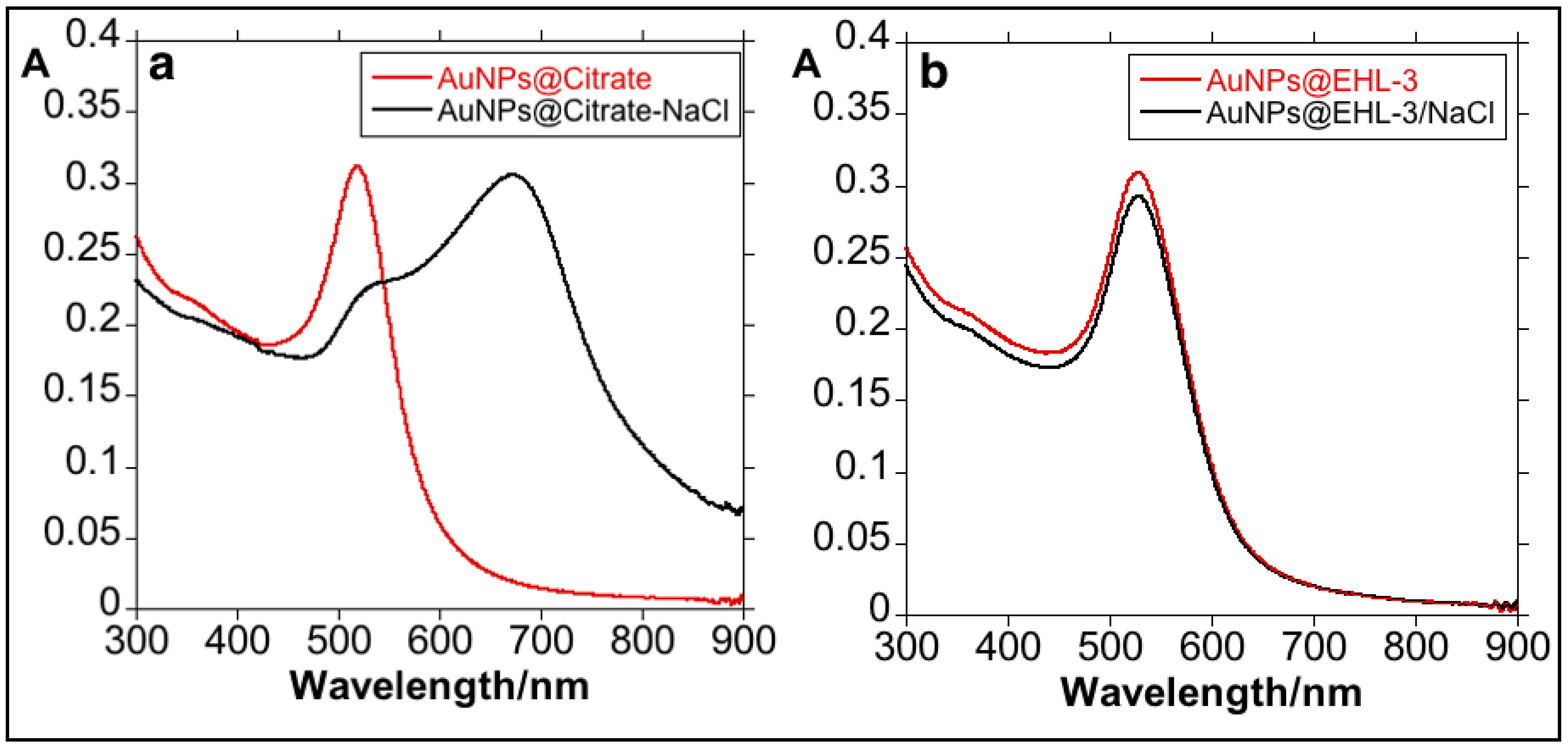
3.1. Synthesis and Characterization of the Bioconjugated Gold Nanoparticles (AuNPs@EHL)
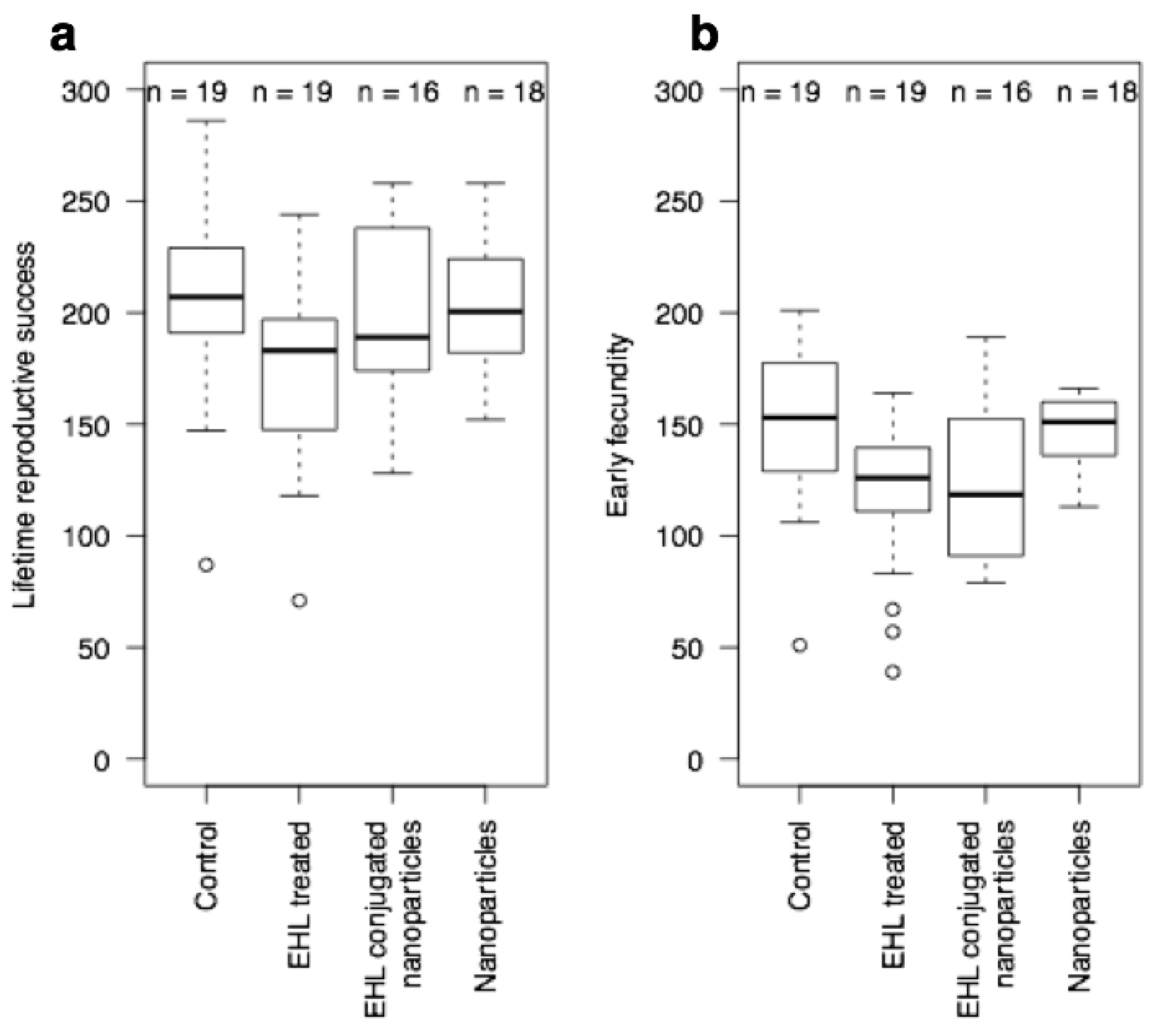
3.2. Biological Activity against C. elegans
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Peumans, W.J.; Van Damme, E. Lectins as plant defense proteins. Plant Physiol. 1995, 109, 347–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharon, N.; Lis, H. History of lectins: From hemagglutinins to biological recognition molecules. Glycobiology 2004, 14, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Bleuler-Martinez, S.; Butschi, A.; Wälti, M.A.; Egloff, P.; Stutz, K.; Yan, S.; Wilson, I.B.H.; Hengartner, M.O.; Aebi, M.; et al. Plasticity of the β-trefoil protein fold in the recognition and control of invertebrate predators and parasites by a fungal defence system. PLoS Pathog. 2012, 8, e1002706. [Google Scholar] [CrossRef]
- Delatorre, P.; Rocha, B.A.; Souza, E.P.; Oliveira, T.M.; Bezerra, G.A.; Moreno, F.B.; Azevedo, W.F. Structure of a lectin from Canavalia gladiata seeds: New structural insights for old molecules. BMC Struct. Biol. 2007, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.A.; Timm, D.; Neet, K.; Owen, W.; Peumans, W.J.; Rao, A.G. Characterization of the lectin from the bulbs of Eranthis hyemalis (winter aconite) as an inhibitor of protein synthesis. J. Biol. Chem. 1993, 268, 25176–25183. [Google Scholar] [PubMed]
- Rao, K.; Rathore, K.S.; Hodges, T.K.; Fu, X.; Stoger, E.; Sudhakar, D.; Bown, D.P. Expression of snowdrop lectin (GNA) in transgenic rice plants confers resistance to rice brown planthopper. Plant J. 1998, 15, 469–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peumans, W.J.; Hao, Q.; van Damme, E.J. Ribosome-inactivating proteins from plants: More than RNA N-glycosidases? FASEB J. 2001, 15, 1493–1506. [Google Scholar] [CrossRef]
- Edwards, M.G.; Gatehouse, A.M. Biotechnology in crop protection: Towards sustainable insect control. In Novel Biotechnologies for Biocontrol Agent Enhancement and Management; Vurro, M., Gressel, J., Eds.; NATO Security through Science Series; Springer: Dordrech, The Netherlands, 2007; pp. 1–23. ISBN 978-1-4020-5797-7. [Google Scholar]
- Cammue, B.P.; Peeters, B.; Peumans, W.J. Isolation and partial characterization of an N-acetylgalactosamine-specific lectin from winter-aconite (Eranthis hyemalis) root tubers. Biochem. J. 1985, 227, 949–955. [Google Scholar] [CrossRef] [PubMed]
- George, O.; Solscheid, C.; Bertolo, E.; Lisgarten, D. Extraction and purification of the lectin found in the tubers of Eranthis hyemalis (winter aconite). J. Integr. OMICS 2011, 1, 268–272. [Google Scholar] [CrossRef]
- McConnell, M.-T. Structural and Functional Characterisation of Eranthis Hyemalis Lectin: A Type II Ribosome Inactivating Protein. Ph.D. Thesis, Canterbury Christ Church University, Canterbury, UK, 2017. [Google Scholar]
- Ju, T.; Otto, V.I.; Cummings, R.D. The Tn antigen—Structural simplicity and biological complexity. Angew. Chem. Int. Ed. 2011, 50, 1770–1791. [Google Scholar] [CrossRef] [PubMed]
- Voss, C.; Eyol, E.; Frank, M.; Von der Lieth, C.W.; Berger, M.R. Identification and characterization of riproximin, a new type II ribosome-inactivating protein with antineoplastic activity from Ximenia americana. FASEB J. 2006, 20, 1194–1196. [Google Scholar] [CrossRef] [PubMed]
- Bayer, H.; Essig, K.; Stanzel, S.; Frank, M.; Gilderseeve, J.C.; Berger, M.R.; Voss, C. Evaluation of Riproximin binding properties reveals a novel mechanism for cellular targeting. J. Biol. Chem. 2012, 287, 35873–35886. [Google Scholar] [CrossRef] [PubMed]
- Adwan, H.; Bayer, H.; Pervaiz, A.; Sagini, M.; Berger, M.R. Riproximin is a recently discovered type II ribosome inactivating protein with potential for treating cancer. Biotechnol. Adv. 2014, 2, 1077–1090. [Google Scholar] [CrossRef] [PubMed]
- McConnell, M.-T.; Lisgarten, D.R.; Byrne, L.J.; Harvey, S.C.; Bertolo, E. Winter Aconite (Eranthis hyemalis) Lectin as a cytotoxic effector in the lifecycle of Caenorhabditis elegans. PeerJ 2015, 3, e1206. [Google Scholar] [CrossRef] [PubMed]
- Boyd, W.A.; Smith, M.V.; Freedman, J.H. Caenorhabditis elegans as a model in developmental toxicology. Methods Mol. Biol. 2012, 889, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A Transparent window into biology: A primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef] [PubMed]
- Cassada, R.C.; Russell, R.L. The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis Elegans. Dev. Biol. 1975, 46, 326–342. [Google Scholar] [CrossRef]
- Dreaden, E.C.; Alkinany, A.M.; Huang, X.; Murphy, C.J.; El-Sayed, M.A. The golden age: Gold nanoparticles for biomedicine. Chem. Soc. Rev. 2012, 41, 2740–2779. [Google Scholar] [CrossRef] [PubMed]
- Hutter, E.; Maysinger, D. Gold nanoparticles and quantum dots for bioimaging. Microsc. Res. Tech. 2011, 74, 592–604. [Google Scholar] [CrossRef] [PubMed]
- Dykman, L.; Khlebstov, N. Gold nanoparticles in biomedical applications: Recent advances and perspectives. Chem. Soc. Rev. 2012, 41, 2256–2282. [Google Scholar] [CrossRef] [PubMed]
- Libralato, G.; Galdiero, E.; Falanga, A.; Carotenuto, R.; de Alteriis, E.; Guida, M. Toxicity effects of functionalized quantum dots, gold and polystyrene nanoparticles on target aquatic biological models: A review. Molecules 2017, 22, 1439. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.Z.; Akhter, S.; Rahman, Z.; Akhter, S.; Anwar, M.; Mallik, N.; Ahmad, F.J. Nanometric gold in cancer nanotechnology: Current status and future prospect. J. Pharm. Pharmacol. 2013, 65, 634–651. [Google Scholar] [CrossRef] [PubMed]
- Rasband, W.S.; ImageJ, U.S. National Institutes of Health, Bethesda, MA, USA, 1997–2016. Available online: https://imagej.nih.gov/ij/ (accessed on 3 August 2018).
- Kimling, J.; Maier, M.; Okenve, B.; Kotaidis, V.; Ballot, H.; Plech, A. Turkevich Method for Gold Nanoparticle Synthesis Revisited. J. Phys. Chem. B 2006, 110, 15700–15707. [Google Scholar] [CrossRef] [PubMed]
- Stiernagle, T. Maintenance of C. elegans. In WormBook (The C. elegans Research Community); Wormbook: Pasadena, CA, USA, 2006. [Google Scholar]
- Shang, L.; Wang, Y.; Jiang, J.; Dong, S. pH-dependent protein conformational changes in albumin: Gold nanoparticle bioconjugates, a spectroscopy study. Langmuir 2007, 23, 2714–2721. [Google Scholar] [CrossRef] [PubMed]
- De Paoli Lacerda, S.H.; Park, J.J.; Meuse, C.; Pristinski, D.; Becker, M.L.; Karim, A.; Douglas, J.F. Interaction of Gold Nanoparticles with Common Human Blood Proteins. ACS Nano 2010, 4, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Tsai, D.H.; Delrio, F.W.; Keene, A.M.; Tyner, K.M.; MacCuspie, R.I.; Cho, T.J.; Zachariah, M.R.; Hackley, V.A. Adsorption and Conformation of Serum Albumin Protein on Gold Nanoparticles Investigated Using Dimensional Measurements and in Situ Spectroscopic Methods. Langmuir 2011, 27, 2464–2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Ni, Y. Combination of UV-Vis Spectroscopy and Chemometrics to Understand Protein-Nanomaterial Conjugate: A Case Study on Human Serum Albumin and Gold Nanoparticles. Talanta 2014, 119, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Hendel, T.; Wuithschick, M.; Kettemann, F.; Birnbaum, A.; Rademann, K.; Polte, J. In Situ Determination of Colloidal Gold Concentrations with UV–Vis Spectroscopy: Limitations and Perspectives. Anal. Chem. 2014, 86, 11115–11124. [Google Scholar] [CrossRef] [PubMed]
- Scarabelli, L.; Grzelczak, M.; Liz-Marzán, L.M. Tuning Gold Nanorod Synthesis through Prereduction with Salicylic Acid. Chem. Mater. 2013, 25, 4232–4238. [Google Scholar] [CrossRef] [Green Version]
- Scarabelli, L.; Sánchez-Iglesias, A.; Pérez-Juste, J.; Liz-Marzán, L.M. A “Tips and Tricks” Practical Guide to the Synthesis of Gold Nanorods. J. Phys. Chem. Lett. 2015, 6, 4270–4279. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, X.; Wang, L.; Hou, X.; Liu, W.; Chen, C. Interaction of gold nanoparticles with proteins and cells. Sci. Technol. Adv. Mater. 2015, 16, 034610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thobhani, S.; Attree, S.; Boyd, R.; Kumarswami, N.; Noble, J.; Szymanski, M.; Porter, R.A. Bioconjugation and characterisation of gold colloid-labelled proteins. J. Immunol. Methods 2010, 356, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.L.; Boyd, W.A.; Williams, P. Assessment of sublethal endpoints for toxicity testing with the nematode Caenorhabditis elegans. Environ. Toxicol. Chem. 2001, 20, 833–838. [Google Scholar] [CrossRef]
- Dhawan, R.; Dusenbery, D.B.; Williams, P.L. Comparison of lethality, reproduction, and behavior as toxicological endpoints in the nematode Caenorhabditis elegans. J. Toxicol. Environ. Health Part A 1999, 58, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Leung, M.C.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Khare, P.; Sonane, M.; Pandey, R.; Ali, S.; Gupta, K.C.; Satish, A. Adverse effects of TiO2 and ZnO nanoparticles in soil nematode, Caenorhabditis elegans. J. Biomed. Nanotechnol. 2011, 7, 116–117. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wick, R.L.; Xing, B. Toxicity of nanoparticulate and bulk ZnO, Al2O3 and TiO2 to the nematode Caenorhabditis elegans. Environ. Pollut. 2009, 157, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Stutz, K.; Kaech, A.; Aebi, M.; Künzler, M.; Hengartner, M.O. Disruption of the C. elegans intestinal brush border by the fungal lectin CCL2 phenocopies dietary lectin toxicity in mammals. PLoS ONE 2015, 10, e0129381. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AuNPs@EHL Sample | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Vol. EHL added (µL) | 25 | 50 | 100 | 200 | 300 | 500 |
| Total vol. of the reaction (µL) | 5025 | 5050 | 5100 | 5200 | 5300 | 5500 |
| Mass EHL in the reaction (µg) | 27.3 | 54.5 | 109 | 218 | 327 | 545 |
| Mass EHL in supernatant (µg/mL) | 3.1 ± 0.1 | 7.4 ± 0.3 | 4.0 ± 0.1 | 40 ± 2 | 61.1 ± 0.4 | 84 ± 3 |
| Mass EHL in VT supernatant (µg) | 15.6 ± 0.5 | 37.4 ± 0.3 | 20.4 ± 0.1 | 208.0 ± 10 | 323.8 ± 2 | 462.0 ± 17 |
| [EHL] in the NPs (µg) | 11.7 | 17.1 | 88.6 | 10.0 | 3.2 | 83.0 |
| Z-Average value (nm) | 266.4 | 90.3 | 54.4 | 51.7 | 60.8 | 51.3 |
| Polydispersity Index (PDI) | 0.29 | 0.26 | 0.28 | 0.42 | 0.44 | 0.44 |
| Zeta Potential (mV/cm) | −23.1 | −19.8 | −27.8 | −24.6 | −20.2 | −29.4 |
| Treatment | No. of Plates | L1s per Plate | Mean % Survival (min. and max.) | Mean % Dauer Formation (min. and max.) |
|---|---|---|---|---|
| Control | 11 | 54.6 ± 3.8 | 68 (53–81) | 0 |
| EHL | 12 | 64.2 ± 7.0 | 23 (11–40) * | 24 (0–45) * |
| AuNPs@Citrate | 11 | 58.6 ± 3.2 | 73 (54–84) | 0 |
| AuNPs@EHL | 12 | 48.7 ± 6.6 | 68 (49–83) | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djafari, J.; McConnell, M.T.; Santos, H.M.; Capelo, J.L.; Bertolo, E.; Harvey, S.C.; Lodeiro, C.; Fernández-Lodeiro, J. Synthesis of Gold Functionalised Nanoparticles with the Eranthis hyemalis Lectin and Preliminary Toxicological Studies on Caenorhabditis elegans. Materials 2018, 11, 1363. https://doi.org/10.3390/ma11081363
Djafari J, McConnell MT, Santos HM, Capelo JL, Bertolo E, Harvey SC, Lodeiro C, Fernández-Lodeiro J. Synthesis of Gold Functionalised Nanoparticles with the Eranthis hyemalis Lectin and Preliminary Toxicological Studies on Caenorhabditis elegans. Materials. 2018; 11(8):1363. https://doi.org/10.3390/ma11081363
Chicago/Turabian StyleDjafari, Jamila, Marie T. McConnell, Hugo M. Santos, José Luis Capelo, Emilia Bertolo, Simon C. Harvey, Carlos Lodeiro, and Javier Fernández-Lodeiro. 2018. "Synthesis of Gold Functionalised Nanoparticles with the Eranthis hyemalis Lectin and Preliminary Toxicological Studies on Caenorhabditis elegans" Materials 11, no. 8: 1363. https://doi.org/10.3390/ma11081363