Investigation of Polyaniline and a Functionalised Derivative as Antimicrobial Additives to Create Contamination Resistant Surfaces


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Activity of PANI and P3ABA Suspensions against E. coli and S. aureus
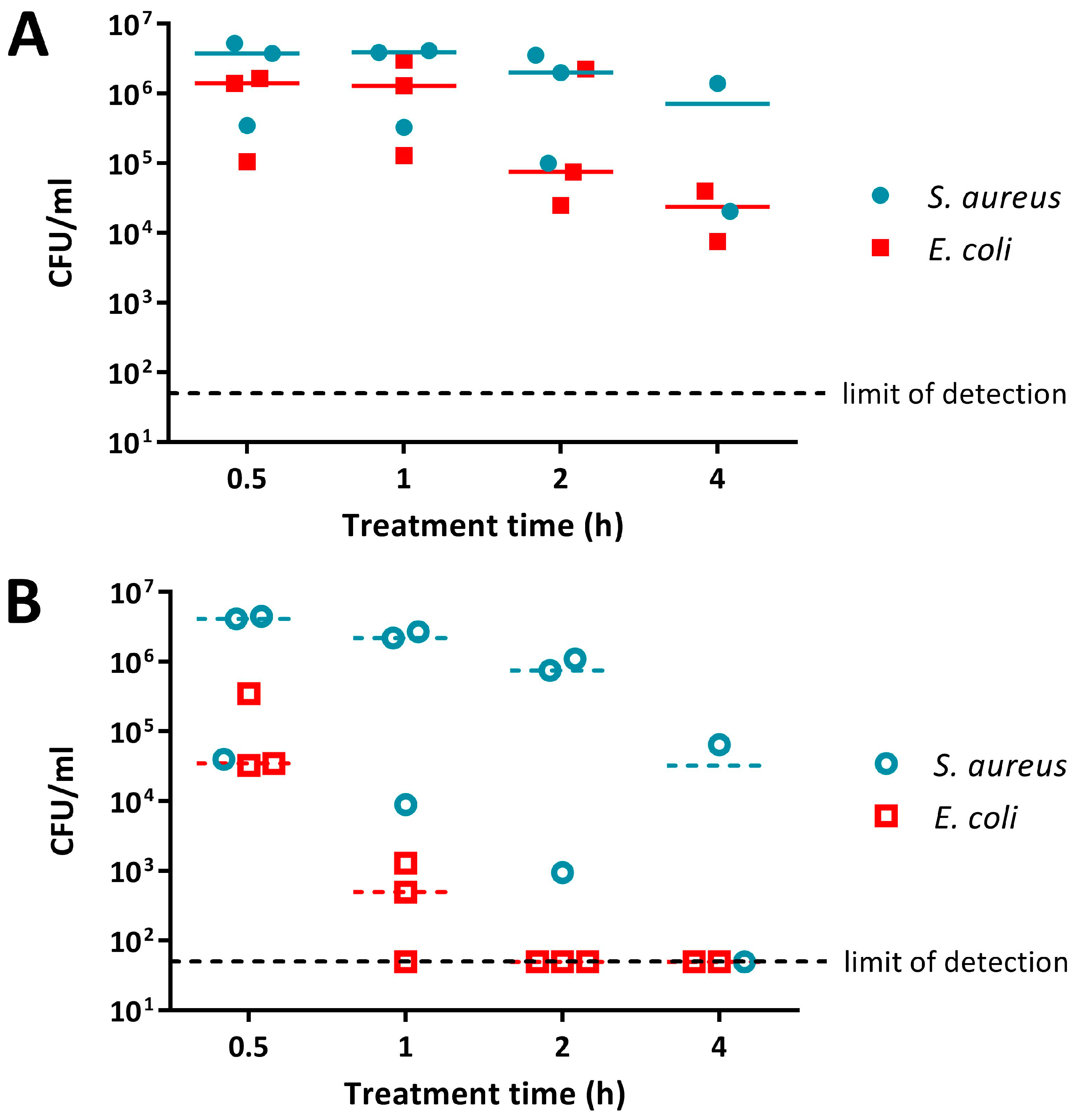
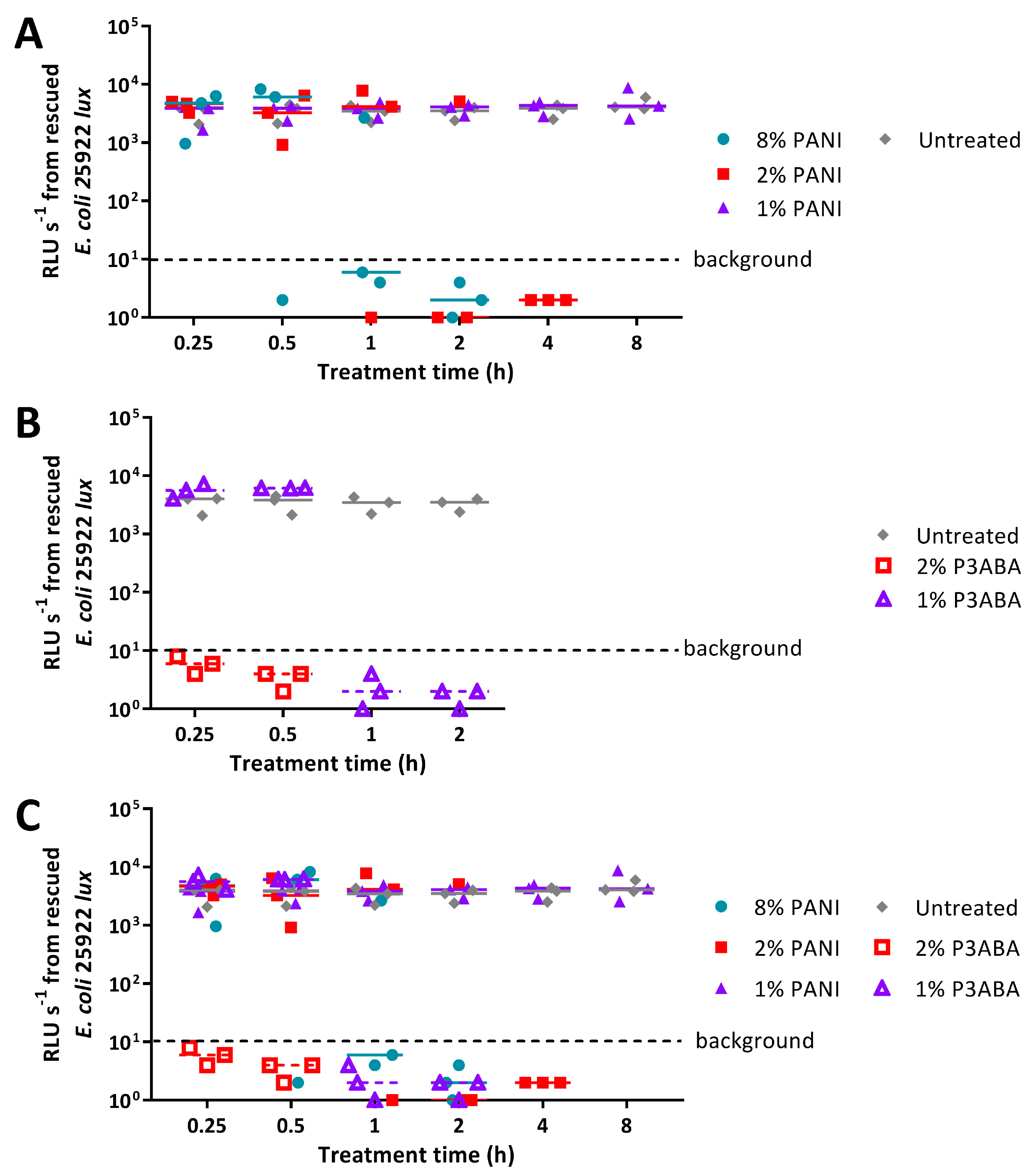
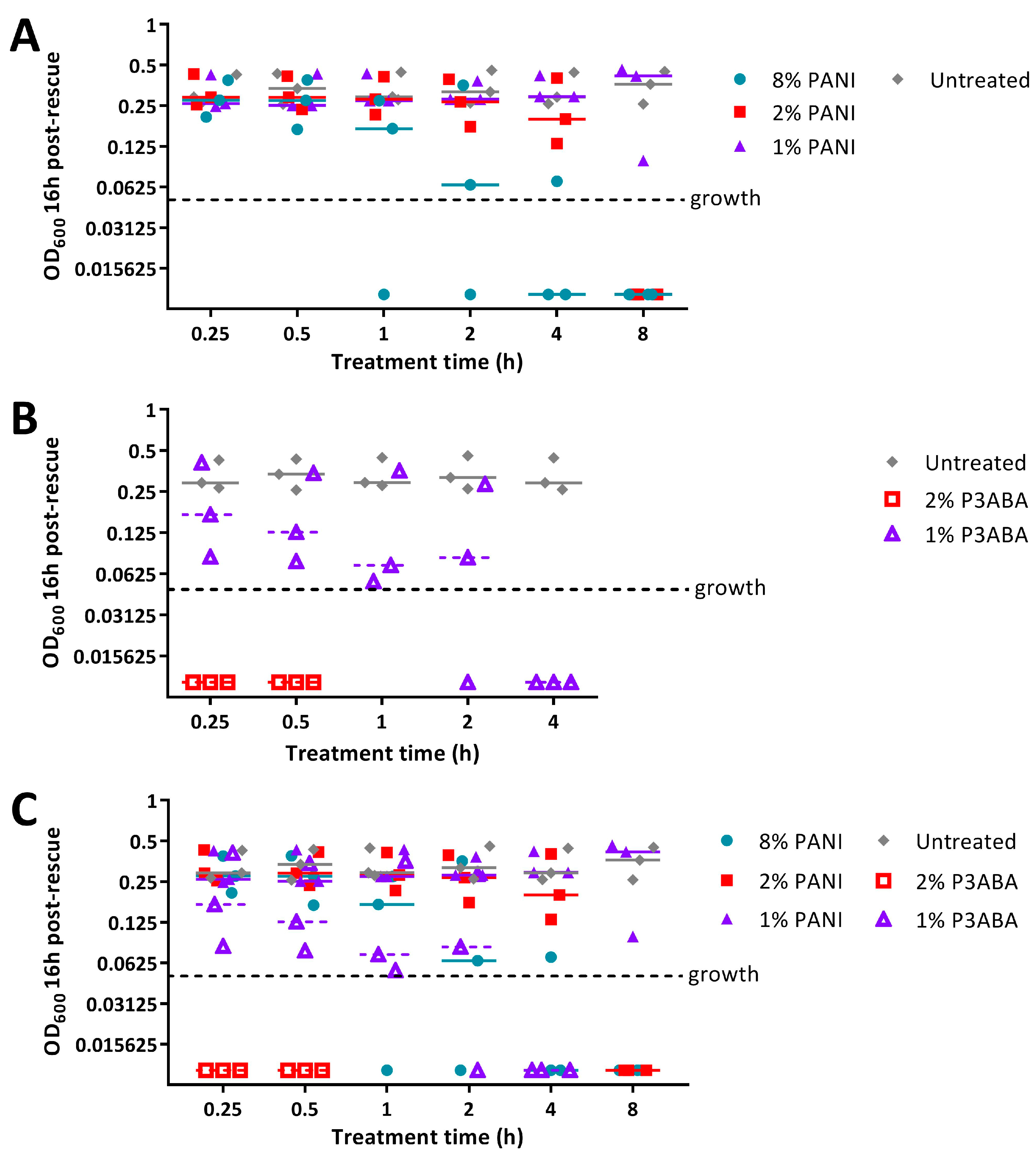
2.2. Activity of Absorbent Surfaces Containing PANI and P3ABA against E. coli and S. aureus
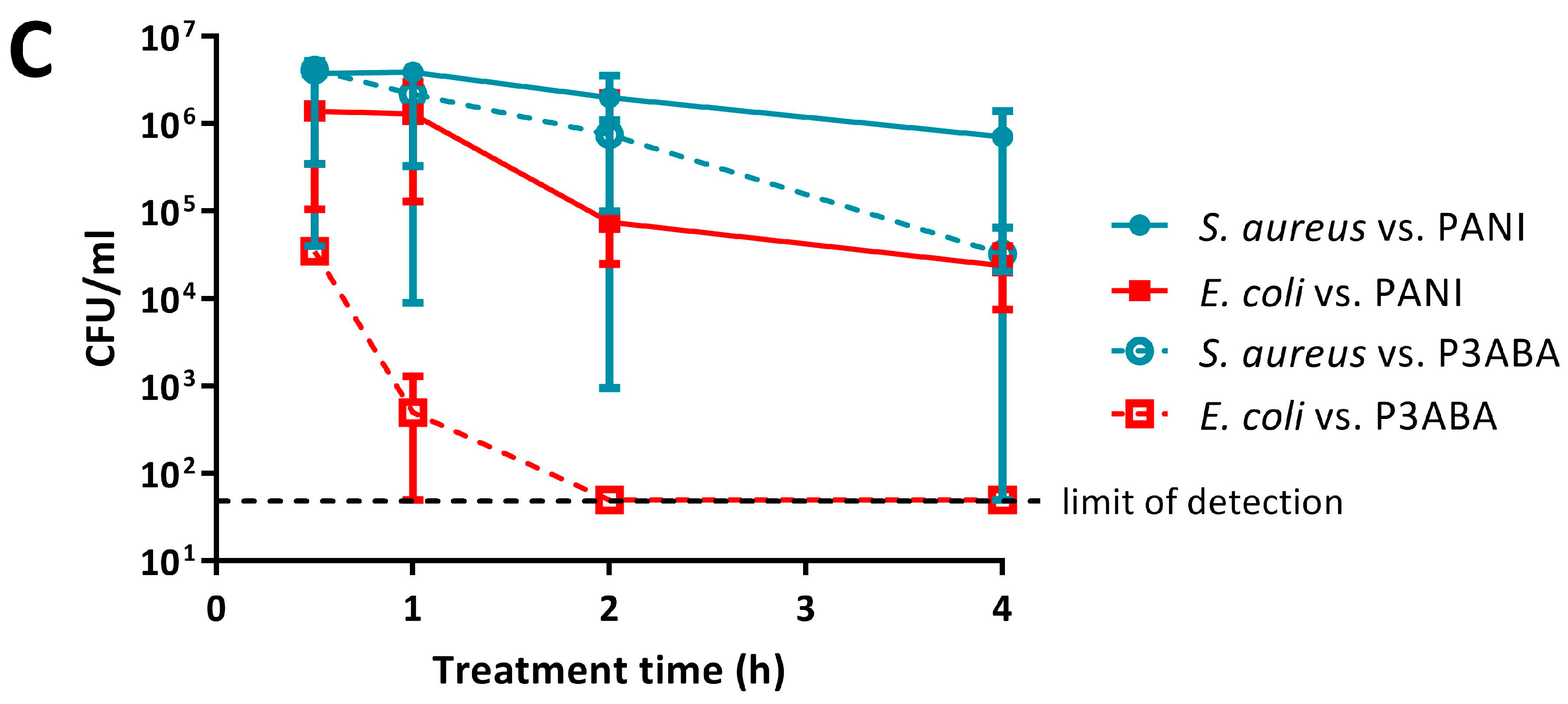
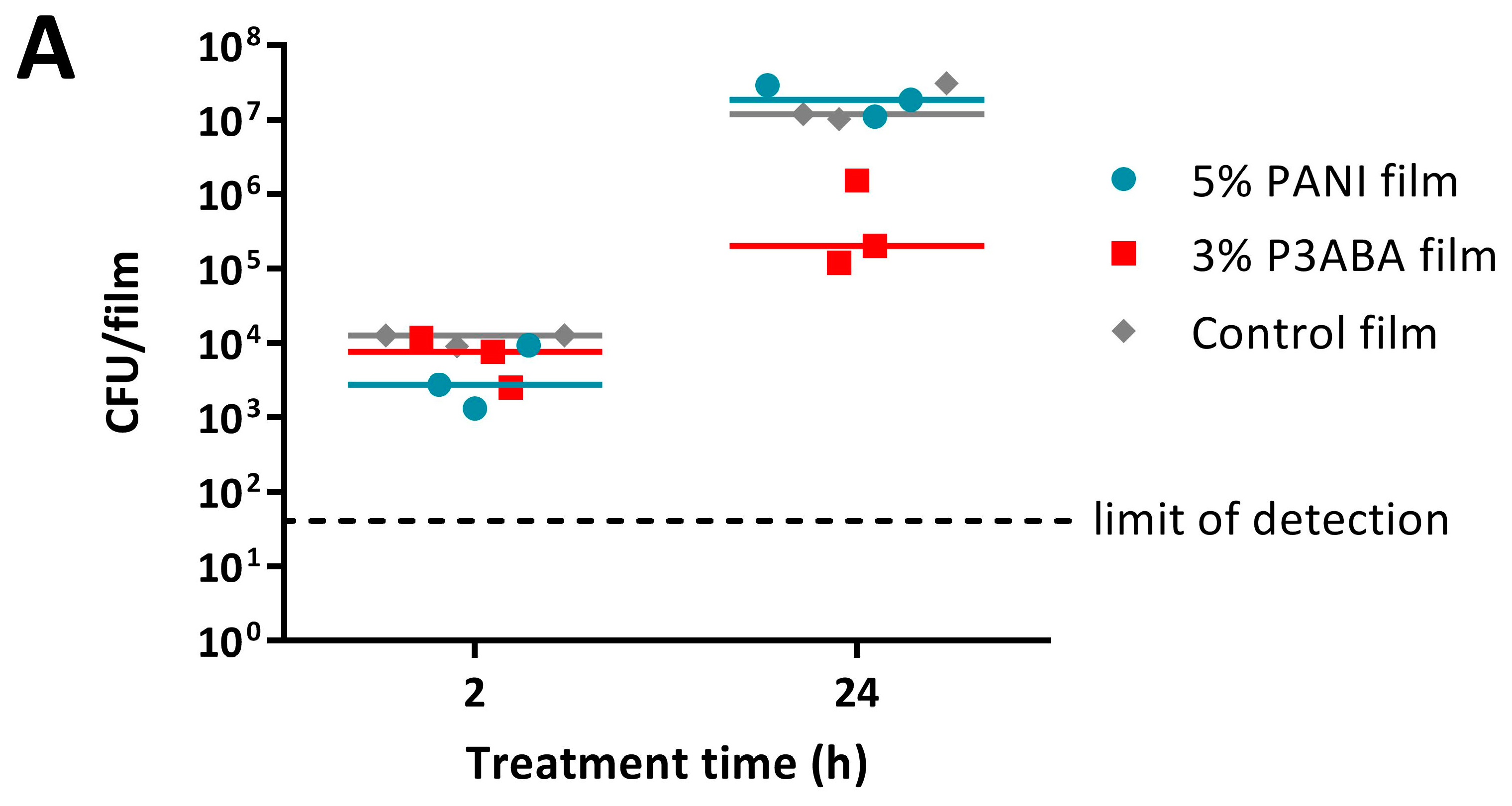
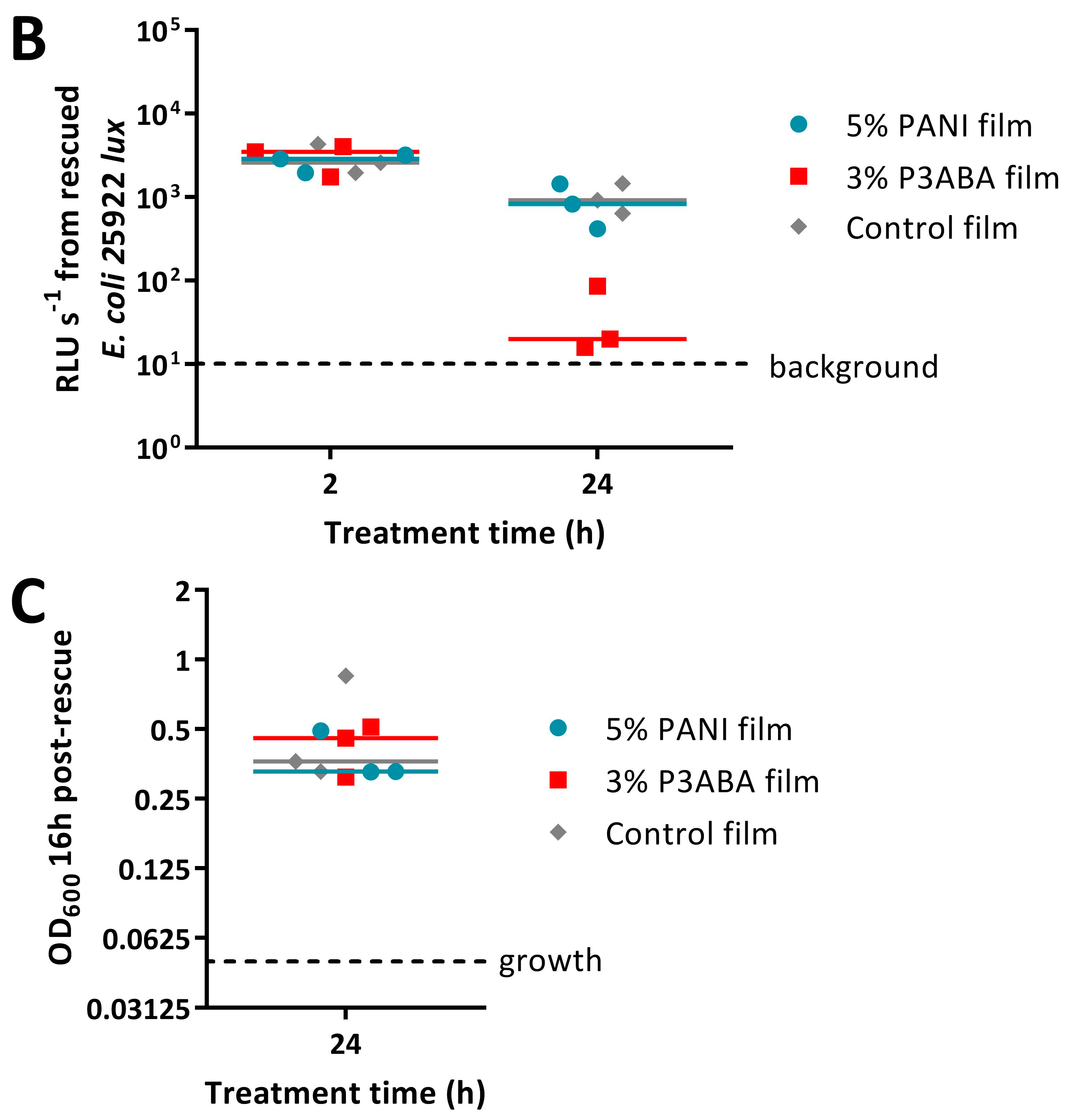
2.3. Activity of Non-Absorbent Surfaces Containing PANI and P3ABA against E. coli and S. aureus
2.4. Characterisation of the Action of PANI and P3ABA Films against E. coli and S. aureus
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Media and Chemicals
4.3. Preparation of PANI and P3ABA Suspensions
4.4. Activity of PANI and P3ABA Suspensions against E. coli and S. aureus
4.5. Activity of Absorbent Surfaces Containing PANI and P3ABA against E. coli and S. aureus
4.6. Activity of Non-Absorbent Surfaces Containing PANI and P3ABA against E. coli and S. aureus
4.7. Characterisation of the Action of PANI and P3ABA Films against E. coli and S. aureus
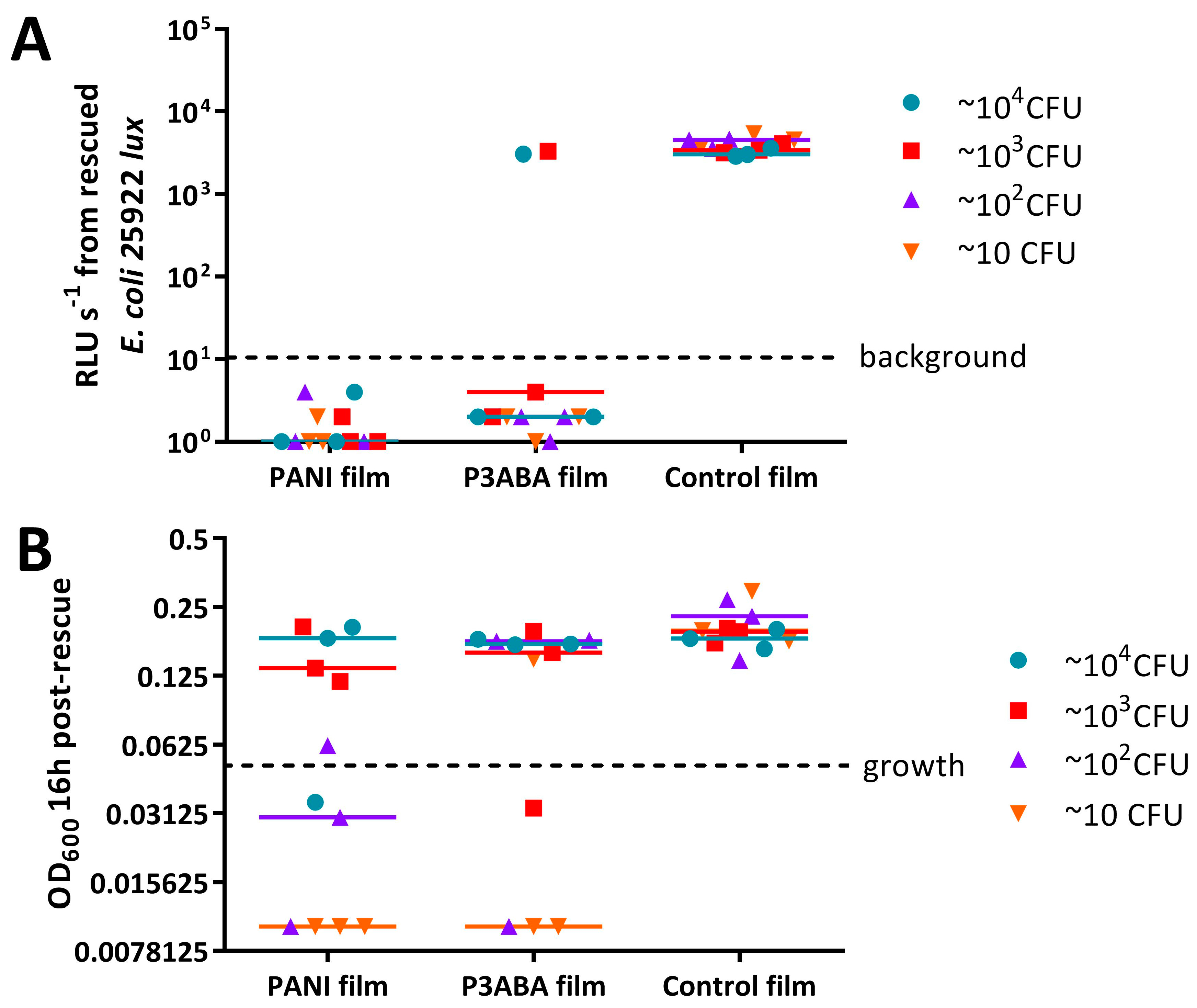
4.7.1. Challenge of PANI and P3ABA Films with a Range of CFU Doses of E. coli 25922 lux and S. aureus 6538 in Saline
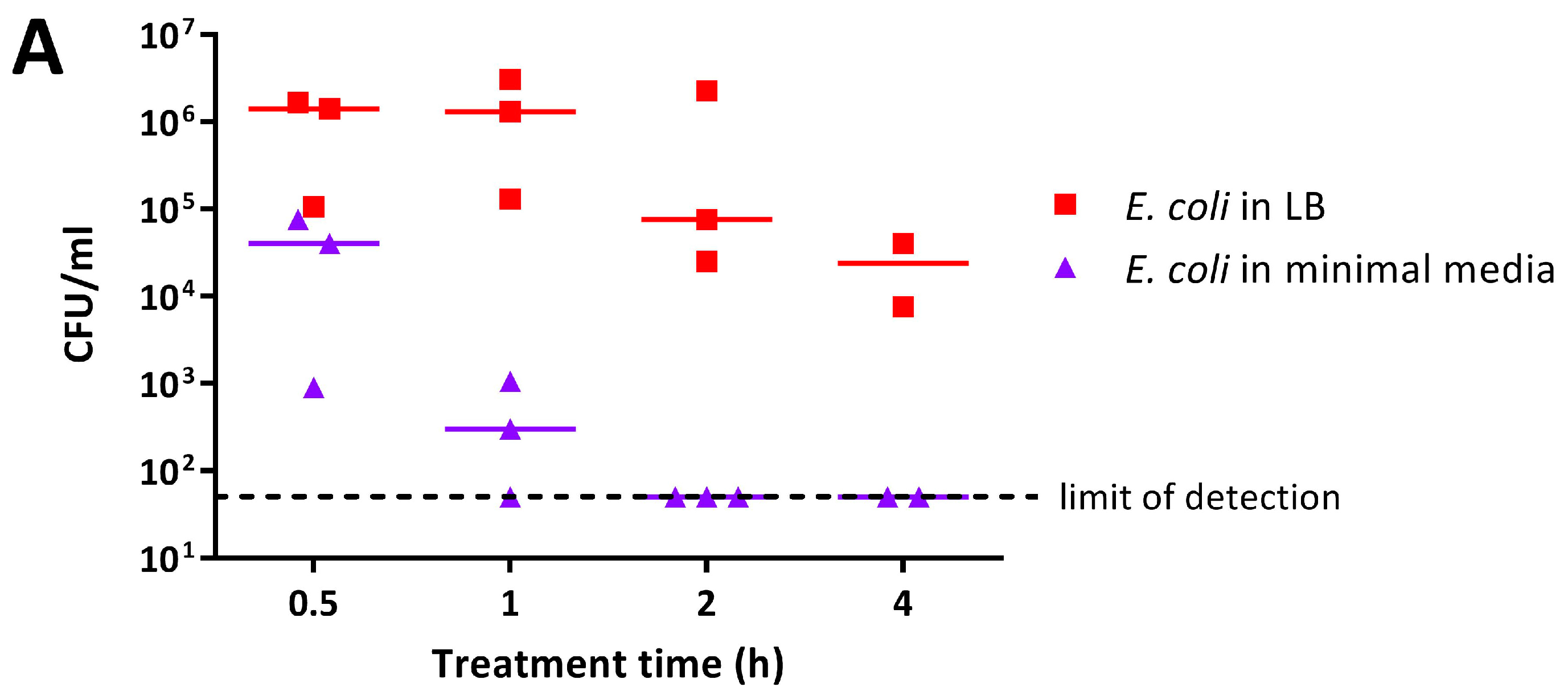
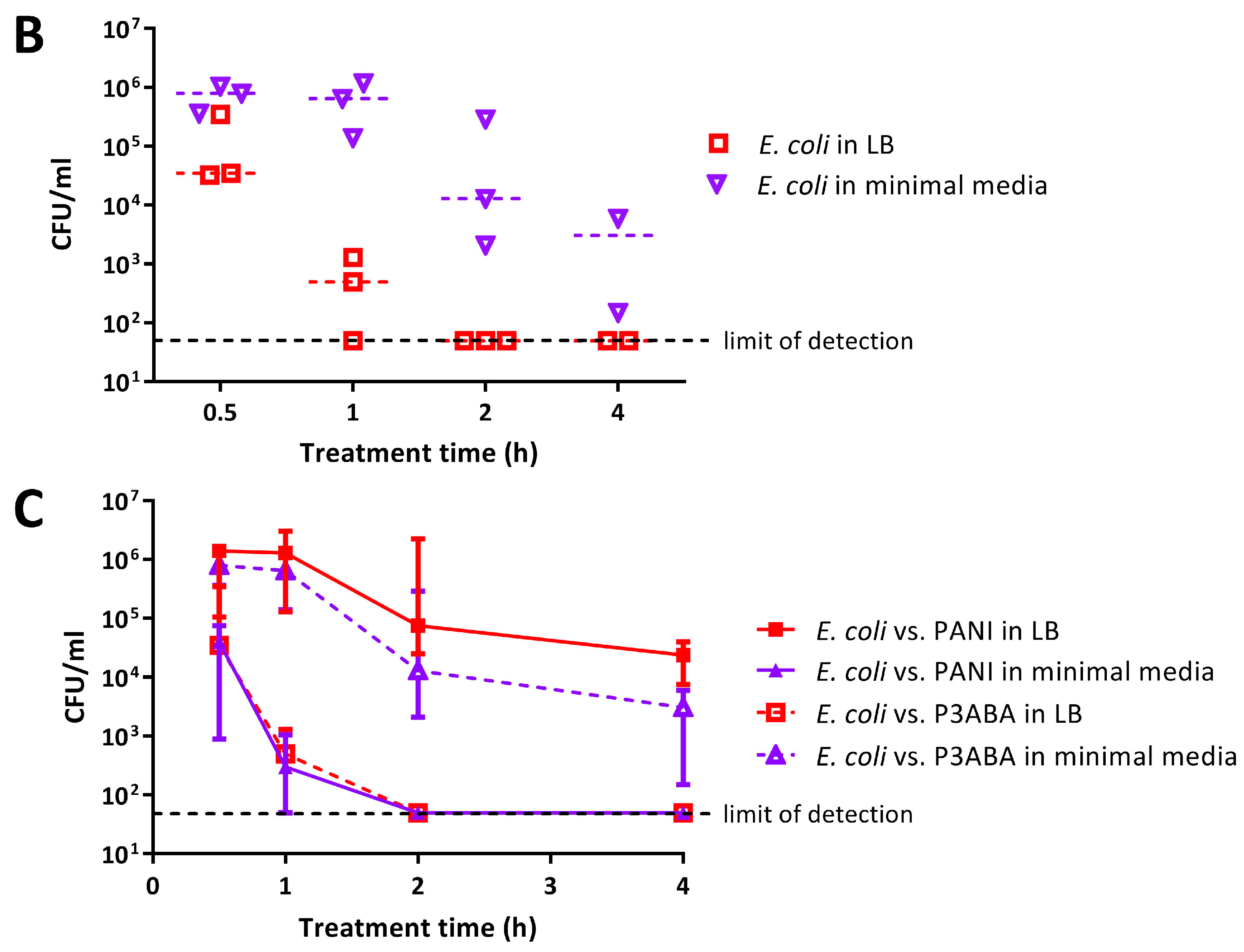
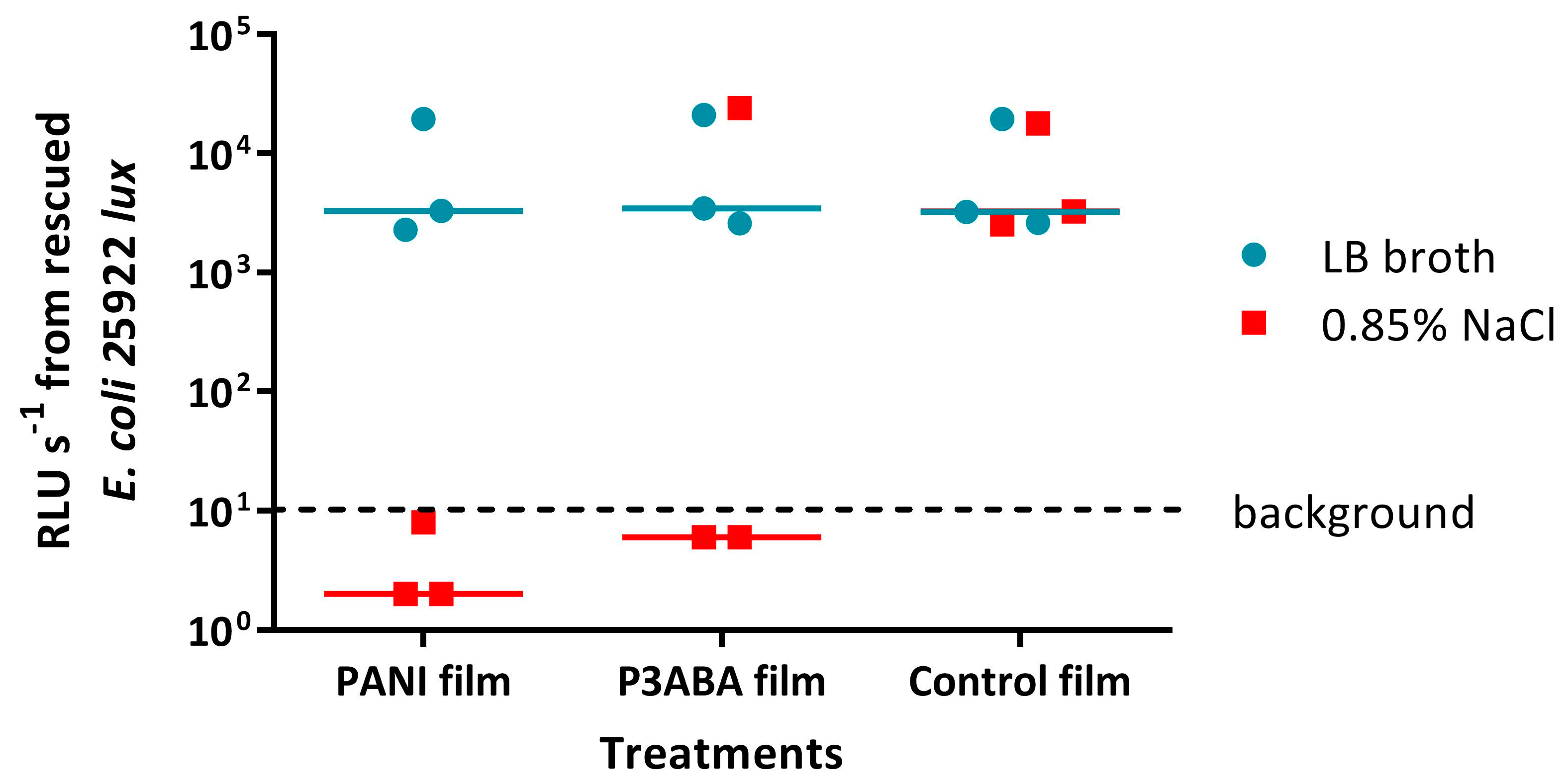
4.7.2. Assay to Evaluate the Influence of the Presence of Organic Matter on the Activity of PANI and P3ABA Films against E. coli 25922 lux
4.8. Appendix A Methods
4.8.1. Validation of Utilisation of E. coli 25922 lux
4.8.2. Determination of the Limit of Detection for E. coli 25922 lux and S. aureus 6538 Growing in a 96 Well Plate
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
Appendix A.1. Validation of Utilisation of E. coli 25922 lux as a Proxy of E. coli 25922 for Investigation of PANI and P3ABA as Surface Antimicrobial Agents
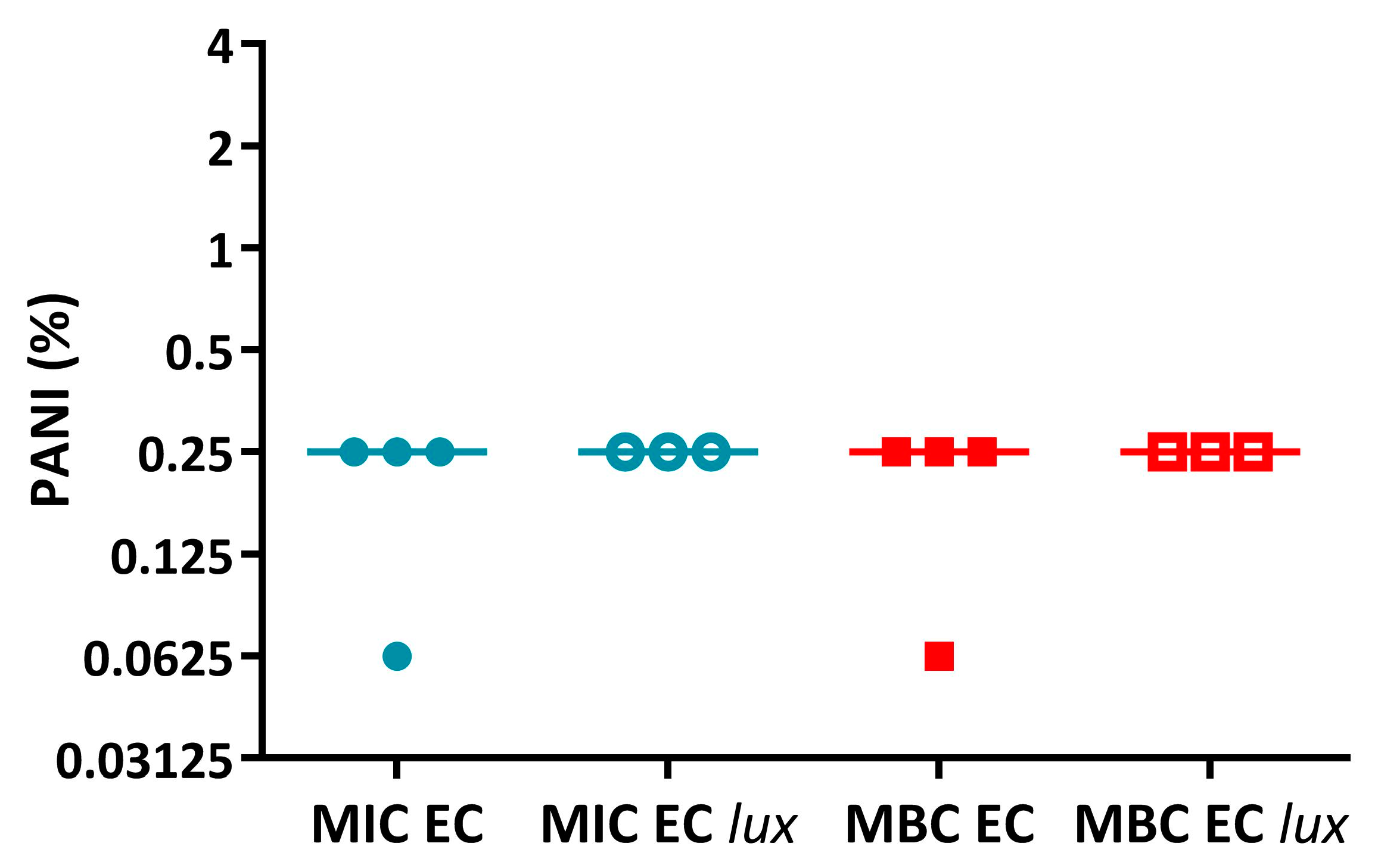
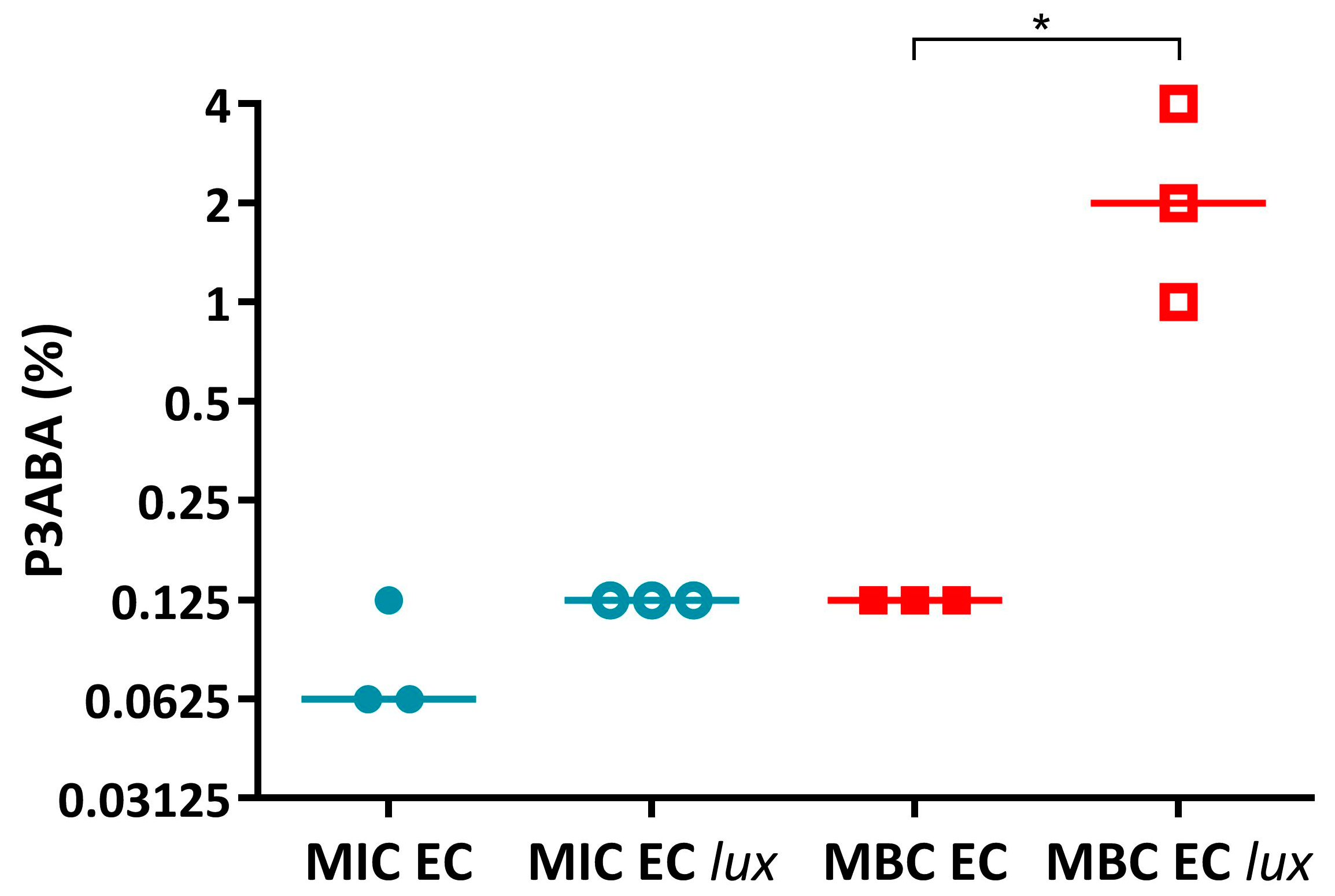
Appendix A.1.1. PANI Has Similar Activity against E. coli 25922 and E. coli 25922 lux While P3ABA Is Less Active against the Latter


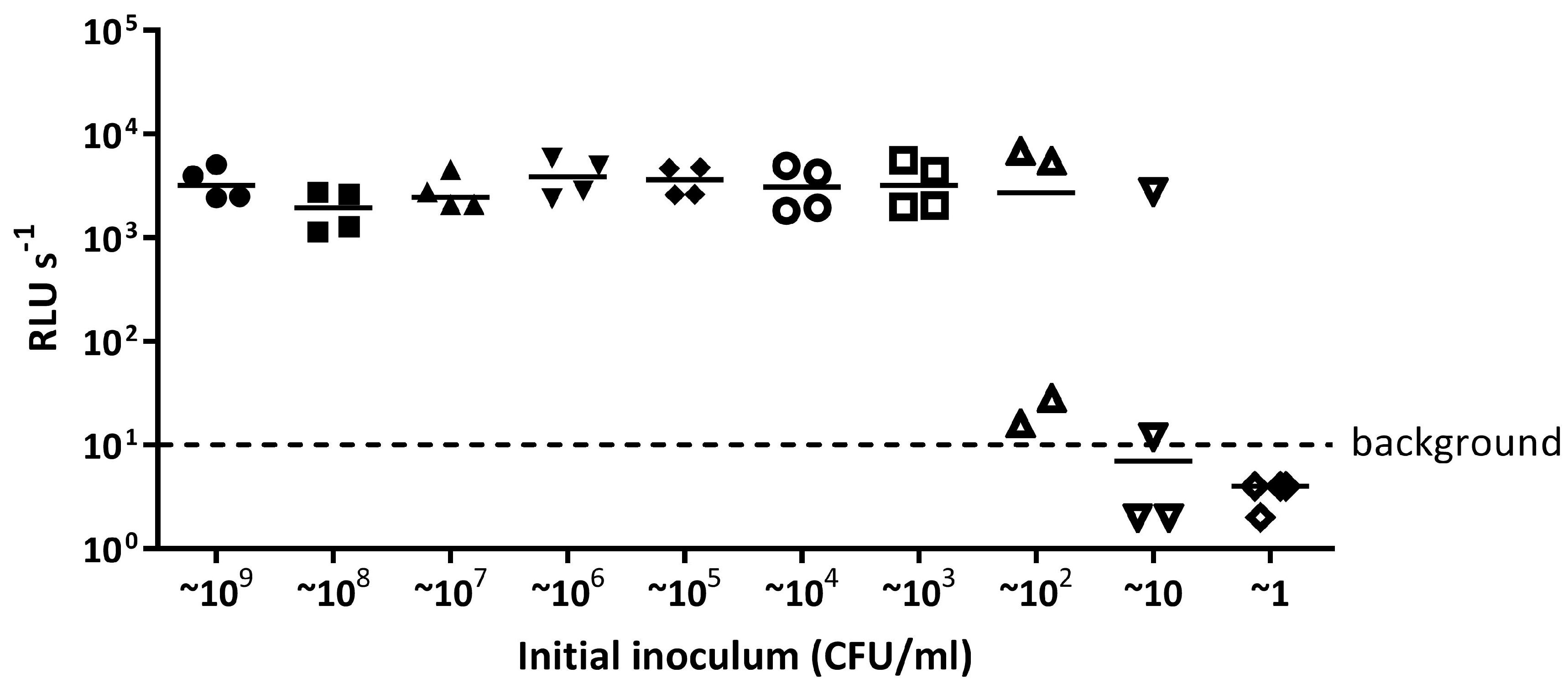
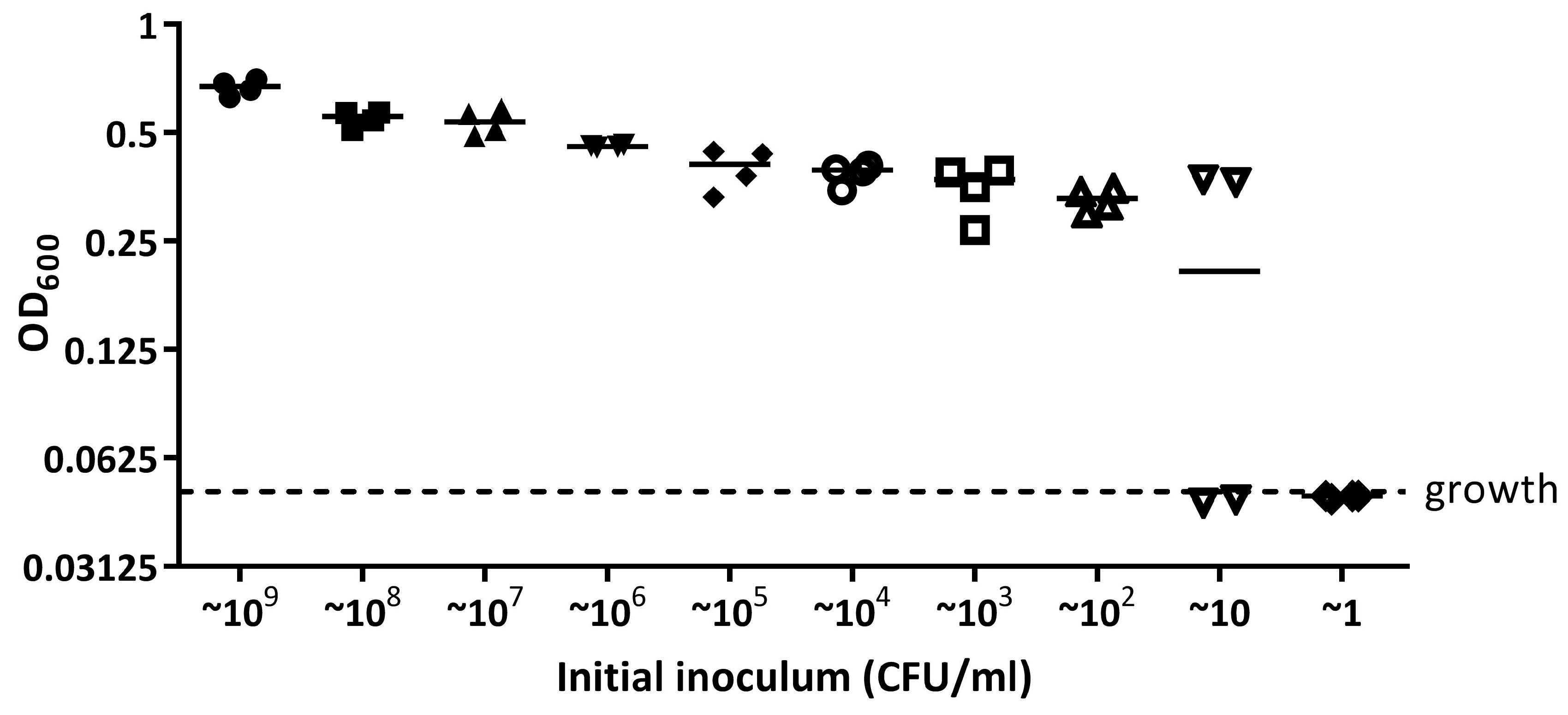
Appendix A.1.2. The Limit of Detection for E. coli 25922 lux and S. aureus 6538 Recovered in a 96 Well Plate Is 100 CFU/mL


References
- WHO. Antimicrobial Resistance: Global Report on Surveillance; WHO Press: Geneva, Switzerland, 2014. [Google Scholar]
- Dancer, S.J. Controlling hospital-acquired infection: Focus on the role of the environment and new technologies for decontamination. Clin. Microbiol. Rev. 2014, 27, 665–690. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.J.; Anderson, D.; Rutala, W.A. The role of the surface environment in healthcare-associated infections. Curr. Opin. Infect. Dis. 2013, 26, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Schwebke, I.; Kampf, G. How long do nosocomial pathogens persist on inanimate surfaces? A systematic review. BMC Infect. Dis. 2006, 6, 130. [Google Scholar] [CrossRef] [PubMed]
- Stiefel, U.; Cadnum, J.L.; Eckstein, B.C.; Guerrero, D.M.; Tima, M.A.; Donskey, C.J. Contamination of hands with methicillin-resistant Staphylococcus aureus after contact with environmental surfaces and after contact with the skin of colonized patients. Infect. Control Hosp. Epidemiol. 2011, 32, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Page, K.; Wilson, M.; Parkin, I. Antimicrobial surfaces and their potential in reducing the role of the inanimate environment in the incidence of hospital-acquired infections. J. Mater. Chem. 2009, 19, 3819–3831. [Google Scholar] [CrossRef]
- Scott, E.; Bloomfield, S.F. The survival and transfer of microbial contamination via cloths, hands and utensils. J. Appl. Bacteriol. 1990, 68, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Rusin, P.; Maxwell, S.; Gerba, C. Comparative surface-to-hand and fingertip-to-mouth transfer efficiency of gram-positive bacteria, gram-negative bacteria, and phage. J. Appl. Microbiol. 2002, 93, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Gerhardts, A.; Hammer, T.R.; Balluff, C.; Mucha, H.; Hoefer, D. A model of the transmission of micro-organisms in a public setting and its correlation to pathogen infection risks. J. Appl. Microbiol. 2012, 112, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Otter, J.A.; Yezli, S.; French, G.L. The role played by contaminated surfaces in the transmission of nosocomial pathogens. Infect. Control Hosp. Epidemiol. 2011, 32, 687–699. [Google Scholar] [CrossRef] [PubMed]
- French, G.L.; Otter, J.A.; Shannon, K.P.; Adams, N.M.T.; Watling, D.; Parks, M.J. Tackling contamination of the hospital environment by methicillin-resistant Staphylococcus aureus (MRSA): A comparison between conventional terminal cleaning and hydrogen peroxide vapour decontamination. J. Hosp. Infect. 2004, 57, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, A.; Pultz, N.J.; Gries, D.M.; Ray, A.J.; Eckstein, E.C.; Aron, D.C.; Donskey, C.J. Acquisition of nosocomial pathogens on hands after contact with environmental surfaces near hospitalized patients. Infect. Control Hosp. Epidemiol. 2004, 25, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.P.; Avery, L.M.; Killham, K.; Jones, D.L. Persistence of Escherichia coli O157 on farm surfaces under different environmental conditions. J. Appl. Microbiol. 2005, 98, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, B.; Cerf, O. Review—Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 2011, 145, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Otter, J.A.; Vickery, K.; Walker, J.T.; de Lancey Pulcini, E.; Stoodley, P.; Goldenberg, S.D.; Salkeld, J.A.G.; Chewins, J.; Yezli, S.; Edgeworth, J.D. Surface-attached cells, biofilms and biocide susceptibility: Implications for hospital cleaning and disinfection. J. Hosp. Infect. 2015, 89, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, F.H. Processes controlling the transmission of bacterial pathogens in the environment. Res. Microbiol. 2007, 158, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Van Houdt, R.; Michiels, C.W. Biofilm formation and the food industry, a focus on the bacterial outer surface. J. Appl. Microbiol. 2010, 109, 1117–1131. [Google Scholar] [CrossRef] [PubMed]
- Hota, B. Contamination, disinfection, and cross-colonization: Are hospital surfaces reservoirs for nosocomial infection? Clin. Infect. Dis. 2004, 39, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, D.M.; Nerandzic, M.M.; Jury, L.A.; Jinno, S.; Chang, S.; Donskey, C.J. Acquisition of spores on gloved hands after contact with the skin of patients with Clostridium difficile infection and with environmental surfaces in their rooms. Am. J. Infect. Control 2012, 40, 556–558. [Google Scholar] [CrossRef] [PubMed]
- Todd, E.C.D.; Greig, J.D.; Bartleson, C.A.; Michaels, B.S. Outbreaks where food workers have been implicated in the spread of foodborne disease. Part 5. Sources of contamination and pathogen excretion from infected persons. J. Food Prot. 2008, 71, 2582–2595. [Google Scholar] [CrossRef] [PubMed]
- Malik, R.E.; Cooper, R.A.; Griffith, C.J. Use of audit tools to evaluate the efficacy of cleaning systems in hospitals. Am. J. Infect. Control 2003, 31, 181–187. [Google Scholar] [CrossRef] [PubMed]
- French, G.L. The continuing crisis in antibiotic resistance. Int. J. Antimicrob. Agents 2010, 36, S3–S7. [Google Scholar] [CrossRef]
- Robine, E.; Boulangé-Petermann, L.; Derangère, D. Assessing bactericidal properties of materials: The case of metallic surfaces in contact with air. J. Microbiol. Methods 2002, 49, 225–234. [Google Scholar] [CrossRef]
- Escalada, M.G.; Russell, A.D.; Maillard, J.-Y.; Ochs, D. Triclosan-bacteria interactions: Single or multiple target sites? Lett. Appl. Microbiol. 2005, 41, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Ojeil, M.; Jermann, C.; Holah, J.; Denyer, S.P.; Maillard, J.-Y. Evaluation of new in vitro efficacy test for antimicrobial surface activity reflecting UK hospital conditions. J. Hosp. Infect. 2013, 85, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Suppakul, P.; Miltz, J.; Sonneveld, K.; Bigger, S.W. Active packaging technologies with an emphasis on antimicrobial packaging and its applications. J. Food Sci. 2003, 68, 408–420. [Google Scholar] [CrossRef] [Green Version]
- Bastarrachea, L.J.; Denis-Rohr, A.; Goddard, J.M. Antimicrobial food equipment coatings: Applications and challenges. Annu. Rev. Food Sci. Technol. 2015, 6, 97–118. [Google Scholar] [CrossRef] [PubMed]
- Green, J.-B.D.; Fulghum, T.; Nordhaus, M. Review of immobilized antimicrobial agents and methods for testing. Biointerphases 2011, 6, CL2–CL43. [Google Scholar] [CrossRef] [PubMed]
- Siedenbiedel, F.; Tiller, J.C. Antimicrobial polymers in solution and on surfaces: Overview and functional principles. Polymers 2012, 4, 46–71. [Google Scholar] [CrossRef]
- Cooksey, K. Effectiveness of antimicrobial food packaging materials. Food Addit. Contam. 2005, 22, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Hasan, J.; Crawford, R.J.; Ivanova, E.P. Antibacterial surfaces: The quest for a new generation of biomaterials. Trends Biotechnol. 2013, 31, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Bastarrachea, L.J.; Dhawan, S.; Sablani, S.S. Engineering Properties of Polymeric-Based Antimicrobial Films for Food Packaging: A Review. Food Eng. Rev. 2011, 3, 79–93. [Google Scholar] [CrossRef]
- Vasilev, K.; Cook, J.; Griesser, H.J. Antibacterial surfaces for biomedical devices. Expert Rev. Med. Devices 2009, 6, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.; Soriano, E.; López-Carballo, G.; Picouet, P.; Lloret, E.; Gavara, R.; Hernández-Muñoz, P. Preservation of aseptic conditions in absorbent pads by using silver nanotechnology. Food Res. Int. 2009, 42, 1105–1112. [Google Scholar] [CrossRef]
- Humphreys, H. Self-disinfecting and microbiocide-impregnated surfaces and fabrics: What potential in interrupting the spread of healthcare-associated infection? Clin. Infect. Dis. 2014, 58, 848–853. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.; Spencer, M.; Edmiston, C. Role of healthcare apparel and other healthcare textiles in the transmission of pathogens: A review of the literature. J. Hosp. Infect. 2015, 90, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Weis, T.L.; Schurr, M.J.; Faith, N.G.; Czuprynski, C.J.; McAnulty, J.F.; Murphy, C.J.; Abbott, N.L. Surfaces modified with nanometer-thick silver-impregnated polymeric films that kill bacteria but support growth of mammalian cells. Biomaterials 2010, 31, 680–690. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Hu, Y.; Li, J.; Li, B. Chitosan/phosvitin antibacterial films fabricated via layer-by-layer deposition. Int. J. Biol. Macromol. 2014, 64, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cen, L.; Neoh, K.G.; Kang, E.T. Antibacterial activity of cloth functionalized with N-alkylated poly(4-vinylpyridine). J. Biomed. Mater. Res. 2004, 71, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Ohl, M.; Schweizer, M.; Graham, M.; Heilmann, K.; Boyken, L.; Diekema, D. Hospital privacy curtains are frequently and rapidly contaminated with potentially pathogenic bacteria. Am. J. Infect. Control 2012, 40, 904–906. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, M.; Graham, M.; Ohl, M.; Heilmann, K.; Boyken, L.; Diekema, D. Novel Hospital Curtains with Antimicrobial Properties: A Randomized, Controlled Trial. Infect. Control Hosp. Epidemiol. 2012, 33, 1081–1085. [Google Scholar] [CrossRef] [PubMed]
- Lemmen, S.; Scheithauer, S.; Haefner, H.; Yezli, S.; Mohr, M.; Otter, J.A. Evaluation of hydrogen peroxide vapor for the inactivation of nosocomial pathogens on porous and nonporous surfaces. Am. J. Infect. Control 2015, 43, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Gizdavic-Nikolaidis, M.R.; Ray, S.; Bennett, J.R.; Swift, S.; Bowmaker, G.A.; Easteal, A.J. Electrospun poly(aniline-co-ethyl 3-aminobenzoate)/poly(lactic acid) nanofibers and their potential in biomedical applications. J. Polym. Sci. Part A Polym. Chem. 2011, 49, 4902–4910. [Google Scholar] [CrossRef]
- Pandey, S.; Annapoorni, S.; Malhotra, B.D. Synthesis and Characterization of Poly(aniline-co-o-anisidine): A Processable Conducting Copolymer. Macromolecules 1993, 26, 3190–3193. [Google Scholar] [CrossRef]
- Gizdavic-Nikolaidis, M.R.; Bennett, J.R.; Swift, S.; Easteal, A.J.; Ambrose, M. Broad spectrum antimicrobial activity of functionalized polyanilines. Acta Biomater. 2011, 7, 4204–4209. [Google Scholar] [CrossRef] [PubMed]
- Gizdavic-Nikolaidis, M.R.; Bennett, J.R.; Zujovic, Z.; Swift, S.; Bowmaker, G.A. Characterization and antimicrobial efficacy of acetone extracted aniline oligomers. Synth. Met. 2012, 162, 1114–1119. [Google Scholar] [CrossRef]
- Dhand, C.; Das, M.; Datta, M.; Malhotra, B.D. Recent advances in polyaniline based biosensors. Biosens. Bioelectron. 2011, 26, 2811–2821. [Google Scholar] [CrossRef] [PubMed]
- Grennan, K.; Killard, A.J.; Hanson, C.J.; Cafolla, A.A.; Smyth, M.R. Optimisation and characterisation of biosensors based on polyaniline. Talanta 2006, 68, 1591–1600. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Jiang, T.; Hu, P.; Han, Z.; Lu, X.; Ye, P. Self-decontaminating properties of fluorinated copolymers integrated with ciprofloxacin for synergistically inhibiting the growth of Escherichia coli. J. Biomater. Sci. Polym. Ed. 2014, 25, 1920–1945. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.L.; Fan, W.J.; Shan, S.Y.; Hu, T.W.; Wang, Y.M.; Jia, Q.M. The Effects of Natural Dopant Acids on Morphologies and Antibacterial Activity of Polyaniline. Adv. Mater. Res. 2013, 650, 249–252. [Google Scholar] [CrossRef]
- Humpolicek, P.; Kasparkova, V.; Saha, P.; Stejskal, J. Biocompatibility of polyaniline. Synth. Met. 2012, 162, 722–727. [Google Scholar] [CrossRef]
- Qi, H.; Liu, M.; Xu, L.; Feng, L.; Tao, L.; Ji, Y.; Zhang, X.; Wei, Y. Biocompatibility evaluation of aniline oligomers with different end-functional groups. Toxicol. Res. 2013, 2, 427–433. [Google Scholar] [CrossRef]
- Seshadri, D.T.; Bhat, N.V. Use of polyaniline as an antimicrobial agent in textiles. Indian J. Fibre Text. Res. 2005, 30, 204–206. [Google Scholar]
- Gizdavic-Nikolaidis, M.R.; Pagnon, J.C.; Ali, N.; Sum, R.; Davies, N.; Roddam, L.F.; Ambrose, M. Functionalized polyanilines disrupt Pseudomonas aeruginosa and Staphylococcus aureus biofilms. Colloids Surf. B. Biointerfaces 2015, 136, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48 (Suppl. 1), 5–16. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Siani, H.; Maillard, J.-Y. Best practice in healthcare environment decontamination. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kram, K.E.; Finkel, S.E. Rich Medium Composition Affects Escherichia coli Survival, Glycation, and Mutation Frequency during Long-Term Batch Culture. Appl. Environ. Microbiol. 2015, 81, 4442–4450. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Bausch, C.; Richmond, C.; Blattner, F.R.; Conway, T. Functional genomics: Expression analysis of Escherichia coli growing on minimal and rich media. J. Bacteriol. 1999, 181, 6425–6440. [Google Scholar] [PubMed]
- Zhang, J.; Greasham, R. Chemically defined media for commercial fermentations. Appl. Microbiol. Biotechnol. 1999, 51, 407–421. [Google Scholar] [CrossRef]
- Sezonov, G.; Joseleau-Petit, D.; D’Ari, R. Escherichia coli physiology in Luria-Bertani broth. J. Bacteriol. 2007, 189, 8746–8749. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.; Schellhorn, H.E. Control of RpoS in global gene expression of Escherichia coli in minimal media. Mol. Genet. Genom. 2009, 281, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.; Dalton, J.; Wiles, S.; Gizdavic-Nikolaidis, M.; Swift, S. The tuberculocidal activity of polyaniline and functionalised polyanilines. PeerJ 2016, 4, e2795. [Google Scholar] [CrossRef] [PubMed]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Liu, J.; Liu, C.; Liu, Y.; Chen, M.; Hu, Y.; Yang, Z. Study on the grafting of chitosan-gelatin microcapsules onto cotton fabrics and its antibacterial effect. Colloids Surf. B Biointerfaces 2013, 109, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Andreu, N.; Zelmer, A.; Fletcher, T.; Elkington, P.T.; Ward, T.H.; Ripoll, J.; Parish, T.; Bancroft, G.J.; Schaible, U.; Robertson, B.D.; et al. Optimisation of bioluminescent reporters for use with mycobacteria. PLoS ONE 2010, 5, e10777. [Google Scholar] [CrossRef] [PubMed]
- Andreu, N.; Zelmer, A.; Sampson, S.L.; Ikeh, M.; Bancroft, G.J.; Schaible, U.E.; Wiles, S.; Robertson, B.D. Rapid in vivo assessment of drug efficacy against Mycobacterium tuberculosis using an improved firefly luciferase. J. Antimicrob. Chemother. 2013, 68, 2118–2127. [Google Scholar] [CrossRef] [PubMed]
- Andreu, N.; Zelmer, A.; Wiles, S. Noninvasive biophotonic imaging for studies of infectious disease. FEMS Microbiol. Rev. 2011, 35, 360–394. [Google Scholar] [CrossRef] [PubMed]
- Appendini, P.; Hotchkiss, J.H. Review of antimicrobial food packaging. Innov. Food Sci. Emerg. Technol. 2002, 3, 113–126. [Google Scholar] [CrossRef]
- Japanese Standards Association. JIS Z 2801:2000 Antimicrobial Products—Test for Antimicrobial Activity and Efficacy; Japanese Standards Association: Tokyo, Japan, 2000. [Google Scholar]
- Humphreys, P.N. Testing standards for sporicides. J. Hosp. Infect. 2011, 77, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Warnes, S.L.; Keevil, C.W. Lack of Involvement of Fenton Chemistry in Death of Methicillin-Resistant and Methicillin-Sensitive Strains of Staphylococcus aureus and Destruction of Their Genomes on Wet or Dry Copper Alloy Surfaces. Appl. Environ. Microbiol. 2016, 82, 2132–2136. [Google Scholar] [CrossRef] [PubMed]
- Lobritz, M.A.; Belenky, P.; Porter, C.B.M.; Gutierrez, A.; Yang, J.H.; Schwarz, E.G.; Dwyer, D.J.; Khalil, A.S.; Collins, J.J. Antibiotic efficacy is linked to bacterial cellular respiration. Proc. Natl. Acad. Sci. USA 2015, 112, 8173–8180. [Google Scholar] [CrossRef] [PubMed]
- Gizdavic-Nikolaidis, M.R.; Easteal, A.J.; Stepanovic, S. Bioactive Aniline Copolymers. International Patent Application No. PCT/NZ2008/000254, 26 September 2008. [Google Scholar]
- Gizdavic-Nikolaidis, M.R.; Bowmaker, G.A.; Zujovic, Z. The Synthesis, Physical Properties, Bioactivity and Potential Applications of Polyanilines; Cambridge Scholars Publishing: Newcastle, UK, 2018; in press. [Google Scholar]
- Fröls, S.; Dyall-Smith, M.; Pfeifer, F. Biofilm formation by haloarchaea. Environ. Microbiol. 2012, 14, 3159–3174. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.; Guo, X.; Jing, H.; Gong, J.; Sun, C.; Yang, K. Antibacterial effect of the conducting polyaniline. J. Mater. Sci. Technol. 2006, 22, 289–290. [Google Scholar]
- Tan, C.; Smith, R.P.; Srimani, J.K.; Riccione, K.A.; Prasada, S.; Kuehn, M.; You, L. The inoculum effect and band-pass bacterial response to periodic antibiotic treatment. Mol. Syst. Biol. 2012, 8, 617. [Google Scholar] [CrossRef] [PubMed]
- Warnes, S.L.; Green, S.M.; Michels, H.T.; Keevil, C.W. Biocidal efficacy of copper alloys against pathogenic enterococci involves degradation of genomic and plasmid DNAs. Appl. Environ. Microbiol. 2010, 76, 5390–5401. [Google Scholar] [CrossRef] [PubMed]
- Oie, S.; Hosokawa, I.; Kamiya, A. Contamination of room door handles by methicillin-sensitive/methicillin-resistant Staphylococcus aureus. J. Hosp. Infect. 2002, 51, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Oie, S.; Suenaga, S.; Sawa, A.; Kamiya, A. Association between isolation sites of methicillin-resistant Staphylococcus aureus (MRSA) in patients with MRSA-positive body sites and MRSA contamination in their surrounding environmental surfaces. Jpn. J. Infect. Dis. 2007, 60, 367–369. [Google Scholar] [PubMed]
- Schmidt, M.G.; Attaway, H.H.; Sharpe, P.A.; John, J.F.; Sepkowitz, K.A.; Morgan, A.; Fairey, S.E.; Singh, S.; Steed, L.L.; Cantey, J.R.; et al. Sustained reduction of microbial burden on common hospital surfaces through introduction of copper. J. Clin. Microbiol. 2012, 50, 2217–2223. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, J.; Humphreys, H. Application of copper to prevent and control infection. Where are we now? J. Hosp. Infect. 2012, 81, 217–223. [Google Scholar] [CrossRef]
- Noyce, J.O.; Michels, H.T.; Keevil, C.W. Use of copper cast alloys to control Escherichia coli O157 cross-contamination during food processing. Appl. Environ. Microbiol. 2006, 72, 4239–4244. [Google Scholar] [CrossRef] [PubMed]
- Rabie, A.J.; McLaren, I.M.; Breslin, M.F.; Sayers, R.; Davies, R.H. Assessment of anti-Salmonella activity of boot dip samples. Avian Pathol. 2015, 44, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Fraise, A. Currently available sporicides for use in healthcare, and their limitations. J. Hosp. Infect. 2011, 77, 210–212. [Google Scholar] [CrossRef] [PubMed]
- Honney, C. Colonisation Resistant Surfaces. MSc Thesis, University of Auckland, Auckland, New Zealand, 2014. [Google Scholar]
- Møretrø, T.; Høiby-pettersen, G.S.; Habimana, O.; Heir, E.; Langsrud, S. Assessment of the antibacterial activity of a triclosan-containing cutting board. Int. J. Food Microbiol. 2011, 146, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Cutter, C.N. The effectiveness of triclosan-incorporated plastic against bacteria on beef surfaces. J. Food Prot. 1999, 62, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Kalyon, B.D.; Olgun, U. Antibacterial efficacy of triclosan-incorporated polymers. Am. J. Infect. Control 2001, 29, 124–125. [Google Scholar] [CrossRef] [PubMed]
- Connolly, P.; Bloomfield, S.F.; Denyer, S.P. The use of impedance for preservative efficacy testing of pharmaceuticals and cosmetic products. J. Appl. Bacteriol. 1994, 76, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.T. Development of a Series of Bioluminescent Sensor Strains for Rapid Screening of Antimicrobials. Ph.D. Thesis, University of Auckland, Auckland, New Zealand, 2013. [Google Scholar]
- Riedel, C.U.; Casey, P.G.; Mulcahy, H.; O’Gara, F.; Gahan, C.G.M.; Hill, C. Construction of p16Slux, a novel vector for improved bioluminescent labeling of gram-negative bacteria. Appl. Environ. Microbiol. 2007, 73, 7092–7095. [Google Scholar] [CrossRef] [PubMed]
- Seaver, L.C.; Imlay, J.A. Are respiratory enzymes the primary sources of intracellular hydrogen peroxide? J. Biol. Chem. 2004, 279, 48742–48750. [Google Scholar] [CrossRef] [PubMed]
- Brewster, J.D. A simple micro-growth assay for enumerating bacteria. J. Microbiol. Methods 2003, 53, 77–86. [Google Scholar] [CrossRef]
- Pankey, G.A.; Sabath, L.D. Clinical relevance of bacteriostatic versus bactericidal mechanisms of action in the treatment of Gram-positive bacterial infections. Clin. Infect. Dis. 2004, 38, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Meighen, E. Genetics of bacterial bioluminescence. Annu. Rev. Genet. 1994, 28, 117–139. [Google Scholar] [CrossRef] [PubMed]











© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robertson, J.; Gizdavic-Nikolaidis, M.; Swift, S. Investigation of Polyaniline and a Functionalised Derivative as Antimicrobial Additives to Create Contamination Resistant Surfaces. Materials 2018, 11, 436. https://doi.org/10.3390/ma11030436
Robertson J, Gizdavic-Nikolaidis M, Swift S. Investigation of Polyaniline and a Functionalised Derivative as Antimicrobial Additives to Create Contamination Resistant Surfaces. Materials. 2018; 11(3):436. https://doi.org/10.3390/ma11030436
Chicago/Turabian StyleRobertson, Julia, Marija Gizdavic-Nikolaidis, and Simon Swift. 2018. "Investigation of Polyaniline and a Functionalised Derivative as Antimicrobial Additives to Create Contamination Resistant Surfaces" Materials 11, no. 3: 436. https://doi.org/10.3390/ma11030436