Molecular Profiling and Optimization Studies for Growth and PHB Production Conditions in Rhodobacter sphaeroides

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Growth Conditions
2.2. PHB Extraction and Analysis
2.3. HPLC Analysis for Organic Acids
2.4. RNA Isolation and Transcript Analysis by qRT-PCR
2.5. Determination of H2O2 Concentration and Peroxidase Activity
2.6. Total Carotenoids Extraction
2.7. Statistical Analysis
3. Results
3.1. Effects of Light and Aeration on PHB Production
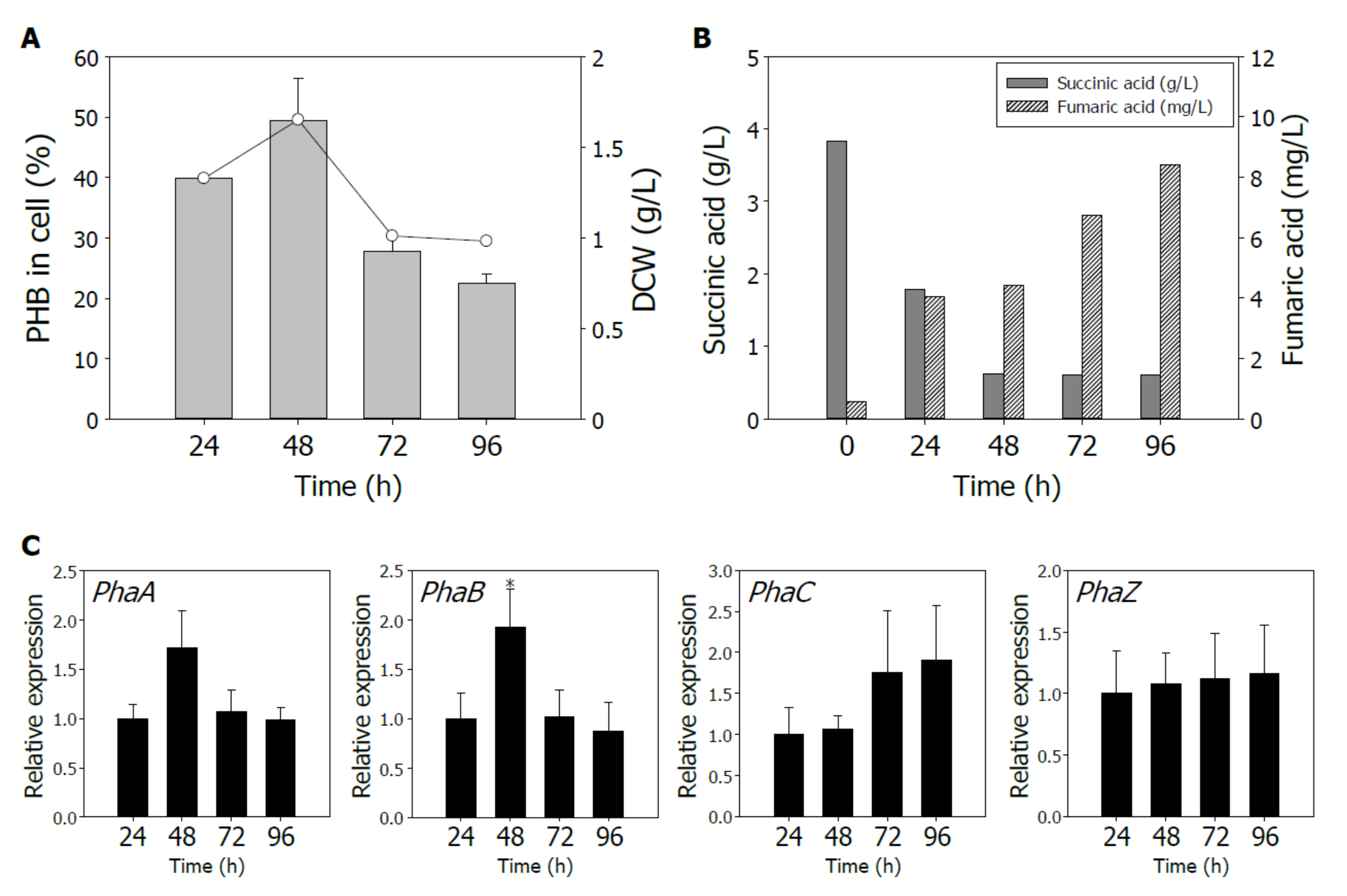
3.2. PHB Production According to Time-Course
3.3. Effects of Temperature on PHB Production
3.4. Effects of C/N Ratio in Growth Medium on PHB Production
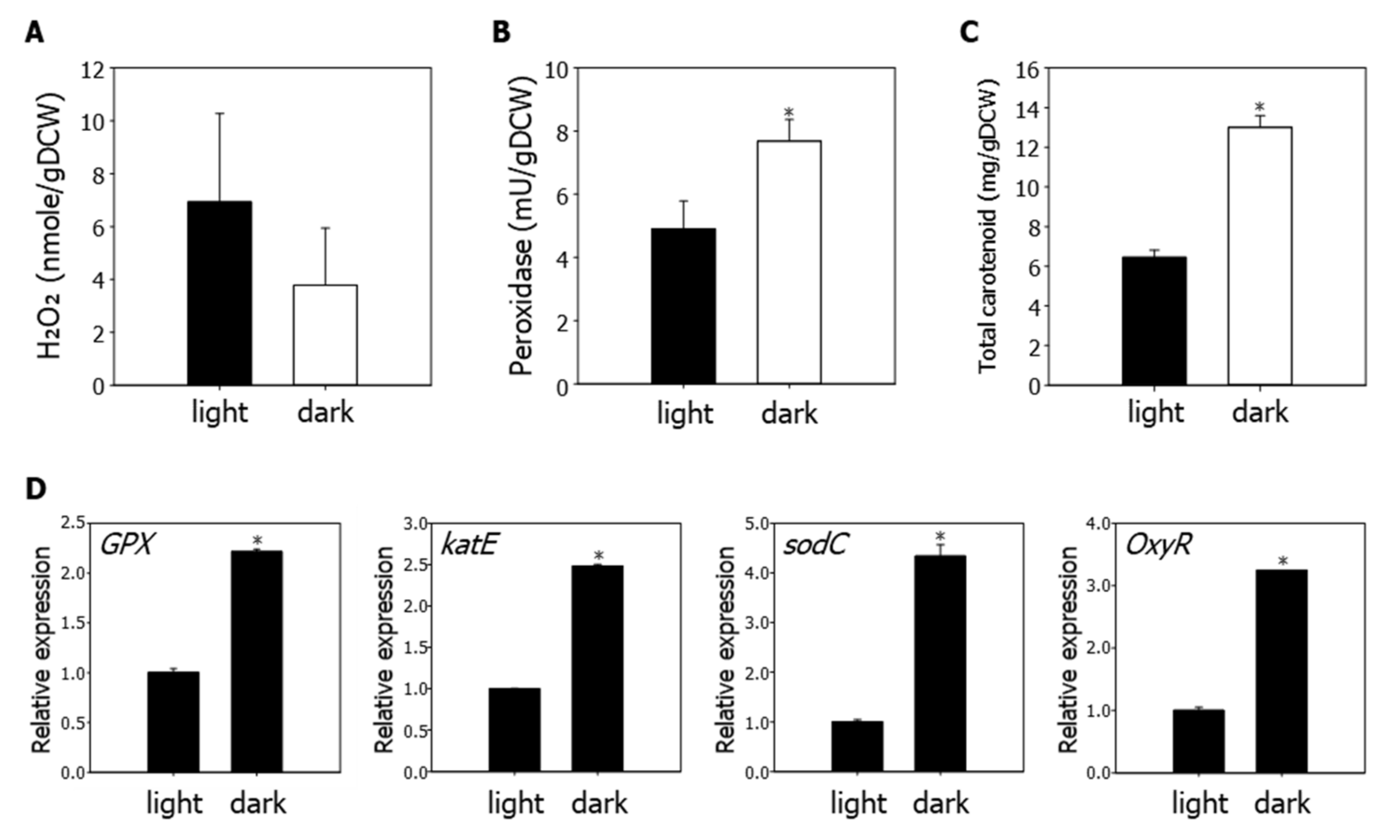
3.5. ROS Regulation in R. sphaeroides
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Luengo, J.M.; García, B.; Sandoval, A.; Naharro, G.; Olivera, E.R. Bioplastics from microorganisms. Curr. Opin. Microbiol. 2003, 6, 251–260. [Google Scholar] [CrossRef]
- Reddy, C.S.K.; Ghai, R.; Rashmi; Kalia, V.C. Polyhydroxyalkanoates: An overview. Bioresour. Technol. 2003, 87, 137–146. [Google Scholar] [CrossRef]
- Grothe, E.; Moo-Young, M.; Chisti, Y. Fermentation optimization for the production of poly(β-hydroxybutyric acid) microbial thermoplastic. Enzym. Microb. Technol. 1999, 25, 132–141. [Google Scholar] [CrossRef] [Green Version]
- García-Torreiro, M.; Lu-Chau, T.A.; Lema, J.M. Effect of nitrogen and/or oxygen concentration on poly(3-hydroxybutyrate) accumulation by Halomonas boliviensis. Bioprocess Biosyst. Eng. 2016, 39, 1365–1374. [Google Scholar] [CrossRef]
- Chou, M.E.; Chang, W.T.; Chang, Y.C.; Yang, M.K. Expression of four pha genes involved in poly-β-hydroxybutyrate production and accumulation in Rhodobacter sphaeroides FJ1. Mol. Genet. Genom. 2009, 282, 97–106. [Google Scholar] [CrossRef]
- Steinbüchel, A.; Hustede, E.; Liebergesell, M.; Pieper, U.; Timm, A.; Valentin, H. Molecular basis for biosynthesis and accumulation of polyhydroxyalkanoic acids in bacteria. FEMS Microbiol. Rev. 1992, 103, 217–230. [Google Scholar] [CrossRef]
- Lenz, R.W.; Marchessault, R.H. Bacterial polyesters: Biosynthesis, biodegradable plastics and biotechnology. Biomacromolecules 2005, 6, 1–8. [Google Scholar] [CrossRef]
- Mahishi, L.H.; Rawal, S.K. Effect of amino acid smentation on the synthesis of poly(3-hydroxybutyrate) by recombinant phaSa+ Escherichia coli. World J. Microbiol. Biotechnol. 2002, 18, 805–810. [Google Scholar] [CrossRef]
- Mahishi, L.H.; Tripathi, G.; Rawal, S.K. Poly(3-hydroxybutyrate) (PHB) synthesis by recombinant Escherichia coli harbouring Streptomyces aureofaciens PHB biosynthesis genes: Effect of various carbon and nitrogen sources. Microbiol. Res. 2003, 158, 19–27. [Google Scholar] [CrossRef]
- Kang, D.K.; Lee, C.R.; Lee, S.H.; Bae, J.H.; Park, Y.K.; Rhee, Y.H.; Sung, B.H.; Sohn, J.H. Production of polyhydroxyalkanoates from sludge palm oil using Pseudomonas putida S12. J. Microbiol. Biotechnol. 2017, 27, 990–994. [Google Scholar] [CrossRef] [Green Version]
- Song, J.H.; Jeon, C.O.; Choi, M.H.; Yoon, S.C.; Park, W. Polyhydroxyalkanoate (PHA) production using waste vegetable oil by Pseudomonas sp. Strain DR2. J. Microbiol. Biotechnol. 2008, 18, 1408–1415. [Google Scholar] [PubMed]
- Khanna, S.; Srivastava, A.K. Statistical media optimization studies for growth and PHB production by Ralstonia eutropha. Process Biochem. 2005, 40, 2173–2182. [Google Scholar] [CrossRef]
- Riedel, S.L.; Jahns, S.; Koenig, S.; Bock, M.C.E.; Brigham, C.J.; Bader, J.; Stahl, U. Polyhydroxyalkanoates production with Ralstonia eutropha from low quality waste animal fats. J. Biotechnol. 2015, 214, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Revelles, O.; Beneroso, D.; Menéndez, J.A.; Arenillas, A.; García, J.L.; Prieto, M.A. Syngas obtained by microwave pyrolysis of household wastes as feedstock for polyhydroxyalkanoate production in Rhodospirillum rubrum. Microb. Biotechnol. 2017, 10, 1412–1417. [Google Scholar] [CrossRef] [Green Version]
- Karmann, S.; Panke, S.; Zinn, M. Fed-batch cultivations of Rhodospirillum rubrum under multiple nutrient-limited growth conditions on syngas as a novel option to produce poly(3-hydroxybutyrate) (PHB). Front. Bioeng. Biotechnol. 2019, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Brandl, H.; Gross, R.A.; Lenz, R.W.; Lloyd, R.; Fuller, R.C. The accumulation of poly(3-hydroxyalkanoates) in Rhodobacter sphaeroides. Arch. Microbiol. 1991, 155, 337–340. [Google Scholar] [CrossRef]
- Sangkharak, K.; Prasertsan, P. Nutrient optimization for production of polyhydroxybutyrate from halotolerant photosynthetic bacteria cultivated under aerobic-dark condition. Electron. J. Biotechnol. 2008, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, J.; Kondo, A. Disruption of poly (3-hydroxyalkanoate) depolymerase gene and overexpression of three poly (3-hydroxybutyrate) biosynthetic genes improve poly (3-hydroxybutyrate) production from nitrogen rich medium by Rhodobacter sphaeroides. Microb. Cell Fact. 2019, 18, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.K.; Lin, Y.C.; Shen, C.H. Identification of two gene loci involved in poly-beta-hydroxybutyrate production in Rhodobacter sphaeroides FJ1. J. Microbiol. Immunol. Infect. 2006, 39, 18–27. [Google Scholar]
- Chou, M.E.; Yang, M.K. Analyses of binding sequences of the PhaR protein of Rhodobacter sphaeroides FJ1. FEMS Microbiol. Lett. 2010, 302, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Zeilstra-Ryalls, J.H.; Kaplan, S. Oxygen intervention in the regulation of gene expression: The photosynthetic bacterial paradigm. Cell. Mol. Life Sci. 2004, 61, 417–436. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, S.; Eraso, J.; Roh, J.H. Interacting regulatory networks in the facultative photosynthetic bacterium, Rhodobacter sphaeroides 2.4.1. Biochem. Soc. Trans. 2005, 33, 51–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegelhoffer, E.C.; Donohue, T.J. Bacterial responses to photo-oxidative stress. Nat. Rev. Microbiol. 2009, 7, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Yuan, J.; Gao, H. Microbial oxidative stress response: Novel insights from environmental facultative anaerobic bacteria. Arch. Biochem. Biophys. 2015, 584, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Abee, T.; Wouters, J.A. Microbial stress response in minimal processing. Int. J. Food Microbiol. 1999, 50, 65–91. [Google Scholar] [CrossRef]
- Licht, M.K.; Nuss, A.M.; Volk, M.; Konzer, A.; Beckstette, M.; Berghoff, B.A.; Klug, G. Adaptation to photooxidative stress: Common and special strategies of the alphaproteobacteria Rhodobacter sphaeroides and Rhodobacter capsulatus. Microorganisms 2020, 8, 283. [Google Scholar] [CrossRef] [Green Version]
- Glaeser, J.; Klug, G. Photo-oxidative stress in Rhodobacter sphaeroides: Protective role of carotenoids and expression of selected genes. Microbiology 2005, 151, 1927–1938. [Google Scholar] [CrossRef] [Green Version]
- Zeller, T.; Klug, G. Detoxification of hydrogen peroxide and expression of catalase genes in Rhodobacter. Microbiology 2004, 150, 3451–3462. [Google Scholar] [CrossRef] [Green Version]
- Kho, D.H.; Yoo, S.B.; Kim, J.S.; Kim, E.J.; Lee, J.K. Characterization of Cu- and Zn-containing superoxide dismutase of Rhodobacter sphaeroides. FEMS Microbiol. Lett. 2004, 234, 261–267. [Google Scholar] [CrossRef]
- Uchoa, A.F.; Knox, P.P.; Turchielle, R.; Seifullina, N.K.; Baptista, M.S. Singlet oxygen generation in the reaction centers of Rhodobacter sphaeroides. Eur. Biophys. J. 2008, 37, 843–850. [Google Scholar] [CrossRef]
- Zeller, T.; Mraheil, M.A.; Moskvin, O.V.; Li, K.; Gomelsky, M.; Klug, G. Regulation of hydrogen peroxide-dependent gene expression in Rhodobacter sphaeroides: Regulatory functions of QxyR. J. Bacteriol. 2007, 189, 3784–3792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sistrom, W.R. The kinetics of the synthesis of photopigments in Rhodopseudomonas sphaeroides. J. Gen. Microbiol. 1962, 28, 607–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Lee, H.J.; Jung, J.H.; Park, C.M. The Arabidopsis thaliana RNA-binding protein FCA regulates thermotolerance by modulating the detoxification of reactive oxygen species. New Phytol. 2015, 205, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, S.G.; Park, C.M. Salicylic acid promotes seed germination under high salinity by modulating antioxidant activity in Arabidopsis. New Phytol. 2010, 188, 626–637. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Modak, H.V.; Tabita, F.R. Photolithoautotrophic growth and control of CO2 fixation in Rhodobacter sphaeroides and Rhodospirillum rubrum in the absence of ribulose bisphosphate carboxylase-oxygenase. J. Bacteriol. 1993, 175, 7109–7114. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Kim, D.H.; Cha, J. Culture conditions affecting H2 production by phototrophic bacterium Rhodobacter sphaeroides KD131. Int. J. Hydrog. Energy 2012, 37, 14055–14061. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, D.H.; Cha, J.; Lee, J.K. Effect of carbon and nitrogen sources on photo-fermentative H2 production associated with nitrogenase, uptake hydrogenase activity, and PHB accumulation in Rhodobacter sphaeroides KD131. Bioresour. Technol. 2012, 116, 179–183. [Google Scholar] [CrossRef]
- Kemavongse, K.; Prasertsan, P.; Upaichit, A.; Methacanon, P. Poly-β-hydroxyalkanoate production by halotolerant Rhodobacter sphaeroides U7. World J. Microbiol. Biotechnol. 2008, 24, 2073–2085. [Google Scholar] [CrossRef]
- Sangkharak, K.; Prasertsan, P. Optimization of polyhydroxybutyrate production from a wild type and two mutant strains of Rhodobacter sphaeroides using statistical method. J. Biotechnol. 2007, 132, 331–340. [Google Scholar] [CrossRef]
- McEwan, A.G. Photosynthetic electron transport and anaerobic metabolism in purple non-sulfur phototrophic bacteria. Antonie Leeuwenhoek 1994, 66, 151–164. [Google Scholar] [CrossRef]
- Morita, R.Y. Bioavailability of energy and its relationship to growth and starvation survival in nature. Can. J. Microbiol. 1988, 34, 436–441. [Google Scholar] [CrossRef]
- Mullineaux, P.M.; Exposito-Rodriguez, M.; Laissue, P.P.; Smirnoff, N. ROS-dependent signalling pathways in plants and algae exposed to high light: Comparisons with other eukaryotes. Free Radic. Biol. Med. 2018, 122, 52–64. [Google Scholar] [CrossRef]
- Nantapong, N.; Murata, R.; Trakulnaleamsai, S.; Kataoka, N.; Yakushi, T.; Matsushita, K. The effect of reactive oxygen species (ROS) and ROS-scavenging enzymes, superoxide dismutase and catalase, on the thermotolerant ability of Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2019, 103, 5355–5366. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, J.; Ríos-Momberg, M.; Hewitt, D.; Hansberg, W. Reactive oxygen species and development in microbial eukaryotes. Trends Microbiol. 2005, 13, 111–118. [Google Scholar] [CrossRef]
- Wong, M.T.; Cheng, D.; Wang, R.; Hsing, I.M. Modifying the endogenous electron fluxes of Rhodobacter sphaeroides 2.4.1 for improved electricity generation. Enzym. Microb. Technol. 2016, 86, 45–51. [Google Scholar] [CrossRef]
- Tamdoğan, N.; Sidal, U. Investigation of poly-β-hydroxybutyrate (PHB) production by Bacillus subtilis ATCC 6633 under different conditions. Kafkas Univ. Vet. Fak. Derg. 2011, 17, 173–176. [Google Scholar] [CrossRef]
- Cui, Y.W.; Shi, Y.P.; Gong, X.Y. Effects of C/N in the substrate on the simultaneous production of polyhydroxyalkanoates and extracellular polymeric substances by Haloferax mediterranei via kinetic model analysis. RSC Adv. 2017, 7, 18953–18961. [Google Scholar] [CrossRef] [Green Version]
- Merugu, R.; Rao, A.S. Optimization of polyhydroxybutyrate production by two phototrophic bacteria Rhodobacter capsulatus KU002 and Rhodopseudomonas palustris KU003. J. Chem. Pharm. Res. 2015, 7, 260–264. [Google Scholar]
- Jin, H.; Nikolau, B.J. Evaluating PHA productivity of bioengineered Rhodosprillum rubrum. PLoS ONE 2014, 9, e96621. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Tobitani, K.; Aoki, S.; Song, Y.; Ooi, T.; Taguchi, S. Improved production of poly(lactic acid)-like polyester based on metabolite analysis to address the rate-limiting step. AMB Express 2014, 4, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gonzalez, L.; De Wever, H. Acetic acid as an indirect sink of CO2 for the synthesis of polyhydroxyalkanoates (PHA): Comparison with PHA production processes directly using CO2 as feedstock. Appl. Sci. 2018, 8, 1416. [Google Scholar] [CrossRef] [Green Version]
- Argun, H.; Kargi, F.; Kapdan, I.K. Light fermentation of dark fermentation effluent for bio-hydrogen production by different Rhodobacter species at different initial volatile fatty acid (VFA) concentrations. Int. J. Hydrog. Energy 2008, 33, 7405–7412. [Google Scholar] [CrossRef]
- Fang, H.H.P.; Zhu, H.; Zhang, T. Phototrophic hydrogen production from glucose by pure and co-cultures of Clostridium butyricum and Rhodobacter sphaeroides. Int. J. Hydrog. Energy 2006, 31, 2223–2230. [Google Scholar] [CrossRef]
- Silva, L.G.; Lorenzetti, A.P.R.; Ribeiro, R.A.; Alves, I.R.; Leaden, L.; Galhardo, R.S.; Koide, T.; Marques, M.V. OxyR and the hydrogen peroxide stress response in Caulobacter crescentus. Gene 2019, 700, 70–84. [Google Scholar] [CrossRef]
- Koskimäki, J.J.; Kajula, M.; Hokkanen, J.; Ihantola, E.L.; Kim, J.H.; Hautajärvi, H.; Hankala, E.; Suokas, M.; Pohjanen, J.; Podolich, O.; et al. Methyl-esterified 3-hydroxybutyrate oligomers protect bacteria from hydroxyl radicals. Nat. Chem. Biol. 2016, 12, 332–338. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.R.; Fitriana, H.N.; Lee, S.Y.; Kim, M.-S.; Moon, M.; Lee, W.-H.; Lee, J.-S.; Lee, S. Molecular Profiling and Optimization Studies for Growth and PHB Production Conditions in Rhodobacter sphaeroides. Energies 2020, 13, 6471. https://doi.org/10.3390/en13236471
Lee YR, Fitriana HN, Lee SY, Kim M-S, Moon M, Lee W-H, Lee J-S, Lee S. Molecular Profiling and Optimization Studies for Growth and PHB Production Conditions in Rhodobacter sphaeroides. Energies. 2020; 13(23):6471. https://doi.org/10.3390/en13236471
Chicago/Turabian StyleLee, Yu Rim, Hana Nur Fitriana, Soo Youn Lee, Min-Sik Kim, Myounghoon Moon, Won-Heong Lee, Jin-Suk Lee, and Sangmin Lee. 2020. "Molecular Profiling and Optimization Studies for Growth and PHB Production Conditions in Rhodobacter sphaeroides" Energies 13, no. 23: 6471. https://doi.org/10.3390/en13236471