Willow Biomass as Energy Feedstock: The Effect of Habitat, Genotype and Harvest Rotation on Thermophysical Properties and Elemental Composition

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Field Experiments
2.2. Examination of Willow Biomass Quality
2.3. Statistical Analysis
3. Results and Discussion
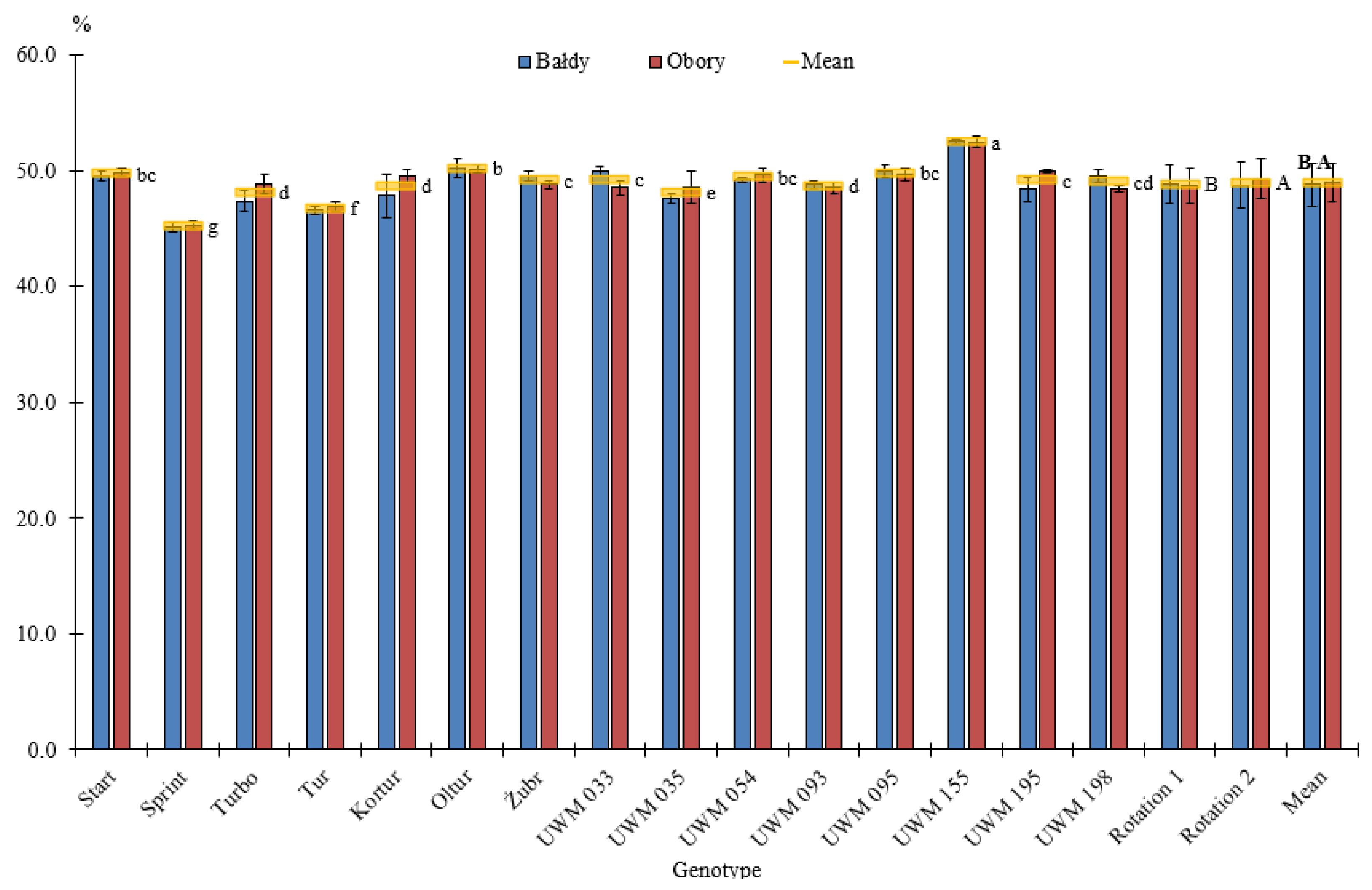
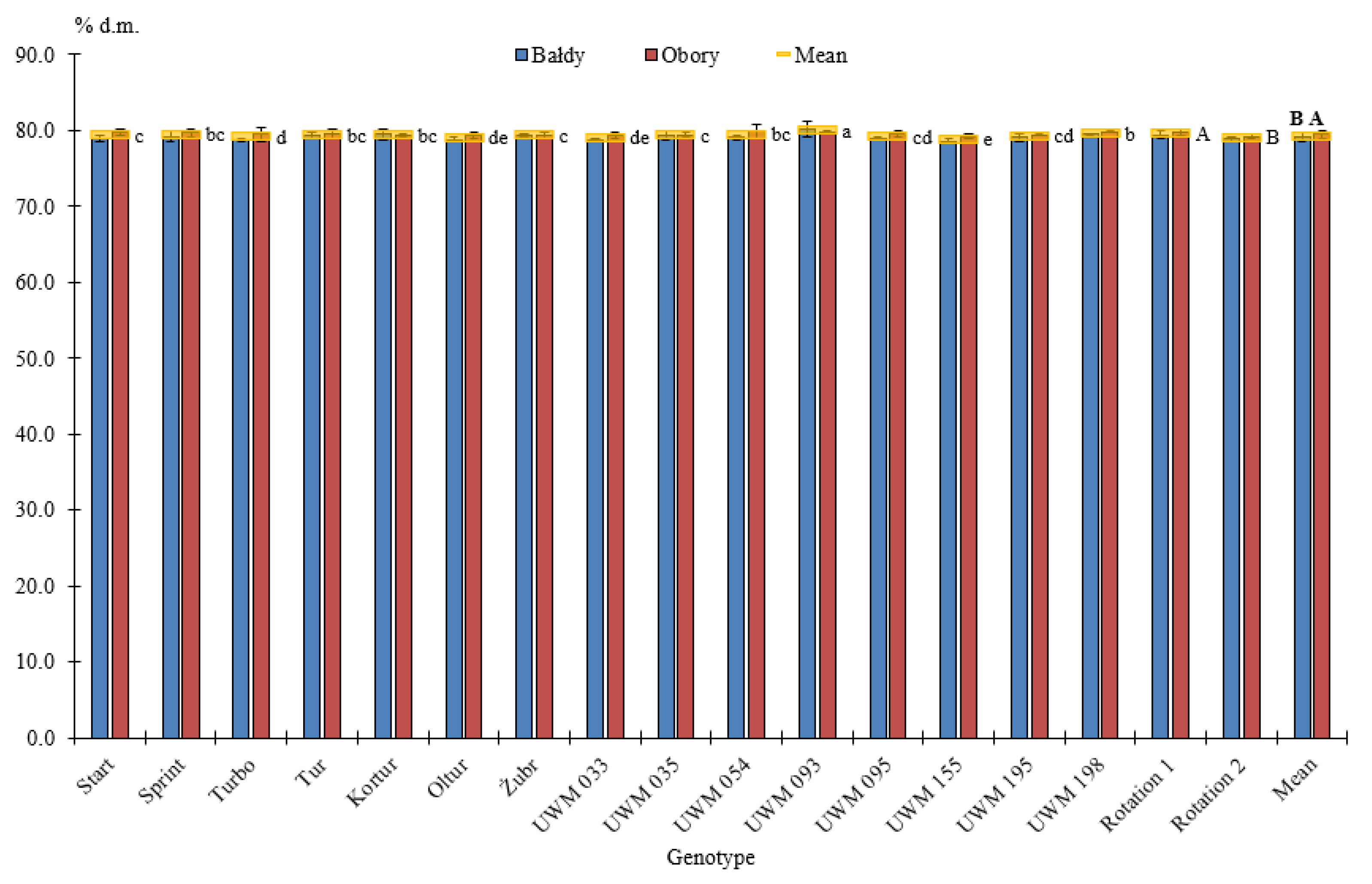
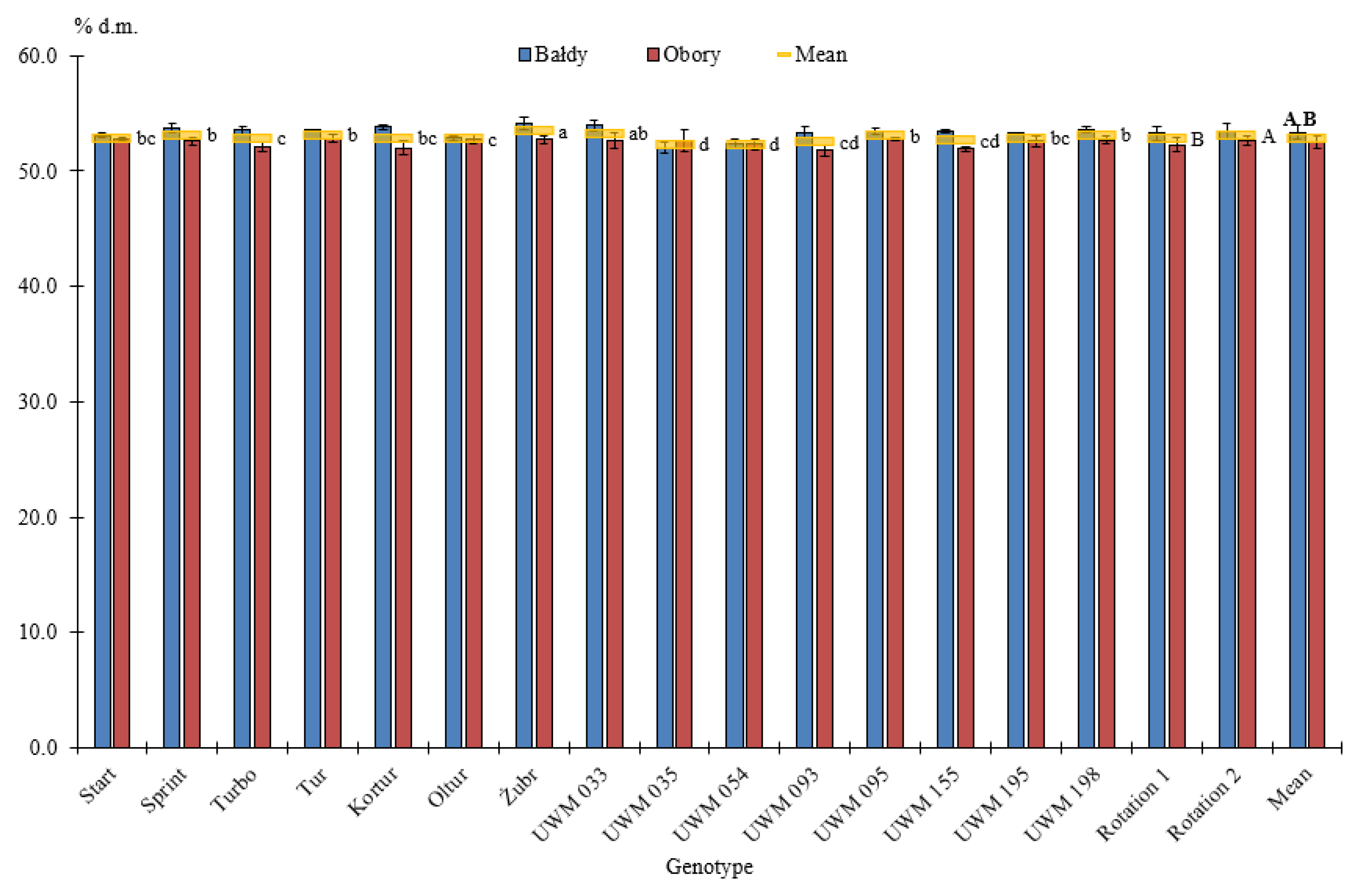
3.1. Thermophysical Properties of Willow Biomass
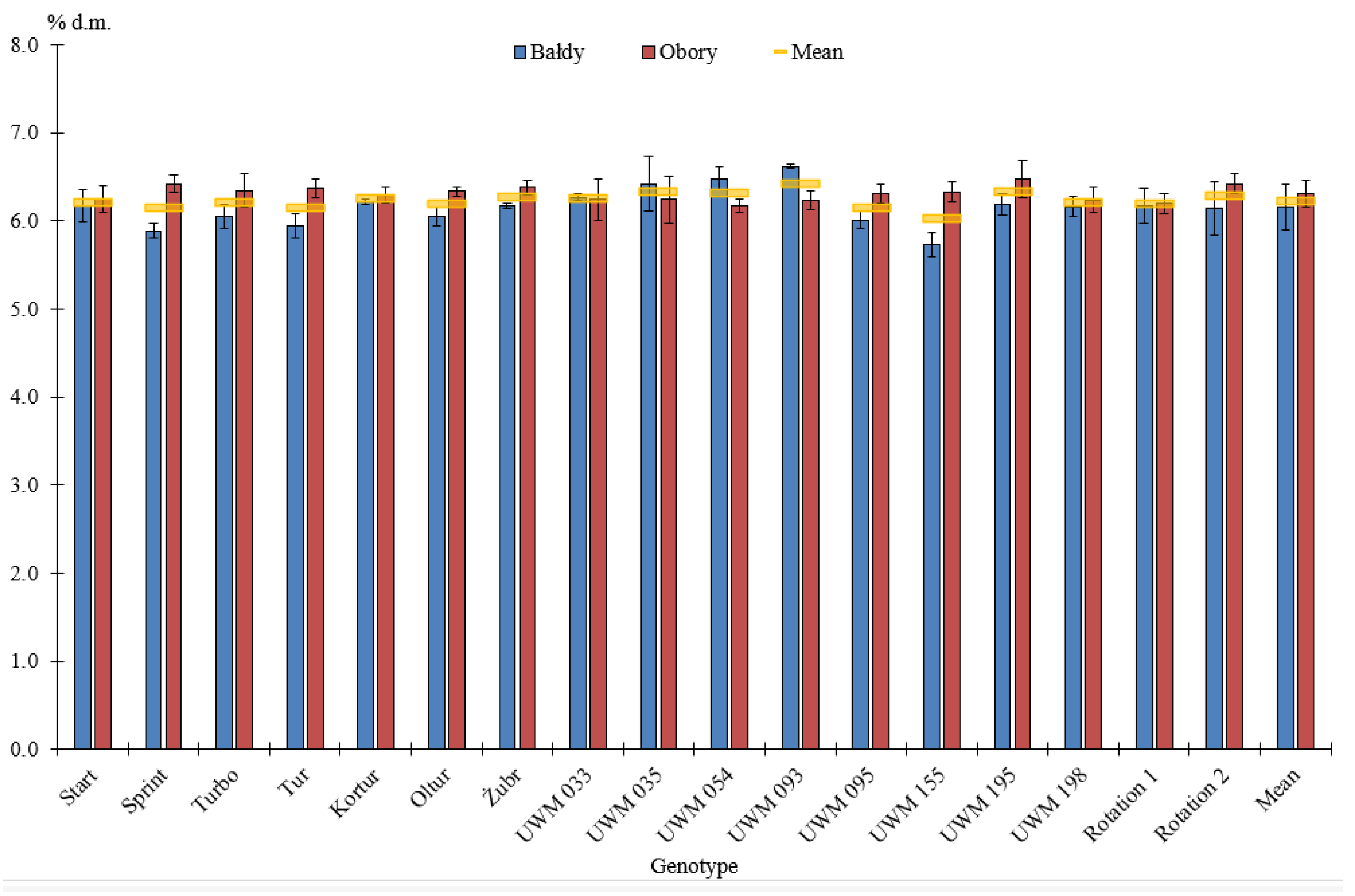
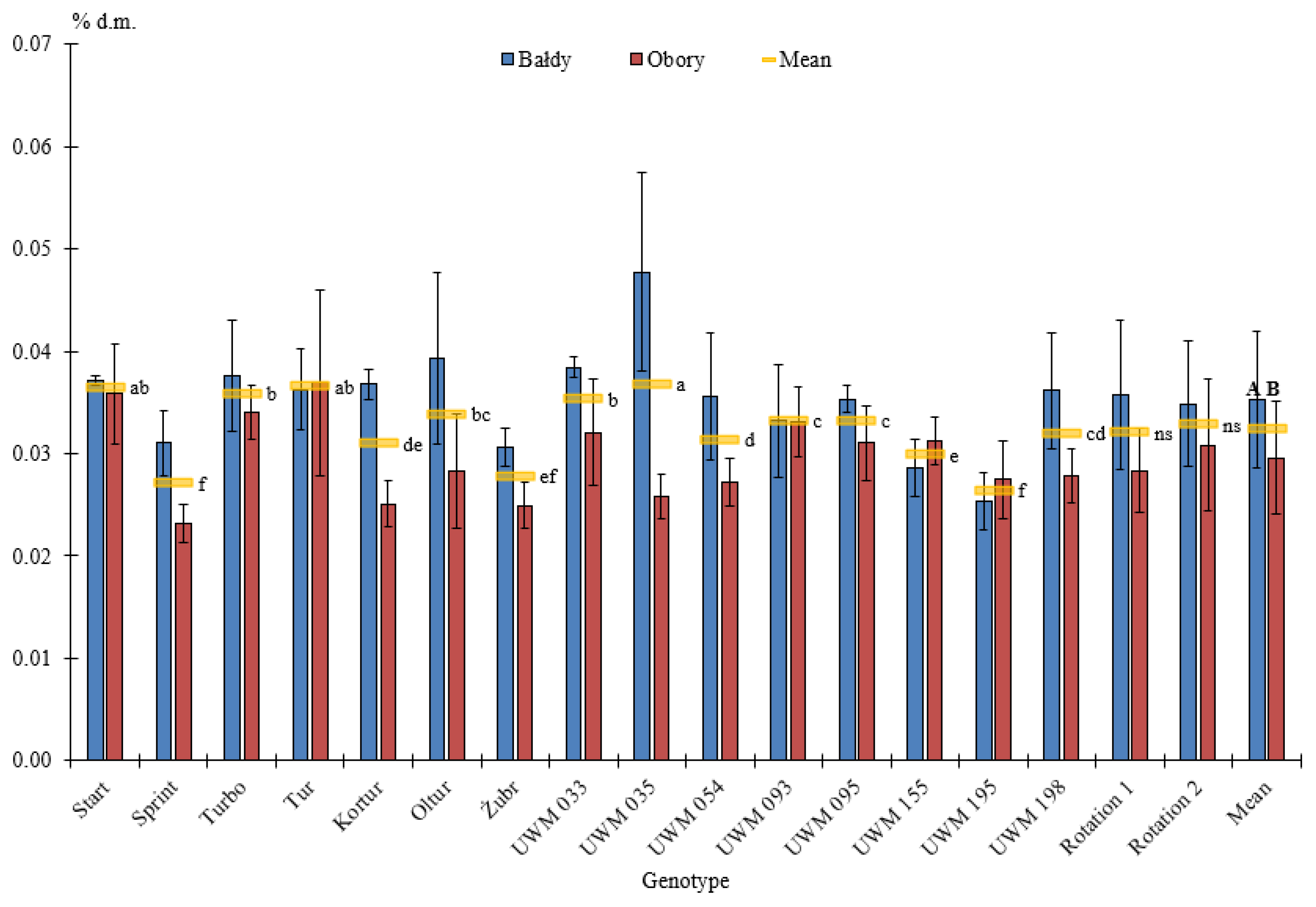
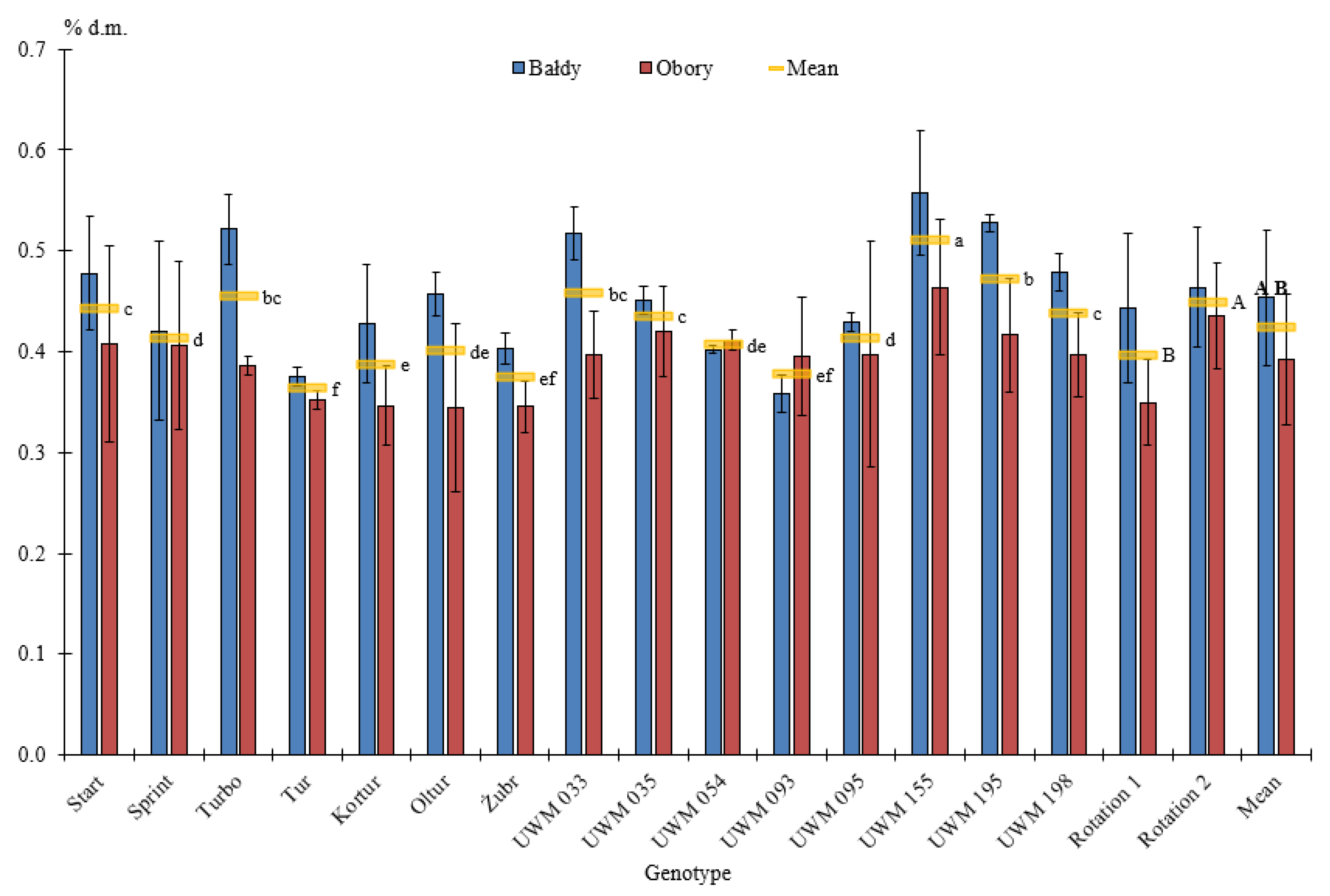
3.2. Elemental Composition of Willow Biomass
3.3. Principal Component Analysis (PCA) and an Assessment of the Impact of the Factors under Analysis on Willow Biomass Properties
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Calvo Buendia, E.; Tanabe, K.; Kranjc, A.; Baasansuren, J.; Fukuda, M.; Ngarize, S.; Osako, A.; Pyrozhenko, Y.; Shermanau, P.; Federici, S. (Eds.) IPCC Intergovernmental Panel on Climate Change (IPCC). In 2019 Refinement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Olabi, A.G. Energy quadrilemma and the future of renewable energy. Energy 2016, 108, 1–6. [Google Scholar] [CrossRef]
- Foley, A.; Olabi, A.G. Renewable energy technology developments, trends and policy implications that can underpin the drive for global climate change. Renew. Sust. Energ. Rev. 2017, 68, 1112–1114. [Google Scholar] [CrossRef] [Green Version]
- Pari, L.; Alfano, V.; Garcia-Galindo, D.; Suardi, A.; Santangelo, E. Pruning biomass potential in Italy related to crop characteristics, agricultural practices and agro-climatic conditions. Energies 2018, 11, 1365. [Google Scholar] [CrossRef] [Green Version]
- Toscano, G.; Alfano, V.; Scarfone, A.; Pari, L. Pelleting vineyard pruning at low cost with a mobile technology. Energies 2018, 11, 2477. [Google Scholar] [CrossRef] [Green Version]
- Eurostat. Energy. Energy statistics-quantities (nrg_quant). Available online: https://ec.europa.eu/eurostat/web/energy/data/database (accessed on 28 January 2020).
- Central Statistical Office. Energy from renewable sources in 2018. Available online: https://stat.gov.pl/en/topics/environment-energy/energy/energy-from-renewable-sources-in-2018,3,11.html (accessed on 21 April 2020).
- Central Statistical Office. Forestry 2017. Available online: https://stat.gov.pl/en/publications/search.html?letter=F&page=2 (accessed on 21 April 2020).
- Stolarski, M.J.; Niksa, D.; Krzyżaniak, M.; Tworkowski, J.; Szczukowski, S. Willow productivity from small and large-scale experimental plantations in Poland from 2000 to 2017. Renew. Sustain. Energy Rev. 2019, 101, 461–475. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Szczukowski, S.; Krzyżaniak, M.; Tworkowski, J. Energy value of yield and biomass quality in a 7-year rotation of willow cultivated on marginal soil. Energies 2020, 13, 2144. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Warmiński, K.; Krzyżaniak, M. Energy value of yield and biomass quality of poplar grown in two consecutive 4-year harvest rotations in the north-east of Poland. Energies 2020, 13, 1495. [Google Scholar] [CrossRef] [Green Version]
- Serapiglia, M.J.; Cameron, K.D.; Stipanovic, A.J.; Abrahamson, L.P.; Volk, T.A.; Smart, L.B. Yield and woody biomass traits of novel shrub willow hybrids at two contrasting sites. Bioenergy Res. 2013, 6, 533–546. [Google Scholar] [CrossRef] [Green Version]
- Manzone, M.; Bergante, S.; Facciotto, G. Energy and economic evaluation of a poplar plantation for woodchips production in Italy. Biomass Bioenergy 2014, 60, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Bioenergy Europe. Statistical Report 2019: Biomass Supply; Bioenergy Europe: Brussels, Belgium, 2019; p. 35. [Google Scholar]
- Aronsson, P.; Rosenqvist, H.; Dimitriou, I. Impact of nitrogen fertilization to short rotation willow coppice plantations grown in Sweden on yield and economy. Bioenergy Res. 2014, 7, 993–1001. [Google Scholar] [CrossRef]
- Cunniff, J.; Purdy, S.J.; Barraclough, T.J.P.; Castle, M.; Maddison, A.L.; Jones, L.E.; Shield, I.F.; Gregory, A.S.; Karp, A. High yielding biomass genotypes of willow (Salix spp.) show differences in below-ground biomass allocation. Biomass Bioenergy 2015, 80, 114–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djomo, S.N.; Witters, N.; Van Dael, M.; Gabrielle, B.; Ceulemans, R. Impact of feedstock, land use change, and soil organic carbon on energy and greenhouse gas performance of biomass cogeneration technologies. Appl. Energy 2015, 154, 122–130. [Google Scholar] [CrossRef]
- Pacaldo, R.; Volk, T.; Briggs, R. Carbon sequestration in fine roots and foliage biomass offsets soil CO2 effluxes along a 19-year chronosequence of shrub willow (Salix x dasyclados) biomass crops. Bioenergy Res. 2014, 7, 769–776. [Google Scholar] [CrossRef]
- Krzyżaniak, M.; Stolarski, M.J.; Szczukowski, S.; Tworkowski, J. Life cycle assessment of new willow cultivars grown as feedstock for integrated biorefineries. Bioenergy Res. 2016, 9, 224–238. [Google Scholar] [CrossRef] [Green Version]
- Monedero, E.; Portero, H.; Lapuerta, M. Pellet blends of poplar and pine sawdust: Effects of material composition, additive, moisture content and compression die on pellet quality. Fuel Process. Technol. 2015, 132, 15–23. [Google Scholar] [CrossRef]
- Nordborg, M.; Berndes, G.; Dimitriou, I.; Henriksson, A.; Mola-Yudego, B.; Rosenqvist, H. Energy analysis of willow production for bioenergy in Sweden. Renew. Sustain. Energy Rev. 2018, 93, 473–482. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Warmiński, K.; Krzyżaniak, M.; Olba–Zięty, E.; Stachowicz, P. Energy consumption and heating costs for a detached house over a 12-year period – Renewable fuels versus fossil fuels. Energy 2020, 204, 117952. [Google Scholar] [CrossRef]
- Aghaalikhani, A.; Savuto, E.; Di Carlo, A.; Borello, D. Poplar from phytoremediation as a renewable energy source: Gasification properties and pollution analysis. Energy Procedia 2017, 142, 924–931. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Szczukowski, S.; Tworkowski, J.; Krzyżaniak, M. Cost of heat energy generation from willow biomass. Renew. Energ. 2013, 59, 100–104. [Google Scholar] [CrossRef]
- Tyśkiewicz, K.; Konkol, M.; Kowalski, R.; Rój, E.; Warmiński, K.; Krzyżaniak, M.; Gil, Ł.; Stolarski, M.J. Characterization of bioactive compounds in the biomass of black locust, poplar and willow. Trees 2019, 33, 1235–1263. [Google Scholar] [CrossRef] [Green Version]
- Karp, A.; Shield, I. Bioenergy from plants and the sustainable yield challenge. New Phytol. 2008, 179, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Stolarski, M.J.; Warmiński, K.; Krzyżaniak, M.; Tyśkiewicz, K.; Olba–Zięty, E.; Graban, Ł.; Lajszner, W.; Załuski, D.; Wiejak, R.; Kamiński, P.; et al. How does extraction of biologically active substances with supercritical carbon dioxide affect lignocellulosic biomass properties? Wood Sci. Technol. 2020, 54, 519–546. [Google Scholar] [CrossRef]
- Fabio, E.S.; Volk, T.A.; Miller, O.R.; Serapiglia, M.J.; Kemanian, A.R.; Montes, F.; Kuzovkina, Y.A.; Kling, G.J.; Smart, L.B. Contributions of environment and genotype to variation in shrub willow biomass composition. Ind. Crops Prod. 2017, 108, 149–161. [Google Scholar] [CrossRef]
- Tharakan, P.J.; Volk, T.A.; Abrahamson, L.P.; White, E.H. Energy feedstock characteristics of willow and hybrid poplar clones at harvest age. Biomass Bioenergy 2003, 25, 571–580. [Google Scholar] [CrossRef]
- Tharakan, P.J.; Volk, T.A.; Nowak, C.A.; Abrahamson, L.P. Morphological traits of 30 willow clones and their relationship to biomass production. Can. J. For. Res. 2005, 35, 421–431. [Google Scholar] [CrossRef]
- Adler, A.; Verwijst, T.; Aronsson, P. Estimation and relevance of bark proportion in a willow stand. Biomass Bioenergy 2005, 29, 102–113. [Google Scholar] [CrossRef]
- Novaes, E.; Osorio, L.; Drost, D.R.; Miles, B.L.; Boaventura-Novaes, C.R.D.; Benedict, C.; Dervinis, C.; Yu, Q.; Sykes, R.; Davis, M.; et al. Quantitative genetic analysis of biomass and wood chemistry of Populus under different nitrogen levels. New Phytol. 2009, 182, 878–890. [Google Scholar] [CrossRef]
- Liu, N.; Jørgensen, U.; Lærke, P.E. Concentrations of chemical elements in willow biomass depend on clone, site and management in the field. Bioenergy Res. 2016, 9, 1216–1230. [Google Scholar] [CrossRef]
- Larsen, S.U.; Jørgensen, U.; Kjeldsen, J.B.; Lærke, P.E. Effect of fertilisation on biomass yield, ash and element uptake in SRC willow. Biomass Bioenergy 2016, 86, 120–128. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Śnieg, M.; Krzyżaniak, M.; Tworkowski, J.; Szczukowski, S.; Graban, Ł.; Lajszner, W. Short rotation coppices, grasses and other herbaceous crops: Biomass properties versus 26 genotypes and harvest time. Ind. Crops Prod. 2018, 119, 22–32. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Krzyżaniak, M.; Załuski, D.; Niksa, D. Evaluation of biomass quality of selected woody species depending on the soil enrichment practice. Int. Agrophys. 2018, 32, 111–121. [Google Scholar] [CrossRef]
- Krzyzaniak, M.; Stolarski, M.J.; Szczukowski, S.; Tworkowski, J. Thermophysical and chemical properties of biomass obtained from willow coppice cultivated in one- and three-year rotation cycles. J. Elem. 2015, 20, 161–175. [Google Scholar] [CrossRef]
- Monedero, E.; Hernández, J.J.; Collado, R. Combustion-related properties of poplar, willow and black locust to be used as fuels in power plants. Energies 2017, 10, 997. [Google Scholar] [CrossRef] [Green Version]
- Kauter, D.; Lewandowski, I.; Claupein, W. Quantity and quality of harvestable biomass from Populus short rotation coppice for solid fuel use—A review of the physiological basis and management influences. Biomass Bioenergy 2003, 24, 411–427. [Google Scholar] [CrossRef]
- Serapiglia, M.J.; Gouker, F.E.; Smart, L.B. Early selection of novel triploid hybrids of shrub willow with improved biomass yield relative to diploids. BMC Plant Biol. 2014, 14, 74. [Google Scholar] [CrossRef] [Green Version]
- Serapiglia, M.J.; Gouker, F.E.; Hart, J.F.; Unda, F.; Mansfield, S.D.; Stipanovic, A.J.; Smart, L.B. Ploidy level affects important biomass traits of novel shrub willow (Salix) hybrids. Bioenergy Res. 2015, 8, 259–269. [Google Scholar] [CrossRef]
- Weger, J.; Hutla, P.; Bubeník, J. Yield and fuel characteristics of willows tested for biomass production on agricultural soil. Res. Agric. Eng. 2016, 4, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Bajcar, M.; Zaguła, G.; Saletnik, B.; Tarapatskyy, M.; Puchalski, C. Relationship between torrefaction parameters and physicochemical properties of torrefied products obtained from selected plant biomass. Energies 2018, 11, 2919. [Google Scholar] [CrossRef] [Green Version]
- Klasnja, B.; Kopitovic, S.; Orlovic, S. Wood and bark of some poplar and willow clones as fuelwood. Biomass Bioenergy 2002, 23, 427–432. [Google Scholar] [CrossRef]
- Komorowicz, M.; Wróblewska, H.; Pawłowski, J. Chemical composition and energetic properties of biomass from selected renewable energy sources. Ochr. Śr. Zasobów Nat. 2009, 40, 402–410. (In Polish) [Google Scholar]
- Jagustyn, B.; Patyna, I.; Skawińska, A. Evaluation of physicochemical properties of Palm Kernel Shell as agro biomass used in the energy industry. Chemik 2013, 67, 552–559. [Google Scholar]
- Brosse, N.; Dufour, A.; Meng, X.Z.; Sun, Q.N.; Ragauskas, A. Miscanthus: A fast-growing crop for biofuels and chemicals production. Biofuel. Bioprod. Biorefin. 2012, 6, 580–598. [Google Scholar] [CrossRef]
- Larsen, S.U.; Jørgensen, U.; Lærke, P.E. Biomass yield, nutrient concentration and nutrient uptake by SRC willow cultivars grown on different sites in Denmark. Biomass Bioenergy 2018, 116, 161–170. [Google Scholar] [CrossRef]
- Forbes, E.G.A.; Johnston, C.R.; Archer, J.E.; McCracken, A.R. SRC willow as a bioremediation medium for a dairy farm effluent with high pollution potential. Biomass Bioenergy 2017, 105, 174–189. [Google Scholar] [CrossRef]
- Larsen, S.U.; Jørgensen, U.; Lærke, P.E. Harvest interval and row spacing of SRC willow influence yield and nutrient content. Biomass Bioenergy 2019, 126, 181–189. [Google Scholar] [CrossRef]
- Otepka, P.; Habán, M.; Habánová, M. Cultivation of fast-growing woody plant basket willow (Salix Viminalis L.) and their bioremedial abilities while fertilized with wood ash. Res. J. Agric. Sci. 2011, 43, 218–222. [Google Scholar]
- Adler, A.; Dimitriou, I.; Aronsson, P.; Verwijst, T.; Weih, M. Wood fuel quality of two Salix viminalis stands fertilised with sludge, ash and sludge-ash mixtures. Biomass Bioenergy 2008, 32, 914–925. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | df | Moisture | Ash | Fixed Carbon | Volatile Matter | HHV | LHV | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | Share (%) | F | Share (%) | F | Share (%) | F | Share (%) | F | Share (%) | F | Share (%) | ||
| Location (Loc) | 1 | 31.3 ** | 0.4 | 11.8 ** | 0.6 | 104.9 ** | 10.3 | 127.4 ** | 9.9 | 763.0 ** | 48.9 | 480.0 ** | 7.9 |
| Rep(Loc) | 4 | 1.5 | 0.1 | 2.1 | 0.4 | 0.1 | 0.1 | 0.5 | 0.2 | 1.0 | 0.3 | 0.2 | 0.0 |
| Genotype (Gen) | 14 | 514.6 ** | 80.9 | 96.1 ** | 65.9 | 11.8 ** | 16.1 | 20.2 ** | 22.0 | 22.7 ** | 20.4 | 325.3 ** | 74.8 |
| Loc × Gen | 14 | 42.8 ** | 6.7 | 3.6 ** | 2.5 | 8.1 ** | 11.1 | 8.2 ** | 9.0 | 5.9 ** | 5.3 | 29.0 ** | 6.7 |
| Error 1 | 56 | - | 0.6 | - | 2.7 | - | 5.5 | - | 4.4 | - | 3.6 | - | 0.9 |
| Rotation (Rot) | 1 | 29.8 ** | 0.3 | 136.0 ** | 11.3 | 221.4 ** | 20.2 | 397.7 ** | 26.0 | 37.5 ** | 2.2 | 2.8 | 0.0 |
| Rot × Loc | 1 | 93.3 ** | 1.0 | 16.9 ** | 1.4 | 1.7 | 0.2 | 0.0 | 0.0 | 56.3 ** | 3.3 | 18.8 ** | 0.3 |
| Rot × Rep (Loc) | 4 | 1.5 | 0.1 | 0.9 | 0.3 | 0.4 | 0.1 | 0.9 | 0.2 | 0.3 | 0.1 | 1.7 | 0.1 |
| Rot × Gen | 14 | 27.0 ** | 4.0 | 5.5 ** | 6.4 | 9.7 ** | 12.4 | 11.6 ** | 10.6 | 5.3 ** | 4.4 | 13.2 ** | 3.2 |
| Rot × Loc × Gen | 14 | 36.6 ** | 5.4 | 3.3 ** | 3.9 | 14.9 ** | 19.1 | 15.4 ** | 14.1 | 9.7 ** | 8.1 | 20.8 ** | 5.1 |
| Error 2 | 56 | - | 0.6 | - | 4.7 | - | 5.1 | - | 3.7 | - | 3.3 | - | 1.0 |
| Total | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | |||||||
| Source of Variation | df | C | H | S | N | ||||
|---|---|---|---|---|---|---|---|---|---|
| F | Share (%) | F | Share (%) | F | Share (%) | F | Share (%) | ||
| Location (Loc) | 1 | 480.8 ** | 35.4 | 140.8 ** | 11.1 | 257.9 ** | 17.7 | 903.3 ** | 17.9 |
| Rep(Loc) | 4 | 1.6 | 0.5 | 0.2 | 0.1 | 1.8 | 0.5 | 1.2 | 0.1 |
| Genotype (Gen) | 14 | 18.1 ** | 18.7 | 15.9 ** | 17.6 | 26.1 ** | 25.1 | 107.0 ** | 29.6 |
| Loc × Gen | 14 | 19.5 ** | 20.2 | 34.7 ** | 38.3 | 21.9 ** | 21.1 | 41.4 ** | 11.5 |
| Error 1 | 56 | - | 4.1 | - | 4.4 | - | 3.8 | - | 1.1 |
| Rotation (Rot) | 1 | 39.4 ** | 3.4 | 88.0 ** | 4.7 | 2.7 | 0.3 | 424.7 ** | 13.3 |
| Rot × Loc | 1 | 4.4 ** | 0.4 | 152.1 ** | 8.2 | 12.7 ** | 1.6 | 166.6 ** | 5.2 |
| Rot × Rep (Loc) | 4 | 0.2 | 0.1 | 1.2 | 0.3 | 1.1 | 0.6 | 2.5 | 0.3 |
| Rot × Gen | 14 | 5.1 ** | 6.0 | 10.8 ** | 8.1 | 8.1 ** | 14.2 | 28.4 ** | 12.4 |
| Rot × Loc × Gen | 14 | 5.4 ** | 6.5 | 5.6 ** | 4.2 | 4.6 ** | 8.0 | 15.5 ** | 6.8 |
| Error 2 | 56 | - | 4.8 | - | 3.0 | - | 7.0 | - | 1.8 |
| Total | 100.0 | 100.0 | 100.0 | 100.0 | |||||
| Variable | PC1 | PC2 | PC3 |
|---|---|---|---|
| Moisture | 0.08 | 0.89 | −0.12 |
| Ash | 0.19 | 0.59 | 0.2 |
| Fixed carbon | 0.84 | 0.19 | 0.17 |
| Volatile matter | −0.8 | −0.39 | −0.22 |
| HHV | 0.26 | −0.22 | 0.77 |
| LHV | 0.03 | −0.89 | 0.31 |
| C | 0.68 | −0.38 | 0.15 |
| H | −0.83 | 0.05 | 0.07 |
| S | 0.05 | 0.03 | 0.87 |
| N | 0.69 | 0.34 | 0.29 |
| Eigenvalue | 3.08 | 2.43 | 1.69 |
| Explained variance (%) | 30.8 | 24.3 | 16.9 |
| Variable | Location | Genotype | Rotation | Total |
|---|---|---|---|---|
| Moisture | 3.36 | 3.28 | 3.31 | 3.54 |
| Ash | 11.01 | 10.96 | 12.13 | 13.08 |
| Fixed carbon | 1.63 | 1.09 | 1.86 | 2.63 |
| Volatile matter | 0.46 | 0.34 | 0.55 | 0.70 |
| HHV | 0.97 | 0.51 | 0.58 | 1.10 |
| LHV | 4.42 | 4.12 | 4.13 | 4.62 |
| C | 1.21 | 0.62 | 0.74 | 1.38 |
| H | 2.99 | 1.56 | 2.02 | 3.60 |
| S | 16.89 | 10.77 | 13.31 | 20.84 |
| N | 13.36 | 9.64 | 12.94 | 17.15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stolarski, M.J.; Krzyżaniak, M.; Warmiński, K.; Załuski, D.; Olba-Zięty, E. Willow Biomass as Energy Feedstock: The Effect of Habitat, Genotype and Harvest Rotation on Thermophysical Properties and Elemental Composition. Energies 2020, 13, 4130. https://doi.org/10.3390/en13164130
Stolarski MJ, Krzyżaniak M, Warmiński K, Załuski D, Olba-Zięty E. Willow Biomass as Energy Feedstock: The Effect of Habitat, Genotype and Harvest Rotation on Thermophysical Properties and Elemental Composition. Energies. 2020; 13(16):4130. https://doi.org/10.3390/en13164130
Chicago/Turabian StyleStolarski, Mariusz Jerzy, Michał Krzyżaniak, Kazimierz Warmiński, Dariusz Załuski, and Ewelina Olba-Zięty. 2020. "Willow Biomass as Energy Feedstock: The Effect of Habitat, Genotype and Harvest Rotation on Thermophysical Properties and Elemental Composition" Energies 13, no. 16: 4130. https://doi.org/10.3390/en13164130